Probiotic Properties and Neuroprotective Effects of Lactobacillus buchneri KU200793 Isolated from Korean Fermented Foods

Abstract
:1. Introduction
2. Results and Discussion
2.1. Tolerance to Artificial Gastric Conditions
2.2. Enzyme Production of LAB Strains
2.3. Antibiotic Susceptibility of LAB Strains
2.4. Adhesion Ability of LAB Strains to HT-29 Intestinal Epithelial Cells
2.5. Antioxidant Effects of Heat-Killed LAB Strains
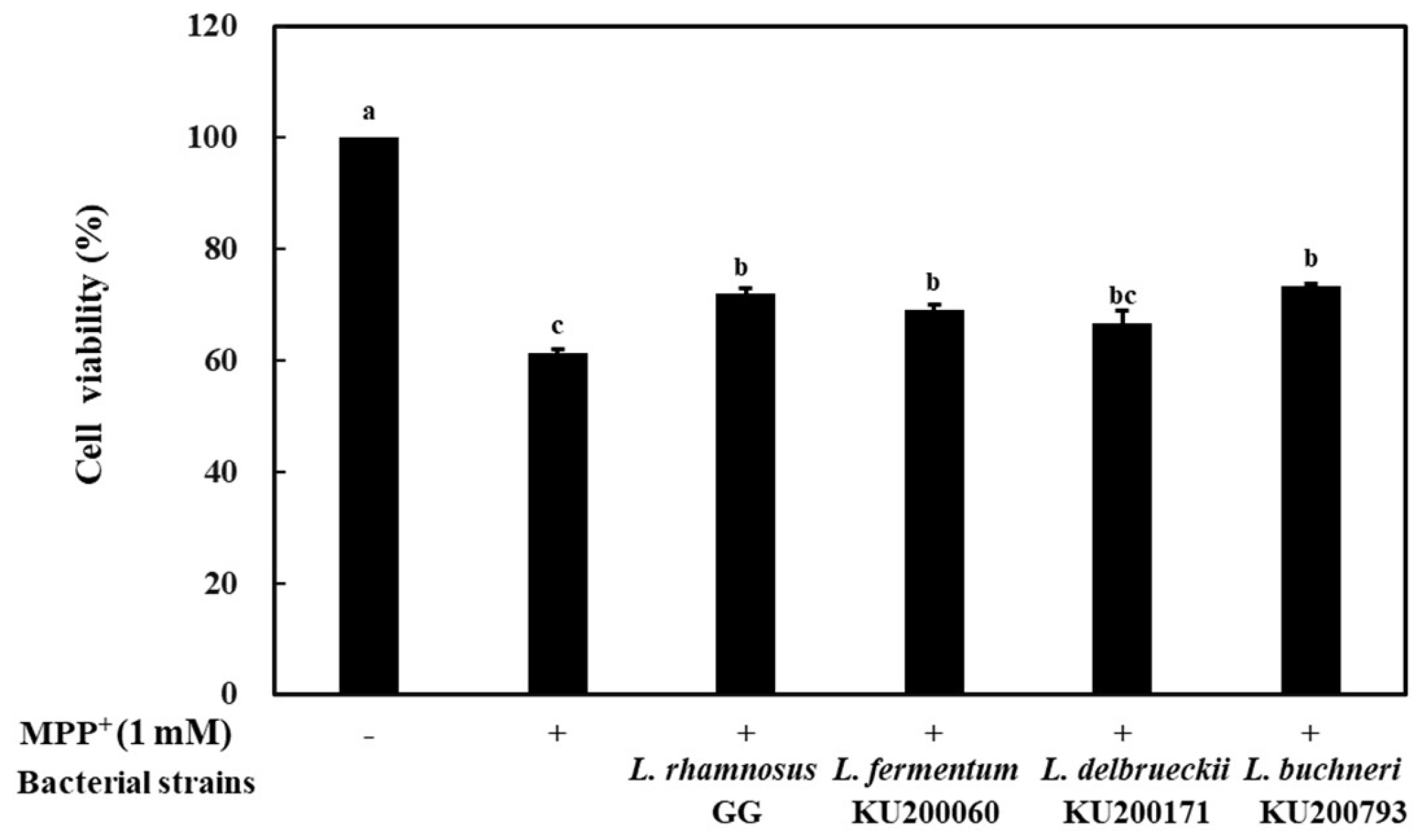
2.6. Neuroprotective Effects of CM Against MPP+ -Induced Cell Death
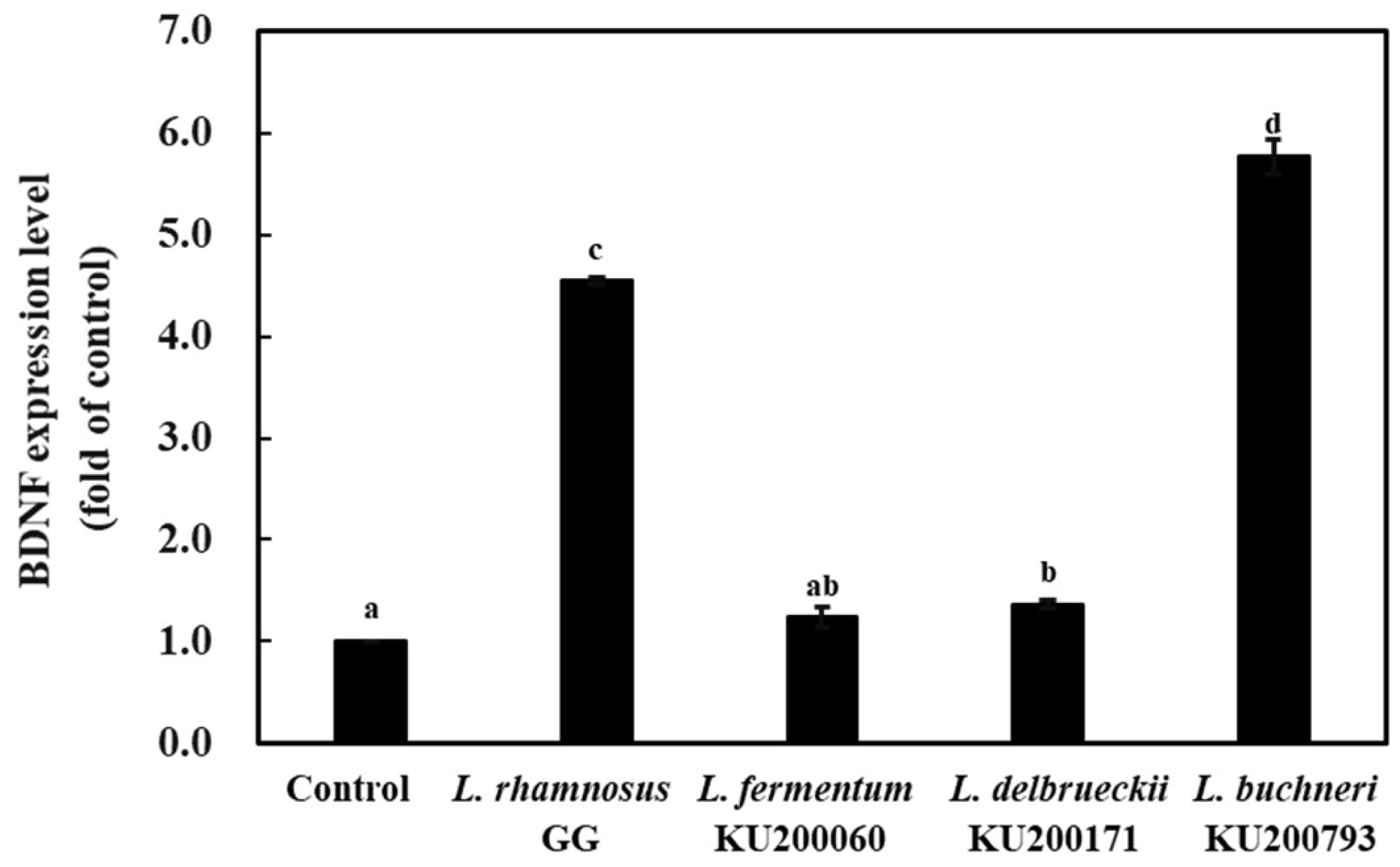
2.7. mRNA Level of the Brain-Derived Neurotropic Factor (BDNF) in HT-29 Cells Treated with Heat-Killed LAB Strains
