Carvone Enantiomers Differentially Modulate IgE-Mediated Airway Inflammation in Mice

 ,
, 

 and
and
Abstract
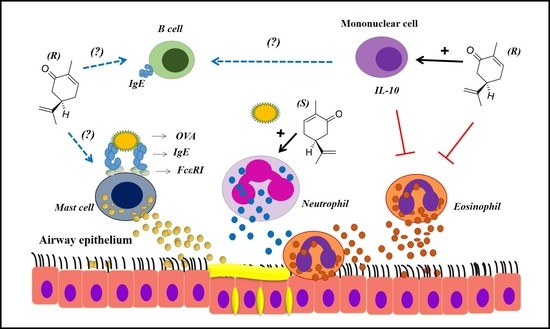
1. Introduction
2. Results
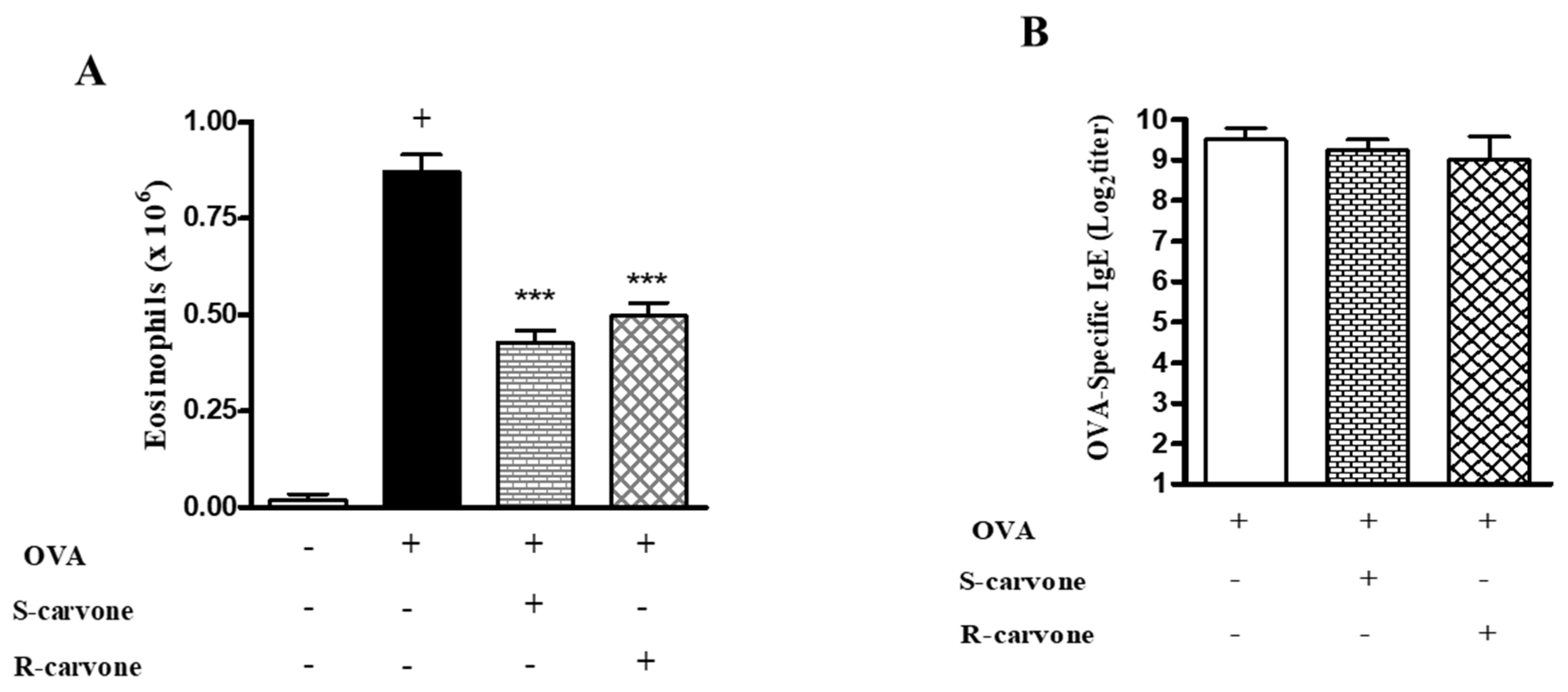
2.1. Carvone Enantiomers Inhibit Eosinophil Recruitment in a Mice Model of Allergic Airway Inflammation
2.2. Effects of Carvone Enantiomers on Leukocyte Infiltration in the Lung of Allergic Mice
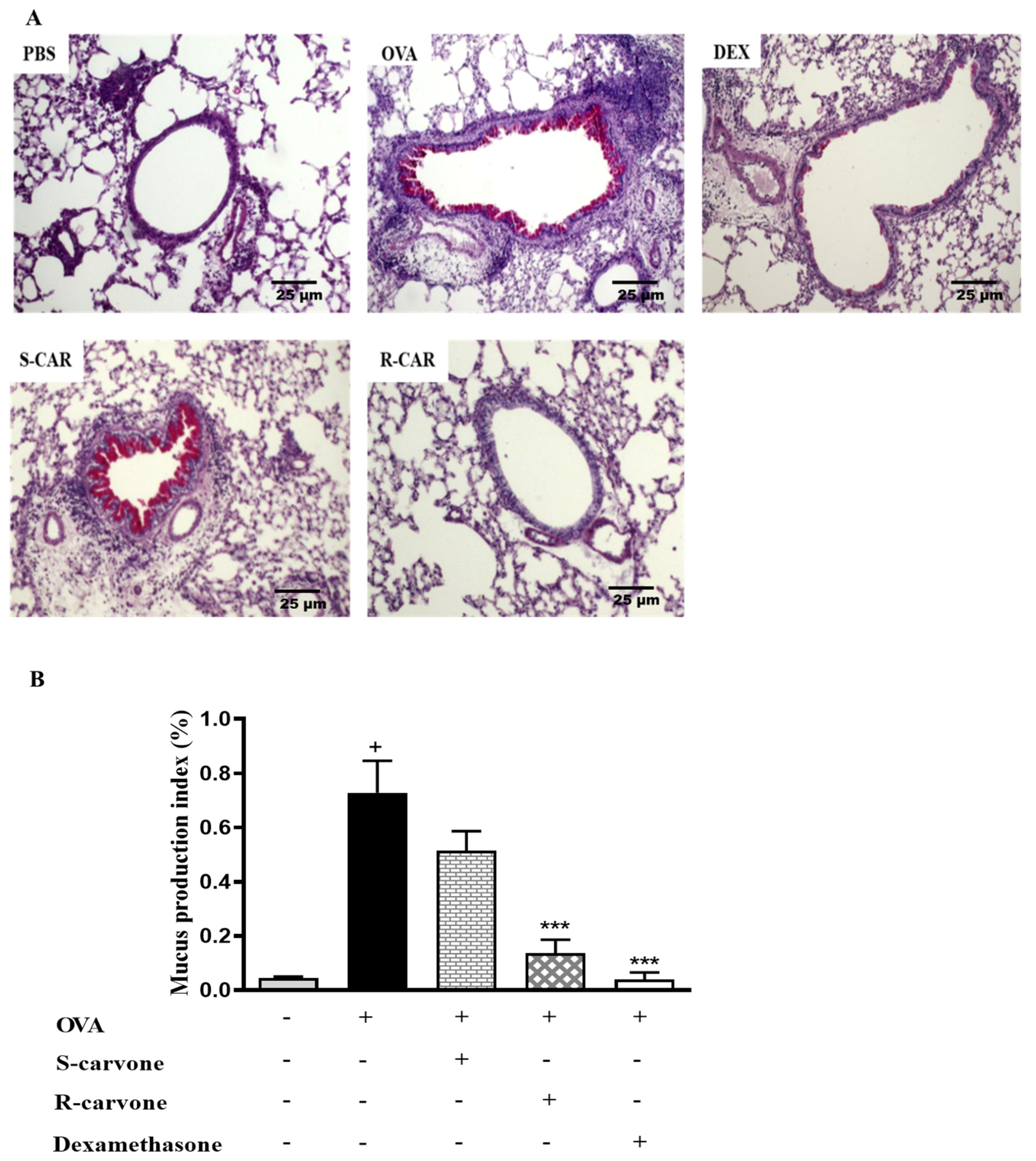
2.3. Effects of Carvone Enantiomers on Mucus Production in the Lung of Allergic Mice
2.4. Carvone Enantiomers Modulate IgE and Cytokine Production In Vivo
2.5. Effects of the Treatment with Carvone Enantiomers on Allergic Sensitization
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Animals
4.3. Airway Allergic Inflammation in Actively Sensitized Mice
4.4. Leukocyte Counting
4.5. Lung Histology
4.6. Quantification of OVA-Specific IgE by Passive Cutaneous Anaphylaxis (PCA)
4.7. Cytokine Quantification
4.8. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BAL | Bronchoalveolar Lavage |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| HE | Hematoxylin-eosin |
| IgE | Immunoglobulin E |
| IFN | Interferon |
| MMP | Matrix metalloproteinase |
| OVA | Ovalbumin |
| PAS | Periodic acid–Schiff |
| PBS | Phosphate-Buffered Saline |
| PCA | Passive cutaneous anaphylaxis |
| R-carvone | (R)-(−)-carvone |
| S-carvone | (S)-(+)-carvone |
| Th2 | T helper 2 |
| TMB | 3,3′,5,5′-Tetramethylbenzidine |
References
- Barnes, P.J. Targeting cytokines to treat asthma and chronic obstructive pulmonary disease. Nat. Rev. Immunol. 2018, 18, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Global Asthma Network. The Global Asthma Report 2018; Global Asthma Network: Auckland, New Zealand, 2018. [Google Scholar]
- Barnes, P.J. Pathophysiology of allergic inflammation. Immunol. Rev. 2011, 242, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Cellular and molecular mechanisms of asthma and COPD. Clin. Sci. 2017, 131, 1541–1558. [Google Scholar] [CrossRef] [PubMed]
- Ebbo, M.; Crinier, A.; Vely, F.; Vivier, E. Innate lymphoid cells: Major players in inflammatory diseases. Nat. Rev. Immunol. 2017, 17, 665. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Glucocorticosteroids: Current and future directions. Br. J. Pharm. 2011, 163, 29–43. [Google Scholar] [CrossRef]
- Ribeiro-Filho, J.; Piuvezam, M.R.; Bozza, P.T. Anti-allergic properties of curine, a bisbenzylisoquinoline alkaloid. Molecules 2015, 20, 4695–4707. [Google Scholar] [CrossRef]
