Engineering Herbicide-Tolerance Rice Expressing an Acetohydroxyacid Synthase with a Single Amino Acid Deletion


Abstract
1. Introduction
2. Results
2.1. Herbicide Tolerance Predicted in AHAS Models
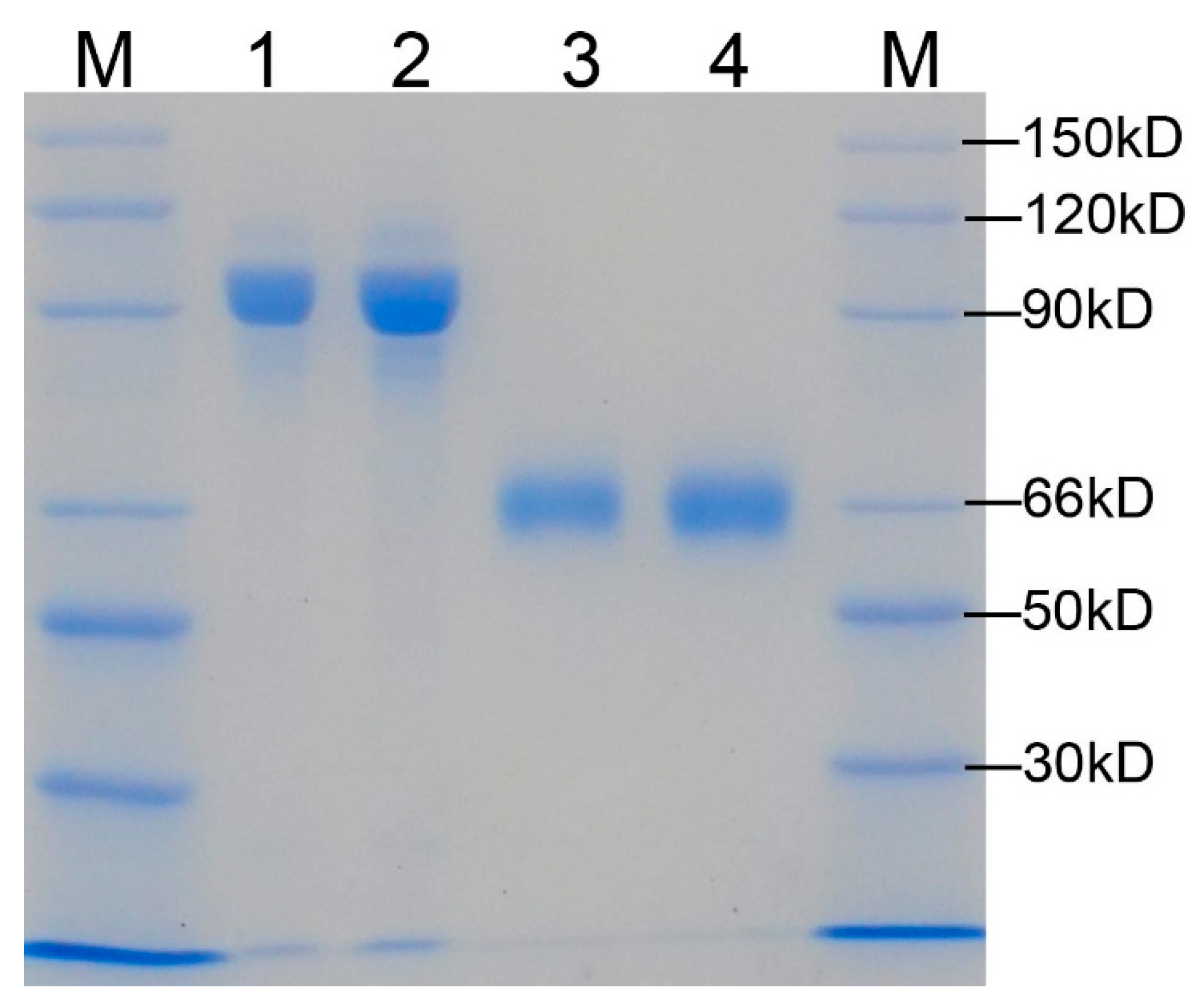
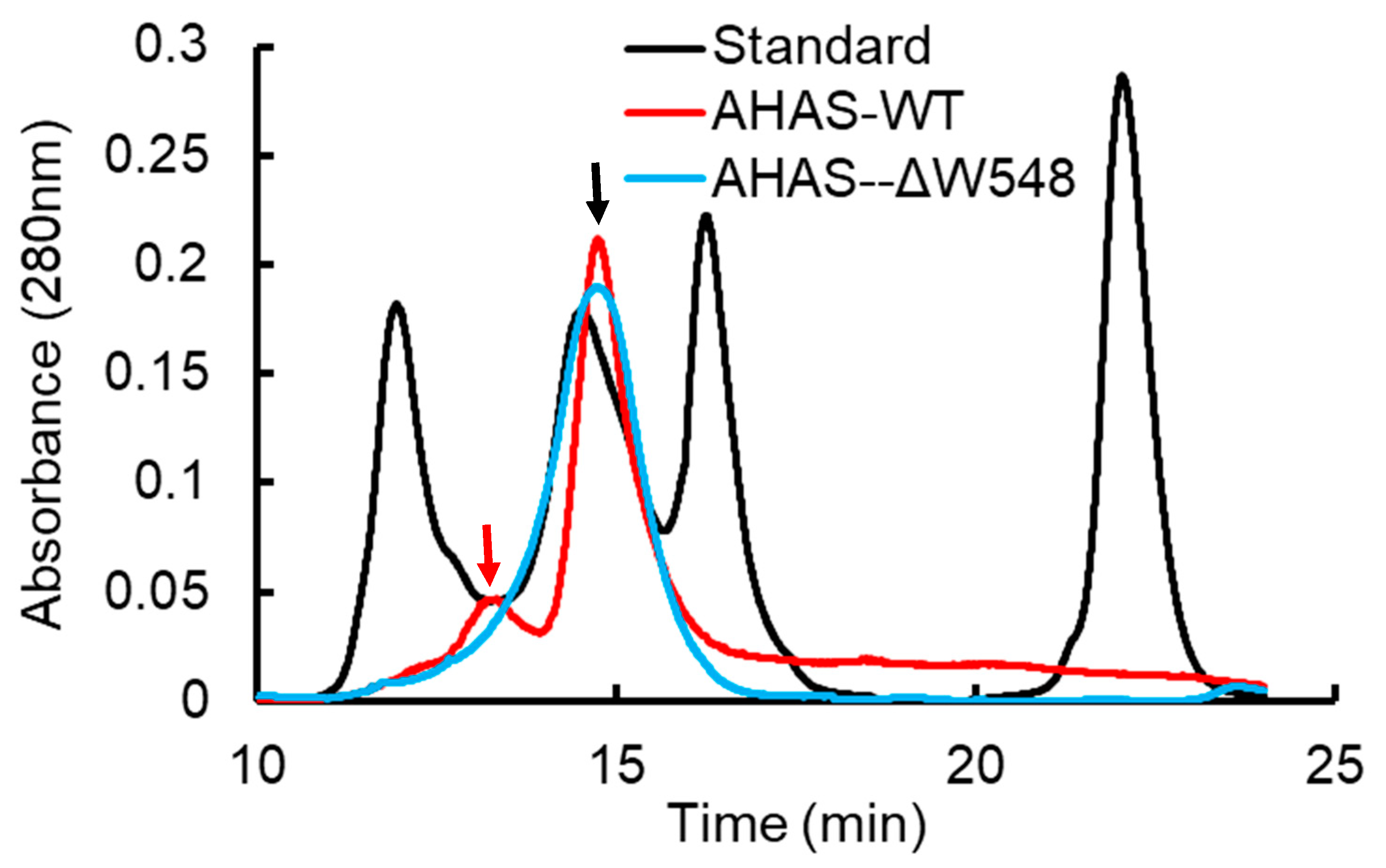
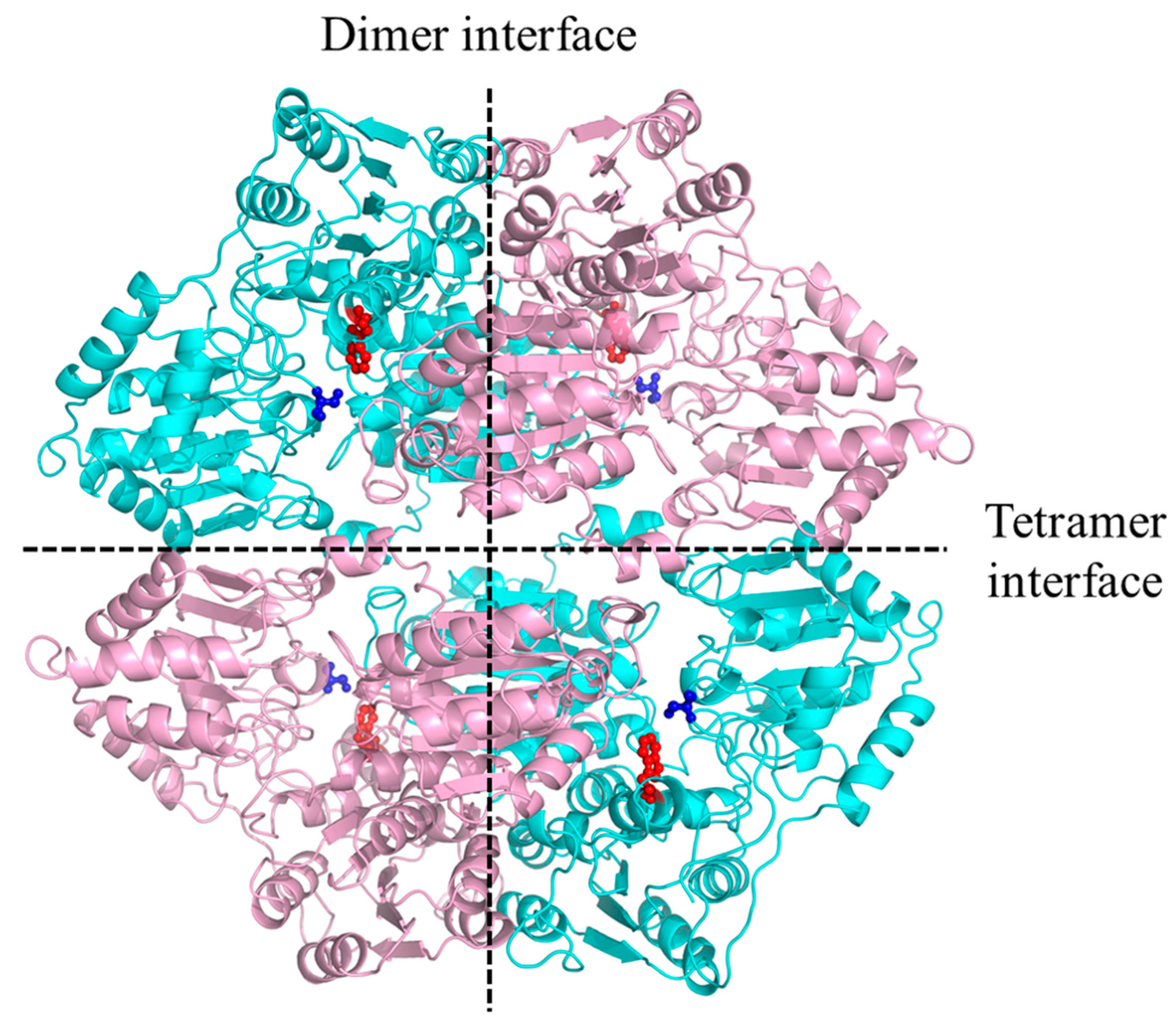
2.2. The W548 Deletion Dissociate the Tetramer in Vitro
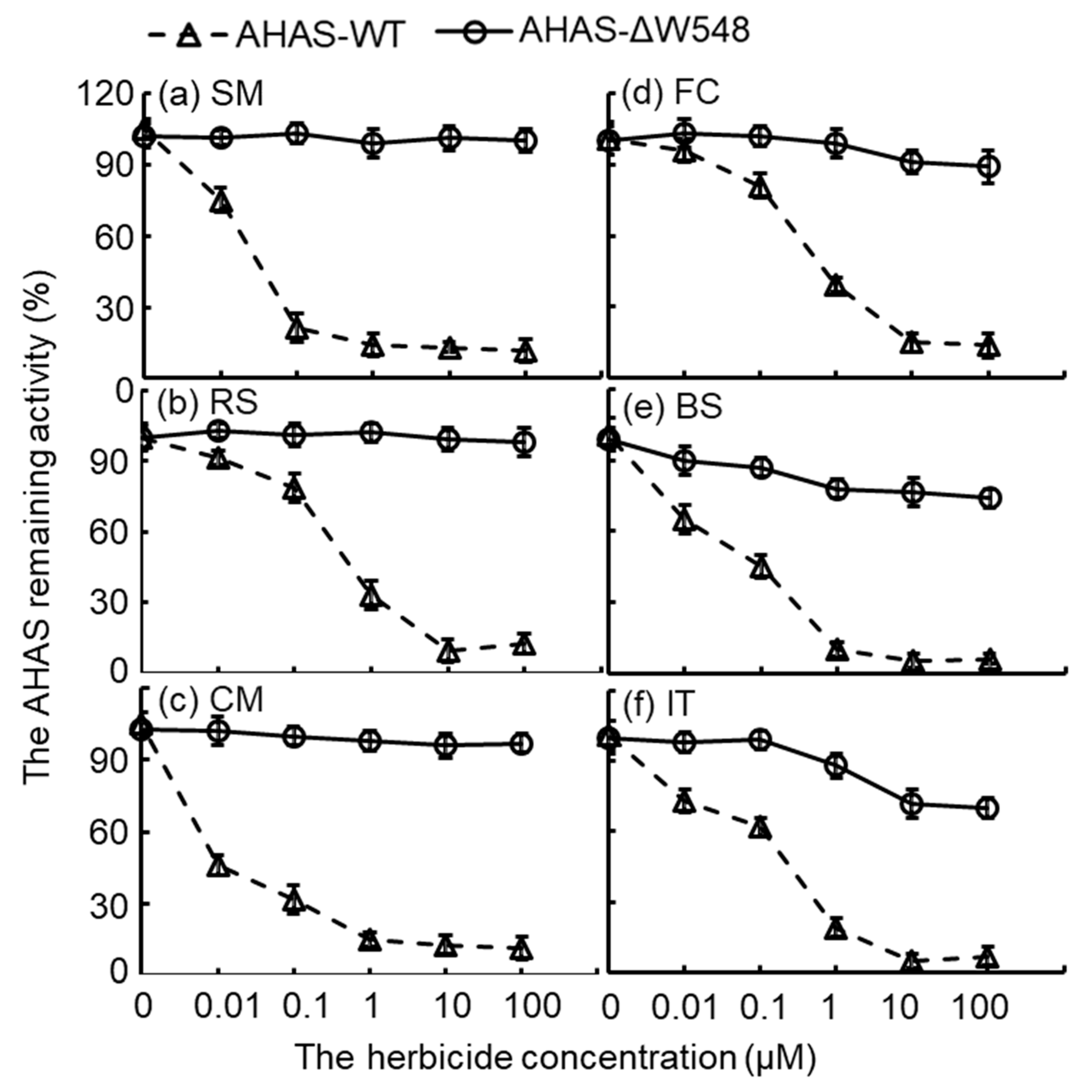
2.3. Multi-Herbicide Tolerance of the AHAS-ΔW548
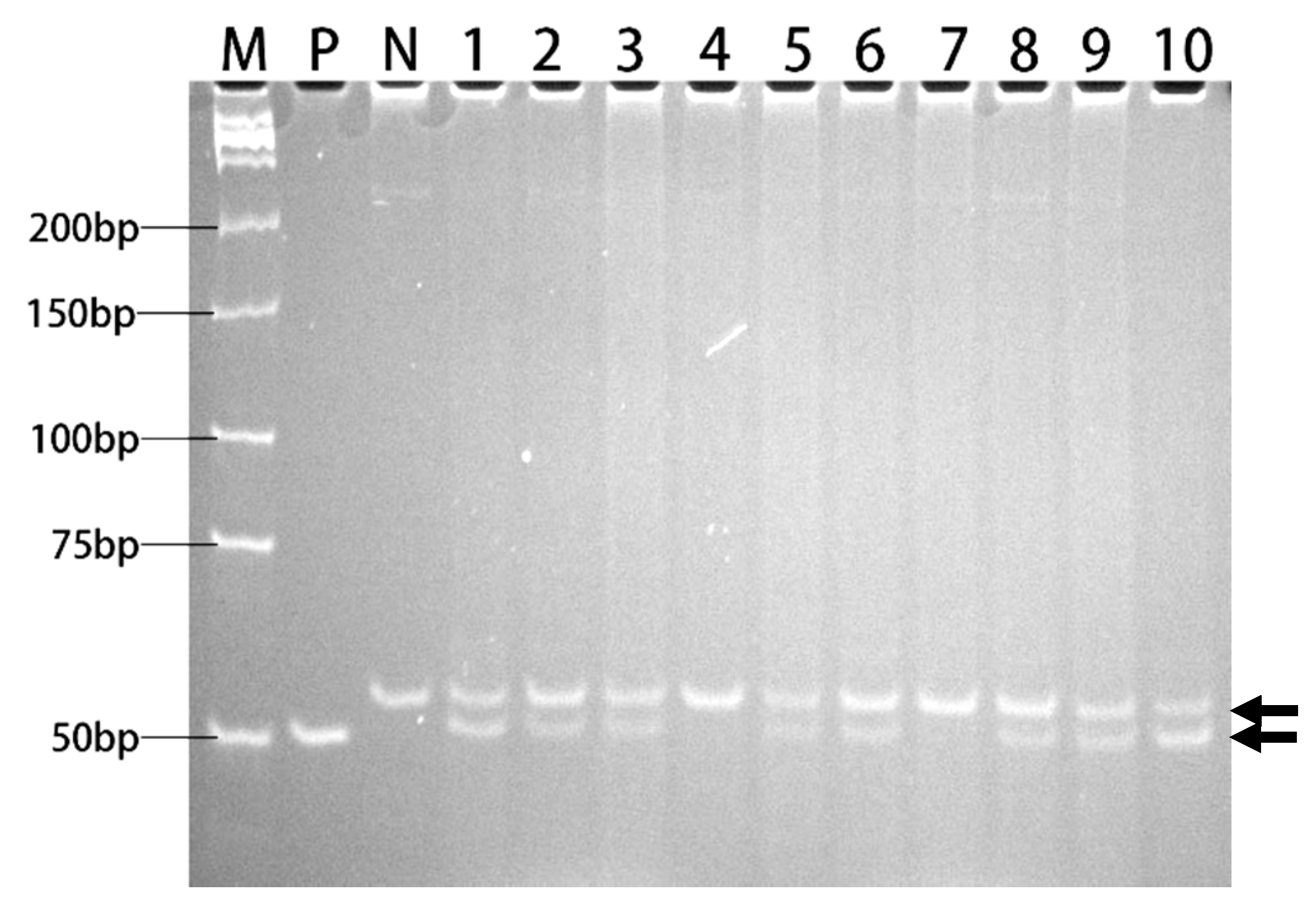
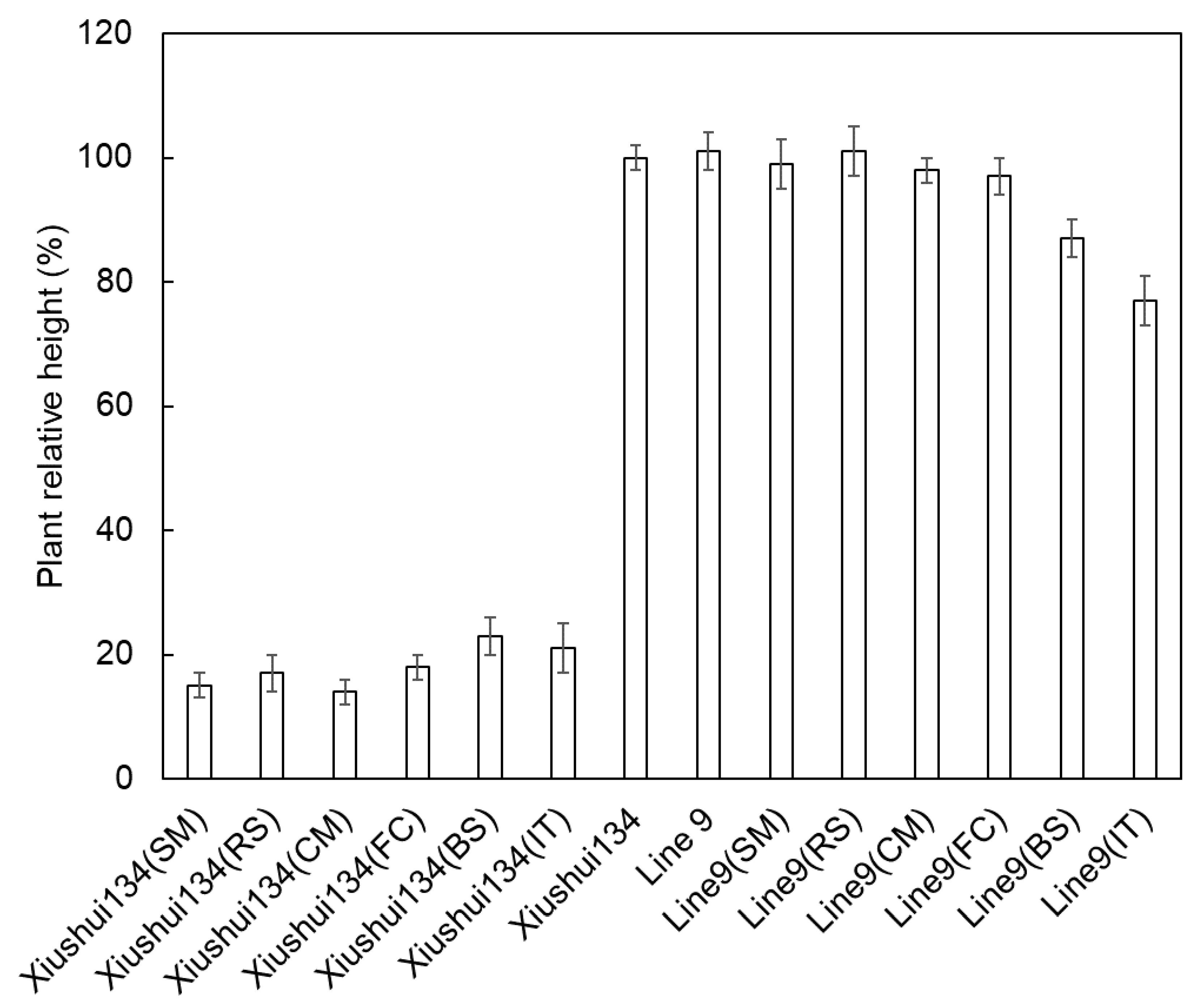
2.4. Transgenic Rice with Herbicide Tolerance
3. Discussion
3.1. Effects of the W548 Deletion on Rice AHAS
3.2. Mechanism of Herbicide Tolerance for the AHAS-ΔW548
3.3. Development of HT Genes Using Residue Deletion
3.4. The Effects of the Ahas-ΔW548 Gene on the Line9
3.5. Novel Herbicide-Tolerant Biotech Crops
4. Materials and Methods
4.1. Structural Simulation of Rice AHAS-Herbicide Complexes
4.2. Purification of the AHAS Proteins
4.3. Activities and Kinetic Assays of Rice AHAS in Vitro
4.4. Development of Transgenic Rice
4.5. Herbicide Tolerance and Agronomic Traits of Transgenic Lines
4.6. Measurement of Total Protein and Free Amino Acids in Rice Seeds
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- ISAAA. Global status of commercialized biotech/GM crops in 2017: Biotech crop adoption surges as economic benefits accumulate in 22 years. ISAAA Brief 2017, 53, 104–106. [Google Scholar]
- Chauhan, B.S. Strategies to manage weedy rice in Asia. Crop Prot. 2013, 48, 51–56. [Google Scholar] [CrossRef]
- Owen, M.D.; Beckie, H.J.; Leeson, J.Y.; Norsworthy, J.K.; Steckel, L.E. Integrated pest management and weed management in the United States and Canada. Pest Manag. Sci. 2015, 71, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Rong, J.; Lu, B.R. Occurence and damage of weedy rice and its threats to rice production in China. Weed Sci. (China) 2015, 33, 1–9. [Google Scholar]