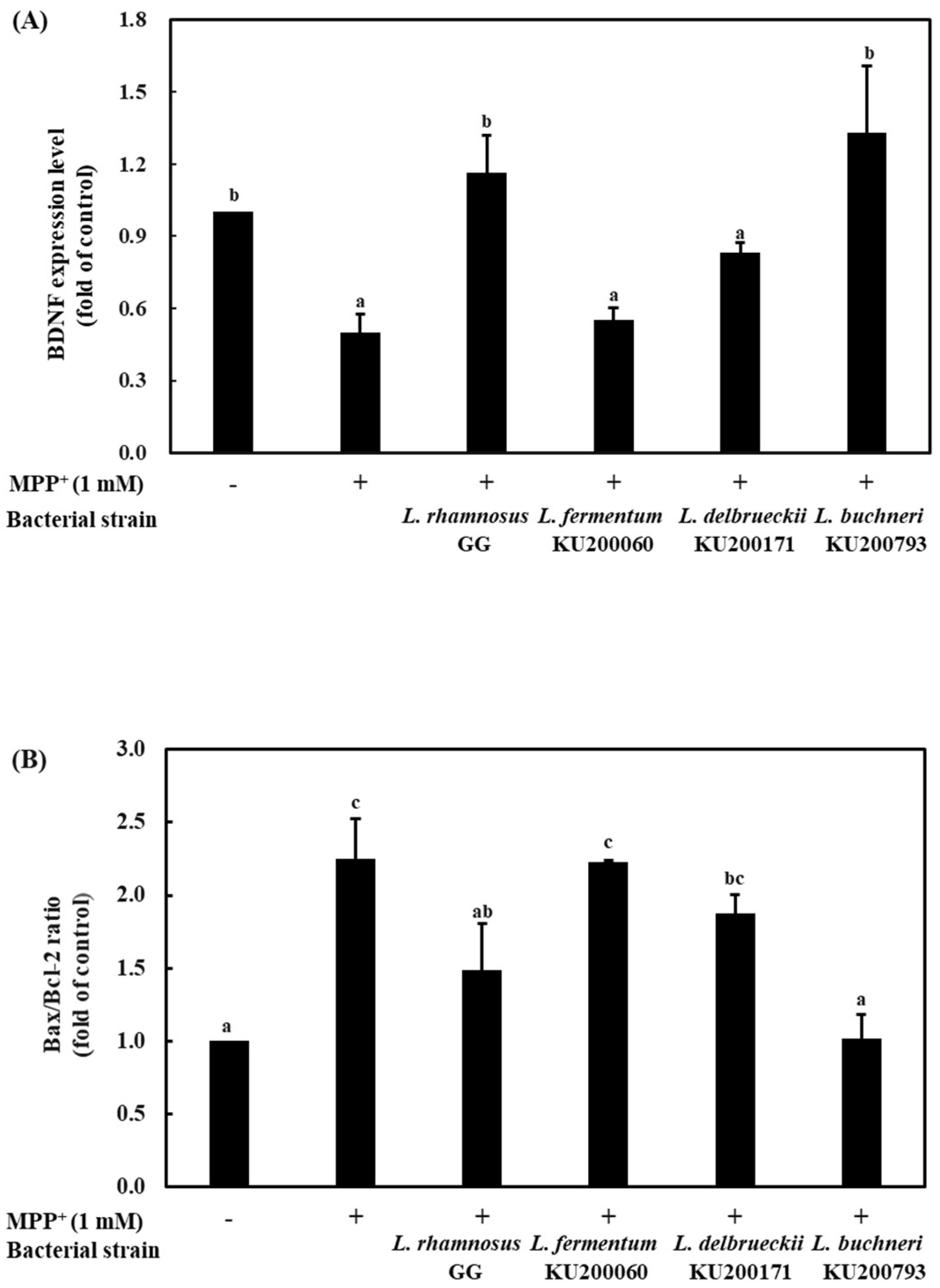
2.8. Bax/Bcl-2 and BDNF Expression Levels of CM in MPP+ Stressed SH-SY5Y Cells
3. Materials and Methods
3.1. Bacterial Strains Culture Condition
3.2. Cell Culture Condition
3.3. Tolerance to Artificial Gastric Conditions
3.4. Adhesion Ability to HT-29 Cells
3.5. Enzyme Production Evaluated Using API ZYM Kit
3.6. Antibiotic Susceptibility
3.7. Preparation of Heat-Killed LAB
3.8. Antioxidant Activity of Heat-Killed LAB Strains
3.9. Preparation of LAB-Conditioned Medium (CM)
3.10. Cytotoxicity Measurement
3.10.1. Cytotoxicity Effect of CM on SH-SY5Y Cell
3.10.2. Neuroprotective Effect of CM on MPP+-stressed SH-SY5Y Cells.
3.11. Relative Quantification of Gene Expression by RT-PCR
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MPP+ | 1-methyl-4-phenylpyridinium |
| GBA | gut-brain-axis |
| PD | Parkinson’s disease |
| LAB | lactic acid bacteria |
| BDNF | brain-derived neurotropic factor |
| GABA | γ-aminobutyric acid |
| CM | conditioned medium |
| ROS | reactive oxygen species |
| TrkB | tyrosine kinase B |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ABTS | 2,2′-azinobis (2-ethylbenzothiazoline-6-sulfonate) |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
References
- Mary, E.S. Probiotics: Considerations for human health. Nutr. Rev. 2003, 61, 91–99. [Google Scholar]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A Novel Class of Psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Perez-Pardo, P.; Kliest, T.; Dodiya, H.B.; Broersen, L.M.; Garssen, J.; Keshavarzian, A.; Kraneveld, A.D. The gut-brain axis in Parkinson’s disease: Possibilities for food-based therapies. Eur. J. Pharmacol. 2017, 817, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Warda, A.K.; Rea, K.; Fitzgerald, P.; Hueston, C.; Gonzalez-Tortuero, E.; Dinan, T.G.; Hill, C. Heat-killed lactobacilli alter both microbiota composition and behavior. Behav. Brain Res. 2019, 362, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Fukushima, H.; Nakakita, Y.; Kado, H.; Kida, S. Dietary heat-killed Lactobacillus brevis SBC8803 (SBL88™) improves hippocampus-dependent memory performance and adult hippocampal neurogenesis. Neuropsychopharmacol. Rep. 2019, 39, 140–145. [Google Scholar]
- Maehata, H.; Kobayashi, Y.; Mitsuyama, E.; Kawase, T.; Kuhara, T.; Xiao, J.Z.; Tsukahara, T.; Toyoda, A. Heat-killed Lactobacillus helveticus strain MCC1848 confers resilience to anxiety or depression-like symptoms caused by subchronic social defeat stress in mice. Biosci. Biotechnol. Biochem. 2019, 83, 1239–1247. [Google Scholar] [CrossRef]
- Sherman, M.P.; Zaghouani, H.; Niklas, V. Gut microbiota, the immune system, and diet influence the neonatal gut-brain axis. Pediatr. Res. 2014, 77, 127–135. [Google Scholar] [CrossRef]