- Srivastava, A.; Lall, R.; Sinha, A.; Gupta, R.C. Essential Oils. In Nutraceuticals in Veterinary Medicine; Springer: Cham, Switzerland, 2019; pp. 157–173. [Google Scholar]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef]
- Younis, Y.M.; Beshir, S.M. Carvone-rich essential oils from Mentha longifolia (L.) Huds. ssp. schimperi Briq. and Mentha spicata L. grown in Sudan. J. Essent. Oil Res. 2004, 16, 539–541. [Google Scholar] [CrossRef]
- Buckingham, J. (Ed.) Dictionary of Natural Products; CRC Press: Boca Raton, FL, USA, 1997; Volume 4, p. 1984. [Google Scholar]
- Silva, M.C.E.; Galhano, C.I.C.; da Silva, A.M. A new sprout inhibitor of potato tuber based on carvone/β-cyclodextrin inclusion compound. J. Incl. Phenom. Macrocycl. Chem. 2007, 57, 121–124. [Google Scholar] [CrossRef][Green Version]
- McGeady, P.; Wansley, D.L.; Logan, D.A. Carvone and Perillaldehyde Interfere with the Serum-Induced Formation of Filamentous Structures in Candida albicans at Substantially Lower Concentrations than Those Causing Significant Inhibition of Growth. J. Nat. Prod. 2002, 65, 953–955. [Google Scholar] [CrossRef]
- Gould, M.N.; Howard, S.P.; Rajesh, D. Monoterpenes and Sesquiterpenes as Chemotherapeutic and Radiation Sensitizers and Immunomodulators. U.S. Patent No. 7,056,491, 6 June 2006. [Google Scholar]
- Goncalves, J.C.R.; de Sousa Oliveira, F.; Benedito, R.B.; de Sousa, D.P.; de Almeida, R.N.; de Araújo, D.A.M. Antinociceptive activity of (−)-carvone: Evidence of association with decreased peripheral nerve excitability. Biol. Pharm. Bull. 2008, 31, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Maruyama, N.; Hayama, K.; Ishibashi, H.; Inoue, S.; Oshima, H.; Yamaguchi, H. Suppression of tumor necrosis factor-alpha-induced neutrophil adherence responses by essential oils. Mediat. Inflamm. 2003, 12, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Juergens, U.R.; Dethlefsen, U.; Steinkamp, G.; Gillissen, A.; Repges, R.; Vetter, H. Anti-inflammatory activity of 1.8-cineol (eucalyptol) in bronchial asthma: A double-blind placebo-controlled trial. Respir. Med. 2003, 97, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Juergens, U.R.; Engelen, T.; Racké, K.; Stöber, M.; Gillissen, A.; Vetter, H. Inhibitory activity of 1, 8-cineol (eucalyptol) on cytokine production in cultured human lymphocytes and monocytes. Pulm. Pharmacol. Ther. 2004, 17, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Filho, J.; Calheiros, A.S.; Vieira-de-Abreu, A.; Carvalho, K.I.M.; Mendes, D.S.; Bandeira-Melo, C.; Martins, M.A.; Dias, C.S.; Piuvezam, M.R.; Bozza, P.T. Curine inibits eosinophil activation and airway hyper-responsiveness in a mouse model of allergic asthma. Toxicol. Appl. Pharm. 2013, 273, 19–26. [Google Scholar] [CrossRef]
