CELSR1 Promotes Neuroprotection in Cerebral Ischemic Injury Mainly through the Wnt/PKC Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
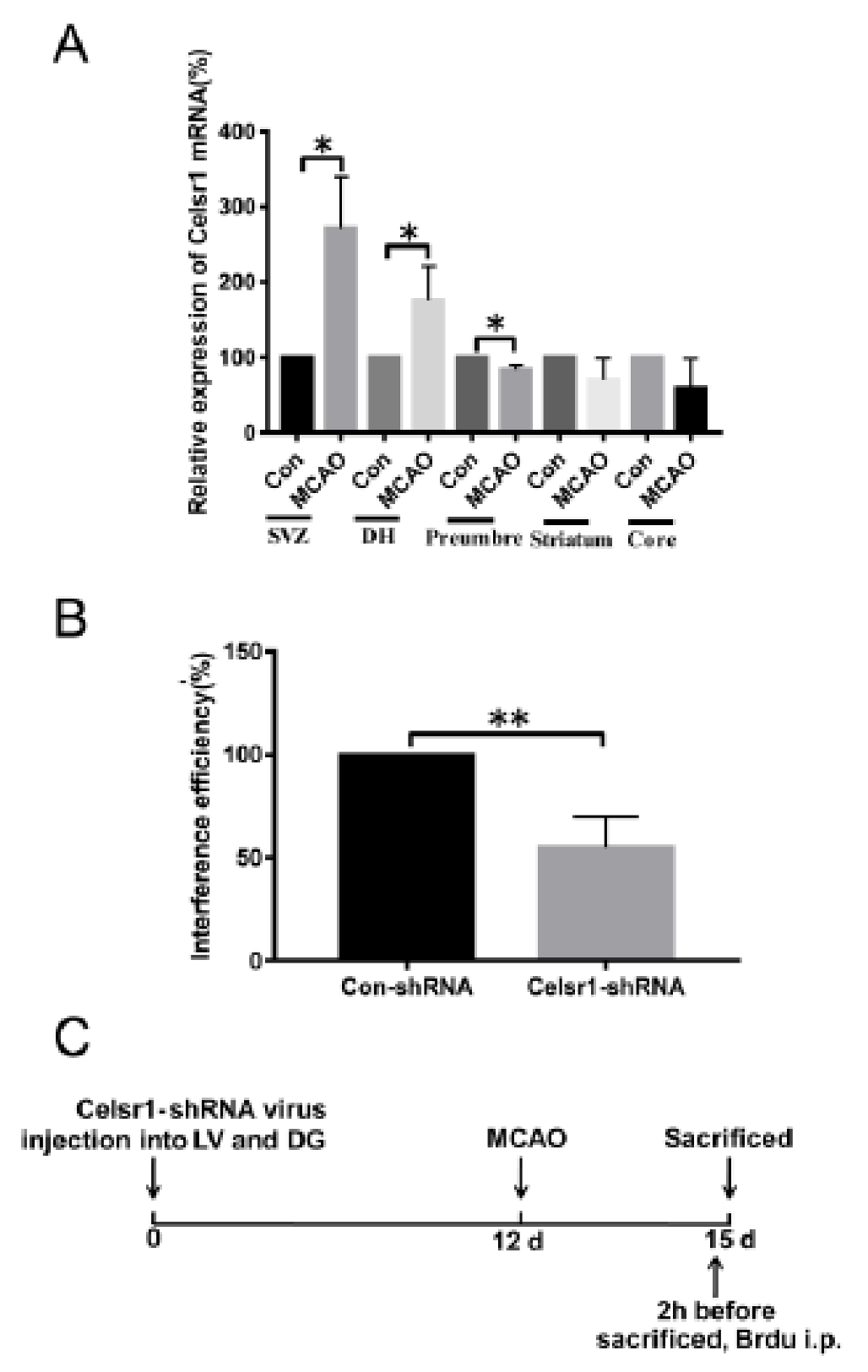
2.1. The Expression Level of Celsr1 Increased Significantly in the Ischemic SVZ and DG
2.2. A Celsr1-shRNA Lentivirus Was Constructed and Microinjected into the Brain
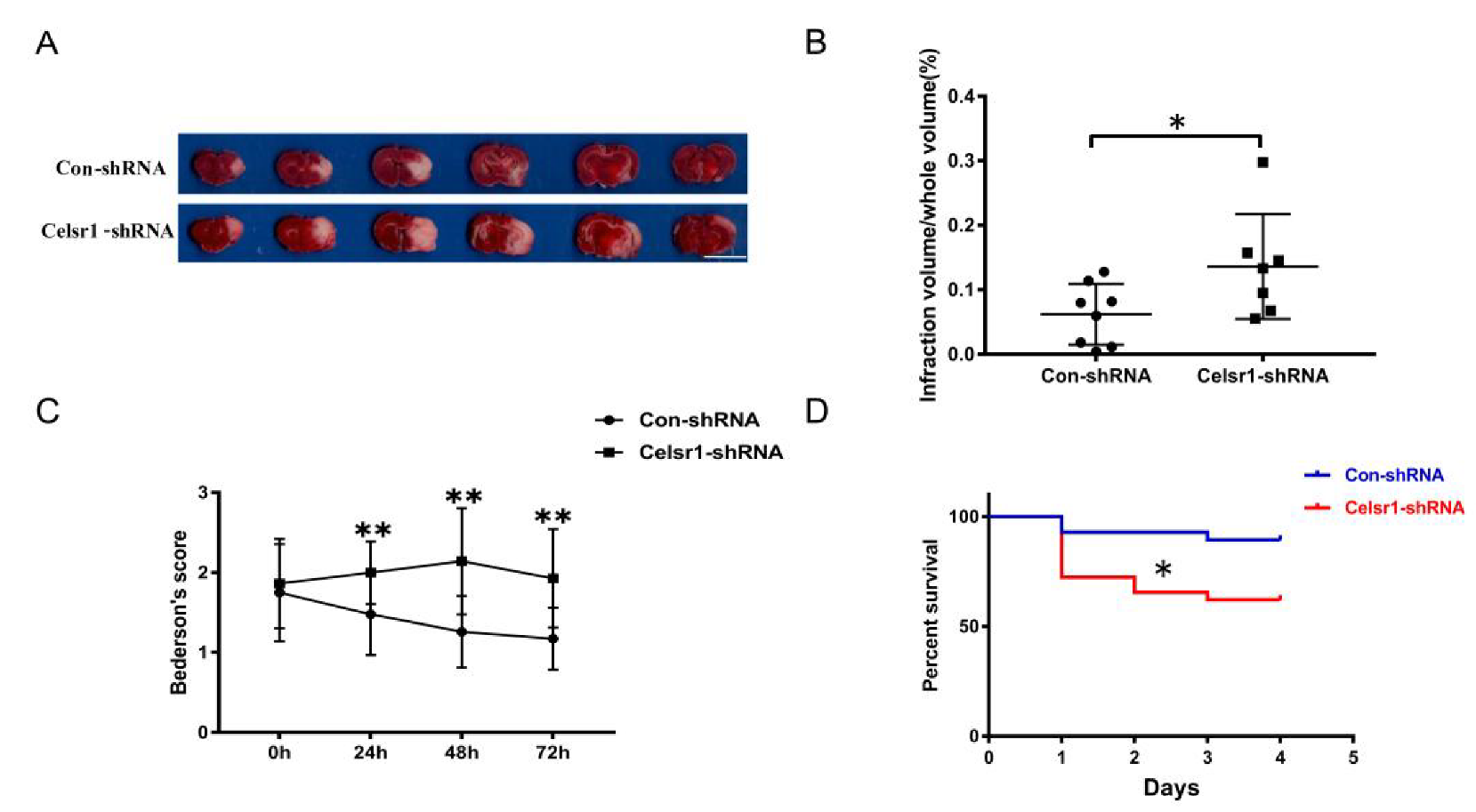
2.3. Knockdown of Celsr1 in the SVZ Accelerated Brain Injury Induced by Ischemia/Reperfusion
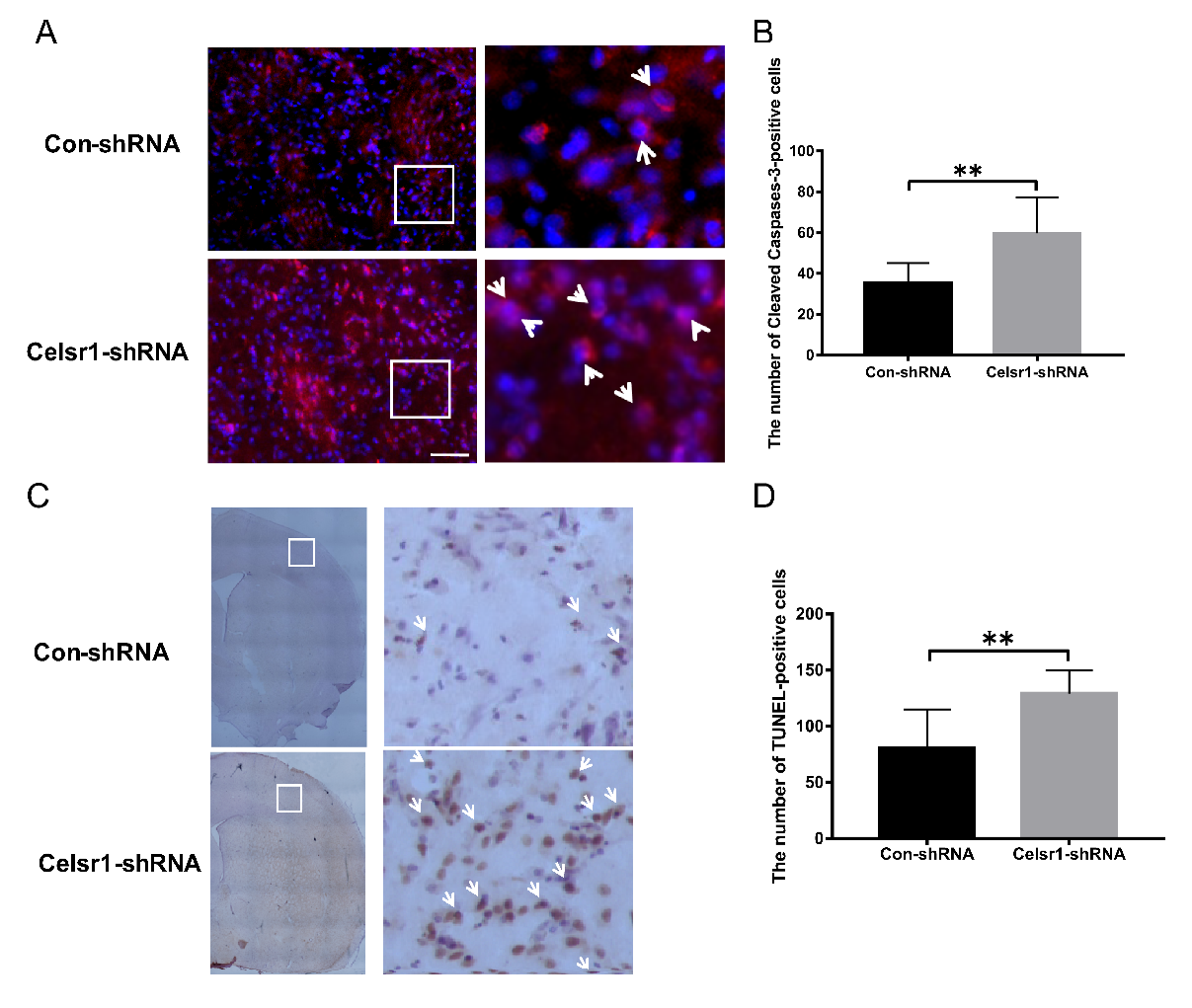
2.4. Celsr1 Knockdown in the SVZ Increased the Percentage of Apoptotic Cells in the Peri-infarct Cerebral Cortex
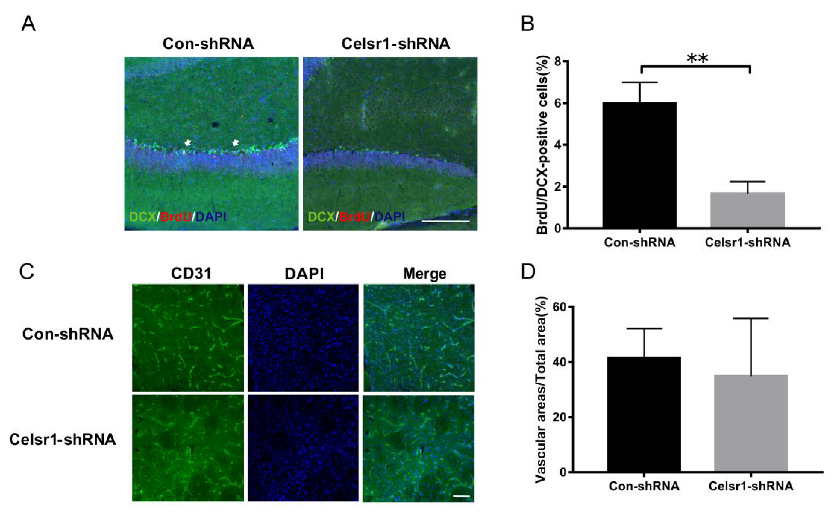
2.5. Celsr1 Knockdown in the SVZ Inhibited Neurogenesis and Angiogenesis after Cerebral Ischemia
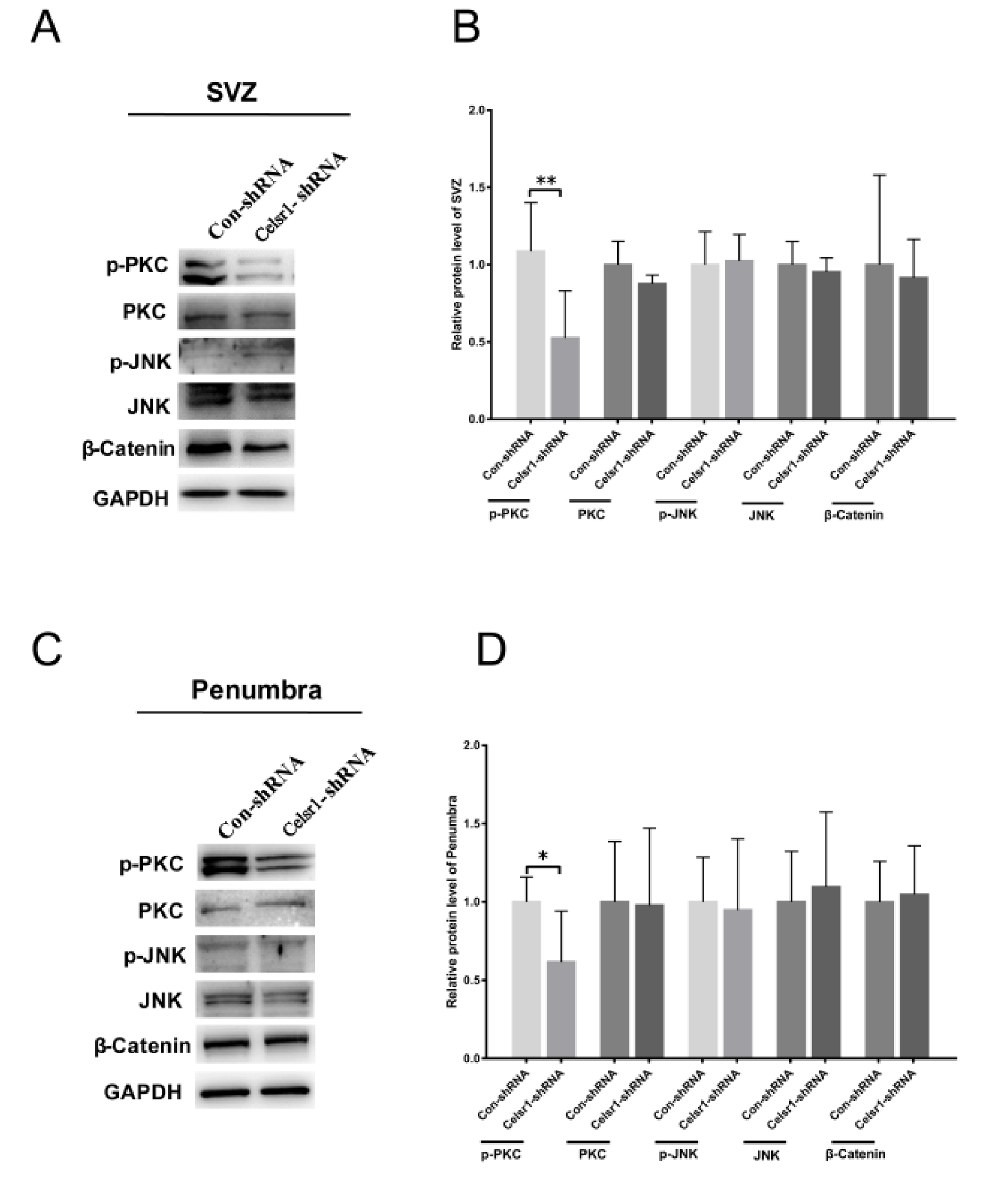
2.6. Celsr1 Knockdown in the SVZ Suppressed the Wnt/PKC Signaling Pathway after Cerebral Ischemia
2.7. Celsr1 Knockdown in the DG Had no Influence on Cell Apoptosis in the Peri-infarct Cerebral Cortex or Neurological Deficit Scores
2.8. Celsr1 Knockdown in the DG Inhibited Neurogenesis but did not Affect Angiogenesis after Cerebral Ischemia
3. Discussion
4. Materials and Methods
4.1. Experimental Model
4.2. Intracerebral Microinjection
4.3. Middle Cerebral Artery (MCA) Occlusion Model
4.4. Examination of Neurological Deficits
4.5. Evaluation of Infarct Volume
4.6. BrdU Labeling
4.7. Immunohistochemistry
4.8. Western Blot Analysis
4.9. Quantitative RT-PCR
4.10. TUNEL Staining
4.11. Cell Transfection