- Foster, J.; McVey Neufeld, K.A. Gut–brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Houser, M.C.; Tansey, M.G. The gut-brain axis: Is intestinal inflammation a silent driver of Parkinson’s disease pathogenesis? NPJ Parkinsons Dis. 2017, 3, 3. [Google Scholar] [CrossRef]
- Jang, H.J.; Song, M.W.; Lee, N.K.; Paik, H.D. Antioxidant effects of live and heat-killed probiotic Lactobacillus plantarum Ln1 isolated from kimchi. J. Food Sci. Technol. 2018, 55, 3174–3180. [Google Scholar] [CrossRef]
- Son, S.H.; Yang, S.J.; Jeon, H.L.; Yu, H.S.; Lee, N.K.; Park, Y.S.; Paik, H.D. Antioxidant and immunostimulatory effect of potential probiotic Lactobacillus paraplantarum SC61 isolated from Korean traditional fermented food, jangajji. Microb. Pathog. 2018, 125, 486–492. [Google Scholar] [CrossRef]
- Lee, N.K.; Kim, S.Y.; Han, K.J.; Eom, S.J.; Paik, H.D. Probiotic potential of Lactobacillus strains with anti-allergic effects from kimchi for yogurt starters. LWT Food Sci. Technol. 2014, 58, 130–134. [Google Scholar] [CrossRef]
- Lee, N.K.; Han, K.J.; Son, S.H.; Eom, S.J.; Lee, S.K.; Paik, H.D. Multifunctional effect of probiotic Lactococcus lactis KC24 isolated from kimchi. LWT Food Sci. Technol. 2015, 64, 1036–1041. [Google Scholar] [CrossRef]
- Kim, B.K.; Song, J.L.; Ju, J.H.; Kang, S.A.; Park, K.Y. Anticancer effects of kimchi fermented for different times and with added ingredients in human HT-29 colon cancer cells. Food Sci. Biotechnol. 2015, 24, 629–633. [Google Scholar] [CrossRef]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of gamma-aminobutyric acid (GABA) by Lactobacillus buchneri isolated from kimchi and its neuroprotective effect on neuronal cells. J. Microbiol. Biotechn. 2007, 17, 104–109. [Google Scholar]
- Zeng, X.Q.; Pan, D.D.; Guo, Y.X. The probiotic properties of Lactobacillus buchneri P2. J. Appl. Microbiol. 2009, 108, 2059–2066. [Google Scholar]
- Fröhlich-Wyder, M.T.; Guggisberg, D.; Badertscher, R.; Wechsler, D.; Wittwer, A.; Irmler, S. The effect of Lactobacillus buchneri and Lactobacillus parabuchneri on the eye formation of semi-hard cheese. Int. Dairy J. 2013, 33, 120–128. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Jang, H.J.; Lee, N.K.; Paik, H.D. Probiotic characterization of Lactobacillus brevis KU15153 showing antimicrobial and antioxidant effect isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 1521–1528. [Google Scholar] [CrossRef]
- Gharbi, Y.; Fhoula, I.; Ruas-Madiedo, P.; Afef, N.; Boudabous, A.; Gueimonde, M.; Ouzari, H.I. In-vitro characterization of potentially probiotic Lactobacillus strains isolated from human microbiota: Interaction with pathogenic bacteria and the enteric cell line HT-29. Ann. Microbiol. 2019, 69, 61–72. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Pandiella, S.S.; Webb, C. Evaluation of the effect of malt, wheat and barley extracts on the viability of potentially probiotic lactic acid bacteria under acidic conditions. Int. J. Food Microbiol. 2003, 82, 133–141. [Google Scholar] [CrossRef]
