Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Venoms through History
2. Venom Evolution
3. Pore-Forming Proteins
4. Cnidarian’s Pore Forming Proteins
5. Sea Anemone Actinoporins
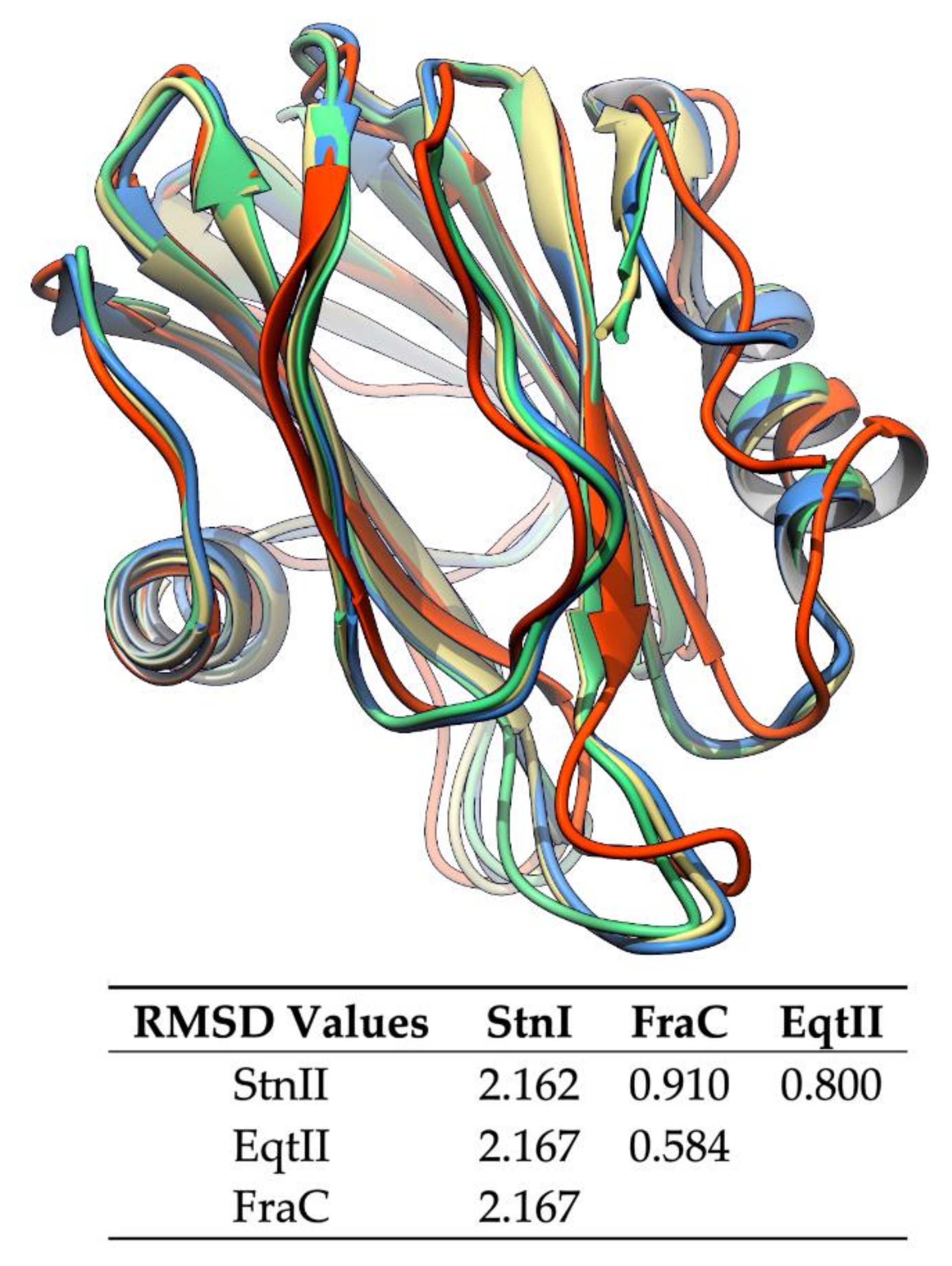
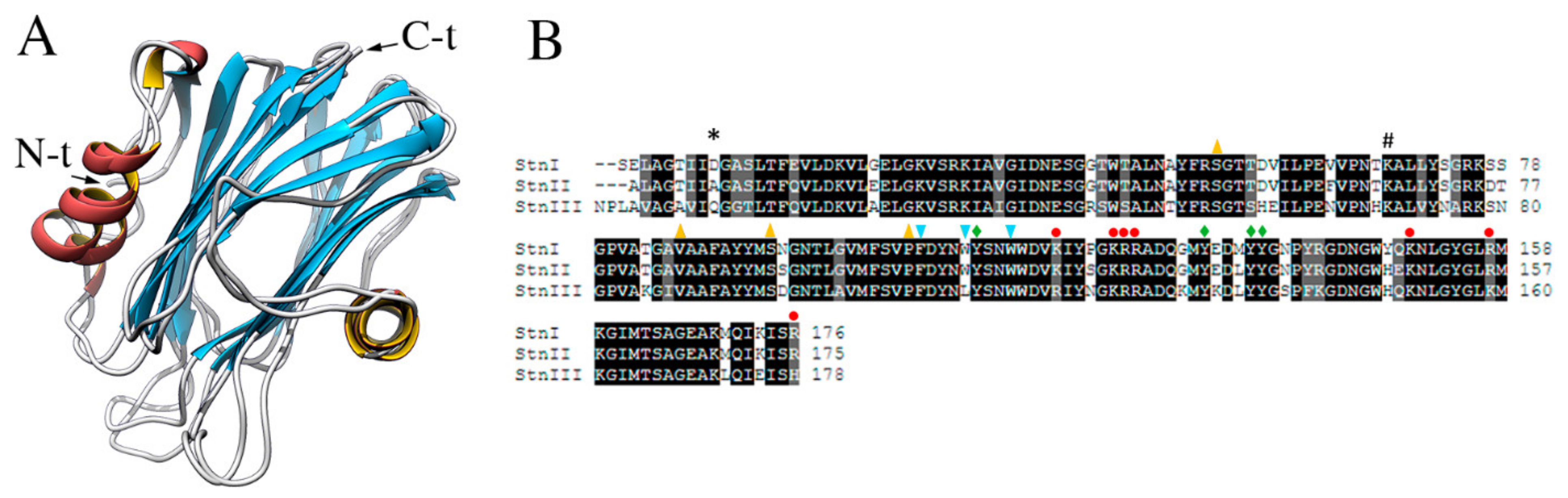
6. Sticholysins I, II, and III
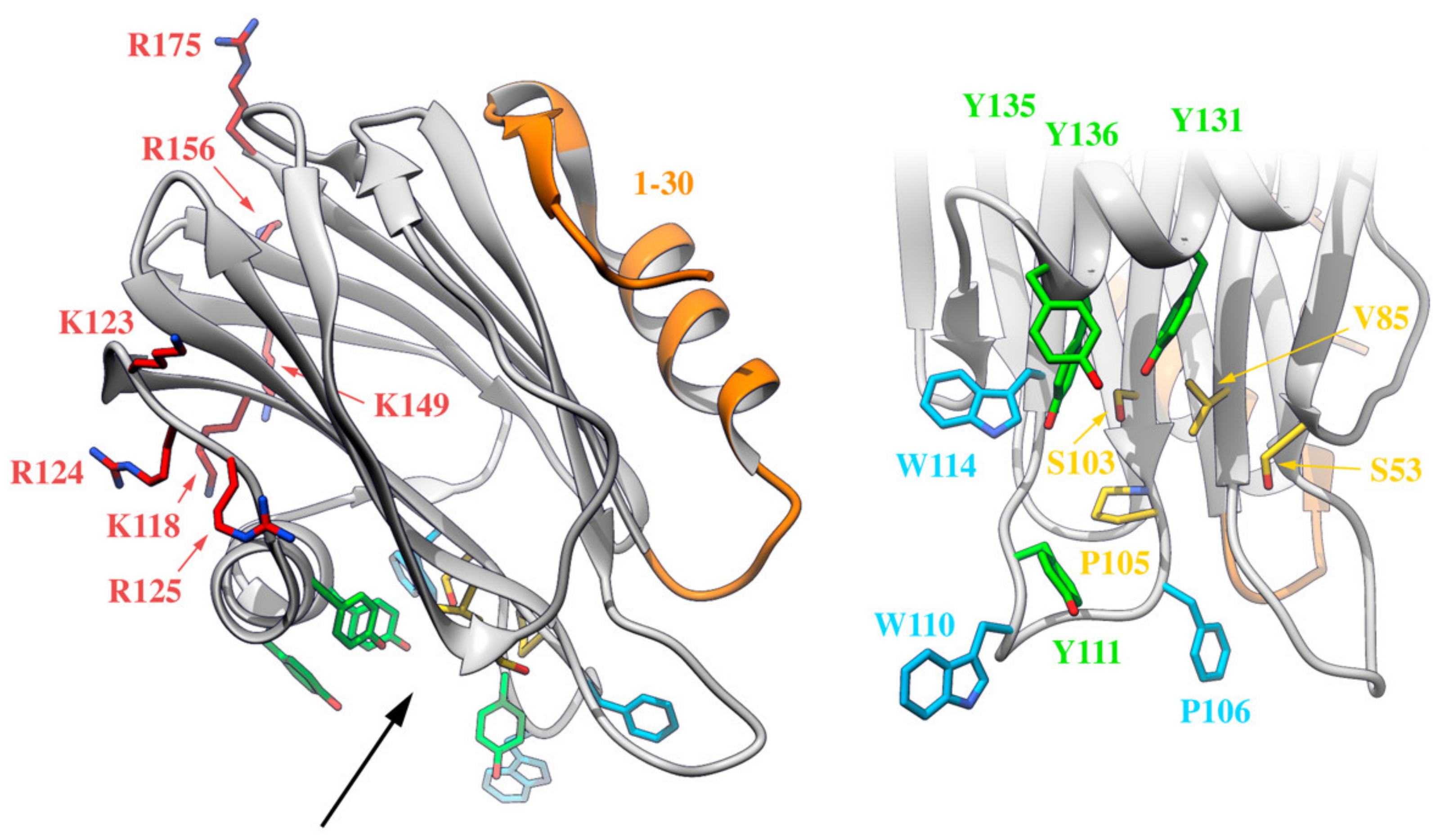
7. The N-Terminal Stretch of the Different Sticholysins
8. The Nature of the Sticholysins’ Functional Pore
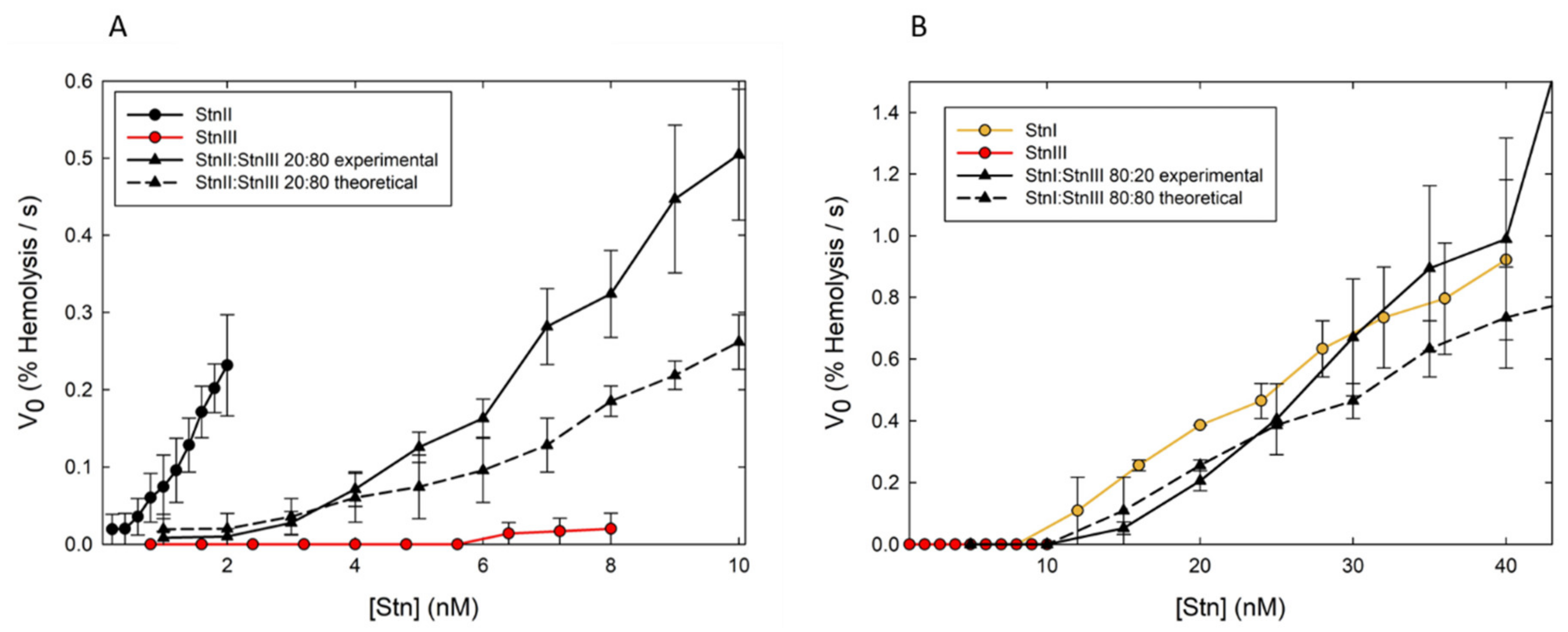
9. Synergy among the Different Sticholysins
10. The Influence of Cholesterol
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BAK | Bcl-2 homologous antagonist/killer |
| BAX | Bcl-2 associated X protein |
| CfTX | Chinorex fleckeri toxin |
| Chol | cholesterol |
| CTL | cholestatrienol |
| DOPC | dioleyl-phosphatidylcholine |
| Eqt | equinatoxin |
| Fra | fragaceatoxin |
| FRET | fluorescence resonance energy transfer |
| MACPF | membrane attack complex/perforin family |
| PFP | pore-forming protein |
| PLA2 | phospholipase A2 |
| POC | phosphorylcholine |
| POPE-NBD | 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(7-nitro-2-1,3-benzoxadiazol-4-yl) |
| R6G | Rhodamine 6G |
| RHA | relative hemolytic activity |
| RMB | relative membrane binding |
| SM | sphingomyelin |
| Stn | sticholysin |
| wt | wild-type |
References
- Plato. The Last Days of Socrates; Penguin: London, UK, 1959. [Google Scholar]
- Burstein, S.M. The Reign of Cleopatra; Greenwood Press: Westport, CT, USA, 2004. [Google Scholar]
- Roller, D.W. Cleopatra: A Biography; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Wink, M. A Short History of Alkaloids. In Alkaloids: Biochemistry, Ecology, and Medicinal Applications; Roberts, M.F., Ed.; Plenum Press: New York, NY, USA, 1998; pp. 11–44. [Google Scholar]
- Timbrell, J. The Poison Paradox: Chemicals as Friends and Foes; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Karamanou, M.; Androutsos, G.; Hayes, A.W.; Tsatsakis, A.M. Toxicology in the Borgias period: The mystery of Cantarella poison. Toxicol. Res. Appl. 2018, 2, 1–3. [Google Scholar] [CrossRef]
- King, G.F. Venoms as a Platform for Human Drugs: Translating Toxins into Therapeutics. Expert Opin. Biol. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Machkour-M’Rabet, S.; Henaut, Y.; Winterton, P.; Rojo, R. A Case of Zootherapy with the Tarantula Brachypelma Vagans Ausserer, 1875 in Traditional Medicine of the Chol Mayan Ethnic Group in Mexico. J. Ethnobiol. Ethnomed. 2011, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wuster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Arbuckle, K.; Rodríguez de la Vega, R.C.; Casewell, N.R. Coevolution Takes the Sting out of It: Evolutionary Biology and Mechanisms of Toxin Resistance in Animals. Toxicon 2017, 140, 118–131. [Google Scholar] [CrossRef]
- Calvete, J.J. Venomics: Integrative venom proteomics and beyond. Biochem. J. 2017, 474, 611–634. [Google Scholar] [CrossRef]
- Anderluh, G.; Krizaj, I.; Strukelj, B.; Gubensek, F.; Maček, P.; Pungercar, J. Equinatoxins, pore-forming proteins from the sea anemone Actinia equina, belong to a multigene family. Toxicon 1999, 37, 1391–1401. [Google Scholar] [CrossRef]
- Hallen, H.E.; Luo, H.; Scott-Craig, J.S.; Walton, J.D. Gene family encoding the major toxins of lethal Amanita mushrooms. Proc. Natl. Acad. Sci. USA 2007, 104, 19097–19101. [Google Scholar] [CrossRef]
- Wang, Y.; Yap, L.L.; Chua, K.L.; Khoo, H.E. A multigene family of Heteractis magnificalysins (HMgs). Toxicon 2008, 51, 1374–1382. [Google Scholar] [CrossRef]
- Clarke, T.H.; Garb, J.E.; Hayashi, C.Y.; Haney, R.A.; Lancaster, A.K.; Corbett, S.; Ayoub, N.A. Multi-tissue transcriptomics of the black widow spider reveals expansions, co-options, and functional processes of the silk gland gene toolkit. BMC Genom. 2014, 15, 365. [Google Scholar] [CrossRef]
- Clarke, T.H.; Garb, J.E.; Hayashi, C.Y.; Arensburger, P.; Ayoub, N.A. Spider Transcriptomes Identify Ancient Large-Scale Gene Duplication Event Potentially Important in Silk Gland Evolution. Genome Biol. Evol 2015, 7, 1856–1870. [Google Scholar] [CrossRef] [PubMed]
- Valle, A.; Alvarado-Mesen, J.; Lanio, M.E.; Alvarez, C.; Barbosa, J.A.; Pazos, I.F. The multigene families of actinoporins (part I): Isoforms and genetic structure. Toxicon 2015, 103, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Leychenko, E.V.; Isaeva, M.; Tkacheva, E.; Zelepuga, E.; Kvetkina, A.; Guzev, K.; Monastyrnaya, M.; Kozlovskaya, E. Multigene Family of Pore-Forming Toxins from Sea Anemone Heteractis crispa. Mar. Drugs 2018, 16, 183. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; Martínez-del-Pozo, A.; Garb, J.E. Stichodactyla helianthus’ de novo transcriptome assembly: Discovery of a new actinoporin isoform. Toxicon 2018, 150, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Trento, M.V.C.; Sales, T.A.; de Abreu, T.S.; Braga, M.A.; Cesar, P.H.S.; Marques, T.R.; Marcussi, S. Exploring the structural and functional aspects of the phospholipase A2 from Naja spp. Int. J. Biol. Macromol. 2019, 140, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Goñi, F.M. The basic structure and dynamics of cell membranes: An update of the Singer-Nicolson model. Biochim. Biophys. Acta 2014, 1838, 1467–1476. [Google Scholar] [CrossRef]
- Abdel Ghani, E.M.; Weis, S.; Walev, I.; Kehoe, M.; Bhakdi, S.; Palmer, M. Streptolysin O: Inhibition of the conformational change during membrane binding of the monomer prevents oligomerization and pore formation. Biochemistry 1999, 38, 15204–15211. [Google Scholar] [CrossRef]
- Parker, M.W.; Feil, S.C. Pore-forming protein toxins: From structure to function. Prog. Biophys. Mol. Biol. 2005, 88, 91–142. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Oñaderra, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Sea anemone actinoporins: The transition from a folded soluble state to a functionally active membrane-bound oligomeric pore. Curr. Protein Pept. Sci. 2007, 8, 558–572. [Google Scholar] [CrossRef]