- Kanapeckas, K.L.; Vigueira, C.C.; Ortiz, A.; Gettler, K.A.; Burgos, N.R.; Fischer, A.J.; Lawton-Rauh, A.L. Escape to ferality: The endoferal origin of weedy rice from crop rice through de-domestication. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Islam, M.; Biswas, J.; Siyoung, L.; Alam, I.; Mooryong, H. Screening of rice varieties for direct seeding method. Aust. J. Crop Sci. 2014, 8, 536–542. [Google Scholar]
- Liang, Y.; Guo, S.; Yin, L. Comparative study on morphological differences between weedy rice and cultivatec rice in Shanghai area. J. Shanghai Norm. Univ. (Nat. Sci.) 2014, 43, 87–97. [Google Scholar]
- Bai, Y.L.; Gu, L.-L. The market and development of three types of pesticides on rice. Mod. Agrochem. 2017, 16, 1–7. [Google Scholar]
- Yasuor, H.; Osuna, M.D.; Ortiz, A.; Saldain, N.E.; Eckert, J.W.; Fischer, A.J. Mechanism of resistance to penoxsulam in late watergrass [Echinochloa phyllopogon (Stapf) Koss.]. J. Agric. Food Chem. 2009, 57, 3653–3660. [Google Scholar] [CrossRef]
- Ruiz-Santaella, J.P.; Heredia, A.; Prado, R.D. Basis of selectivity of cyhalofop-butyl in Oryza sativa L. Planta 2006, 223, 191–199. [Google Scholar] [CrossRef]
- Rashid, M.H.; Alam, M.M.; Rao, A.N.; Ladha, J.K. Comparative efficacy of pretilachlor and hand weeding in managing weeds and improving the productivity and net income of wet-seeded rice in Bangladesh. Field Crop. Res. 2012, 128, 17–26. [Google Scholar] [CrossRef]
- Tan, S.; Evans, R.R.; Dahmer, M.L.; Singh, B.K.; Shaner, D.L. Imidazolinone-tolerant crops: History, current status and future. Pest Manag. Sci. 2005, 61, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, S.; Thompson, M.E.; Ort, O.; Shaner, D.L.; Stidham, M.; Singh, B.; Tan, S.; Johnson, T.C.; Mann, R.K.; Schmitzer, P.R.; et al. Acetohydroxyacid synthase inhibitors (AHAS/ALS). Mod. Crop Prot. Compd. Second Ed. 2012. [Google Scholar] [CrossRef]
