The Impact of Human Lipoaspirate and Adipose Tissue-Derived Stem Cells Contact Culture on Breast Cancer Cells: Implications in Breast Reconstruction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
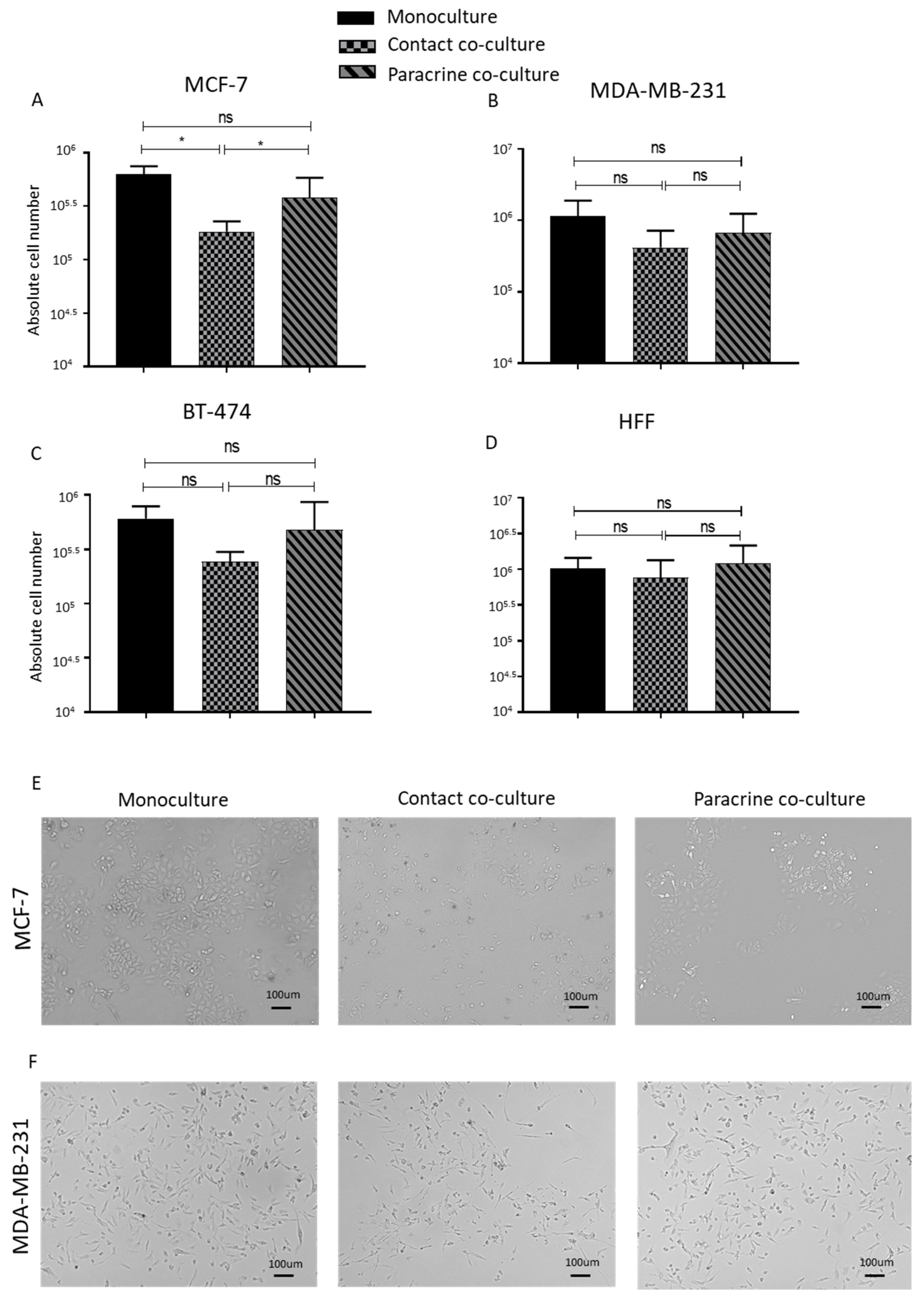
2.1. Lipoaspirate Contact Co-Culture Reduces the Proliferation Rate of Breast Cancer Cell Lines
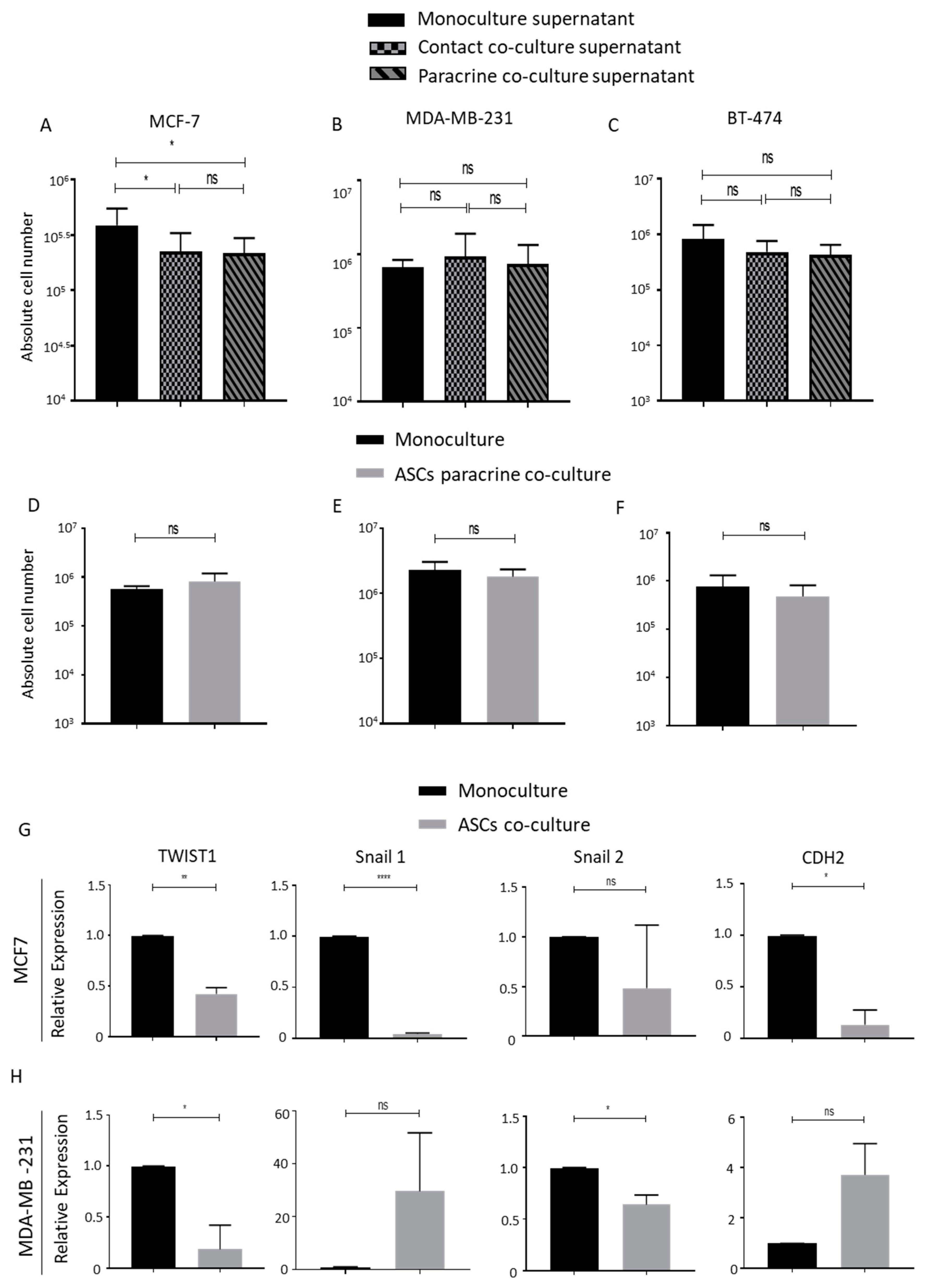
2.2. Conditioned Mediums from Lipoaspirate Co-Culture Do Not Promote the Proliferation of Breast Cancer Cells in Culture
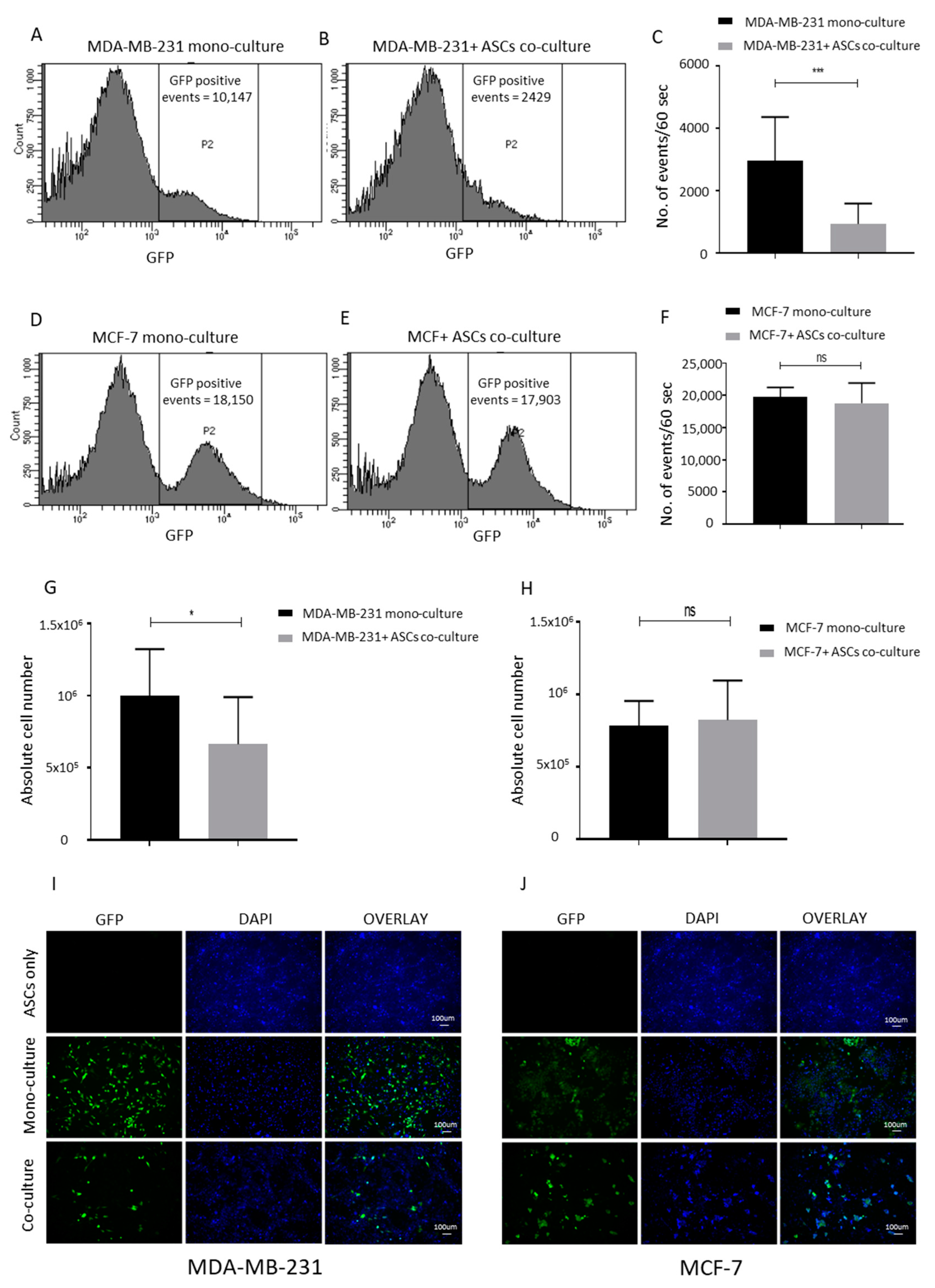
2.3. ASCs Contact Culture Inhibit the Proliferation of MDA-MB-231 Cells
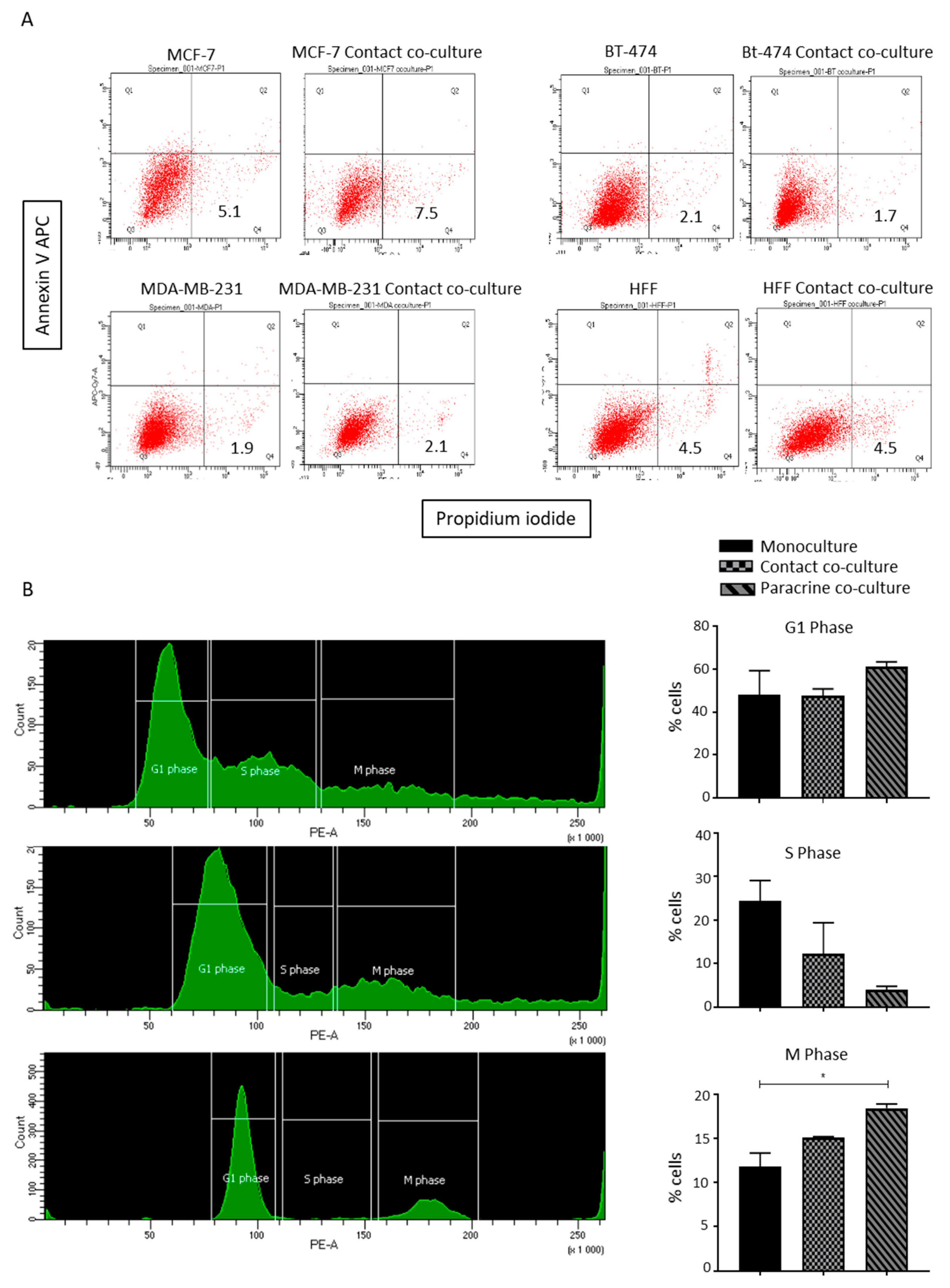
2.4. Lipoaspirate Contact Co-Culture Inhibit Cell Cycle in MCF-7 Cells
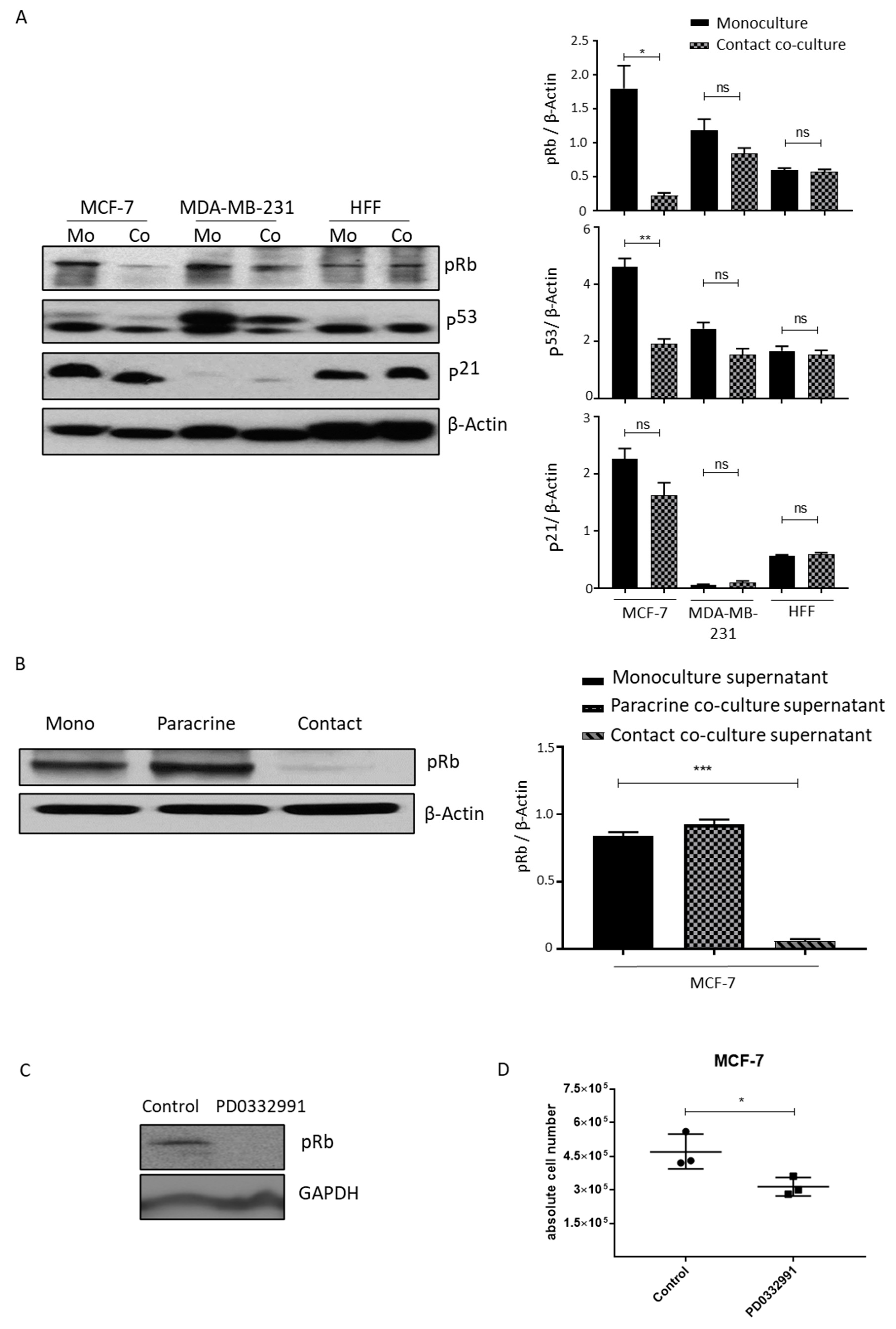
2.5. Lipoaspirate Contact Co-Culture and Supernatant Activate Retinoblastoma Protein (RB) Mediated Cell Cycle Arrest in MCF-7 Cells
3. Discussion
4. Materials and Methods
4.1. Donor Specification
4.2. Isolation of Human Adipose-Derived Stem Cells
4.3. Culture of ASCs
4.4. Culture of Breast Cancer Cell lines
4.5. Culture of Human Foreskin Fibroblasts (HFF)
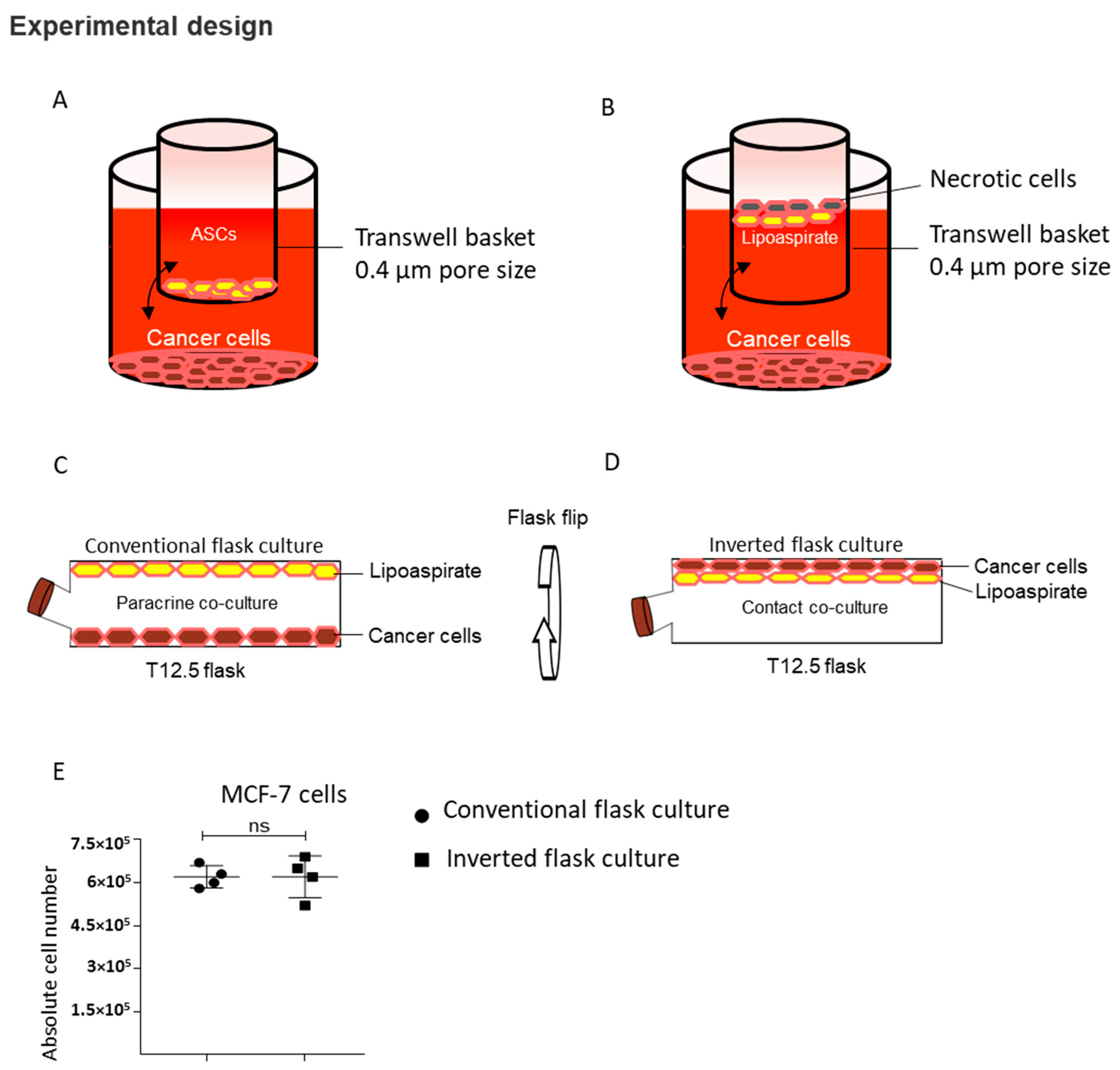
4.6. Inverted Flask Culture: Contact Co-Culture
4.7. Conventional Flask Culture: Paracrine Co-Culture
4.8. Transwell Co-Culture Experiments
4.9. Generation of GFP-Expressing Breast Cancer Cell Lines
4.10. ASCs Breast Cancer Cells Contact Co-Culture
4.11. GFP Positive Cell Count by FACS
4.12. CyQUANT Cell Proliferation Assay
4.13. Fluorescent Microscopy
4.14. Annexin/Propidium Iodide Live Dead staining
4.15. Cell Cycle Analyses
4.16. Western Blotting
4.17. Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.18. Statistical Analyses