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CELSR1 | Cadherin epidermal growth factor (EGF) laminin G (LAG) seven-pass G-type receptor 1 |
| MCAO | middle cerebral artery occlusion |
| SVZ | subventricular zone |
| tPA | tissue plasminogen activator |
| SGZ | subgranular zone |
| NSCs | neural stem cells |
| PCP | planar cell polarity |
| GFP | green fluorescent protein |
| DG | dentate gyrus |
| CCA | common carotid artery |
| ECA | external carotid artery |
| ICA | internal carotid artery |
| MCA | middle cerebral artery |
| TTC | triphenyltetrazolium chloride |
| TUNEL | TdT-mediated dUTP-biotin nick end labeling |
References
- Hui, Z.; Sha, D.J.; Wang, S.L.; Li, C.S.; Qian, J.; Wang, J.Q.; Zhao, Y.; Zhang, J.H.; Cheng, H.Y.; Yang, H.; et al. Panaxatriol saponins promotes angiogenesis and enhances cerebral perfusion after ischemic stroke in rats. BMC Complement. Altern. Med. 2017, 17, 70. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.R.; Khatri, P.; Broderick, J.P.; Grotta, J.C.; Kasner, S.E.; Kim, D.; Meyer, B.C.; Panagos, P.; Romano, J.; Scott, P. Review, historical context, and clarifications of the NINDS rt-PA stroke trials exclusion criteria: Part 1: Rapidly improving stroke symptoms. Stroke 2013, 44, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Tilley, B.C.; Lyden, P.D.; Brott, T.G.; Lu, M.; Levine, S.R.; Welch, K.M. Total quality improvement method for reduction of delays between emergency department admission and treatment of acute ischemic stroke. The National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group. Arch. Neurol. 