Calmodulin and Calmodulin Binding Proteins in Dictyostelium: A Primer


Abstract
1. Introduction
Dictyostelium Calcium Binding Proteins
2. Dictyostelium Versus Human Calmodulin
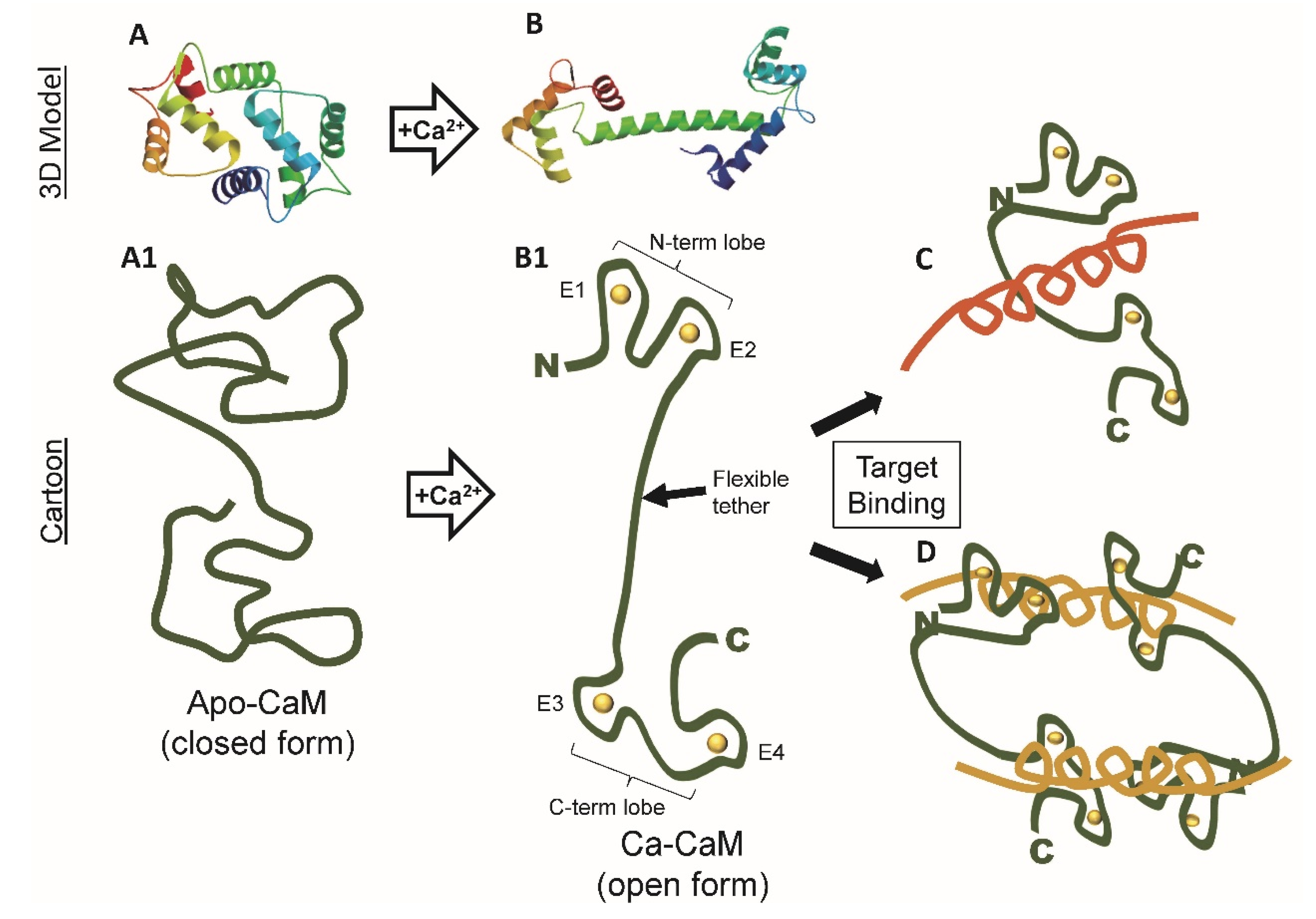
3. Calcium-Binding Causes Dramatic Conformational Changes in Calmodulin
4. Calcium-Dependent Calmodulin Binding
Calmodulin Binding Domains and Motifs
5. Calcium-Independent Calmodulin Binding via IQ Motifs
6. Detecting Calmodulin-Binding Proteins
7. Calmodulin Antagonists Provide Some Insight
8. Dictyostelium as a Potential Model to Study Calmodulin Mutations
9. CaM Kinases and Calcineurin
10. Conclusions, Comments, and Speculation
Funding
Conflicts of Interest
References
- Cai, X.; Wang, X.; Patel, S.; Clapham, D.E. Insights into the early evolution of animal calcium signaling machinery: A unicellular point of view. Cell Calcium 2015, 57, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Krebs, J. Why calcium? How calcium became the best communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [PubMed]
- Marchadier, E.; Oates, M.E.; Fang, H.; Donoghue, P.C.J.; Hetherington, A.M.; Gough, J. Evolution of the calcium-based intracellular signaling system. Genome Biol. Evol. 2016, 8, 2118–2132. [Google Scholar] [CrossRef] [PubMed]
- Plattner, H.; Verkhratsky, A. Evolution of calcium signaling. Cell Calcium 2015, 57, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Aubry, L.; Mattei, S.; Blot, B.; Sadoul, R.; Satre, M.; Klein, G. Biochemical characterization of two analogues of the apoptosis-linked gene 2 protein in Dictyostelium discoideum and interaction with a physiological partner in mammals, murine Alix. J. Biol. Chem. 2002, 277, 21947–21954. [Google Scholar] [CrossRef] [PubMed]
- Damer, C.K.; Bayeva, M.; Kim, P.S.; Ho, L.K.; Eberhardt, E.S.; Socec, C.I.; Lee, J.S.; Bruce, A.E.; Goldman-Yassen, E.A.; Naliboff, L.C. Copine A is required for cytokinesis, contractile vacuole function, and development in Dictyostelium. Euk. Cell 2007, 6, 430–442. [Google Scholar] [CrossRef]
- Rösel, D.; Půta, F.; Blahůšková, A.; Smýkal, P.; Folk, P. Molecular characterization of a calmodulin-like Dictyostelium protein CalB. FEBS Lett. 2000, 473, 323–327. [Google Scholar] [CrossRef]
- Kollmar, M. Thirteen is enough: The myosins of Dictyostelium and their light chains. BMC Genom. 2006, 7, 183. [Google Scholar] [CrossRef]
- Hoeflich, K.P.; Ikura, M. Calmodulin in action. Cell 2002, 108, 739–742. [Google Scholar] [CrossRef]
- Kelly, K.L.; Dalton, S.R.; Wai, R.B.; Ramchandani, K.; Xu, R.J.; Linse, S.; Londergan, C.H. Conformational ensembles of calmodulin revealed by nonperturbing site-specific vibrational probe groups. J. Phys.Chem. 2018, 122, 2917–2955. [Google Scholar] [CrossRef]