- Nunez, M.C.; Garcia-Rubino, M.E.; Conejo-Garcia, A.; Cruz-Lopez, O.; Kimatrai, M.; Gallo, M.A.; Campos, J.M. Homochiral drugs: A demanding tendency of the pharmaceutical industry. Curr. Med. Chem. 2009, 16, 2064–2074. [Google Scholar] [CrossRef]
- Kasprzyk-Hordern, B. Pharmacologically active compounds in the environment and their chirality. Chem. Soc. Rev. 2010, 39, 4466–4503. [Google Scholar] [CrossRef]
- Casy, A.F. The Steric Factor in Medicinal Chemistry: Dissymmetric Probes of Pharmacological Receptors; Springer: New York, NY, USA, 2013. [Google Scholar]
- Bertucci, C.; Pistolozzi, M.; de Simone, A. Circular dichroism in drug discovery and development: An abridged review. Anal. Bioanal. Chem. 2010, 398, 155–166. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, L.; Zhou, H.; Yu, L.S.; Zeng, S. Stereoselective binding of chiral drugs to plasma proteins. Acta Pharm. Sin. 2013, 34, 998–1006. [Google Scholar] [CrossRef]
- McBrien, C.N.; Menzies-Gow, A. The biology of eosinophils and their role in asthma. Front. Med. 2017, 4, 93. [Google Scholar] [CrossRef]
- Serhan, C.; Chiang, N.; Van Dyke, T. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Lampinen, M.; Carlson, M.; Håkansson, L.D.; Venge, P. Cytokine-regulated accumulation of eosinophils in inflammatory disease. Allergy 2004, 59, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.F.; Berdnikovs, S.; Simon, H.U.; Bochner, B.S.; Rosenwasser, L.J. Eosinophilic bioactivities in severe asthma. World Allergy Organ. J. 2016, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Corry, D.B.; Rishi, K.; Kanellis, J.; Kiss, A.; Song, L.Z.; Xu, J.; Kheradmand, F. Decreased allergic lung inflammatory cell egression and increased susceptibility to asphyxiation in MMP2-deficiency. Nat. Immunol. 2002, 3, 347–353. [Google Scholar] [CrossRef]
- Dai, J.P.; Chen, J.; Bei, Y.F.; Han, B.X.; Wang, S. Influence of borneol on primary mice oral fibroblasts: A penetration enhancer may be used in oral submucous fibrosis. J. Oral Pathol. Med. 2009, 38, 276–281. [Google Scholar] [CrossRef]
- Fahy, J.V. Goblet cell and mucin gene abnormalities in asthma. Chest 2002, 122, 320S–326S. [Google Scholar] [CrossRef]