- Duggleby, R.G.; McCourt, J.A.; Guddat, L.W. Structure and mechanism of inhibition of plant acetohydroxyacid synthase. Plant Physiol. Biochem. 2008, 46, 309–324. [Google Scholar] [CrossRef]
- Garcia, M.D.; Nouwens, A.; Lonhienne, T.G.; Guddat, L.W. Comprehensive understanding of acetohydroxyacid synthase inhibition by different herbicide families. Proc. Natl. Acad. Sci. USA 2017, 114, 1091–1100. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S.B. Resistance to AHAS inhibitor herbicides: Current understanding. Pest Manag. Sci. 2014, 70, 1340–1350. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Eberlein, C.V.; Mallory-Smith, C.A.; Hoffman, D.L. DNA-sequence variation in domain a of the acetolactate synthase genes of herbicide-resistant and herbicide-susceptible weed biotypes. Weed Sci. 1992, 40, 670–676. [Google Scholar] [CrossRef]
- MallorySmith, C.A.; Thill, D.C.; Dial, M.J. Identification of sulfonylurea herbicide-resistant prickly lettuce (Lactuca serriola). Weed Technol. 1990, 4, 163–168. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Wang, X. Acetohydroxyacid synthases: Evolution, structure, and function. Appl. Microbiol. Biotechnol. 2016, 100, 8633–8649. [Google Scholar] [CrossRef]
- Shivraina, V.K.; Burgosa, N.R.; Andersb, M.M.; Rajgurua, S.N.; Moorea, J.; Sales, M.A. Gene flow between ClearfieldTM rice and red rice. Crop Prot. 2007, 26, 349–356. [Google Scholar] [CrossRef]
- Jones, J.M.; Dionne, L.; Dell’Orco, J.; Parent, R.; Krueger, J.N.; Cheng, X.; Dib-Hajj, S.D.; Bunton-Stasyshyn, R.K.; Sharkey, L.M.; Dowling, J.J.; et al. Single amino acid deletion in transmembrane segment D4S6 of sodium channel Scn8a (Nav1.6) in a mouse mutant with a chronic movement disorder. Neurobiol. Dis. 2016, 89, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Pagadala, N.S.; Syed, K.; Tuszynski, J. Software for molecular docking: A review. Biophys. Rev. 2017, 9, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Piao, Z.; Wang, W.; Wei, Y.; Zonta, F.; Wan, C.; Bai, J.; Wu, S.; Wang, X.; Fang, J. Characterization of an acetohydroxy acid synthase mutant conferring tolerance to imidazolinone herbicides in rice (Oryza sativa). Planta 2018, 247, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.S.; Guddat, L.W.; Duggleby, R.G. Crystallization of Arabidopsis thaliana acetohydroxyacid synthase in complex with the sulfonylurea herbicide chlorimuron ethyl. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 153–155. [Google Scholar] [CrossRef]
- Kawai, K.; Kaku, K.; Izawa, N.; Shimizu, T.; Fukuda, A.; Tanaka, Y. A novel mutant acetolactase synthase gene from rice cells which confers resistance to ALS-inhibiting herbicides. J. Pest. Sci. 2007, 32, 89–98. [Google Scholar] [CrossRef]
- Chang, A.K.; Duggleby, R.G. Herbicide-resistant forms of Arabidopsis thaliana acetohydroxyacid synthase: Characterization of the catalytic properties and sensitivity to inhibitors of four defined mutants. Biochem. J. 1998, 333, 765–777. [Google Scholar] [CrossRef]
- Lonhienne, T.; Garcia, M.D.; Pierens, G.; Mobli, M.; Nouwens, A.; Guddat, L.W. Structural insights into the mechanism of inhibition of AHAS by herbicides. Proc. Natl. Acad. Sci. USA 2018, 115, 1945–1954. [Google Scholar] [CrossRef]
- Garcia, M.D.; Wang, J.G.; Lonhienne, T.; Guddat, L.W. Crystal structure of plant acetohydroxyacid synthase, the target for several commercial herbicides. FEBS J. 2017, 284, 2037–2051. [Google Scholar] [CrossRef]
- Falco, S.C.; McDevitt, R.E.; Chui, C.F.; Hartnett, M.E.; Knowlton, S.; Mauvais, C.J.; Smith, J.K.; Mazur, B.J. Engineering herbicide-resistant acetolactate synthase. Dev. Ind. Microbiol. 1989, 30, 187–194. [Google Scholar]
- Duggleby, R.G.; Pang, S.S.; Yu, H.; Guddat, L.W. Systematic characterization of mutations in yeast acetohydroxyacid synthase. Interpretation of herbicide-resistance data. Eur. J. Biochem. 2003, 270, 2895–2904. [Google Scholar] [CrossRef]