- Berchtold, M.W.; Villalobo, A. The many faces of calmodulin in cell proliferation, programmed cell death, autophagy and cancer. Biochim. Biophys. Acta 2014, 1843, 398–435. [Google Scholar] [CrossRef] [PubMed]
- O’Day, D.H.; Keszei, A. Signalling and sex in the social amoebozoans. Biol. Rev. 2012, 87, 313–329. [Google Scholar] [CrossRef] [PubMed]
- O’Day, D.H.; Sabateeshan, M.; Myre, M.A.; Huber, R.J. Calmodulin-mediated events during the life cycle of the amoebozoan Dictyostelium discoideum. Biol. Rev. 2019. (ACCEPTED). [Google Scholar]
- Catalano, A.; O’Day, D.H. Calmodulin-binding proteins in the model organism Dictyostelium: A complete & critical review. Cell. Signal. 2008, 20, 277–291. [Google Scholar] [PubMed]
- Clarke, M.; Bazari, W.L.; Kayman, S.C. Isolation and properties of calmodulin of Dictyostelium discoideum. J. Bacteriol. 1980, 141, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Bazari, W.L.; Clarke, M. Characterization of a novel calmodulin from Dictyostelium discoideum. J. Biol. Chem. 1981, 256, 3598–3603. [Google Scholar]
- Marshak, D.R.; Clarke, M.; Roberts, D.M.; Watterson, D.M. Structural and functional properties of calmodulin from the eukaryotic microorganism Dictyostelium discoideum. Biochemistry 1984, 23, 1892–1899. [Google Scholar] [CrossRef]
- Villalobo, A. The multifunctional role of phospho-calmodulin in pathophysiological processes. Biochem. J. 2018, 475, 4011–4023. [Google Scholar] [CrossRef]
- Urrutia, J.; Aguado, A.; Muguruza-Montero, A.; Nunez, E.; Malo, C.; Casis, O.; Villarroel, A. The crossroad of ion channels and calmodulin in disease. Int. J. Molec. Sci. 2019, 20, 400. [Google Scholar] [CrossRef]
- Friedberg, F.; Rhoads, A.R. Evolutionary aspects of calmodulin. Life 2001, 51, 215–221. [Google Scholar]
- Halling, D.B.; Liebeskind, B.J.; Hall, A.W.; Aldrich, R.W. Conserved properties of individual Ca2+-binding sites in calmodulin. Proc. Natl. Acad. Sci. USA 2016, 113, E1216–E1225. [Google Scholar] [CrossRef]
- Faas, G.C.; Raghavachari, S.; Lisman, J.E.; Mody, I. Calmodulin as a direct detector of Ca2+ signals. Nat. Neurosci. 2011, 14, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Tidow, H.; Nissen, P. Structural diversity of calmodulin binding to its target sites. FEBS J. 2013, 280, 5551–5565. [Google Scholar] [CrossRef] [PubMed]