- Shimizu, T.; Shimizu, S.; Hattori, R.; Majima, Y. A mechanism of antigen-induced goblet cell degranulation in the nasal epithelium of sensitized rats. J. Allergy Clin. Immunol. 2003, 112, 119–125. [Google Scholar] [CrossRef]
- Bloemen, K.; Verstraelen, S.; Van Den Heuvel, R.; Witters, H.; Nelissen, I.; Schoeters, G. The allergic cascade: Review of the most important molecules in the asthmatic lung. Immunol. Lett. 2007, 113, 6–18. [Google Scholar] [CrossRef]
- Fu, C.L.; Ye, Y.L.; Lee, Y.L.; Chiang, B.L. Effects of overexpression of IL-10, IL-12, TGF-β and IL-4 on allergen induced change in bronchial responsiveness. Respir. Res. 2006, 7, 72. [Google Scholar] [CrossRef]
- Coomes, S.; Kannan, Y.; Pelly, V.; Entwistle, L.J.; Guidi, R.; Perez-Lloret, J.; Nikolov, N.; Müller, W.; Wilson, M.S. CD4+ Th2 cells are directly regulated by IL-10 during allergic airway inflammation. Mucosal Immunol. 2017, 10, 150–161. [Google Scholar] [CrossRef]
- Patel, T.R.; Sur, S. IgE and eosinophils as therapeutic targets in asthma. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Haworth, O.; Cernadas, M.; Yang, R.; Serhan, C.N.; Levy, B.D. Resolvin E1 regulates interleukin 23, interferon-gamma and lipoxin A4 to promote the resolution of allergic airway inflammation. Nat. Immunol. 2008, 9, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Hachem, P.; Lisbonne, M.; Michel, M.L.; Diem, S.; Roongapinun, S.; Lefort, J.; Leite-de-Moraes, M.C. α-galactosylceramide-induced iNKT cells suppress experimental allergic asthma in sensitized mice: Role of IFN-γ. Eur. J. Immunol. 2005, 35, 2793–2802. [Google Scholar] [CrossRef]
- Reisinger, J.; Triendl, A.; Küchler, E.; Bohle, B.; Krauth, M.T.; Rauter, I.; Niederberger, V. IFN-γ–enhanced allergen penetration across respiratory epithelium augments allergic inflammation. J. Allergy Clin. Immunol. 2005, 115, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T.; Polosa, R. Treatment strategies for allergy and asthma. Nat. Rev. Immunol. 2008, 8, 218–230. [Google Scholar] [CrossRef]
- Ethuin, F.; Gérard, B.; Benna, J.E.; Boutten, A.; Gougereot-Pocidalo, M.-A.; Jacob, L.; Chollet-Martin, S. Human neutrophils produce interferon gamma upon stimulation by interleukin-12. Lab. Investig. 2004, 84, 1363–1371. [Google Scholar] [CrossRef]
- Sturge, C.R.; Burger, E.; Raetz, M.; Hooper, L.V.; Yarovinsky, F. Cutting Edge: Developmental Regulation of IFN-γ Production by Mouse Neutrophil Precursor Cells. J. Immunol. 2015, 195, 36–40. [Google Scholar] [CrossRef]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion on the safety assessment of carvone, considering all sources of exposure. EFSA scientific committee. EFSA J. 2014, 12, 3806. [Google Scholar] [CrossRef]