- Ibdah, M.; Bar-Ilan, A.; Livnah, O.; Schloss, J.V.; Barak, Z.; Chipman, D.M. Homology modeling of the structure of bacterial acetohydroxy acid synthase and examination of the active site by site-directed mutagenesis. Biochemistry 1996, 35, 16282–16291. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Chen, C.Y.; Yang, B. TALEN-mediated homologous recombination produces site-directed DNA base change and herbicide-resistant Rice. J. Genet. Genom. 2016, 43, 291–305. [Google Scholar] [CrossRef]
- Feldmann, K.A.; Marks, M.D.; Christianson, M.L.; Quatrano, R.S. A dwarf mutant of Arabidopsis generated by T-DNA insertion mutagenesis. Science 1989, 243, 1351–1354. [Google Scholar] [CrossRef]
- Liu, M.; Shi, Z.; Zhang, X.; Wang, M.; Zhang, L.; Zheng, K.; Liu, J.; Hu, X.; Di, C.; Qian, Q.; et al. Inducible overexpression of ideal plant Architecture1 improves both yield and disease resistance in rice. Nat. Plants 2019, 5, 389–400. [Google Scholar] [CrossRef]
- Saika, H.; Horita, J.; Taguchi-Shiobara, F.; Nonaka, S.; Nishizawa-Yokoi, A.; Iwakami, S.; Hori, K.; Matsumoto, T.; Tanaka, T.; Itoh, T.; et al. A novel rice cytochrome P450 gene, CYP72A31, confers tolerance to acetolactate synthase-inhibiting herbicides in rice and Arabidopsis. Plant Physiol. 2014, 166, 1232–1240. [Google Scholar] [CrossRef]
- Endo, M.; Shimizu, T.; Fujimori, T.; Yanagisawa, S.; Toki, S. Herbicide-resistant mutations in acetolactate synthase can reduce feedback inhibition and lead to accumulation of branched-chain amino acids. Food Nutr. Sci. 2013, 4, 522–528. [Google Scholar] [CrossRef][Green Version]
- Zhao, C.J.; Schieber, A.; Ganzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations-a review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Lioe, H.N.; Apriyantono, A.; Takara, K.; Wada, K.; Yasuda, M. Umami taste enhancement of MSG/NaCl mixtures by subthreshold L-a-Aromatic amino acids. J. Food Sci. 2005, 70, 401–405. [Google Scholar] [CrossRef]