- Westerlund, A.M.; Delemotte, L. Effect of Ca2+ on the promiscuous target-protein binding of calmodulin. PLoS Comput. Biol. 2018, 14, e1006072. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.B. IQGAP family members in yeast, Dictyostelium and mammalian cells. Int. J. Cell Biol. 2012, 894817. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.J.; O’Day, D.H. Extracellular matrix dynamics and functions in the social amoeba Dictyostelium: A critical review. Biochimica et Biophysica Acta 2017, 1861, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Myre, M.A.; O’Day, D.H. Calmodulin binds to and inhibits the activity of phosphoglycerate kinase. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1693, 177–183. [Google Scholar] [CrossRef]
- O’Day D., H. CaMBOT: Profiling and characterizing calmodulin binding proteins. Cell. Signal. 2003, 15, 347–355. [Google Scholar] [CrossRef]
- Myre, M.A.; O’Day, D.H. Dictyostelium calcium-binding protein 4a interacts with nucleomorphin, a BRCT-domain protein that regulates nuclear number. Biochem. Biophys. Res. Commun. 2004, 322, 665–671. [Google Scholar] [CrossRef]
- Shukla, D.; Peck, A.; Pande, V.S. Conformational heterogeneity of the calmodulin binding interface. Nature Commun. 2016, 7, 10910. [Google Scholar] [CrossRef]
- Yap, K.L.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Func. Genom. 2000, 1, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Bahler, M.; Rhoads, A. Calmodulin signaling via the IQ motif. FEBS Lett. 2002, 513, 107–113. [Google Scholar] [CrossRef]
- Heissler, S.M.; Sellers, J.R. Myosin light chains: Teaching old dogs new tricks. BioArchitecture 2014, 4, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Crawley, S.W.; Liburd, L.; Shaw, K.; Jung, Y.; Smith, S.P.; Coté, G.P. Identification of calmodulin and MlcC as light chains for Dictyostelium myosin-1 isozymes. Biochem. 2011, 50, 6579–6588. [Google Scholar] [CrossRef]
- Langelaan, D.N.; Liburd, J.; Yang, Y.; Miller, E.; Chitayat, S.; Crawley, S.W.; Côté, G.P.; Smith, S.P. Structure of the single-lobe myosin light chain C in complex with the light chain-binding domains of myosin-1-C provides insights into divergent IQ motif recognition. J. Biol. Chem. 2016, 291, 19607–19617. [Google Scholar] [CrossRef]
- Vlahou, G.; Rivero, F. Rho GTPase signaling in Dictyostelium discoideum: Insights from the genome. Eur. J. Cell Biol. 2006, 85, 947–959. [Google Scholar] [CrossRef]
- Dammann, H.; Hellstern, S.; Husain, Q.; Mutzel, R. Primary structure, expression and developmental regulation of a Dictyostelium calcineurin A homologue. Eur. J. Biochem. 1996, 238, 391–399. [Google Scholar] [CrossRef]
- Malchow, D.; Mutzel, R.; Schlatterer, C. On the role of calcium during chemotactic signalling and differentiation of the cellular slime mould Dictyostelium discoideum. Int. J. Develop. Biol. 1996, 40, 135–139. [Google Scholar]
- Lydan, M.A.; O’Day, D.H. 35S labelling of recombinant calmodulin in E. coli. Methods Mol. Biol. Protoc. Gene Anal. 1994, 31, 389–396. [Google Scholar]