4.19. Ethics Approval and Consent to Participate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krastev, T.; van Turnhout, A.; Vriens, E.; Smits, L.; van der Hulst, R. Long-term follow-up of autologous fat transfer vs conventional breast reconstruction and association with cancer relapse in patients with breast cancer. JAMA Surg. 2019, 154, 56–63. [Google Scholar] [CrossRef]
- Koellensperger, E.; Bonnert, L.C.; Zoernig, I.; Marmé, F.; Sandmann, S.; Germann, G.; Gramley, F.; Leimer, U. The impact of human adipose tissue-derived stem cells on breast cancer cells: Implications for cell-assisted lipotransfers in breast reconstruction. Stem Cell Res. Ther. 2017, 8, 121. [Google Scholar] [CrossRef]
- Coleman, S.R. Structural fat grafting. Aesthet. Surg. J. 1998, 18, 386–388. [Google Scholar] [CrossRef]
- Tsuji, W.; Valentin, J.E.; Marra, K.G.; Donnenberg, A.D.; Donnenberg, V.S.; Rubin, J.P. An animal model of local breast cancer recurrence in the setting of autologous fat grafting for breast reconstruction. Stem Cells Transl. Med. 2018, 7, 125–134. [Google Scholar] [CrossRef]
- Krastev, T.K.; Alshaikh, G.A.; Hommes, J.; Piatkowski, A.; van der Hulst, R.R. Efficacy of autologous fat transfer for the correction of contour deformities in the breast: A systematic review and meta-analysis. J. Plast. Reconstr. Aesthet. Surg. 2018, 71, 1392–1409. [Google Scholar] [CrossRef]
- Klinger, M.; Caviggioli, F.; Klinger, F.M.; Giannasi, S.; Bandi, V.; Banzatti, B.; Forcellini, D.; Maione, L.; Catania, B.; Vinci, V. Autologous fat graft in scar treatment. J. Craniofacial Surg. 2013, 24, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, M.E.; Brouwer, K.M.; Van Trier, A.J.; Groot, M.L.; Middelkoop, E.; Van Zuijlen, P.P. Effectiveness of autologous fat grafting in adherent scars: Results obtained by a comprehensive scar evaluation protocol. Plast. Reconstr. Surg. 2017, 139, 212–219. [Google Scholar] [CrossRef]