- Funke, T.; Han, H.; Healy-Fried, M.L.; Fischer, M.; Schonbrunn, E. Molecular basis for the herbicide resistance of Roundup Ready crops. Proc. Natl. Acad. Sci. USA 2006, 103, 13010–13015. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on application (EFSA-GMO-NL-2009-64) for the placing on the market of herbicide-tolerant genetically modified soybean BPS-CV127-9 for food and feed uses, import and processing under regulation (EC) No 1829/2003 from BASF plant science. EFSA J. 2014, 12, 3505. [Google Scholar]
- Sun, Y.; Zhang, X.; Wu, C.; He, Y.; Ma, Y.; Hou, H.; Guo, X.; Du, W.; Zhao, Y.; Xia, L. Engineering herbicide-resistant rice plants through CRISPR/Cas9-mediated homologous recombination of acetolactate synthase. Mol. Plant 2016, 9, 628–631. [Google Scholar] [CrossRef]
- Li, J.; Meng, X.; Zong, Y.; Chen, K.; Zhang, H.; Liu, J.; Li, J.; Gao, C. Gene replacements and insertions in rice by intron targeting using CRISPR-Cas9. Nat. Plants 2016, 2. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.K.; Duggleby, R.G. Expression, purification and characterization of Arabidopsis thaliana acetohydroxyacid synthase. Biochem. J. 1997, 327, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Robards, K.; Haddad, P.R.; Jackson, P.E. High-performance liquid chromatography-separations. Princ. Pract. Mod. Chromatogr. Methods 2004, 6, 305–380. [Google Scholar]
- Yoon, T.Y.; Chung, S.M.; Chang, S.I.; Yoon, M.Y.; Hahn, T.R.; Choi, J.D. Roles of lysine 219 and 255 residues in tobacco acetolactate synthase. Biochem. Biophys. Res. Commun. 2002, 293, 433–439. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Mei, F.; Zhang, W.; Shen, Z.; Fang, J. Creation of Bt rice expressing a fusion protein of Cry1Ac and Cry1I-like using a green tissue-specific promoter. J. Econ. Entomol. 2014, 107, 1674–1679. [Google Scholar] [CrossRef]