- Gauthier, M.L.; O’Day, D.H. Detection of calmodulin-binding proteins and calmodulin-dependent phosphorylation linked to calmodulin-dependent chemotaxis to folic and cAMP in Dictyostelium. Cell. Signal. 2001, 13, 575–584. [Google Scholar] [CrossRef]
- Catalano, A.; Poloz, Y.; O’Day, D.H. Dictyostelium puromycin-sensitive aminopeptidase A is a nucleoplasmic nucleomorphin-binding protein that relocates to the cytoplasm during mitosis. Histochem. Cell Biol. 2011, 136, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Catalano, A.C.; O’Day, D.H. Rad53 homologue forkhead-associated kinase A (FhkA) and Ca2+-binding protein 4a (CBP4a) are nucleolar proteins that differentially redistribute during mitosis in Dictyostelium. Cell Division 2013, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.J.; Catalano, A.; O’Day, D.H. Cyclin-dependent kinase 5 is a calmodulin-binding protein that associates with puromycin-sensitive aminopeptidase in the nucleus of Dictyostelium, Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 11–20. [Google Scholar] [CrossRef] [PubMed][Green Version]
- O’Day, D.H.; Huber, R.J.; Suarez, A. Extracellular calmodulin regulates growth and cAMP-mediated chemotaxis in Dictyostelium discoideum. Biochem. Biophys. Res. Commun. 2012, 425, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, A.; Uyeda, T.Q.P. DWWA, a novel protein containing two WW domains and an IQ motif, is required for scission of the residual cytoplasmic bridge during cytokinesis in Dictyostelium. Mol. Biol. Cell 2004, 15, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.; Huber, R.; Myre, M.; O’Day, D.H. An extracellular matrix, calmodulin-binding protein with EGF-Like repeats that enhance cell motility. Cell. Signal. 2011, 23, 1197–1206. [Google Scholar] [CrossRef]
- Sriskanthadevan, S.; Zhu, Y.; Manoharan, K.; Yang, C.; Siu, C.H. The cell adhesion molecule DdCAD-1 regulates morphogenesis through differential spatiotemporal expression in Dictyostelium discoideum. Development 2011, 138, 2487–2497. [Google Scholar] [CrossRef]
- Annesley, S.J.; Bago, R.; Bosnar, M.; Filic, V.; Marinović, M.; Weber, I.; Mehta, A.; Fisher, P.R. Dictyostelium discoideum nucleoside diphosphate kinase C plays a negative regulatory role in phagocytosis, macropinocytosis and exocytosis. PLoS ONE 2011, 6, e26024. [Google Scholar] [CrossRef]
- Myre, M.A.; Lumsden, A.L.; Thompson, M.N.; Wasco, W.; MacDonald, M.E.; Gusella, J.F. Deficiency of huntingtin has pleiotropic effects in the social amoeba Dictyostelium discoideum. PLoS Genet. 2011, 7, 4. [Google Scholar] [CrossRef]