- Juhl, A.A.; Karlsson, P.; Damsgaard, T.E. Fat grafting for alleviating persistent pain after breast cancer treatment: A randomized controlled trial. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 1192–1202. [Google Scholar] [CrossRef]
- Maione, L.; Vinci, V.; Caviggioli, F.; Klinger, F.; Banzatti, B.; Catania, B.; Lisa, A.; Klinger, M. Autologous fat graft in postmastectomy pain syndrome following breast conservative surgery and radiotherapy. Aesthet. Plast. Surg. 2014, 38, 528–532. [Google Scholar] [CrossRef]
- Rigotti, G.; Marchi, A.; Galie, M.; Baroni, G.; Benati, D.; Krampera, M.; Pasini, A.; Sbarbati, A. Clinical treatment of radiotherapy tissue damage by lipoaspirate transplant: A healing process mediated by adipose-derived adult stem cells. Plast. Reconstr. Surg. 2007, 119, 1409–1422. [Google Scholar] [CrossRef]
- Panettiere, P.; Marchetti, L.; Accorsi, D. The serial free fat transfer in irradiated prosthetic breast reconstructions. Aesthet. Plast. Surg. 2009, 33, 695–700. [Google Scholar] [CrossRef]
- Brown, S.A.; Levi, B.; Lequex, C.; Wong, V.W.; Mojallal, A.; Longaker, M.T. Basic science review on adipose tissue for clinicians. Plast. Reconstr. Surg. 2010, 126, 1936–1946. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, G.; Marchi, A.; Galie, M.; Baroni, G.; Benati, D.; Krampera, M.; Pasini, A.; Sbarbati, A. Human adipose-derived mesenchymal stromal cells may promote breast cancer progression and metastatic spread. Plast. Reconstr. Surg. 2015, 136, 76–84. [Google Scholar]