- Bastos, V.P.; Gomes, A.S.; Lima, F.J.; Brito, T.S.; Soares, P.M.; Pinho, J.P.; Magalhães, P.J. Inhaled 1, 8-cineole reduces inflammatory parameters in airways of Ovalbumin-challenged Guinea Pigs. Basic Clin. Pharm. Toxicol. 2011, 108, 34–39. [Google Scholar] [CrossRef]
- Juergens, U.R.; Stöber, M.; Vetter, H. The anti-inflammatory activity of L-menthol compared to mint oil in human monocytes in vitro: A novel perspective for its therapeutic use in inflammatory diseases. Eur. J. Med. Res. 1998, 3, 539. [Google Scholar] [PubMed]
- Khan, S.; Weterings, J.J.; Britten, C.M.; de Jong, A.R.; Graafland, D.; Melief, C.J.; Ossendorp, F. Chirality of TLR-2 ligand Pam3CysSK4 in fully synthetic peptide conjugates critically influences the induction of specific CD8+ T-cells. Mol. Immunol. 2009, 46, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Yu, E.S.; Han, A.R.; Nam, J.W.; Seo, E.K.; Hwang, E.S. Isoeleutherin and eleutherinol, naturally occurring selective modulators of Th cell-mediated immune responses. Biochem. Biophys. Res. Commun. 2008, 371, 278–282. [Google Scholar] [CrossRef]
- Henderson, W.R., Jr.; Banerjee, E.R.; Chi, E.Y. Differential effects of (S)-and (R)-enantiomers of albuterol in a mouse asthma model. J. Allergy Clin. Immunol. 2005, 116, 332–340. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Zhou, Y.; Wang, Y.; Sun, X.; Zhao, Y.; Guan, Y.; Zhang, Y.; Wang, W. PPARγ as an E3 Ubiquitin-Ligase Impedes Phosphate-Stat6 Stability and Promotes Prostaglandins E2-Mediated Inhibition of IgE Production in Asthma. Front. Immunol. 2020, 11, 1224. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, C.M.; Christiansen, S.B.; Duncan, I.J.; Erhard, H.W.; Lay, D.C., Jr.; Mench, J.A.; O’Connor, C.E.; Petherick, J.C. Guidelines for the ethical use of animals in applied ethology studies. Appl. Anim. Behav. Sci. 2003, 81, 291–305. [Google Scholar] [CrossRef]
- Lloyd, C.M.; Gonzalo, J.A.; Nguyen, T.; Delaney, T.; Tian, J.; Oettgen, H.; Gutierrez-Ramos, J.C. Resolution of bronchial hyperresponsiveness and pulmonary inflammation is associated with IL-3 and tissue leukocyte apoptosis. J. Immunol. 2001, 166, 2033–2040. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, N.; Fu, X.; Wei, L.; Ding, M.; Pang, Y.; Liu, D.; Ren, Y.; Guo, M. Butylphthalide ameliorates airway inflammation and mucus hypersecretion via NF-κB in a murine asthma model. Int. Immunopharmacol. 2019, 76, 105873. [Google Scholar] [CrossRef]