- Lydan, M.A.; Cotter, D.A.; O’Day, D.H. Calmodulin function and calmodulin-binding proteins during autoactivation and spore germination in Dictyostelium discoideum. Cell. Signal. 1994, 6, 751–762. [Google Scholar] [CrossRef]
- Gröner, M.; Malchow, D. Calmodulin-antagonists inhibit vesicular Ca2+ uptake in Dictyostelium. Cell Calcium 1996, 19, 105–111. [Google Scholar] [CrossRef]
- Malchow, D.; Lusche, D.F.; Schlatterer, C. A link of Ca2+ to cAMP oscillations in Dictyostelium: The calmodulin antagonist W-7 potentiates cAMP relay and transiently inhibits the acidic Ca2+-store. BMC Develop. Biol. 2004, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Sugden, C.; Urbaniak, M.D.; Araki, T.; Williams, J.G. The Dictyostelium prestalk inducer differentiation-inducing factor-1 (DIF-1) triggers unexpectedly complex global phosphorylation changes. Mol. Biol. Cell 2014, 26, 805–820. [Google Scholar] [CrossRef]
- Horn, F.; Gross, J. A role for calcineurin in Dictyostelium discoideum development. Differentiation 1996, 60, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Kobel-Höller, K.; Gley, K.; Jochinke, J.; Heider, K.; Fritsch, V.N.; Nguyen, H.V.D.; Lischke, T.; Radek, R.; Baugrass, R.; Mutzel, R.S.; et al. Calcineurin silencing in Dictyostelium discoideum leads to cellular alterations affecting mitochondria, gene expression, and oxidative stress response. Protist 2018, 169, 584–602. [Google Scholar] [CrossRef]
- Nyegaard, M.; Overgaard, M.T.; Søndergaard, M.T.; Vranas, M.; Behr, E.R.; Hindlegrandt, L.L.; Lund, J.; Hedley, P.L.; Camm, A.J.; Wettrell, G.; et al. Mutations in calmodulin cause ventricular tachycardia and sudden cardiac death. Am. J. Hum. Genet. 2012, 91, 703–712. [Google Scholar] [CrossRef]
- Marsman, R.F.; Barc, J.; Beekman, L.; Alders, M.; Dooijes, D.; van den Wijngaard, A.; Ratbi, I.; Sefiani, A.; Bhuiyan, Z.A.; Wilde, A.A.; et al. A mutation in CALM1 encoding calmodulin in familial idiopathic ventricular fibrillation in childhood and adolescence. J. Am. Coll. Cardiol. 2014, 63, 259–266. [Google Scholar] [CrossRef]
- Wang, K.; Holt, C.; Lu, J.; Brohus, M.; Larsen, K.T.; Overgaard, M.T.; Wimmer, R.; Van Petegem, F. Arrhythmia mutations in calmodulin cause conformational changes that affect interactions with the cardiac voltage-gated calcium channel. Proc. Natl. Acad. Sci. USA 2018, 115, E10556–E10565. [Google Scholar] [CrossRef]
- Jensen, H.H.; Brohus, M.; Nyegaard, M.; Overgaard, M.T. Human calmodulin mutations. Front. Mol. Neurosci. 2018, 11, 396–404. [Google Scholar] [CrossRef]
- Iriki, H.; Kawata, T.; Muramoto, T. Generation of deletions and precise point mutations in Dictyostelium discoideum using the CRISPR nickase. PLoS ONE 2019, 14, e0224128. [Google Scholar] [CrossRef]
