Maternal Malnutrition Affects Hepatic Metabolism through Decreased Hepatic Taurine Levels and Changes in HNF4A Methylation

 and
and
Abstract
1. Introduction
2. Results
2.1. Characteristics of 3-Month-Old Male Offspring
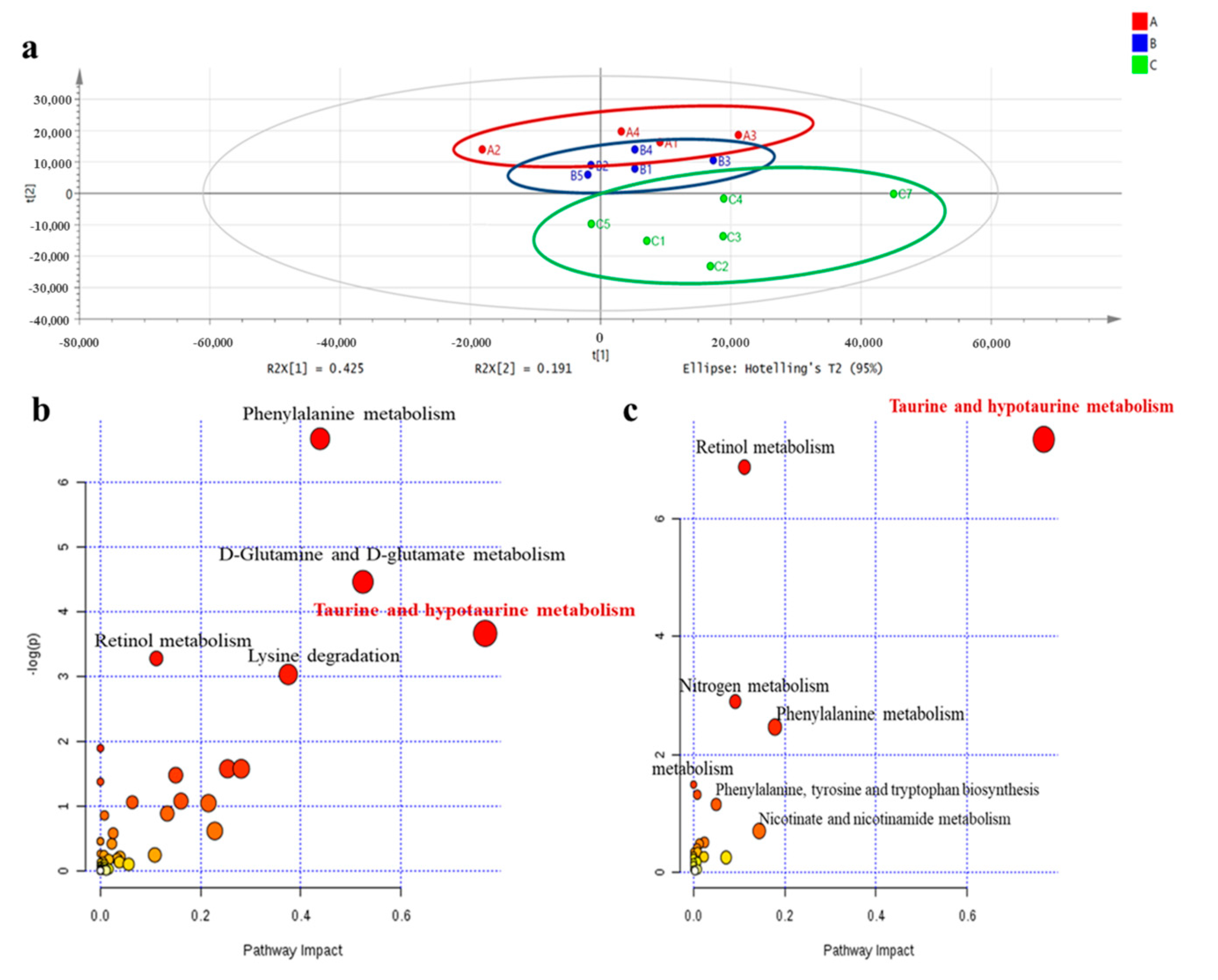
2.2. Metabolomic Analysis in Plasma in 3-Month-Old Male Offspring
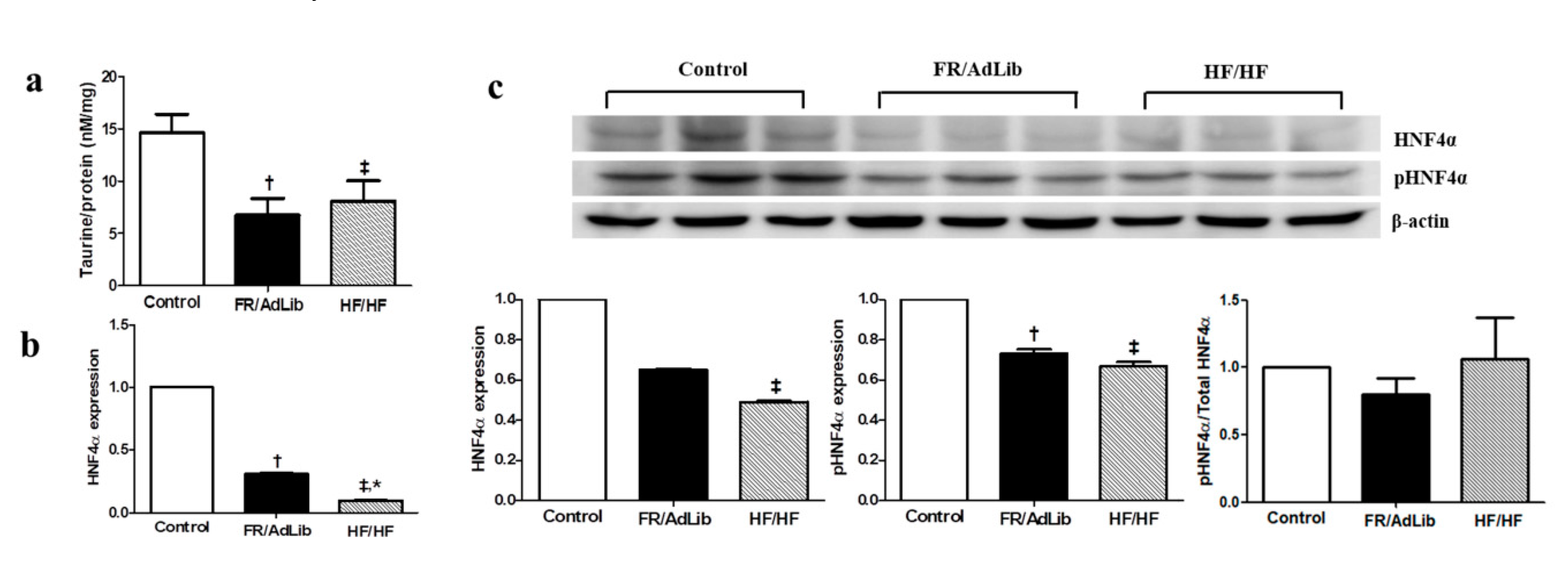
2.3. Hepatic HNF4A Methylation and Expression in 3-Month-Old Male Offspring
2.4. Hepatic Histology and Hepatic Triglyceride Analysis
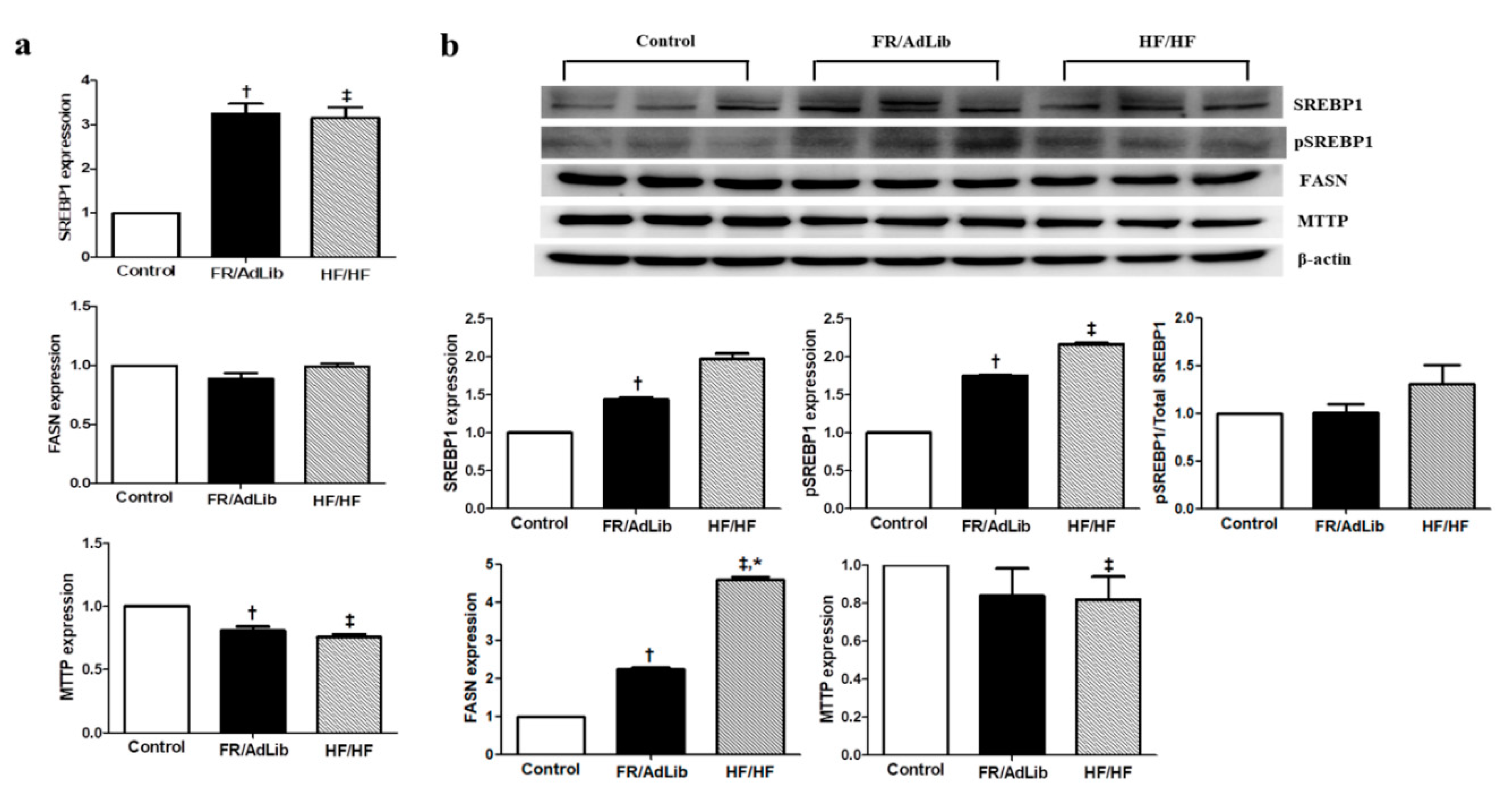
2.5. Maternal Diet Altered Hepatic Lipogenesis and Gluconeogenesis
3. Discussion
4. Materials and Methods
4.1. Animals and Study Design
4.2. Plasma and Hepatic Biochemical Analysis
4.3. Plasma Metabolomics Analysis
4.4. Hepatic Histological Analysis
4.5. DNA Methylation Analysis by Bisulfite Amplicon Sequencing
4.6. Gene Expression Analysis by Quantitative Real-Time PCR
4.7. Protein Expression Analysis by Western Blot
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Barker, D.J.; Hales, C.N.; Fall, C.H.; Osmond, C.; Phipps, K.; Clark, P.M. Type 2 (non-insulin-dependent) diabetes mellitus, hypertension and hyperlipidaemia (syndrome X): Relation to reduced fetal growth. Diabetologia 1993, 36, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Parlee, S.D.; MacDougald, O.A. Maternal nutrition and risk of obesity in offspring: The Trojan horse of developmental plasticity. Biochim. Biophys. Acta 2014, 1842, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Coupe, B.; Grit, I.; Darmaun, D.; Parnet, P. The timing of “catch-up growth” affects metabolism and appetite regulation in male rats born with intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R813–R824. [Google Scholar] [CrossRef] [PubMed]
- Lobstein, T.; Baur, L.; Uauy, R.; TaskForce, I.I.O. Obesity in children and young people: A crisis in public health. Obes. Rev. 2004, 5 (Suppl. 1), 4–104. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, P.D.; Buss, C.; Entringer, S.; Swanson, J.M. Developmental origins of health and disease: Brief history of the approach and current focus on epigenetic mechanisms. Semin. Reprod. Med. 2009, 27, 358–368. [Google Scholar] [CrossRef]