- Patel, A.K.; Singhania, R.R.; Pandey, A.; Chincholkar, S.B. Probiotic bile salt hydrolase: Current developments and perspectives. Appl. Biochem. Biotechnol. 2009, 162, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Kõll, P.; Mändar, R.; Smidt, I.; Hütt, P.; Truusalu, K.; Mikelsaar, R.H.; Shchepetova, J.; Krogh-Andersen, K.; Marcotte, H.; Hammarström, L.; et al. Screening and evaluation of human intestinal lactobacilli for the development of novel gastrointestinal probiotics. Curr. Microbiol. 2010, 61, 560–566. [Google Scholar] [CrossRef]
- Shin, H.J.; Choi, H.J.; Kim, D.W.; Ahn, C.S.; Lee, Y.G.; Jeong, Y.K.; Joo, W.H. Probiotic potential of Pediococcus pentosaceus BCNU 9070. J. Life Sci. 2012, 22, 1194–1200. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria-a review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef]
- Son, S.H.; Jeon, H.L.; Jeon, E.B.; Lee, N.K.; Park, Y.S.; Kang, D.K.; Paik, H.D. Potential probiotic Lactobacillus plantarum Ln4 from kimchi: Evaluation of β-galactosidase and antioxidant activities. LWT Food Sci. Technol. 2017, 85, 181–186. [Google Scholar] [CrossRef]
- Morales, G.; Paredes, A. Antioxidant activities of Lampaya medicinalis extracts and their main chemical constituents. BMC Complement. Altern. Med. 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.Y.; Chang, F.J. Antioxidative effect of intestinal bacteria Bifidobacterium longum ATCC 15708 and Lactobacillus acidophilus ATCC 4356. Dig. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Fan, M.; Liao, Q. Study on effect elements of exopolysaccharide production of Lactobacillus kimchi SR8 and DPPH radical scavenging activity. J. Food Nutr. Res. 2017, 5, 928–934. [Google Scholar] [CrossRef] [Green Version]
- An, H.; Kim, I.S.; Koppula, S.; Kim, B.W.; Park, P.J.; Lim, B.O.; Choi, W.S.; Lee, K.H.; Choi, D.K. Protective effects of Gastrodia elata Blume on MPP+-induced cytotoxicity in human dopaminergic SH-SY5Y cells. J. Ethnopharmacol. 2010, 130, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Michael, D.R.; Davies, T.S.; Loxley, K.E.; Allen, M.D.; Good, M.A.; Hughes, T.R.; Plummer, S.F. In vitro neuroprotective activities of two distinct probiotic consortia. Benef. Microbes. 2019, 10, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szapacs, M.E.; Mathews, T.A.; Tessarollo, L.; Ernest Lyons, W.; Mamounase, L.A.; Andrews, A.M. Exploring the relationship between serotonin and brain-derived neurotrophic factor: Analysis of BDNF protein and extra neuronal 5-HT in mice with reduced serotonin transporter or BDNF expression. J. Neurosci. Methods. 