- García-Ortega, L.; Alegre-Cebollada, J.; García-Linares, S.; Bruix, M.; Martínez-del-Pozo, A.; Gavilanes, J.G. The behavior of sea anemone actinoporins at the water-membrane interface. Biochim. Biophys. Acta 2011, 1808, 2275–2288. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.J.C.; Dalla Serra, M.; Froelich, C.J.; Wallace, M.I.; Anderluh, G. Membrane pore formation at protein-lipid interfaces. Trends Biochem. Sci. 2014, 39, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Rojko, N.; Anderluh, G. How Lipid Membranes Affect Pore Forming Toxin Activity. ACC Chem. Res. 2015, 48, 3073–3079. [Google Scholar] [CrossRef] [PubMed]
- Ros, U.; García-Sáez, A.J. More than a pore: The interplay of pore-forming proteins and lipid membranes. J. Membr. Biol. 2015, 248, 545–561. [Google Scholar] [CrossRef]
- Y Yamaji-Hasegawa, A.; Hullin-Matsuda, F.; Greimel, P.; Kobayashi, T. Pore-forming toxins: Properties, diversity, and uses as tools to image sphingomyelin and ceramide phosphoethanolamine. Biochim. Biophys. Acta 2015, 1858, 576–592. [Google Scholar] [CrossRef]
- Cosentino, K.; Ros, U.; García-Saez, A.J. Assembling the puzzle: Oligomerization of a-pore forming proteins in membranes. Biochim. Biophys. Acta 2016, 1858, 457–466. [Google Scholar] [CrossRef]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; Gavilanes, J.G.; Martínez-del-Pozo, A.; García-Linares, S. Pore-Forming Proteins from Cnidarians and Arachnids as Potential Biotechnological Tools. Toxins 2019, 11, 370. [Google Scholar] [CrossRef]
- Lahiani, A.; Yavin, E.; Lazarovici, P. The Molecular Basis of Toxins’ Interactions with Intracellular Signaling via Discrete Portals. Toxins 2017, 9, 107. [Google Scholar] [CrossRef]
- Tanaka, K.; Caaveiro, J.M.; Tsumoto, K. Bidirectional Transformation of a Metamorphic Protein between the Water-Soluble and Transmembrane Native States. Biochemistry 2015, 54, 6863–6866. [Google Scholar] [CrossRef]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- García-Linares, S.; Rivera-de-Torre, E.; Palacios-Ortega, J.; Gavilanes, J.G.; Martínez-del-Pozo, A. The metamorphic transformation of a water-soluble monomeric protein into an oligomeric transmembrane pore. In Advances in Biomembranes and Lipid Self-Assembly; Iglič, A., Rappolt, M., García-Sáez, A.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 26, pp. 51–97. [Google Scholar]
- Lella, M.; Mahalakshmi, R. Metamorphic Proteins: Emergence of Dual Protein Folds from One Primary Sequence. Biochemistry 2017, 56, 2971–2984. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, A.W.; Avigad, L.S. Properties of a toxin from the sea anemone Stoichactis helianthus, including specific binding to sphingomyelin. Proc. Natl. Acad. Sci. USA 1976, 73, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Linder, R.; Bernheimer, A.W.; Kim, K.S. Interaction between sphingomyelin and a cytolysin from the sea anemone Stoichactis helianthus. Biochim. Biophys. Acta 1977, 467, 290–300. [Google Scholar] [CrossRef]
- Maček, P. Polypeptide cytolytic toxins from sea anemones (Actiniaria). FEMS Microbiol. Immunol. 1992, 5, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Flores-Romero, H.; Ros, U.; García-Sáez, A.J. Pore formation in regulated cell death. EMBO J. 2020, e105753. [Google Scholar] [CrossRef] [PubMed]
- Panchal, R.G.; Smart, M.L.; Bowser, D.N.; Williams, D.A.; Petrou, S. Pore-forming proteins and their application in biotechnology. Curr. Pharm. Biotechnol. 2002, 3, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Praper, T.; Sonnen, A.; Viero, G.; Kladnik, A.; Froelich, C.J.; Anderluh, G.; Dalla Serra, M.; Gilbert, R.J. Human perforin employs different avenues to damage membranes. J. Biol. Chem. 2011, 286, 2946–2955. [Google Scholar] [CrossRef]
- Anderluh, G.; Lakey, J.H. Disparate proteins use similar architectures to damage membranes. Trends Biochem. Sci. 2008, 33, 482–490. [Google Scholar] [CrossRef]
- Kondos, S.C.; Hatfaludi, T.; Voskoboinik, I.; Trapani, J.A.; Law, R.H.; Whisstock, J.C.; Dunstone, M.A. The structure and function of mammalian membrane-attack complex/perforin-like proteins. Tissue Antigens 2010, 76, 341–351. [Google Scholar] [CrossRef]
- Law, R.H.; Lukoyanova, N.; Voskoboinik, I.; Caradoc-Davies, T.T.; Baran, K.; Dunstone, M.A.; D’Angelo, M.E.; Orlova, E.V.; Coulibaly, F.; Verschoor, S.; et al. The structural basis for membrane binding and pore formation by lymphocyte perforin. Nature 2010, 468, 447–451. [Google Scholar] [CrossRef]
- Dorca-Arevalo, J.; Blanch, M.; Pradas, M.; Blasi, J. Epsilon toxin from Clostridium perfringens induces cytotoxicity in FRT thyroid epithelial cells. Anaerobe 2018, 53, 43–49. [Google Scholar] [CrossRef] [PubMed]
- González-Bullón, D.; Uribe, K.B.; Largo, E.; Guembelzu, G.; García-Arribas, A.B.; Martín, C.; Ostolaza, H. Membrane Permeabilization by Bordetella Adenylate Cyclase Toxin Involves Pores of Tunable Size. Biomolecules 2019, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Ostolaza, H.; Gonzalez-Bullon, D.; Uribe, K.B.; Martin, C.; Amuategi, J.; Fernandez-Martinez, X. Membrane Permeabilization by Pore-Forming RTX Toxins: What Kind of Lesions Do These Toxins Form? Toxins 2019, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Hadders, M.A.; Beringer, D.X.; Gros, P. Structure of C8a-MACPF Reveals Mechanism of Membrane Attack in Complement Immune Defense. Science 2007, 317, 1552–1554. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.J. Protein-lipid interactions and non-lamellar lipidic structures in membrane pore formation and membrane fusion. Biochim. Biophys. Acta 2015. [Google Scholar] [CrossRef] [PubMed]