- Kwon, E.J.; Kim, Y.J. What is fetal programming? A lifetime health is under the control of in utero health. Obstet. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef]
- Choi, S.S.; Diehl, A.M. Hepatic triglyceride synthesis and nonalcoholic fatty liver disease. Curr. Opin. Lipidol. 2008, 19, 295–300. [Google Scholar] [CrossRef]
- Mensenkamp, A.R.; Havekes, L.M.; Romijn, J.A.; Kuipers, F. Hepatic steatosis and very low density lipoprotein secretion: The involvement of apolipoprotein E. J. Hepatol. 2001, 35, 816–822. [Google Scholar] [CrossRef]
- Cote, I.; Chapados, N.A.; Lavoie, J.M. Impaired VLDL assembly: A novel mechanism contributing to hepatic lipid accumulation following ovariectomy and high-fat/high-cholesterol diets? Br. J. Nutr. 2014, 112, 1592–1600. [Google Scholar] [CrossRef][Green Version]
- Li, M.; Reynolds, C.M.; Segovia, S.A.; Gray, C.; Vickers, M.H. Developmental Programming of Nonalcoholic Fatty Liver Disease: The Effect of Early Life Nutrition on Susceptibility and Disease Severity in Later Life. BioMed Res. Int. 2015, 2015, 437107. [Google Scholar] [CrossRef] [PubMed]
- Hoet, J.J.; Hanson, M.A. Intrauterine nutrition: Its importance during critical periods for cardiovascular and endocrine development. J. Physiol. 1999, 514, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Ozanne, S.E.; Hales, C.N. The long-term consequences of intra-uterine protein malnutrition for glucose metabolism. Proc. Nutr. Soc. 1999, 58, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Bautista, C.J.; Deas, M.; Martinez-Samayoa, P.M.; Gonzalez-Zamorano, M.; Ledesma, H.; Morales, J.; Larrea, F.; Nathanielsz, P.W. A low maternal protein diet during pregnancy and lactation has sex- and window of exposure-specific effects on offspring growth and food intake, glucose metabolism and serum leptin in the rat. J. Physiol. 2006, 571, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Guo, L.; Edmison, J.; Dasarathy, S.; McCullough, A.J.; Hanson, R.W.; Milburn, M. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metabolism 2011, 60, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Williams, S.; Grove, K.; Lane, R.H.; Aagaard-Tillery, K.M. A maternal high-fat diet is accompanied by alterations in the fetal primate metabolome. Am. J. Obstet. Gynecol. 2009, 201, 281.e1–281.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, X. Power of metabolomics in biomarker discovery and mining mechanisms of obesity. Obes. Rev. 2013, 14, 344–349. [Google Scholar] [CrossRef]
- Xie, B.; Waters, M.J.; Schirra, H.J. Investigating potential mechanisms of obesity by metabolomics. J. Biomed Biotechnol. 2012, 2012, 805683. [Google Scholar] [CrossRef]
- Van Vliet, E.; Eixarch, E.; Illa, M.; Arbat-Plana, A.; Gonzalez-Tendero, A.; Hogberg, H.T.; Zhao, L.; Hartung, T.; Gratacos, E. Metabolomics reveals metabolic alterations by intrauterine growth restriction in the fetal rabbit brain. PLoS ONE 2013, 8, e64545. [Google Scholar] [CrossRef]
- Chu, H.K.; Duan, Y.; Yang, L.; Schnabl, B. Small metabolites, possible big changes: A microbiota-centered view of non-alcoholic fatty liver disease. Gut 2019, 68, 359–370. [Google Scholar] [CrossRef]
- Desai, M.; Gayle, D.; Babu, J.; Ross, M.G. The timing of nutrient restriction during rat pregnancy/lactation alters metabolic syndrome phenotype. Am. J. Obstet. Gynecol. 2007, 196, 555.e1–555.e7. [Google Scholar] [CrossRef] [PubMed]
- Grattan, D.R. Fetal programming from maternal obesity: Eating too much for two? Endocrinology 2008, 149, 5345–5347. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, K.A.; Choi, G.Y.; Desai, M.; Lee, S.H.; Pang, M.G.; Jo, I.; Kim, Y.J. Feed restriction during pregnancy/lactation induces programmed changes in lipid, adiponectin and leptin levels with gender differences in rat offspring. J. Matern. Fetal Neonatal. Med. 2013, 26, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; You, Y.A.; Kwon, E.J.; Jung, S.C.; Jo, I.; Kim, Y.J. Maternal Food Restriction during Pregnancy and Lactation Adversely Affect Hepatic Growth and Lipid Metabolism in Three-Week-Old Rat Offspring. Int. J. Mol. Sci. 2016, 17, 2115. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoo, J.Y.; You, Y.A.; Kwon, W.S.; Lee, S.M.; Pang, M.G.; Kim, Y.J. Proteomic analysis of fetal programming-related obesity markers. Proteomics 2015, 15, 2669–2677. [Google Scholar] [CrossRef]