1997, 54, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Hayakawa, K.; Maki, T.; Arai, K.; Lo, E.H. Effects of Postconditioning on Neurogenesis and Angiogenesis during the Recovery Phase after Focal Cerebral Ischemia. Stroke 2015, 46, 2691–2694. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Investig. 2000, 106, 829–838. [Google Scholar] [CrossRef]
- Ruan, L.; Wang, B.; ZhuGe, Q.; Jin, K. Coupling of neurogenesis and angiogenesis after ischemic stroke. Brain Res. 2015, 1623, 166–173. [Google Scholar] [CrossRef]
- Woitke, F.; Ceanga, M.; Rudolph, M.; Niv, F.; Witte, O.W.; Redecker, C.; Kunze, A.; Keiner, S. Adult hippocampal neurogenesis poststroke: More new granule cells but aberrant morphology and impaired spatial memory. PLoS ONE 2017, 12, e0183463. [Google Scholar] [CrossRef]
- Lu, J.; Manaenko, A.; Hu, Q. Targeting Adult Neurogenesis for Poststroke Therapy. Stem Cells Int. 2017, 2017, 5868632. [Google Scholar] [CrossRef]
- Zhang, R.L.; Zhang, Z.G.; Zhang, L.; Chopp, M. Proliferation and differentiation of progenitor cells in the cortex and the subventricular zone in the adult rat after focal cerebral ischemia. Neuroscience 2001, 105, 33–41. [Google Scholar] [CrossRef]
- Vallee, A.; Guillevin, R.; Vallee, J.N. Vasculogenesis and angiogenesis initiation under normoxic conditions through Wnt/beta-catenin pathway in gliomas. Rev. Neurosci. 2018, 29, 71–91. [Google Scholar] [CrossRef]