- Jiang, B.; Tsao, R.; Li, Y.; Miao, M. Food safety: Food analysis technologies/techniques. Encycl. Agric. Food Syst. 2014, 273–288. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicides | AHAS-WT | AHAS-ΔW548 |
|---|---|---|
| SM | 7.159 | 5.731 |
| RS | 7.491 | 5.834 |
| CM | 7.507 | 5.784 |
| FC | 7.681 | 6.619 |
| BS | 7.208 | 6.389 |
| IT | 7.641 | 7.544 |
| Enzyme/Herbicide a | Km (mM) b | Kcat (s−1) | Catalytic Efficiency Kcat/Km (S−1mM−1) |
|---|---|---|---|
| AHAS-WT | 9.2 ± 0.4 | 6.7 ± 0.3 | 0.71 ± 0.03 |
| AHAS-ΔW548 | 9.4 ± 0.4 | 6.6 ± 0.5 | 0.70 ± 0.05 |
| AHAS-ΔW548 (SM) | 9.3 ± 0.6 | 6.6 ± 0.3 | 0.72 ± 0.05 |
| AHAS-ΔW548 (RS) | 9.4 ± 0.5 | 6.5 ± 0.4 | 0.69 ± 0.04 |
| AHAS-ΔW548 (CM) | 9.2 ± 0.3 | 6.5 ± 0.4 | 0.71 ± 0.03 |
| AHAS-ΔW548 (FC) | 9.4 ± 0.3 | 5.9 ± 0.4 * | 0.63 ± 0.03 * |
| AHAS-ΔW548 (BS) | 11.3 ± 0.6 ** | 5.8 ± 0.3 ** | 0.51 ± 0.04 ** |
| AHAS-ΔW548 (IT) | 12.1 ± 0.3 ** | 5.1 ± 0.3 ** | 0.42 ± 0.03 ** |
| Line Name | Plant Height (cm) | Panicle Numbers Per Plant | Length of Panicle (cm) | Grains Per Panicle | Seed-Setting Rate (%) | 1000-Grain Weight (g) | Yield Per Plant (g) | Grain Width(cm) | Grain Length(cm) |
|---|---|---|---|---|---|---|---|---|---|
| Xiushui134 | 94.1 ± 1.3 | 8.6 ± 0.4 | 14.2 ± 1.8 | 151.2 ± 9.6 | 90.2 ± 1.5 | 26.9 ± 0.7 | 28.5 ± 2.9 | 3.3 ± 0.2 | 6.8 ± 0.3 |
| Line9 | 92.2 ± 2.1 | 8.9 ± 0.5 | 14.8 ± 1.6 | 145.9 ± 8.7 | 91.5 ± 1.3 | 27.2 ± 1.5 | 29.8 ± 3.6 | 3.3 ± 0.2 | 6.9 ± 0.5 |
| Line9 (SM) | 93.5 ± 1.8 | 8.5 ± 0.5 | 15.1 ± 1.2 | 153.5 ± 10.8 | 90.5 ± 2.1 | 26.3 ± 1.3 | 27.3 ± 2.1 | 3.3 ± 0.2 | 6.8 ± 0.4 |
| Compound | Xiushui134 | Line9 | Line9 (SM) | Compound | Xiushui134 | Line9 | Line9 (SM) |
|---|---|---|---|---|---|---|---|
| Ile a | 18 ± 8 | 79 ± 26 ** | 75 ± 23 ** | Trp | 89 ± 29 | 95 ± 21 | 85 ± 31 |
| Leu | 23 ± 11 | 82 ± 25 ** | 87 ± 29 ** | Gly | 103 ± 20 | 105 ± 23 | 99 ± 31 |