- Picon-Ruiz, M.; Pan, C.; Drews-Elger, K.; Jang, K.; Besser, A.H.; Zhao, D.; Morata-Tarifa, C.; Kim, M.; Ince, T.A.; Azzam, D.J.; et al. Interactions between adipocytes and breast cancer cells stimulate cytokine production and drive Src/Sox2/miR-302b–mediated malignant progression. Cancer Res. 2016, 76, 491–504. [Google Scholar] [CrossRef]
- Charvet, H.J.; Orbay, H.; Harrison, L.; Devi, K.; Sahar, D.E. In vitro effects of adipose-derived stem cells on breast cancer cells harvested from the same patient. Ann. Plast. Surg. 2016, 76, S241–S245. [Google Scholar] [CrossRef]
- Goto, H.; Shimono, Y.; Funakoshi, Y.; Imamura, Y.; Toyoda, M.; Kiyota, N.; Kono, S.; Takao, S.; Mukohara, T.; Minami, H. Adipose-derived stem cells enhance human breast cancer growth and cancer stem cell-like properties through adipsin. Oncogene 2019, 38, 767–779. [Google Scholar] [CrossRef]
- Balaban, S.; Shearer, R.F.; Lee, L.S.; van Geldermalsen, M.; Schreuder, M.; Shtein, H.C.; Cairns, R.; Thomas, K.C.; Fazakerley, D.J.; Grewal, T.; et al. Adipocyte lipolysis links obesity to breast cancer growth: Adipocyte-derived fatty acids drive breast cancer cell proliferation and migration. Cancer Metab. 2017, 5, 1. [Google Scholar] [CrossRef]
- Jotzu, C.; Alt, E.; Welte, G.; Li, J.; Hennessy, B.T.; Devarajan, E.; Krishnappa, S.; Pinilla, S.; Droll, L.; Song, Y.H. Adipose tissue derived stem cells differentiate into carcinoma-associated fibroblast-like cells under the influence of tumor derived factors. Cell. Oncol. 2011, 34, 55–67. [Google Scholar] [CrossRef]