- Boutin, C.; Goffinet, A.M.; Tissir, F. Celsr1-3 cadherins in PCP and brain development. Curr. Top. Dev. Biol. 2012, 101, 161–183. [Google Scholar] [PubMed]
- Hadjantonakis, A.K.; Sheward, W.J.; Harmar, A.J.; de Galan, L.; Hoovers, J.M.; Little, P.F. Celsr1, a neural-specific gene encoding an unusual seven-pass transmembrane receptor, maps to mouse chromosome 15 and human chromosome 22qter. Genomics 1997, 45, 97–104. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Duncan, J.S.; Stoller, M.L.; Francl, A.F.; Tissir, F.; Devenport, D.; Deans, M.R. Celsr1 coordinates the planar polarity of vestibular hair cells during inner ear development. Dev. Biol. 2017, 423, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Zhang, D.L.; Xu, Z.G.; Ma, M.L.; Wang, W.B.; Li, L.L.; Han, X.L.; Huo, Y.; Yu, X.; Sun, J.P. Understanding cadherin EGF LAG seven-pass G-type receptors. J. Neurochem. 2014, 131, 699–711. [Google Scholar] [CrossRef]
- Yamada, Y.; Fuku, N.; Tanaka, M.; Aoyagi, Y.; Sawabe, M.; Metoki, N.; Yoshida, H.; Satoh, K.; Kato, K.; Watanabe, S.; et al. Identification of CELSR1 as a susceptibility gene for ischemic stroke in Japanese individuals by a genome-wide association study. Atherosclerosis 2009, 207, 144–149. [Google Scholar] [CrossRef]
- Zhan, Y.H.; Lin, Y.; Tong, S.J.; Ma, Q.L.; Lu, C.X.; Fang, L.; Wei, W.; Cai, B.; Wang, N. The CELSR1 polymorphisms rs6007897 and rs4044210 are associated with ischaemic stroke in Chinese Han population. Ann. Hum. Biol. 2015, 42, 26–30. [Google Scholar] [CrossRef]