- Jones, R.H.; Ozanne, S.E. Fetal programming of glucose-insulin metabolism. Mol. Cell. Endocrinol. 2009, 297, 4–9. [Google Scholar] [CrossRef]
- Yan, C.C.; Bravo, E.; Cantafora, A. Effect of taurine levels on liver lipid metabolism: An in vivo study in the rat. Proc. Soc. Exp. Biol. Med. 1993, 202, 88–96. [Google Scholar] [CrossRef]
- Miyata, M.; Funaki, A.; Fukuhara, C.; Sumiya, Y.; Sugiura, Y. Taurine attenuates hepatic steatosis in a genetic model of fatty liver disease. J. Toxicol. Sci. 2020, 45, 87–94. [Google Scholar] [CrossRef]
- Gentile, C.L.; Nivala, A.M.; Gonzales, J.C.; Pfaffenbach, K.T.; Wang, D.; Wei, Y.; Jiang, H.; Orlicky, D.J.; Petersen, D.R.; Pagliassotti, M.J.; et al. Experimental evidence for therapeutic potential of taurine in the treatment of nonalcoholic fatty liver disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1710–R1722. [Google Scholar] [CrossRef]
- Chiang, J.Y. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar]
- Qi, Y.; Jiang, C.; Cheng, J.; Krausz, K.W.; Li, T.; Ferrell, J.M.; Gonzalez, F.J.; Chiang, J.Y. Bile acid signaling in lipid metabolism: Metabolomic and lipidomic analysis of lipid and bile acid markers linked to anti-obesity and anti-diabetes in mice. Biochim. Biophys. Acta 2015, 1851, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Hayhurst, G.P.; Lee, Y.H.; Lambert, G.; Ward, J.M.; Gonzalez, F.J. Hepatocyte nuclear factor 4alpha (nuclear receptor 2A1) is essential for maintenance of hepatic gene expression and lipid homeostasis. Mol. Cell. Biol. 2001, 21, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Si-Tayeb, K.; Lemaigre, F.P.; Duncan, S.A. Organogenesis and development of the liver. Dev. Cell 2010, 18, 175–189. [Google Scholar] [CrossRef]