2004, 140, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, J.P.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Lee, J.Y.; Yeom, Z.; Heo, D.H.; Lim, Y.H. Neuroprotective effect of Ruminococcus albus on oxidatively stressed SH-SY5Y cells and animals. Sci. Rep. 2017, 7, 14520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howells, D.W.; Porritt, M.J.; Wong, J.Y.; Batchelor, P.E.; Kalnins, R.; Hughes, A.J.; Donnan, G.A. Reduced BDNF mRNA expression in the Parkinson’s disease substantia nigra. Exp. Neurol. 2000, 166, 127–135. [Google Scholar] [CrossRef]
- Cheng, R.; Xu, T.; Zhang, Y.; Wang, F.; Zhao, L.; Jiang, Y.; He, F. Lactobacillus rhamnosus GG and Bifidobacterium bifidum TMC3115 Can Affect Development of Hippocampal Neurons Cultured In Vitro in a Strain-Dependent Manner. Probiotics Antimicrob. Proteins 2019, 1–11. [Google Scholar] [CrossRef]
- Crompton, M. Bax, Bid and the permeabilization of the mitochondrial outer membrane in apoptosis. Curr. Opin. Cell Biol. 2000, 12, 414–419. [Google Scholar] [CrossRef]
- Jung, H.W.; Jin, G.Z.; Kim, S.Y.; Kim, Y.S.; Park, Y.K. Neuroprotective effect of methanol extract of Phellodendri cortex against 1-methyl-4-phenylpyridinium (MPP+)-induced apoptosis in PC-12 cells. Cell Biol. Int. 2009, 33, 957–963. [Google Scholar] [CrossRef]
- Jung, T.W.; Lee, J.Y.; Shim, W.S.; Kang, E.S.; Kim, J.S.; Ahn, C.W.; Lee, H.C.; Cha, B.S. Adiponectin protects human neuroblastoma SH-SY5Y cells against MPP+-induced cytotoxicity. Biochem. Biophys. Res. Commun. 2006, 343, 564–570. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| LAB Strains | Survival Rate (%) | Adhesion Ability to HT-29 Cells (%) | |
|---|---|---|---|
| Gastric Acid Tolerance (0.3% Pepsin, pH 2.5) | Bile Salts Tolerance (0.3% Oxgall) | ||
| L. rhamnosus GG | 99.68 ± 0.26 c (1) | 100.41 ± 0.15 b | 2.34 ± 0.15 b |
| L. fermentum KU200060 | 100.47 ± 0.07 b | 102.24 ± 0.70 a | 1.18 ± 0.04 c |
| L. delbrueckii KU200171 | 62.68 ± 0.12 d | 102.43 ± 0.50 a | 1.83 ± 0.01 bc |
| L. buchneri KU200793 | 102.12 ± 0.35 a | 91.47 ± 0.46 c | 14.10 ± 0.45 a |
| Enzyme | Enzyme Activity | |||
|---|---|---|---|---|
| L. rhamnosus GG | L. fermentum KU200060 | L. delbrueckii KU200171 | L. buchneri KU200793 | |
| Control | 0 (1) | 0 | 0 | 0 |
| Alkaline phosphate | 0 | 0 | 0 | 0 |
| Esterase | 2 | 1 | 0 | 0 |
| Esterase lipase | 1 | 1 | 0 | 0 |
| Lipase | 0 | 0 | 0 | 0 |
| Leucine arylamidase | 3 | 2 | 4 | 3 |
| Valine arylamidase | 3 | 0 | 3 | 2 |
| Cystine arylamidase | 0 | 0 | 0 | 0 |
| Trypsin | 0 | 0 | 0 | 0 |
| α-Chymotrypsin | 0 | 0 | 0 | 0 |
| Acid phosphatase | 1 | 1 | 1 | 0 |
| Naphthol-AS-BI-phosphohydrolase | 2 | 1 | 2 | 1 |