- Reboul, C.F.; Whisstock, J.C.; Dunstone, M.A. Giant MACPF/CDC pore forming toxins: A class of their own. Biochim. Biophys. Acta 2015, 1858, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Wade, K.R.; Tweten, R.K. The Apicomplexan CDC/MACPF-like pore-forming proteins. Curr. Opin. Microbiol. 2015, 26, 48–52. [Google Scholar] [CrossRef]
- Grosse, L.; Wurm, C.A.; Bruser, C.; Neumann, D.; Jans, D.C.; Jakobs, S. Bax assembles into large ring-like structures remodeling the mitochondrial outer membrane in apoptosis. EMBO J 2016, 35, 402–413. [Google Scholar] [CrossRef]
- Cosentino, K.; García-Sáez, A.J. MIM through MOM: The awakening of Bax and Bak pores. EMBO J. 2018, 37. [Google Scholar] [CrossRef]
- McArthur, K.; Whitehead, L.W.; Heddleston, J.M.; Li, L.; Padman, B.S.; Oorschot, V.; Geoghegan, N.D.; Chappaz, S.; Davidson, S.; San Chin, H.; et al. BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 2018, 359. [Google Scholar] [CrossRef]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond—Mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Cowan, A.D.; Smith, N.A.; Sandow, J.J.; Kapp, E.A.; Rustam, Y.H.; Murphy, J.M.; Brouwer, J.M.; Bernardini, J.P.; Roy, M.J.; Wardak, A.Z.; et al. BAK core dimers bind lipids and can be bridged by them. Nat. Struct. Mol. Biol. 2020, 27, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Flores-Romero, H.; García-Sáez, A.J. Lipids glue BAK dimers together. Nat. Struct. Mol. Biol. 2020, 27, 1003–1004. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Feil, S.C.; Mckinstry, W.J.; Tweten, R.K.; Parker, M.W. Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 1997, 89, 685–692. [Google Scholar] [CrossRef]
- Shatursky, O.; Heuck, A.P.; Shepard, L.A.; Rossjohn, J.; Parker, M.W.; Johnson, A.E.; Tweten, R.K. The mechanism of membrane insertion for a cholesterol-dependent cytolysin: A novel paradigm for pore-forming toxins. Cell 1999, 99, 293–299. [Google Scholar] [CrossRef]
- Giddings, K.S.; Johnson, A.E.; Tweten, R.K. Redefining cholesterol’s role in the mechanism of the cholesterol-dependent cytolysins. Proc. Natl. Acad. Sci. USA 2003, 100, 11315–11320. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Tweten, R.K.; Johnson, A.E. The domains of a cholesterol-dependent cytolysin undergo a major FRET-detected rearrangement during pore formation. Proc. Natl. Acad. Sci. USA 2005, 102, 7139–7144. [Google Scholar] [CrossRef]
- Giesemann, T.; Jank, T.; Gerhard, R.; Maier, E.; Just, I.; Benz, R.; Aktories, K. Cholesterol-dependent Pore Formation of Clostridium difficile Toxin A. J. Biol. Chem. 2006, 281, 10808–10815. [Google Scholar] [CrossRef]
- Bavdek, A.; Gekara, N.O.; Priselac, D.; Aguirre, I.G.; Darji, A.; Chakraborty, T.; Maček, P.; Lakey, J.H.; Weiss, S.; Anderluh, G. Sterol and pH Interdependence in the Binding, Oligomerization, and Pore Formation of Listeriolysin O. Biochemistry 2007, 46, 4425–4437. [Google Scholar] [CrossRef]
- Rosado, C.J.; Buckle, A.M.; Law, R.H.; Butcher, R.E.; Kan, W.T.; Bird, C.H.; Ung, K.; Browne, K.A.; Baran, K.; Bashtannyk-Puhalovich, T.A.; et al. A common fold mediates vertebrate defense and bacterial attack. Science 2007, 317, 1548–1551. [Google Scholar] [CrossRef]
- Rossjohn, J.; Polekhina, G.; Feil, S.C.; Morton, C.J.; Tweten, R.K.; Parker, M.W. Structures of Perfringolysin O Suggest a Pathway for Activation of Cholesterol-dependent Cytolysins. J. Mol. Biol. 2007, 367, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Hotze, E.M.; Tweten, R.K. Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim. Biophys. Acta 2011, 1818, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Ota, K.; Leonardi, A.; Mikelj, M.; Skocaj, M.; Wohlschlager, T.; Kunzler, M.; Aebi, M.; Narat, M.; Krizaj, I.; Anderluh, G.; et al. Membrane cholesterol and sphingomyelin, and ostreolysin A are obligatory for pore-formation by a MACPF/CDC-like pore-forming protein, pleurotolysin B. Biochimie 2013, 95, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Shewell, L.K.; Harvey, R.M.; Higgins, M.A.; Day, C.J.; Hartley-Tassell, L.E.; Chen, A.Y.; Gillen, C.M.; James, D.B.; Alonzo, F., III; Torres, V.J.; et al. The cholesterol-dependent cytolysins pneumolysin and streptolysin O require binding to red blood cell glycans for hemolytic activity. Proc. Natl. Acad. Sci. USA 2014, 111, E5312–E5320. [Google Scholar] [CrossRef] [PubMed]
- Ellisdon, A.M.; Reboul, C.F.; Panjikar, S.; Huynh, K.; Oellig, C.A.; Winter, K.L.; Dunstone, M.A.; Hodgson, W.C.; Seymour, J.; Dearden, P.K.; et al. Stonefish toxin defines an ancient branch of the perforin-like superfamily. Proc. Natl. Acad. Sci. USA 2015, 112, 15360–15365. [Google Scholar] [CrossRef]
- Kathuria, R.; Mondal, A.K.; Sharma, R.; Bhattacharyya, S.; Chattopadhyay, K. Revisiting the role of cholesterol in regulating the pore-formation mechanism of Vibrio cholerae cytolysin, a membrane-damaging beta-barrel pore-forming toxin. Biochem. J. 2018, 475, 3039–3055. [Google Scholar] [CrossRef]
- Vogele, M.; Bhaskara, R.M.; Mulvihill, E.; van Pee, K.; Yildiz, O.; Kuhlbrandt, W.; Muller, D.J.; Hummer, G. Membrane perforation by the pore-forming toxin pneumolysin. Proc. Natl. Acad. Sci. USA 2019, 116, 13352–13357. [Google Scholar] [CrossRef]
- Caaveiro, J.M.; Echabe, I.; Gutiérrez-Aguirre, I.; Nieva, J.L.; Arrondo, J.L.; González-Mañas, J.M. Differential interaction of equinatoxin II with model membranes in response to lipid composition. Biophys. J. 2001, 80, 1343–1353. [Google Scholar] [CrossRef]
- Valcarcel, C.A.; Dalla Serra, M.; Potrich, C.; Bernhart, I.; Tejuca, M.; Martínez, D.; Pazos, F.; Lanio, M.E.; Menestrina, G. Effects of lipid composition on membrane permeabilization by sticholysin I and II, two cytolysins of the sea anemone Stichodactyla helianthus. Biophys. J. 2001, 80, 2761–2774. [Google Scholar] [CrossRef]
- Bonev, B.B.; Lam, Y.H.; Anderluh, G.; Watts, A.; Norton, R.S.; Separovic, F. Effects of the Eukaryotic Pore-Forming Cytolysin Equinatoxin II on Lipid Membranes and the Role of Sphingomyelin. Biophys. J. 2003, 84, 2382–2392. [Google Scholar] [CrossRef]