- Sandovici, I.; Smith, N.H.; Nitert, M.D.; Ackers-Johnson, M.; Uribe-Lewis, S.; Ito, Y.; Jones, R.H.; Marquez, V.E.; Cairns, W.; Tadayyon, M.; et al. Maternal diet and aging alter the epigenetic control of a promoter-enhancer interaction at the Hnf4a gene in rat pancreatic islets. Proc. Natl. Acad. Sci. USA 2011, 108, 5449–5454. [Google Scholar] [CrossRef]
- Holliday, R. DNA methylation and epigenetic mechanisms. Cell Biophys. 1989, 15, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Cretney, E.C.; Kropp, J.; Khateeb, K.; Berg, M.A.; Penagaricano, F.; Magness, R.; Radunz, A.E.; Khatib, H. Maternal Diet during Pregnancy Induces Gene Expression and DNA Methylation Changes in Fetal Tissues in Sheep. Front. Genet. 2013, 4, 49. [Google Scholar] [CrossRef]
- Krahmer, N.; Farese, R.V., Jr.; Walther, T.C. Balancing the fat: Lipid droplets and human disease. EMBO Mol. Med. 2013, 5, 973–983. [Google Scholar] [CrossRef]
- Naik, A.; Kosir, R.; Rozman, D. Genomic aspects of NAFLD pathogenesis. Genomics 2013, 102, 84–95. [Google Scholar] [CrossRef]
- Shimano, H. Sterol regulatory element-binding proteins (SREBPs): Transcriptional regulators of lipid synthetic genes. Prog. Lipid Res. 2001, 40, 439–452. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Hussain, M.M.; Shi, J.; Dreizen, P. Microsomal triglyceride transfer protein and its role in apoB-lipoprotein assembly. J. Lipid Res. 2003, 44, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.M.; Rava, P.; Walsh, M.; Rana, M.; Iqbal, J. Multiple functions of microsomal triglyceride transfer protein. Nutr. Metab. 2012, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Dettlaff-Pokora, A.; Sledzinski, T.; Swierczynski, J. Up-Regulation Mttp and Apob Gene Expression in Rat Liver is Related to Post-Lipectomy Hypertriglyceridemia. Cell. Physiol. Biochem. 2015, 36, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Thorn, S.R.; Rozance, P.J.; Brown, L.D.; Hay, W.W., Jr. The intrauterine growth restriction phenotype: Fetal adaptations and potential implications for later life insulin resistance and diabetes. Semin. Reprod. Med. 2011, 29, 225–236. [Google Scholar] [CrossRef]