- Schafer, S.T.; Han, J.; Pena, M.; von Bohlen Und Halbach, O.; Peters, J.; Gage, F.H. The Wnt adaptor protein ATP6AP2 regulates multiple stages of adult hippocampal neurogenesis. J. Neurosci. 2015, 35, 4983–4998. [Google Scholar] [CrossRef]
- Boucherie, C.; Boutin, C.; Jossin, Y.; Schakman, O.; Goffinet, A.M.; Ris, L.; Gailly, P.; Tissir, F. Neural progenitor fate decision defects, cortical hypoplasia and behavioral impairment in Celsr1-deficient mice. Mol. Psychiatry 2018, 23, 723–734. [Google Scholar] [CrossRef]
- Zhan, Y.H.; Luo, Q.C.; Zhang, X.R.; Xiao, N.A.; Lu, C.X.; Yue, C.; Wang, N.; Ma, Q.L. CELSR1 Is a Positive Regulator of Endothelial Cell Migration and Angiogenesis. Biochemistry 2016, 81, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Tissir, F.; Goffinet, A.M. Atypical cadherins Celsr1-3 and planar cell polarity in vertebrates. Prog. Mol. Biol. Transl. Sci. 2013, 116, 193–214. [Google Scholar] [PubMed]
- Tissir, F.; De-Backer, O.; Goffinet, A.M.; Lambert de Rouvroit, C. Developmental expression profiles of Celsr (Flamingo) genes in the mouse. Mech. Dev. 2002, 112, 157–160. [Google Scholar] [CrossRef]
- Qu, Z.; Su, F.; Qi, X.; Sun, J.; Wang, H.; Qiao, Z.; Zhao, H.; Zhu, Y. Wnt/beta-catenin signalling pathway mediated aberrant hippocampal neurogenesis in kainic acid-induced epilepsy. Cell Biochem. Funct. 2017, 35, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Chen, S.; Cai, S.; Dong, L.; Liu, L.; Yang, Y.; Guo, F.; Lu, X.; He, H.; Chen, Q.; et al. Wnt5a through noncanonical Wnt/JNK or Wnt/PKC signaling contributes to the differentiation of mesenchymal stem cells into type II alveolar epithelial cells in vitro. PLoS ONE 2014, 9, e90229. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Park, H.; Lee, H.W.; Kwon, Y.G. The Wnt pathway and the roles for its antagonists, DKKS, in angiogenesis. Iubmb Life 2012, 64, 724–731. [Google Scholar] [CrossRef]
- Alvarez-Buylla, A.; Lim, D.A. For the long run: Maintaining germinal niches in the adult brain. Neuron 2004, 41, 683–686. [Google Scholar] [CrossRef]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and functional implications of adult neurogenesis. Cell 2008, 132, 645–660. [Google Scholar] [CrossRef]
- Jin, K.; Minami, M.; Lan, J.Q.; Mao, X.O.; Batteur, S.; Simon, R.P.; Greenberg, D.A. Neurogenesis in dentate subgranular zone and rostral subventricular zone after focal cerebral ischemia in the rat. Proc. Natl. Acad. Sci. USA 2001, 98, 4710–4715. [Google Scholar] [CrossRef]
- Kokaia, Z.; Lindvall, O. Neurogenesis after ischaemic brain insults. Curr. Opin. Neurobiol. 2003, 13, 127–132. [Google Scholar] [CrossRef]
- Wang, X.J.; Sun, T.; Kong, L.; Shang, Z.H.; Yang, K.Q.; Zhang, Q.Y.; Jing, F.M.; Dong, L.; Xu, X.F.; Liu, J.X.; et al. Gypenosides pre-treatment protects the brain against cerebral ischemia and increases neural stem cells/progenitors in the subventricular zone. Int. J. Dev. Neurosci. 2014, 33, 49–56. [Google Scholar] [CrossRef]