- Brzozowski, J.S.; Skelding, K.A. The multi-functional calcium/calmodulin stimulated protein kinase (CaMK) family: Emerging targets for anti-cancer therapeutic intervention. Pharmaceuticals 2019, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M.; Manning, G.; Liu, A.; Fey, P.; Pilcher, K.E.; Xu, Y.; Smith, J.L. The Dictyostelium kinome—Analysis of the protein kinases from a simple model organism. PLoS Genet. 2006, 2, e38. [Google Scholar] [CrossRef] [PubMed]
- Betapudi, V.; Mason, C.; Licate, L.; Egelhoff, T.T. Identification and characterization of a novel α-kinase with a von Willebrand Factor A-like motif localized to the contractile vacuole and golgi complex in Dictyostelium discoideum. Mol. Biol. Cell 2005, 16, 2248–2262. [Google Scholar] [CrossRef] [PubMed]
- Swan, D.G.; Hale, R.S.; Dhillon, N.; Leadlay, P.F. A bacterial calcium-binding protein homologous to calmodulin. Nature 1987, 329, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Myre, M.A.; Huber, R.J.; O’Day, D.H. Functional analysis of proteins involved in neurodegeneration using the model organism Dictyostelium: Alzheimer’s, Huntington’s and Batten disease. In Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research; Wim, E.C., Robert, T.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 491–518. [Google Scholar]
- Liburd, J.; Chitayat, S.; Crawley, S.W.; Munro, K.; Miller, E.; Denis, C.M.; Spencer, H.L.; Côté, G.P.; Smith, S.P. Structure of the small Dictyostelium discoideum light chain MlcB provides insights into MyoB IQ motif recognition. J. Biol. Chem. 2014, 289, 17030–17042. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| I. CaMBPs with Canonical CaM-Binding Domains (CaMBDs). | ||||
|---|---|---|---|---|
| CaMBP | ORF/Dictybase | CaMBDs | Verified | Reference |
| Atg13 | DDB_G0269162 | 104FYKNVIILIRTIYAML129 | N | [13] |
| Atg101 | DDB_G0288287 | 72LTSIMKRKAKTAQISI86 | N | [13] |
| CanA/CAN | DDB0185021 | 445RQMLRAKVKSVSKMMRMFSLLRQE468 | Y | [26] |
| 529LLRMNSRGELFRINSKGDLF548 | N | [14] | ||
| Cdk5 | DDB_G0288677 | 17IVYKAKNRETGEIVALK33 | Y | [27] |
| 132LLINRKGELKLADFGLARAFGIP154 | N | [27] | ||
| cmbB | DDB0201645 | 179IPKSLRSLFLGKGYNQPLEF198 | N | [13] |
| DDB_G0285767 | DDB_G0285767 | 172FFEYRENLIKTANKSF191 | N | [13] |
| 240WYKHLKEEFSKVKLKV255 | N | [13] | ||
| Drainin | DDB_G0269130 | 380RTALSILRYFIS391 | N | [13] |
| fAR1 | DDB_G0281211 | 611CVLIIFGAKFWKIYKPVEDD630 | N | [13] |
| Htt | DDB_G0272344 | 1885LDLRKKQLLRLLSL1896 | N | [13] |
| IplA | DDB_G0292564 | 841VSKGRNYNGI850 | N | [13] |
| 850IRLVGQRITHKECL863 | N | [13] | ||
| MIHCK/pakB | DDB0191345 | 150AFRKAYHTLDLSKSGASGRY169 | N | [14] |
| MHCK-A/mhkA | DDB0216274 | 535FVSLARIVKINVGTREIRV553 | N | [14] |