- Rowan, B.G.; Gimble, J.M.; Sheng, M.; Anbalagan, M.; Jones, R.K.; Frazier, T.P.; Asher, M.; Lacayo, E.A.; Friedlander, P.L.; Kutner, R.; et al. Human adipose tissue-derived stromal/stem cells promote migration and early metastasis of triple negative breast cancer xenografts. PLoS ONE 2014, 9, e89595. [Google Scholar] [CrossRef]
- Almarzouqi, F.; Rennekampff, H.-O.; Stromps, J.-P.; Alharbi, Z.; Pallua, N. The effect of lipoaspirates vs. dissected abdominal fat on breast cancer cells in vitro. Eur. J. Med. Res. 2017, 22, 10. [Google Scholar] [CrossRef][Green Version]
- Massa, M.; Gasparini, S.; Baldelli, I.; Scarabelli, L.; Santi, P.; Quarto, R.; Repaci, E. Interaction between breast cancer cells and adipose tissue cells derived from fat grafting. Aesthet. Surg. J. 2016, 36, 358–363. [Google Scholar] [CrossRef]
- Orbay, H.; Hinchcliff, K.M.; Charvet, H.J.; Sahar, D.E. Fat graft safety after oncologic surgery: Addressing the contradiction between in vitro and clinical studies. Plast. Reconstr. Surg. 2018, 142, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.M.; Kokai, L.E.; Donnenberg, V.S.; Fine, J.L.; Marra, K.G.; Donnenberg, A.D.; Neto, M.S.; Rubin, J.P. Oncologic Safety of Fat Grafting for Autologous Breast Reconstruction in an Animal Model of Residual Breast Cancer. Plast. Reconstr. Surg. 2019, 143, 103–112. [Google Scholar] [CrossRef]
- Silva-Vergara, C.; Fontdevila, J.; Descarrega, J.; Burdio, F.; Yoon, T.-S.; Grande, L. Oncological outcomes of lipofilling breast reconstruction: 195 consecutive cases and literature review. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 475–481. [Google Scholar] [CrossRef]