- Barlič, A.; Gutiérrez-Aguirre, I.; Caaveiro, J.M.; Cruz, A.; Ruiz-Argüello, M.B.; Pérez-Gil, J.; González-Mañas, J.M. Lipid phase coexistence favors membrane insertion of equinatoxin-II, a pore-forming toxin from Actinia equina. J. Biol. Chem. 2004, 279, 34209–34216. [Google Scholar] [CrossRef] [PubMed]
- Martínez, D.; Otero, A.; Álvarez, C.; Pazos, F.; Tejuca, M.; Lanio, M.E.; Gutiérrez-Aguirre, I.; Barlič, A.; Iloro, I.; Arrondo, J.L.; et al. Effect of sphingomyelin and cholesterol on the interaction of St II with lipidic interfaces. Toxicon 2007, 49, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Bakrač, B.; Gutierrez-Aguirre, I.; Podlesek, Z.; Sonnen, A.F.; Gilbert, R.J.; Maček, P.; Lakey, J.H.; Anderluh, G. Molecular determinants of sphingomyelin specificity of a eukaryotic pore-forming toxin. J. Biol. Chem. 2008, 283, 18665–18677. [Google Scholar] [CrossRef] [PubMed]
- Schön, P.; García-Saez, A.J.; Malovrh, P.; Bacia, K.; Anderluh, G.; Schwille, P. Equinatoxin II permeabilizing activity depends on the presence of sphingomyelin and lipid phase coexistence. Biophys. J. 2008, 95, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Mancheño, J.M.; Martínez, D.; Tejuca, M.; Pazos, F.; Lanio, M.E. Sticholysins, two pore-forming toxins produced by the Caribbean sea anemone Stichodactyla helianthus: Their interaction with membranes. Toxicon 2009, 54, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Bakrač, B.; Anderluh, G. Molecular mechanism of sphingomyelin-specific membrane binding and pore formation by actinoporins. Adv. Exp. Med. Biol. 2009, 677, 106–115. [Google Scholar]
- Razpotnik, A.; Krizaj, I.; Kem, W.R.; Maček, P.; Turk, T. A new cytolytic protein from the sea anemone Urticina crassicornis that binds to cholesterol- and sphingomyelin-rich membranes. Toxicon 2009, 53, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.S.; Chieppa, G.; Desideri, A.; Cannata, S.; Romano, E.; Luly, P.; Rufini, S. Sticholysin II: A pore-forming toxin as a probe to recognize sphingomyelin in artificial and cellular membranes. Toxicon 2012, 60, 724–733. [Google Scholar] [CrossRef]
- Maula, T.; Isaksson, Y.J.; García-Linares, S.; Niinivehmas, S.; Pentikainen, O.T.; Kurita, M.; Yamaguchi, S.; Yamamoto, T.; Katsumura, S.; Gavilanes, J.G.; et al. 2NH and 3OH are crucial structural requirements in sphingomyelin for sticholysin II binding and pore formation in bilayer membranes. Biochim. Biophys. Acta 2013, 1828, 1390–1395. [Google Scholar] [CrossRef]
- García-Linares, S.; Palacios-Ortega, J.; Yasuda, T.; Astrand, M.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Toxin-induced pore formation is hindered by intermolecular hydrogen bonding in sphingomyelin bilayers. Biochim. Biophys. Acta 2016, 1858, 1189–1195. [Google Scholar] [CrossRef]
- García-Linares, S.; Rivera-de-Torre, E.; Morante, K.; Tsumoto, K.; Caaveiro, J.M.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, Á. Differential effect of membrane composition on the pore-forming ability of four different sea anemone actinoporins. Biochemistry 2016, 55, 6630–6641. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; García-Linares, S.; Astrand, M.; Al Sazzad, M.A.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Regulation of Sticholysin II-Induced Pore Formation by Lipid Bilayer Composition, Phase State, and Interfacial Properties. Langmuir 2016, 32, 3476–3484. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Yamaji-Hasegawa, A.; Hullin-Matsuda, F.; Kobayashi, T. Molecular mechanisms of action of sphingomyelin-specific pore-forming toxin, lysenin. Semin. Cell Dev. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Endapally, S.; Frias, D.; Grzemska, M.; Gay, A.; Tomchick, D.R.; Radhakrishnan, A. Molecular Discrimination between Two Conformations of Sphingomyelin in Plasma Membranes. Cell 2019, 176, 1040–1053.e17. [Google Scholar] [CrossRef]
- Palacios-Ortega, J.; García-Linares, S.; Rivera-de-Torre, E.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Sticholysin, Sphingomyelin, and Cholesterol: A Closer Look at a Tripartite Interaction. Biophys. J. 2019, 116, 2253–2265. [Google Scholar] [CrossRef]
- Dulubova, I.E.; Krasnoperov, V.G.; Khvotchev, M.V.; Pluzhnikov, K.A.; Volkova, T.M.; Grishin, E.V.; Vais, H.; Bell, D.R.; Usherwood, P.N. Cloning and structure of d-latroinsectotoxin, a novel insect-specific member of the latrotoxin family: Functional expression requires C-terminal truncation. J. Biol. Chem. 1996, 271, 7535–7543. [Google Scholar] [CrossRef]
- Ales, E.; Gabilan, N.H.; Cano-Abad, M.F.; Garcia, A.G.; Lopez, M.G. The sea anemone toxin Bc2 induces continuous or transient exocytosis, in the presence of sustained levels of high cytosolic Ca2+ in chromaffin cells. J. Biol. Chem. 2000, 275, 37488–37495. [Google Scholar] [CrossRef]
- Ashton, A.C.; Rahman, M.A.; Volynski, K.E.; Manser, C.; Orlova, E.V.; Matsushita, H.; Davletov, B.A.; van Heel, M.; Grishin, E.V.; Ushkaryov, Y.A. Tetramerisation of a-latrotoxin by divalent cations is responsible for toxin-induced non-vesicular release and contributes to the Ca+2-dependent vesicular exocytosis from synaptosomes. Biochimie 2000, 82, 453–468. [Google Scholar] [CrossRef]
- Orlova, E.V.; Rahman, M.A.; Gowen, B.; Volynski, K.E.; Ashton, A.C.; Manser, C.; van Heel, M.; Ushkaryov, Y.A. Structure of a-latrotoxin oligomers reveals that divalent cation-dependent tetramers form membrane pores. Nat. Struct. Biol. 2000, 7, 48–53. [Google Scholar]
- Yan, S.; Wang, X.C. Recent Advances in Research on Widow Spider Venoms and Toxins. Toxins 2015, 7, 5055–5067. [Google Scholar] [CrossRef]