| MHCK-B/mhkB | DDB0191333 | 150AFRKAYHTLDLSKSGASGRY169 | N | [14] |
| NdkC | DDB_G0273069 | 23GLVGEIIARYEKKGFVLVGLKQLV46 | N | [13] |
| NumA | DDB0231257 | 171EDVSRFIKGKLLQKQQKIYKDLERF195 | Y | [28] |
| PakA/ DPAKa | DDB0191313 | 291EIEKIQREERIKIEKEYDDK310 | N | [14] |
| PatA | DDB_G0277861 | 1064WQIVRQTHKKLVVINALKE1085 | N | [13] |
| PgkA | DDB0191349 | 209KPFLAILGGAKVSDKIKLIF228 | Y | [29] |
| PsaA | DDB_G0270994 | 108LSLVFTGLLNDKLKGFYRSKYTV130 | N | [30] |
| Rab11A | DDB_G0269238 | 69TAGQERYRAITSAYYRGAVGALLV92 | N | [13] |
| Rab11C | DDB_G0277101 | 68GQERFRAVTSGYYRGAVGAMI88 | N | [13] |
| RhgA | DDB_G0283389 | 425VILLQLKKIKGLKSKEY442 | N | [13] |
| RhgB | DDB_G0280059 | 431ILKALKKVGGLKAKQYY447 | N | [13] |
| rgaA | DDB0191437 | 267MIITYNKRKQGTDYLKAVIG286 | N | [14] |
| Rpl19 | DDB0214854 | 123LYLKAKGNVFKNKRTLIEYIV143 | N | [14] |
| ThyB/DdTK1 | DDB0191436 | 20GKTTELIRRIKRFNFANKKC30 | N | [13] |
| VwkA | DDB0216405 | 420LGIDEHGKKVVLKQSKYIGGR440 | N | [14] |
| 536SKNKAIVVDIQGVKTSKGYLL556 | N | [14] | ||
| II. CaMBPs with Non-Canonical CaMBDs | ||||
| Atg1 | DDB_G0292390 | 110EKALYFMKQLAN121 | N | [13] |
| Rab8A | DDB_G0280043 | 71TAGQERFRTITTAYYRGAMGI91 | N | [13] |
| VatM | DDB_G0291858 | 291DHKRQTLAGIV301 | N | [13] |
| Subclass | Motif | Proteins with Motif |
|---|---|---|
| 1-10 Subclasses | ||
| 1-10 | (FILVW)xxxxxxxx(FILVW) | Atg13; CanA; Cdk5; CmbB; fAR1; IplA; NdkC; PatA; PsaA; Rab11A; Rab11C; RhgA; RhgB; VwkA |
| 1-5-10 | (FILVW)xxx(FAILVW)xxxx(FILVW) | fAR1; NdkC; NumA; PgkA; Rab11a; Rab11a; RhgB; ThyB |
| Basic 1-5-10 | (RK)(RK)(RK)(FAILVW)xxx(FILV)xxxx(FILVW) | L9 |
| 1-12 Subclass | ||
| 1-12 | (FILVW)xxxxxxxxxxXXxx(FILVW) | Cdk5; PsaA; RhgB |
| 1-14 Subclasses | ||
| 1-14 | (FILVW)xxxxxxxxxxxx(FILVW) | Atg101; CanA; Cdk5; fAR1; IplA; NdkC; NumA; PatA; PsaA; Rab11a; Rab11a; VwkA |
| 1-8-14 | (FILVW)xxxxxx(FAILVW)xxxxx(FILVW) | Htt; L9; PgkA; Rab11a |
| Basic1-8-14 | (RK)(RK)(RK)(FILVW)xxxxxx(FAILVW)xxxxx(FILVW) | none? |
| 1-5-8-14 | (FILVW)xxx(FAILVW)xx(FAILVW)xxxxx(FILVW) | none? |
| 1-16 Subclasses | ||
| 1-16 | (FILVW)xxxxxxxxxxxxx(FILVW) | Atg1; Atg13; Atg101; Cdk5; CmbB; fAR1; L9; NdkC; NumA; PsaA; Rab11a; RgaA;VwkA |
| Calmodulin-Binding Motifs in Dictyostelium | |||
|---|---|---|---|
| Full IQ motifs ((F/I/L/V)Qxxx(R/K)Gxxx(R/K)xx(F/I/L/V/W/Y)) | |||
| CaMBP (gene) | Dictybase | IQ | Binding Sequence |
| MhcA (mhcA) | DDB0191444 | IQ1 | IQAA TRGWIARKVYK |
| Myo1E(myoE) | DDB0216200 | IQ1 | IQKTWRGYRARSKWN |
| Myo1F(myoF) | DDB0220021 | IQ1 | IQRVWRGYKVRKWYD |
| Myo5A(myoH) | DDB0233685 | IQ1 | IQKIWRGYTDRKAYI |
| Myo5B(myoJ) | DDB0185050 | IQ1 | IQKRWKGYLYRKRYK |
| IQ-like motifs ((F/I/L/V)Qxxx(R/K)xxxxxxxx) | |||
| DwwA (dwwA) | DDB0216188 | IQ1 | IQRT FRNHKKQSY |