- Lee, J.S.; Eo, P.; Kim, M.C.; Kim, J.B.; Jin, H.K.; Bae, J.S.; Jeong, J.H.; Park, H.Y.; Yang, J.D. Effects of Stromal Vascular Fraction on Breast Cancer Growth and Fat Engraftment in NOD/SCID Mice. Aesthet. Plast. Surg. 2019, 43, 498–513. [Google Scholar] [CrossRef]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef]
- Shukla, L.; Morrison, W.A.; Shayan, R. Adipose-derived stem cells in radiotherapy injury: A new frontier. Front. Surg. 2015, 2, 1. [Google Scholar] [CrossRef]
- Gebremeskel, S.; Gencarelli, J.; Gareau, A.J.; Levatte, T.; Dugandzic, A.; Johnston, B.; Bezuhly, M. Promotion of primary murine breast cancer growth and metastasis by adipose-derived stem cells is reduced in the presence of autologous fat graft. Plast. Reconstr. Surg. 2019, 143, 137–147. [Google Scholar] [CrossRef]
- Vahmani, P.; Rolland, D.C.; Gzyl, K.E.; Baines, D.D.; Dugan, M.E. The Growth-Inhibiting Effects of Beef Fatty Acids on MCF-7 Cells Are Influenced Mostly by the Depot Location and Inconsistently by the Biohydrogenation Intermediate Content. Lipids 2018, 53, 699–708. [Google Scholar] [CrossRef]
- Fain, J.N.; Cheema, P.; Madan, A.K.; Tichansky, D.S. Dexamethasone and the inflammatory response in explants of human omental adipose tissue. Mol. Cell. Endocrinol. 2010, 315, 292–298. [Google Scholar] [CrossRef]
- Gesta, S.; Lolmède, K.; Daviaud, D.; Berlan, M.; Bouloumié, A.; Lafontan, M.; Valet, P.; Saulnier-Blache, J.S. Culture of human adipose tissue explants leads to profound alteration of adipocyte gene expression. Horm. Metab. Res. 2003, 35, 158–163. [Google Scholar] [CrossRef]
- Harms, M.J.; Li, Q.; Lee, S.; Zhang, C.; Kull, B.; Hallen, S.; Thorell, A.; Alexandersson, I.; Hagberg, C.E.; Peng, X.-R.; et al. Mature human white adipocytes cultured under membranes maintain identity, function, and can transdifferentiate into brown-like adipocytes. Cell Rep. 2019, 27, 213–225. [Google Scholar] [CrossRef]