- Yamamoto, T.; Shimano, H.; Nakagawa, Y.; Ide, T.; Yahagi, N.; Matsuzaka, T.; Nakakuki, M.; Takahashi, A.; Suzuki, H.; Sone, H.; et al. SREBP-1 interacts with hepatocyte nuclear factor-4 alpha and interferes with PGC-1 recruitment to suppress hepatic gluconeogenic genes. J. Biol. Chem. 2004, 279, 12027–12035. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTORC1 activates SREBP-1c and uncouples lipogenesis from gluconeogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3281–3282. [Google Scholar] [CrossRef]
- Bonora, E.; Formentini, G.; Calcaterra, F.; Lombardi, S.; Marini, F.; Zenari, L.; Saggiani, F.; Poli, M.; Perbellini, S.; Raffaelli, A.; et al. HOMA-estimated insulin resistance is an independent predictor of cardiovascular disease in type 2 diabetic subjects—Prospective data from the Verona Diabetes Complicated Study. Diabetes Care 2002, 25, 1135–1141. [Google Scholar] [CrossRef]
- Tabachnik, T.; Kisliouk, T.; Marco, A.; Meiri, N.; Weller, A. Thyroid Hormone-Dependent Epigenetic Regulation of Melanocortin 4 Receptor Levels in Female Offspring of Obese Rats. Endocrinology 2017, 158, 842–851. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 9) | FR/AdLib (n = 9) | HF/HF (n = 9) | |
|---|---|---|---|
| Birth weight (g) | 7.09 ± 0.06 | 6.30 ± 0.08 † | 6.66 ± 0.07 ‡,* |
| Weight (g) | 466.78 ± 5.83 | 445.50 ± 10.96 | 602.11 ± 17.65 ‡,* |
| Weight gain (%) | 6474.33 ± 82.07 | 6971.42 ± 173.99 † | 8886.71 ± 263.41 ‡,* |
| Liver (g) | 14.18 ± 0.28 | 14.27 ± 0.74 | 17.17 ± 0.68 ‡,* |
| Liver/Weight (%) | 3.06 ± 0.06 | 3.18 ± 0.07 | 2.85 ± 0.05 ‡,* |
| Epididymal fat pad (g) | 4.90 ± 0.27 | 5.01 ± 0.56 | 15.58 ± 1.62 ‡,* |
| Control (n = 9) | FR/AdLib (n = 9) | HF/HF (n = 9) | |
|---|---|---|---|
| Plasma metabolic profiles | - | - | - |
| Glucose (mg/dl) | 142.22 ± 7.20 | 173.38 ± 11.61 † | 174.67 ± 7.17 ‡ |
| Insulin (ng/mL) | 0.15 ± 0.03 | 0.14 ± 0.01 | 0.38 ± 0.06 ‡,* |
| HOMA-IR | 1.31 ± 0.21 | 1.79 ± 0.18 † | 4.02 ± 0.58 ‡,* |
| Total cholesterol (mg/dl) | 59.38 ± 2.82 | 63.13 ± 3.68 | 82.33 ± 5.38 ‡,* |
| HDL -cholesterol (mg/dl) | 30.00 ± 5.28 | 43.75 ± 6.04 | 46.11 ± 10.08 |
| LDL -cholesterol (mg/dl) | 6.00 ± 0.66 | 6.38 ± 0.53 | 7.44 ± 1.12 |
| Triglyceride (mg/dl) | 58.22 ± 8.24 | 63.63 ± 13.86 | 62.44 ± 8.62 |