- Rohou, A.; Nield, J.; Ushkaryov, Y.A. Insecticidal toxins from black widow spider venom. Toxicon 2007, 49, 531–549. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martínez-Del-Pozo, A. Evaluation of different approaches used to study membrane permeabilization by actinoporins on model lipid vesicles. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183311. [Google Scholar] [CrossRef] [PubMed]
- Sher, D.; Knebel, A.; Bsor, T.; Nesher, N.; Tal, T.; Morgenstern, D.; Cohen, E.; Fishman, Y.; Zlotkin, E. Toxic polypeptides of the hydra-a bioinformatic approach to cnidarian allomones. Toxicon 2005, 45, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.A.; Mayhew, M.L.; Jin, J.; Herzig, V.; Undheim, E.A.B.; Sombke, A.; Fry, B.G.; Meritt, D.J.; King, G.F. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nat. Commun. 2018, 9, 755. [Google Scholar] [CrossRef]
- Clark, G.C.; Casewell, N.R.; Elliott, C.T.; Harvey, A.L.; Jamieson, A.G.; Strong, P.N.; Turner, A.D. Friends or Foes? Emerging Impacts of Biological Toxins. Trends Biochem. Sci. 2019, 44, 365–379. [Google Scholar] [CrossRef]
- Steele, R.E.; David, C.N.; Technau, U. A genomic view of 500 million years of cnidarian evolution. Trends Genet. 2011, 27, 7–13. [Google Scholar] [CrossRef]
- Technau, U.; Steele, R.E. Evolutionary crossroads in developmental biology: Cnidaria. Development 2011, 138, 1447–1458. [Google Scholar] [CrossRef]
- Kayal, E.; Bentlage, B.; Sabrina Pankey, M.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Koch, T.L.; Grimmelikhuijzen, C.J.P. Global Neuropeptide Annotations From the Genomes and Transcriptomes of Cubozoa, Scyphozoa, Staurozoa (Cnidaria: Medusozoa), and Octocorallia (Cnidaria: Anthozoa). Front. Endocrinol. 2019, 10, 831. [Google Scholar] [CrossRef]
- Beadnell, C.E.; Rider, T.A.; Williamson, J.A.; Fenner, P.J. Management of a major box jellyfish (Chironex fleckeri) sting. Lessons from the first minutes and hours. Med. J. Aust. 1992, 156, 655–658. [Google Scholar] [CrossRef]
- Andreosso, A.; Bansal, P.S.; Smout, M.J.; Wilson, D.; Seymour, J.E.; Daly, N.L. Structural Characterisation of Predicted Helical Regions in the Chironex fleckeri CfTX-1 Toxin. Mar. Drugs 2018, 16, 201. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.T.; Manion, J.; Littleboy, J.B.; Oyston, L.; Khuong, T.M.; Wang, Q.P.; Nguyen, D.T.; Hesselson, D.; Seymour, J.E.; Neely, G.G. Molecular dissection of box jellyfish venom cytotoxicity highlights an effective venom antidote. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-seq approach to identify putative toxins from acrorhagi in aggressive and non-aggressive Anthopleura elegantissima polyps. BMC Genom. 2015, 16, 221. [Google Scholar] [CrossRef] [PubMed]
- Rojko, N.; Dalla Serra, M.; Maček, P.; Anderluh, G. Pore formation by actinoporins, cytolysins from sea anemones. Biochim. Biophys. Acta 2016, 1858, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Broe, M.; Daly, M. Multi-copy venom genes hidden in de novo transcriptome assemblies, a cautionary tale with the snakelocks sea anemone Anemonia sulcata (Pennant, 1977). Toxicon 2015, 108, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; Undheim, E.A.B.; King, G.F. Revisiting venom of the sea anemone Stichodactyla haddoni: Omics techniques reveal the complete toxin arsenal of a well-studied sea anemone genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef]
- Brinkman, D.; Burnell, J. Identification, cloning and sequencing of two major venom proteins from the box jellyfish, Chironex fleckeri. Toxicon 2007, 50, 850–860. [Google Scholar] [CrossRef]
- Brinkman, D.L.; Konstantakopoulos, N.; McInerney, B.V.; Mulvenna, J.; Seymour, J.E.; Isbister, G.K.; Hodgson, W.C. Chironex fleckeri (box jellyfish) venom proteins: Expansion of a cnidarian toxin family that elicits variable cytolytic and cardiovascular effects. J. Biol. Chem. 2014, 289, 4798–4812. [Google Scholar] [CrossRef]
- Anderluh, G.; Maček, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar] [CrossRef]
- Glasser, E.; Rachamim, T.; Aharonovich, D.; Sher, D. Hydra actinoporin-like toxin-1, an unusual hemolysin from the nematocyst venom of Hydra magnipapillata which belongs to an extended gene family. Toxicon 2014, 91, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; García-Linares, S.; Alegre-Cebollada, J.; Lacadena, J.; Gavilanes, J.G.; Martínez-del-Pozo, A. Synergistic action of actinoporin isoforms from the same sea anemone species assembled into functionally active heteropores. J. Biol. Chem. 2016, 291, 14109–14119. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Daly, M. Evolution of the Cytolytic Pore-Forming Proteins (Actinoporins) in Sea Anemones. Toxins 2016, 8, 368. [Google Scholar] [CrossRef] [PubMed]
- Bjorkbom, A.; Rog, T.; Kaszuba, K.; Kurita, M.; Yamaguchi, S.; Lonnfors, M.; Nyholm, T.K.; Vattulainen, I.; Katsumura, S.; Slotte, J.P. Effect of Sphingomyelin Headgroup Size on Molecular Properties and Interactions with Cholesterol. Biophys. J. 2010, 99, 3300–3308. [Google Scholar] [CrossRef] [PubMed]
- Alm, I.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Cholesterol stimulates and ceramide inhibits sticholysin II-induced pore formation in complex bilayer membranes. Biochim. Biophys. Acta Biomembr. 2015, 1848, 925–931. [Google Scholar] [CrossRef]
- García-Linares, S.; Alm, I.; Maula, T.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A. The effect of cholesterol on the long-range network of interactions established among sea anemone Sticholysin II residues at the water-membrane interface. Mar. Drugs 2015, 13, 1647–1665. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadis, A.; Anderluh, G.; Maček, P.; Turk, D. Crystal structure of the soluble form of equinatoxin II, a pore-forming toxin from the sea anemone Actinia equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef]
- Hinds, M.G.; Zhang, W.; Anderluh, G.; Hansen, P.E.; Norton, R.S. Solution structure of the eukaryotic pore-forming cytolysin equinatoxin II: Implications for pore formation. J. Mol. Biol. 2002, 315, 1219–1229. [Google Scholar] [CrossRef]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef]