- Mazur, S.; Zołocińska, A.; Siennicka, K.; Janik-Kosacka, K.; Chrapusta, A.; Pojda, Z. Safety of adipose-derived cell (stromal vascular fraction–SVF) augmentation for surgical breast reconstruction in cancer patients. Adv. Clin. Exp. Med. 2018, 27, 1085–1090. [Google Scholar] [CrossRef]
- Ritter, A.; Friemel, A.; Fornoff, F.; Adjan, M.; Solbach, C.; Yuan, J.; Louwen, F. Characterization of adipose-derived stem cells from subcutaneous and visceral adipose tissues and their function in breast cancer cells. Oncotarget 2015, 6, 34475. [Google Scholar] [CrossRef]
- Ryu, H.; Oh, J.E.; Rhee, K.J.; Baik, S.K.; Kim, J.; Kang, S.J.; Sohn, J.H.; Choi, E.; Shin, H.C.; Kim, Y.M.; et al. Adipose tissue-derived mesenchymal stem cells cultured at high density express IFN-β and suppress the growth of MCF-7 human breast cancer cells. Cancer Lett. 2014, 352, 220–227. [Google Scholar] [CrossRef]
- Yoon, N.; Park, M.; Shigemoto, T.; Peltier, G.; Lee, R. Activated human mesenchymal stem/stromal cells suppress metastatic features of MDA-MB-231 cells by secreting IFN-β. Cell Death Dis. 2016, 7, e2191. [Google Scholar] [CrossRef][Green Version]
- Sherr, C.J.; McCormick, F. The RB and p53 pathways in cancer. Cancer Cell 2002, 2, 103–112. [Google Scholar] [CrossRef]
- Cavinato, M.; Koziel, R.; Romani, N.; Weinmüllner, R.; Jenewein, B.; Hermann, M.; Dubrac, S.; Ratzinger, G.; Grillari, J.; Schmuth, M. UVB-Induced Senescence of Human Dermal Fibroblasts Involves Impairment of Proteasome and Enhanced Autophagic Activity. J. Gerontol. Ser. A 2017, 72, 632–639. [Google Scholar] [CrossRef][Green Version]