| MhcA (mhcA) | DDB0191444 | IQ2 | IQQNLRAYIDFKSW |
| Myo1A(myoA) | DDB0215392 | IQ2 | IQRT YRGWLLVRECV |
| Myo1B(myoB) | DDB0191351 | IQ1 | IQKAFRNWKAKKHSL |
| Myo1C(myoC) | DDB0215355 | IQ3 | IQGYFRAWKQASPFF |
| Myo1E(myoE) | DDB0216200 | IQ2 | IQLF YRSYRYKKWFR |
| Myo5A(myoH) | DDB0233685 | IQ2 | FQSL IRSYLQQLEYN |
| Myo5A(myoH) | DDB0233685 | IQ3 | LQSL IRTNELEKQFN |
| Myo5A(myoH) | DDB0233685 | IQ4 | FQSL LRRLEDSKEFN |
| Myo5A(myoH) | DDB0233685 | IQ5 | IQSLWRSNLAKKQLK |
| Myo5B(myoJ) | DDB0185050 | IQ2 | IQTK LRSVHAKQQLS |
| Myo5B(myoJ) | DDB0185050 | IQ3 | IQKVWRAHRDRVQYQ |
| Myo5B(myoJ) | DDB0185050 | IQ4 | IQTVMRRHLFSEQVH |
| Myo5B(myoJ) | DDB0185050 | IQ5 | LQTK IRQILSKREVD |
| Myo5B(myoJ) | DDB0185050 | IQ6 | IQARWRMKLAKRVYI |
| MyoG (myoG) | DDB0232322 | IQ1 | IQSTWRMYLIRKRFI |
| MyoG (myoG) | DDB0232322 | IQ2 | IQKNTRRWLVQKRYQ |
| MyoG (myoG) | DDB0232322 | IQ4 | IQTHLRSLLSKDYSY |
| MyoI (myoI) | DDB0185049 | IQ1 | IQSVWRMYRCKKRYQ |
| MyoI (myoI) | DDB0185049 | IQ4 | VQNNIRSFIARRHSR |
| MyoM (myoM) | DDB0191100 | IQ1 | IQAF FKMIKIRNQYK |
| MyoM (myoM) | DDB0191100 | IQ2 | LQTL IRAQRAKKDFE |
| NumA (numA) | DDB0231257 | IQ1 | LQKQQKIYKDLERF |
| RgaA (rgaA) | DDB0191437 | IQ1 | IQEL KRNLVSEVRR |
| RgaA (rgaA) | DDB0191437 | IQ2 | LQTE PKYLAGLVYL |
| VatM | IQLA LRTATTRSGA | ||
| Non-IQ motifs | |||
| Myo1A(myoA) | DDB0215392 | IQ1 | IGSVWKMYKQRIKWYL |
| Myo1C(myoC) | DDB0215355 | IQ1 | IKNAYRNYKAFQFEC |
| Myo1C(myoC) | DDB0215355 | IQ2 | IKNAFRNYKLYRQRC |
| Myo1D(myoD) | DDB0191347 | IQ1 | LQRFFLRFTLMSYYY |
| Myo1F(myoF) | DDB0220021 | IQ2 | IQTYYLRYKVLTYIK |
| MyoG (myoG) | DDB0232322 | IQ3 | LESFSRMVIFRAPYL |
| MyoI (myoI) | DDB0185049 | IQ2 | LGAAMLSHSSRRDFQ |
| MyoI (myoI) | DDB0185049 | IQ3 | IKGFFKMLTYQKQFK |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Day, D.H.; Taylor, R.J.; Myre, M.A. Calmodulin and Calmodulin Binding Proteins in Dictyostelium: A Primer. Int. J. Mol. Sci. 2020, 21, 1210. https://doi.org/10.3390/ijms21041210
O’Day DH, Taylor RJ, Myre MA. Calmodulin and Calmodulin Binding Proteins in Dictyostelium: A Primer. International Journal of Molecular Sciences. 2020; 21(4):1210. https://doi.org/10.3390/ijms21041210
Chicago/Turabian StyleO’Day, Danton H., Ryan J. Taylor, and Michael A. Myre. 2020. "Calmodulin and Calmodulin Binding Proteins in Dictyostelium: A Primer" International Journal of Molecular Sciences 21, no. 4: 1210. https://doi.org/10.3390/ijms21041210
APA StyleO’Day, D. H., Taylor, R. J., & Myre, M. A. (2020). Calmodulin and Calmodulin Binding Proteins in Dictyostelium: A Primer. International Journal of Molecular Sciences, 21(4), 1210. https://doi.org/10.3390/ijms21041210