| Val | 51 ± 17 | 224 ± 32 ** | 231 ± 28 ** | Gln | 145 ± 29 | 167 ± 35 | 154 ± 31 |
| Phe | 22 ± 9 | 25 ± 7 | 29 ± 6 | Pro | 154 ± 31 | 176 ± 30 | 181 ± 27 |
| Lys | 35 ± 11 | 28 ± 9 | 31 ± 12 | Ala | 266 ± 29 | 311 ± 35 | 304 ± 33 |
| His | 38 ± 16 | 47 ± 8 | 46 ± 9 | Ser | 270 ± 21 | 302 ± 29 | 298 ± 37 |
| Thr | 42 ± 9 | 45 ± 11 | 49 ± 13 | Arg | 340 ± 31 | 362 ± 49 | 388 ± 37 |
| Met | 46 ± 20 | 51 ± 12 | 41 ± 22 | Asp | 513 ± 46 | 501 ± 56 | 481 ± 43 |
| His | 48 ± 17 | 41 ± 21 | 59 ± 15 | Asn | 759 ± 57 | 780 ± 62 | 776 ± 69 |
| Tyr | 75 ± 36 | 89 ± 22 | 95 ± 32 | Glu | 1527 ± 246 | 1382 ± 321 | 1659 ± 356 |
| Total Protein b | 8.9 ± 0.5 | 9.1 ± 0.7 | 9.2 ± 0.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang,, J.; Wan, C.; Wang, W.; Ma, L.; Wang, X.; Cheng, C.; Zhou, J.; Qiao, Y.; Wang, X. Engineering Herbicide-Tolerance Rice Expressing an Acetohydroxyacid Synthase with a Single Amino Acid Deletion. Int. J. Mol. Sci. 2020, 21, 1265. https://doi.org/10.3390/ijms21041265
Fang, J, Wan C, Wang W, Ma L, Wang X, Cheng C, Zhou J, Qiao Y, Wang X. Engineering Herbicide-Tolerance Rice Expressing an Acetohydroxyacid Synthase with a Single Amino Acid Deletion. International Journal of Molecular Sciences. 2020; 21(4):1265. https://doi.org/10.3390/ijms21041265
Chicago/Turabian StyleFang,, Jun, Changzhao Wan, Wei Wang, Liuyin Ma, Xinqi Wang, Can Cheng, Jihua Zhou, Yongjin Qiao, and Xiao Wang. 2020. "Engineering Herbicide-Tolerance Rice Expressing an Acetohydroxyacid Synthase with a Single Amino Acid Deletion" International Journal of Molecular Sciences 21, no. 4: 1265. https://doi.org/10.3390/ijms21041265
APA StyleFang,, J., Wan, C., Wang, W., Ma, L., Wang, X., Cheng, C., Zhou, J., Qiao, Y., & Wang, X. (2020). Engineering Herbicide-Tolerance Rice Expressing an Acetohydroxyacid Synthase with a Single Amino Acid Deletion. International Journal of Molecular Sciences, 21(4), 1265. https://doi.org/10.3390/ijms21041265