| HNF4A-P1_3M | |||||
|---|---|---|---|---|---|
| Control | FR/AdLib | HF/HF | Control vs. FR/AdLib (p-Value) | Control vs. HF/HF (p-Value) | |
| CpG1 | 0.2 ± 0.02 | 0.31 ± 0.04 | 0.31 ± 0.04 ‡ | 0.059 | 0.025 |
| CpG2 | 0.2 ± 0.05 | 0.31 ± 0.04 † | 0.32 ± 0.04 ‡ | 0.028 | 0.016 |
| CpG3 | 0.21 ± 0.01 | 0.32 ± 0.04 † | 0.33 ± 0.04 ‡ | 0.028 | 0.027 |
| CpG4 | 0.21 ± 0.02 | 0.32 ± 0.04 † | 0.33 ± 0.04 ‡ | 0.002 | 0.019 |
| CpG5 | 0.21 ± 0.02 | 0.33 ± 0.04 † | 0.32 ± 0.04 ‡ | 0.02 | 0.025 |
| CpG6 | 0.21 ± 0.02 | 0.31 ± 0.04 † | 0.31 ± 0.04 ‡ | 0.055 | 0.033 |
| CpG7 | 0.16 ± 0.02 | 0.25 ± 0.03 † | 0.28 ± 0.03 ‡ | 0.047 | 0.007 |
| CpG8 | 0.21 ± 0.02 | 0.33 ± 0.04 † | 0.35 ± 0.04 ‡ | 0.031 | 0.01 |
| CpG9 | 0.16 ± 0.02 | 0.25 ± 0.03 | 0.27 ± 0.03 ‡ | 0.058 | 0.011 |
| CpG10 | 0.2 ± 0.02 | 0.31 ± 0.04 | 0.33 ± 0.04 ‡ | 0.088 | 0.028 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, J.E.; You, Y.A.; Kwon, E.J.; Kim, S.M.; Lee, J.; Han, K.H.; Kim, Y.J. Maternal Malnutrition Affects Hepatic Metabolism through Decreased Hepatic Taurine Levels and Changes in HNF4A Methylation. Int. J. Mol. Sci. 2020, 21, 9060. https://doi.org/10.3390/ijms21239060
Du JE, You YA, Kwon EJ, Kim SM, Lee J, Han KH, Kim YJ. Maternal Malnutrition Affects Hepatic Metabolism through Decreased Hepatic Taurine Levels and Changes in HNF4A Methylation. International Journal of Molecular Sciences. 2020; 21(23):9060. https://doi.org/10.3390/ijms21239060
Chicago/Turabian StyleDu, Ji Eun, Young Ah You, Eun Jin Kwon, Soo Min Kim, Jeongae Lee, Ki Hwan Han, and Young Ju Kim. 2020. "Maternal Malnutrition Affects Hepatic Metabolism through Decreased Hepatic Taurine Levels and Changes in HNF4A Methylation" International Journal of Molecular Sciences 21, no. 23: 9060. https://doi.org/10.3390/ijms21239060
APA StyleDu, J. E., You, Y. A., Kwon, E. J., Kim, S. M., Lee, J., Han, K. H., & Kim, Y. J. (2020). Maternal Malnutrition Affects Hepatic Metabolism through Decreased Hepatic Taurine Levels and Changes in HNF4A Methylation. International Journal of Molecular Sciences, 21(23), 9060. https://doi.org/10.3390/ijms21239060