- Holt, P.G.; Rose, A.H.; Batty, J.E.; Turner, K.J. Induction of adjuvant independent IgE responses in inbred mice: Primary, secondary, and persistent IgE responses to ovalbumin and ovomucoid. Int. Arch. Allergy Immunol. 1981, 65, 42–50. [Google Scholar] [CrossRef]
- Costa, H.F.; Bezerra-Santos, C.R.; Barbosa Filho, J.M.; Martins, M.A.; Piuvezam, M.R. Warifteine, a bisbenzylisoquinoline alkaloid, decreases immediate allergic and thermal hyperalgesic reactions in sensitized animals. Int. Immunopharmacol. 2008, 8, 519–525. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Not Detectable (0) | Mild (1) | Moderate (2) | Severe (3) |
|---|---|---|---|---|
| PBS | 4 | 1 | - | - |
| OVA | - | - | 2 | 3 |
| S-carvone | - | 1 | 4 | - |
| R-carvone | 1 | 2 | 2 | - |
| Dexamethasone | 1 | 4 | - | - |
| Group | Not Detectable (0) | Mild (1) | Moderate (2) | Severe (3) |
|---|---|---|---|---|
| PBS | - | - | - | - |
| OVA | - | 2 | 3 | - |
| S-carvone | - | 1 | 3 | 1 |
| R-carvone | 1 | 2 | 2 | - |
| Dexamethasone | 1 | 4 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro-Filho, J.; da Silva Brandi, J.; Ferreira Costa, H.; Carla de Paula Medeiros, K.; Alves Leite, J.; Pergentino de Sousa, D.; Regina Piuvezam, M. Carvone Enantiomers Differentially Modulate IgE-Mediated Airway Inflammation in Mice. Int. J. Mol. Sci. 2020, 21, 9209. https://doi.org/10.3390/ijms21239209
Ribeiro-Filho J, da Silva Brandi J, Ferreira Costa H, Carla de Paula Medeiros K, Alves Leite J, Pergentino de Sousa D, Regina Piuvezam M. Carvone Enantiomers Differentially Modulate IgE-Mediated Airway Inflammation in Mice. International Journal of Molecular Sciences. 2020; 21(23):9209. https://doi.org/10.3390/ijms21239209
Chicago/Turabian StyleRibeiro-Filho, Jaime, Juliana da Silva Brandi, Hermann Ferreira Costa, Karina Carla de Paula Medeiros, Jacqueline Alves Leite, Damião Pergentino de Sousa, and Márcia Regina Piuvezam. 2020. "Carvone Enantiomers Differentially Modulate IgE-Mediated Airway Inflammation in Mice" International Journal of Molecular Sciences 21, no. 23: 9209. https://doi.org/10.3390/ijms21239209
APA StyleRibeiro-Filho, J., da Silva Brandi, J., Ferreira Costa, H., Carla de Paula Medeiros, K., Alves Leite, J., Pergentino de Sousa, D., & Regina Piuvezam, M. (2020). Carvone Enantiomers Differentially Modulate IgE-Mediated Airway Inflammation in Mice. International Journal of Molecular Sciences, 21(23), 9209. https://doi.org/10.3390/ijms21239209