- Mechaly, A.E.; Bellomio, A.; Gil-Carton, D.; Morante, K.; Valle, M.; González-Mañas, J.M.; Guerin, D.M. Structural insights into the oligomerization and architecture of eukaryotic membrane pore-forming toxins. Structure 2011, 19, 181–191. [Google Scholar] [CrossRef]
- García-Linares, S.; Castrillo, I.; Bruix, M.; Menéndez, M.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G. Three-dimensional structure of the actinoporin sticholysin I. Influence of long-distance effects on protein function. Arch. Biochem. Biophys. 2013, 532, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Caaveiro, J.M.; Morante, K.; González-Mañas, J.M.; Tsumoto, K. Structural basis for self-assembly of a cytolytic pore lined by protein and lipid. Nat. Commun. 2015, 6, 6337. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, I.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Bruix, M. 1H, 13C, and 15N NMR assignments of StnII-R29Q, a defective lipid binding mutant of the sea anemone actinoporin sticholysin II. Biomol. NMR Assign. 2009, 3, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Goormaghtigh, E. Infrared spectroscopy study on the conformational changes leading to pore formation of the toxin sticholysin II. Biophys. J. 2007, 93, 3191–3201. [Google Scholar] [CrossRef] [PubMed]
- Subburaj, Y.; Ros, U.; Hermann, E.; Tong, R.; García-Sáez, A.J. Toxicity of an a-pore-forming toxin depends on the assembly mechanism on the target membrane as revealed by single-molecule imaging. J. Biol. Chem. 2015, 290, 4856–4865. [Google Scholar] [CrossRef]
- Morante, K.; Bellomio, A.; Gil-Cartón, D.; Redondo-Morata, L.; Sot, J.; Scheuring, S.; Valle, M.; González-Mañas, J.M.; Tsumoto, K.; Caaveiro, J.M.M. Identification of a membrane-bound prepore species clarifies the lytic mechanism of actinoporins. J. Biol. Chem. 2016, 291, 19210–19219. [Google Scholar] [CrossRef]
- Rojko, N.; Kristan, K.C.; Viero, G.; Zerovnik, E.; Maček, P.; Dalla Serra, M.; Anderluh, G. Membrane damage by an a-helical pore-forming protein, Equinatoxin II, proceeds through a succession of ordered steps. J. Biol. Chem. 2013, 288, 23704–23715. [Google Scholar] [CrossRef]
- Martín-Benito, J.; Gavilanes, F.; de Los Ríos, V.; Mancheño, J.M.; Fernández, J.J.; Gavilanes, J.G. Two-dimensional crystallization on lipid monolayers and three-dimensional structure of sticholysin II, a cytolysin from the sea anemone Stichodactyla helianthus. Biophys. J. 2000, 78, 3186–3194. [Google Scholar] [CrossRef]
- Mancheño, J.M.; Martín-Benito, J.; Gavilanes, J.G.; Vázquez, L. A complementary microscopy analysis of Sticholysin II crystals on lipid films: Atomic force and transmission electron characterizations. Biophys. Chem. 2005, 119, 219–223. [Google Scholar] [CrossRef]
- Maček, P.; Belmonte, G.; Pederzolli, C.; Menestrina, G. Mechanism of action of equinatoxin II, a cytolysin from the sea anemone Actinia equina L. belonging to the family of actinoporins. Toxicology 1994, 87, 205–227. [Google Scholar] [CrossRef]
- Kristan, K.C.; Viero, G.; Dalla Serra, M.; Maček, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Fauth, E.V.; Cilli, E.M.; Ligabue-Braun, R.; Verli, H. Differential Effect of Solution Conditions on the Conformation of the Actinoporins Sticholysin II and Equinatoxin II. Anais Acad. Bras. Cienc. 2014, 86, 1949–1962. [Google Scholar] [CrossRef] [PubMed]
- García-Linares, S.; Richmond, R.; García-Mayoral, M.F.; Bustamante, N.; Bruix, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. The sea anemone actinoporin (Arg-Gly-Asp) conserved motif is involved in maintaining the competent oligomerization state of these pore-forming toxins. FEBS J. 2014, 281, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Anderluh, G.; Razpotnik, A.; Podlesek, Z.; Maček, P.; Separovic, F.; Norton, R.S. Interaction of the eukaryotic pore-forming cytolysin equinatoxin II with model membranes: 19F NMR studies. J. Mol. Biol. 2005, 347, 27–39. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Cunietti, M.; Herrero-Galán, E.; Gavilanes, J.G.; Martínez-del-Pozo, A. Calorimetric scrutiny of lipid binding by sticholysin II toxin mutants. J. Mol. Biol. 2008, 382, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, I.; Araujo, N.A.; Alegre-Cebollada, J.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Bruix, M. Specific interactions of sticholysin I with model membranes: An NMR study. Proteins 2010, 78, 1959–1970. [Google Scholar] [CrossRef]
- Pedrera, L.; Fanani, M.L.; Ros, U.; Lanio, M.E.; Maggio, B.; Álvarez, C. Sticholysin I-membrane interaction: An interplay between the presence of sphingomyelin and membrane fluidity. Biochim. Biophys. Acta 2014, 1838, 1752–1759. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Aguirre, I.; Barlič, A.; Podlesek, Z.; Maček, P.; Anderluh, G.; González-Mañas, J.M. Membrane insertion of the N-terminal a-helix of equinatoxin II, a sea anemone cytolytic toxin. Biochem. J. 2004, 384 Pt 2, 421–428. [Google Scholar] [CrossRef]
- Kristan, K.; Podlesek, Z.; Hojnik, V.; Gutiérrez-Aguirre, I.; Guncar, G.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; Maček, P.; Anderluh, G. Pore Formation by Equinatoxin, a Eukaryotic Pore-forming Toxin, Requires a Flexible N-terminal Region and a Stable b-Sandwich. J. Biol. Chem. 2004, 279, 46509–46517. [Google Scholar] [CrossRef]
- Kristan, K.; Viero, G.; Maček, P.; Dalla Serra, M.; Anderluh, G. The equinatoxin N-terminus is transferred across planar lipid membranes and helps to stabilize the transmembrane pore. FEBS J. 2007, 274, 539–550. [Google Scholar] [CrossRef]