| α-Galactosidase | 0 | 4 | 0 | 0 |
| β-Galactosidase | 0 | 5 | 0 | 4 |
| β-Glucuronidase | 0 | 0 | 0 | 0 |
| α-Glucosidase | 3 | 1 | 0 | 0 |
| β-Glucosidase | 4 | 0 | 0 | 1 |
| N-Acetyl-β-glucosaminidase | 0 | 0 | 0 | 2 |
| α-Mannosidase | 0 | 0 | 0 | 0 |
| α-Fucosidase | 0 | 0 | 0 | 0 |
| Antioxidant Activity | LAB Strains | |||
|---|---|---|---|---|
| L. rhamnosus GG | L. fermentum KU200060 | L. delbrueckii KU200171 | L. buchneri KU200793 | |
| Ampicillin | S (1) | S | S | S |
| Gentamicin | R | R | R | R |
| Kanamycin | R | R | R | R |
| Streptomycin | R | R | R | R |
| Tetracycline | S | S | S | S |
| Ciprofloxacin | R | R | R | R |
| Chloramphenicol | S | S | S | S |
| Doxycycline | S | S | S | S |
| Antioxidant Activity | LAB Strains | |||
|---|---|---|---|---|
| L. rhamnosus GG | L. fermentum KU200060 | L. delbrueckii KU200171 | L. buchneri KU200793 | |
| DPPH radical scavenging activity (%) | 23.76 ± 1.53 a (1) | 21.81 ± 1.62 a | 17.20 ± 0.48 b | 23.04 ± 0.88 a |
| ABTS radical scavenging activity (%) | 90.92 ± 1.91 a | 91.87 ± 3.27 a | 68.33 ± 1.91 b | 90.05 ± 3.27 a |
| Inhibition rate of β-carotene and linoleic acid oxidation (%) | 33.63 ± 2.60 ab | 28.49 ± 2.79 b | 16.09 ± 2.13 c | 38.42 ± 3.56 a |
| Primer (1) | Sequence (5′-3′) | Reference |
|---|---|---|
| BDNF | (sense) CAAACATCCGAGGACAAGGTGG | [35] |
| (antisense) CTCATGGACATGTTTGCAGCATCT | ||
| Bax | (sense) GTGGTTGCCCTCTTCTACTTTGC | |
| (antisense) GAGGACTCCAGCCACAAAGATG | ||
| Bcl-2 | (sense) CGGCTGAAGTCTCCATTAGC | |
| (antisense) CCAGGGAAGTTCTGGTGTGT | ||
| GAPDH | (sense) GAGTCAACGGATTTGGTCGT | |
| (antisense) GACAAGCTTCCCGTTCTCAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheon, M.-J.; Lim, S.-M.; Lee, N.-K.; Paik, H.-D. Probiotic Properties and Neuroprotective Effects of Lactobacillus buchneri KU200793 Isolated from Korean Fermented Foods. Int. J. Mol. Sci. 2020, 21, 1227. https://doi.org/10.3390/ijms21041227
Cheon M-J, Lim S-M, Lee N-K, Paik H-D. Probiotic Properties and Neuroprotective Effects of Lactobacillus buchneri KU200793 Isolated from Korean Fermented Foods. International Journal of Molecular Sciences. 2020; 21(4):1227. https://doi.org/10.3390/ijms21041227
Chicago/Turabian StyleCheon, Min-Jeong, Sung-Min Lim, Na-Kyoung Lee, and Hyun-Dong Paik. 2020. "Probiotic Properties and Neuroprotective Effects of Lactobacillus buchneri KU200793 Isolated from Korean Fermented Foods" International Journal of Molecular Sciences 21, no. 4: 1227. https://doi.org/10.3390/ijms21041227
APA StyleCheon, M.-J., Lim, S.-M., Lee, N.-K., & Paik, H.-D. (2020). Probiotic Properties and Neuroprotective Effects of Lactobacillus buchneri KU200793 Isolated from Korean Fermented Foods. International Journal of Molecular Sciences, 21(4), 1227. https://doi.org/10.3390/ijms21041227