- Sun, L.; Lee, J.; Fine, H.A. Neuronally expressed stem cell factor induces neural stem cell migration to areas of brain injury. J. Clin. Investig. 2004, 113, 1364–1374. [Google Scholar] [CrossRef]
- Thored, P.; Arvidsson, A.; Cacci, E.; Ahlenius, H.; Kallur, T.; Darsalia, V.; Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Persistent production of neurons from adult brain stem cells during recovery after stroke. Stem Cells 2006, 24, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef]
- Bendel, O.; Bueters, T.; von Euler, M.; Ogren, S.O.; Sandin, J.; von Euler, G. Reappearance of hippocampal CA1 neurons after ischemia is associated with recovery of learning and memory. J. Cerebr. Blood Flow Met. 2005, 25, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, T.; Wulff, P. The hippocampus in aging and disease: From plasticity to vulnerability. Neuroscience 2015, 309, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Han, C.L.; Li, J.Y.; Wang, C.X.; Luo, L.X. Wnt5a through noncanonical Wnt/JNK signaling promotes the differentiation of human embryonic stem cells to lentoid bodies. Investig. Ophth. Vis. Sci. 2018, 59, 891. [Google Scholar]
- Doudney, K.; Stanier, P. Epithelial cell polarity genes are required for neural tube closure. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 135, 42–47. [Google Scholar] [CrossRef]
- Nishimura, T.; Honda, H.; Takeichi, M. Planar cell polarity links axes of spatial dynamics in neural-tube closure. Cell 2012, 149, 1084–1097. [Google Scholar] [CrossRef]