- Antonini, V.; Perez-Barzaga, V.; Bampi, S.; Penton, D.; Martinez, D.; Dalla Serra, M.; Tejuca, M. Functional Characterization of Sticholysin I and W111C Mutant Reveals the Sequence of the Actinoporin’s Pore Assembly. PLoS ONE 2014, 9, e110824. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; García-Linares, S.; Rivera-de-Torre, E.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Differential Effect of Bilayer Thickness on Sticholysin Activity. Langmuir 2017, 33, 11018–11027. [Google Scholar] [CrossRef] [PubMed]
- Carretero, G.P.B.; Vicente, E.F.; Cilli, E.M.; Alvarez, C.M.; Jenssen, H.; Schreier, S. Dissecting the mechanism of action of actinoporins. Role of the N-terminal amphipathic alpha-helix in membrane binding and pore activity of sticholysins I and II. PLoS ONE 2018, 13, e0202981. [Google Scholar] [CrossRef] [PubMed]
- Mechaly, A.E.; Bellomio, A.; Morante, K.; Agirre, J.; Gil-Carton, D.; Valle, M.; González-Mañas, J.M.; Guerin, D.M. Pores of the toxin FraC assemble into 2D hexagonal clusters in both crystal structures and model membranes. J. Struct. Biol. 2012, 180, 312–317. [Google Scholar] [CrossRef]
- Varanda, W.; Finkelstein, A. Ion and nonelectrolyte permeability properties of channels formed in planar lipid bilayer membranes by the cytolytic toxin from the sea anemone, Stoichactis helianthus. J. Membr. Biol. 1980, 55, 203–211. [Google Scholar] [CrossRef]
- Rojko, N.; Cronin, B.; Danial, J.S.; Baker, M.A.; Anderluh, G.; Wallace, M.I. Imaging the lipid-phase-dependent pore formation of equinatoxin II in droplet interface bilayers. Biophys. J. 2014, 106, 1630–1637. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Lacadena, V.; Oñaderra, M.; Mancheño, J.M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Phenotypic selection and characterization of randomly produced non-haemolytic mutants of the toxic sea anemone protein sticholysin II. FEBS Lett. 2004, 575, 14–18. [Google Scholar] [CrossRef]
- Hong, Q.; Gutiérrez-Aguirre, I.; Barlič, A.; Malovrh, P.; Kristan, K.; Podlesek, Z.; Maček, P.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; et al. Two-step Membrane Binding by Equinatoxin II, a Pore-forming Toxin from the Sea Anemone, Involves an Exposed Aromatic Cluster and a Flexible Helix. J. Biol. Chem. 2002, 277, 41916–41924. [Google Scholar] [CrossRef]
- Casallanovo, F.; de Oliveira, F.J.; de Souza, F.C.; Ros, U.; Martínez, Y.; Penton, D.; Tejuca, M.; Martínez, D.; Pazos, F.; Pertinhez, T.A.; et al. Model peptides mimic the structure and function of the N-terminus of the pore-forming toxin sticholysin II. Biopolymers 2006, 84, 169–180. [Google Scholar] [CrossRef]
- Ros, U.; Pedrera, L.; Diaz, D.; Karam, J.C.; Sudbrack, T.P.; Valiente, P.A.; MartInez, D.; Cilli, E.M.; Pazos, F.; Itri, R.; et al. The membranotropic activity of N-terminal peptides from the pore-forming proteins sticholysin I and II is modulated by hydrophobic and electrostatic interactions as well as lipid composition. J. Biosci. 2012, 36, 781–791. [Google Scholar] [CrossRef]
- Doyle, J.W.; Kem, W.R. Binding of a radiolabeled sea anemone cytolysin to erythrocyte membranes. Biochim. Biophys. Acta 1989, 987, 181–186. [Google Scholar] [CrossRef]
- Lanio, M.E.; Morera, V.; Álvarez, C.; Tejuca, M.; Gómez, T.; Pazos, F.; Besada, V.; Martínez, D.; Huerta, V.; Padrón, G.; et al. Purification and characterization of two hemolysins from Stichodactyla helianthus. Toxicon 2001, 39, 187–194. [Google Scholar] [CrossRef]
- Stevens, S.M., Jr.; Kem, W.R.; Prokai, L. Investigation of cytolysin variants by peptide mapping: Enhanced protein characterization using complementary ionization and mass spectrometric techniques. Rapid Commun. Mass Spectrom. 2002, 16, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.; Martinez, D.; Palmero, A.; Soto, C.; Tejuca, M.; Pazos, F.; Menendez, R.; Alvarez, C.; Garateix, A. Pharmacological effects of two cytolysins isolated from the sea anemone Stichodactyla helianthus. J. Biosci. 2009, 34, 891–898. [Google Scholar] [CrossRef]
- Del Monte-Martínez, A.; González-Bacerio, J.; Romero, L.; Aragon, C.; Martínez, D.; Chávez, M.D.; Álvarez, C.; Lanio, M.E.; Guisan, J.M.; Díaz, J. Improved purification and enzymatic properties of a mixture of sticholysin I and II: Isotoxins with hemolytic and phospholipase A activities from the sea anemone Stichodactyla helianthus. Protein Expr. Purif. 2013, 95, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; Garb, J.E.; Slotte, J.P.; Gavilanes, J.G.; Martínez-del-Pozo, A. Structural and functional characterization of sticholysin III: A newly discovered actinoporin within the venom of the sea anemone Stichodactyla helianthus. Arch. Biochem. Biophys. 2020, 689, 108435. [Google Scholar] [CrossRef] [PubMed]
- Martínez, D.; Morera, V.; Álvarez, C.; Tejuca, M.; Pazos, F.; García, Y.; Raida, M.; Padrón, G.; Eliana Lanio, M. Identity between cytolysins purified from two morphos of the Caribbean sea anemone Stichodactyla helianthus. Toxicon 2002, 40, 1219. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Clementi, G.; Cunietti, M.; Porres, C.; Oñaderra, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Silent mutations at the 5′-end of the cDNA of actinoporins from the sea anemone Stichodactyla helianthus allow their heterologous overproduction in Escherichia coli. J. Biotechnol. 2007, 127, 211–221. [Google Scholar] [CrossRef]
- Wiseman, T.S.; Williston, J.F.; Brandts, J.F.; Lin, L.N. Rapid measurements of binding constants and heats of binding using a new titration calorimeter. Anal. Chem. 1989, 179, 131–137. [Google Scholar] [CrossRef]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A. One single salt bridge explains the different cytolytic activities shown by actinoporins sticholysin I and II from the venom of Stichodactyla helianthus. Arch. Biochem. Biophys. 2017, 636, 79–89. [Google Scholar] [CrossRef]
- Prato, E.; Biandolino, F. Total lipid content and fatty acid composition of commercially important fish species from the Mediterranean, Mar Grande Sea. Food Chem. 2012, 131, 1233–1239. [Google Scholar] [CrossRef]