- Ejaz, A.; Mitterberger, M.C.; Lu, Z.; Mattesich, M.; Zwierzina, M.E.; Hörl, S.; Kaiser, A.; Viertler, H.-P.; Rostek, U.; Meryk, A. Weight loss upregulates the small GTPase DIRAS3 in human white adipose progenitor cells, which negatively regulates adipogenesis and activates autophagy via Akt–mTOR inhibition. EBioMedicine 2016, 6, 149–161. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ejaz, A.; Yang, K.S.; Venkatesh, K.P.; Chinnapaka, S.; Kokai, L.E.; Rubin, J.P. The Impact of Human Lipoaspirate and Adipose Tissue-Derived Stem Cells Contact Culture on Breast Cancer Cells: Implications in Breast Reconstruction. Int. J. Mol. Sci. 2020, 21, 9171. https://doi.org/10.3390/ijms21239171
Ejaz A, Yang KS, Venkatesh KP, Chinnapaka S, Kokai LE, Rubin JP. The Impact of Human Lipoaspirate and Adipose Tissue-Derived Stem Cells Contact Culture on Breast Cancer Cells: Implications in Breast Reconstruction. International Journal of Molecular Sciences. 2020; 21(23):9171. https://doi.org/10.3390/ijms21239171
Chicago/Turabian StyleEjaz, Asim, Katherine S. Yang, Kaushik P. Venkatesh, Somaiah Chinnapaka, Lauren E. Kokai, and J. Peter Rubin. 2020. "The Impact of Human Lipoaspirate and Adipose Tissue-Derived Stem Cells Contact Culture on Breast Cancer Cells: Implications in Breast Reconstruction" International Journal of Molecular Sciences 21, no. 23: 9171. https://doi.org/10.3390/ijms21239171
APA StyleEjaz, A., Yang, K. S., Venkatesh, K. P., Chinnapaka, S., Kokai, L. E., & Rubin, J. P. (2020). The Impact of Human Lipoaspirate and Adipose Tissue-Derived Stem Cells Contact Culture on Breast Cancer Cells: Implications in Breast Reconstruction. International Journal of Molecular Sciences, 21(23), 9171. https://doi.org/10.3390/ijms21239171