- Zhang, G.L.; Wang, L.H.; Liu, X.Y.; Zhang, Y.X.; Hu, M.Y.; Liu, L.; Fang, Y.Y.; Mu, Y.; Zhao, Y.; Huang, S.H.; et al. Cerebral Dopamine Neurotrophic Factor (CDNF) Has Neuroprotective Effects against Cerebral Ischemia That May Occur through the Endoplasmic Reticulum Stress Pathway. Int. J. Mol. Sci. 2018, 19, 1905. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-H.; Zhang, G.-L.; Liu, X.-Y.; Peng, A.; Ren, H.-Y.; Huang, S.-H.; Liu, T.; Wang, X.-J. CELSR1 Promotes Neuroprotection in Cerebral Ischemic Injury Mainly through the Wnt/PKC Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1267. https://doi.org/10.3390/ijms21041267
Wang L-H, Zhang G-L, Liu X-Y, Peng A, Ren H-Y, Huang S-H, Liu T, Wang X-J. CELSR1 Promotes Neuroprotection in Cerebral Ischemic Injury Mainly through the Wnt/PKC Signaling Pathway. International Journal of Molecular Sciences. 2020; 21(4):1267. https://doi.org/10.3390/ijms21041267
Chicago/Turabian StyleWang, Li-Hong, Geng-Lin Zhang, Xing-Yu Liu, Ai Peng, Hai-Yuan Ren, Shu-Hong Huang, Ting Liu, and Xiao-Jing Wang. 2020. "CELSR1 Promotes Neuroprotection in Cerebral Ischemic Injury Mainly through the Wnt/PKC Signaling Pathway" International Journal of Molecular Sciences 21, no. 4: 1267. https://doi.org/10.3390/ijms21041267
APA StyleWang, L.-H., Zhang, G.-L., Liu, X.-Y., Peng, A., Ren, H.-Y., Huang, S.-H., Liu, T., & Wang, X.-J. (2020). CELSR1 Promotes Neuroprotection in Cerebral Ischemic Injury Mainly through the Wnt/PKC Signaling Pathway. International Journal of Molecular Sciences, 21(4), 1267. https://doi.org/10.3390/ijms21041267