- Zorec, R.; Tester, M.; Maček, P.; Mason, W.T. Cytotoxicity of equinatoxin II from the sea anemone Actinia equina involves ion channel formation and an increase in intracellular calcium activity. J. Membr. Biol. 1990, 118, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, G.; Pederzolli, C.; Maček, P.; Menestrina, G. Pore formation by the sea anemone cytolysin equinatoxin-II in red blood cells and model lipid membranes. J. Membr. Biol. 1993, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Serra, M.D.; Ferreras, M.; Lanio, M.E.; Menestrina, G. Mechanism of membrane permeabilization by sticholysin I, a cytolysin isolated from the venom of the sea anemone Stichodactyla helianthus. Biochemistry 1996, 35, 14947–14957. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.; Casallanovo, F.; Shida, C.S.; Nogueira, L.V.; Martínez, D.; Tejuca, M.; Pazos, I.F.; Lanio, M.E.; Menestrina, G.; Lissi, E.; et al. Binding of sea anemone pore-forming toxins sticholysins I and II to interfaces-Modulation of conformation and activity, and lipid-protein interaction. Chem. Phys. Lipids 2003, 122, 97–105. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, H.; Yang, W.; Liu, Y.; Liu, W.; Wei, J.; Tu, H.; Xie, X.; Wang, L.; Xu, A. Functional expression and characterization of an acidic actinoporin from sea anemone Sagartia rosea. Biochem. Biophys. Res. Commun. 2003, 312, 562–570. [Google Scholar] [CrossRef] [PubMed]
- García-Linares, S.; Maula, T.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A. Role of the tryptophan residues in the specific interaction of the sea anemone Stichodactyla helianthus’s actinoporin Sticholysin II with biological membranes. Biochemistry 2016, 55, 6406–6420. [Google Scholar] [CrossRef]
- Ahumada, M.; Calderon, C.; Lissi, E.; Alvarez, C.; Lanio, M.E.; Pazos, F. The pore forming capacity of Sticholysin I in dipalmitoyl phosphatidyl vesicles is tuned by osmotic stress. Chem. Phys. Lipids 2017, 203, 87–93. [Google Scholar] [CrossRef]
- Wimley, W.C. Determining the Effects of Membrane-Interacting Peptides on Membrane Integrity. Methods Mol. Biol. 2015, 1324, 89–106. [Google Scholar]
- Shin, M.L.; Michaels, D.W.; Mayer, M.M. Membrane damage by a toxin from the sea anemone Stoichactis helianthus. II. Effect of membrane lipid composition in a liposome system. Biochim. Biophys. Acta 1979, 555, 79–88. [Google Scholar] [CrossRef]
- Kanazawa, A. Sterols in marine invertebrates. Fish. Sci. 2001, 67, 997–1007. [Google Scholar] [CrossRef]
- Tziakas, D.N.; Chalikias, G.K.; Boudoulas, H. Significance of the cholesterol content of erythrocyte membranes in atherosclerosis. Clin. Lipidol. 2010, 5, 449–452. [Google Scholar] [CrossRef]
- Sepcic, K.; Berne, S.; Potrich, C.; Turk, T.; Maček, P.; Menestrina, G. Interaction of ostreolysin, a cytolytic protein from the edible mushroom Pleurotus ostreatus, with lipid membranes and modulation by lysophospholipids. Eur. J. Biochem. 2003, 270, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Sepcic, K.; Berne, S.; Rebolj, K.; Batista, U.; Plemenitas, A.; Sentjurc, M.; Maček, P. Ostreolysin, a pore-forming protein from the oyster mushroom, interacts specifically with membrane cholesterol-rich lipid domains. FEBS Lett. 2004, 575, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Buboltz, J.T.; Feigenson, G.W. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochim. Biophys. Acta 1999, 1417, 89–100. [Google Scholar] [CrossRef]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef]
- Palacios-Ortega, J. Interaction of Sticholysins with Model Bilayers. Effects of Lipid Composition and Phase State on Pore Formation; Trabajo de Fin de Grado; Abo Akademi University: Turku, Finland; Complutense University of Madrid: Madrid, Spain, 2016. [Google Scholar]
- Yasuda, T.; Al Sazzad, A.; Jäntti, N.Z.; Pentikäinen, O.T.; Slotte, J.P. The Influence of Hydrogen Bonding on Sphingomyelin/Colipid Interactions in Bilayer Membranes. Biophys. J. 2016, 110, 431–440. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Miranda-Zaragoza, B.; Rodríguez-Almazan, C. Actinoporins: From the Structure and Function to the Generation of Biotechnological and Therapeutic Tools. Biomolecules 2020, 10, 539. [Google Scholar] [CrossRef]
- Tejuca, M.; Díaz, I.; Figueredo, R.; Roque, L.; Pazos, F.; Martínez, D.; Iznaga-Escobar, N.; Perez, R.; Álvarez, C.; Lanio, M.E. Construction of an immunotoxin with the pore forming protein StI and ior C5, a monoclonal antibody against a colon cancer cell line. Int. Immunopharmacol. 2004, 4, 731–744. [Google Scholar] [CrossRef]
- Potrich, C.; Tomazzolli, R.; Dalla Serra, M.; Anderluh, G.; Malovrh, P.; Maček, P.; Menestrina, G.; Tejuca, M. Cytotoxic activity of a tumor protease-activated pore-forming toxin. Bioconjug. Chem. 2005, 16, 369–376. [Google Scholar] [CrossRef]
- Tejuca, M.; Anderluh, G.; Dalla Serra, M. Sea anemone cytolysins as toxic components of immunotoxins. Toxicon 2009, 54, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Zhang, J.; Xu, R.; Dong, Y.; Sun, A.; Shen, Y.; Wei, D. Gigantoxin-4-4D5 scFv is a novel recombinant immunotoxin with specific toxicity against HER2/neu-positive ovarian carcinoma cells. Appl. Microbiol. Biotechnol. 2016, 100, 6403–6413. [Google Scholar] [CrossRef] [PubMed]
- Mutter, N.L.; Soskine, M.; Huang, G.; Albuquerque, I.S.; Bernardes, G.J.L.; Maglia, G. Modular Pore-Forming Immunotoxins with Caged Cytotoxicity Tailored by Directed Evolution. ACS Chem. Biol. 2018, 13, 3153–3160. [Google Scholar] [CrossRef] [PubMed]
- Wloka, C.; Mutter, N.L.; Soskine, M.; Maglia, G. a-Helical Fragaceatoxin C Nanopore Engineered for Double-Stranded and Single-Stranded Nucleic Acid Analysis. Angew. Chem. Int. Ed. Engl. 2016, 55, 12494–12498. [Google Scholar] [CrossRef] [PubMed]
- Soletti, R.C.; de Faria, G.P.; Vernal, J.; Terenzi, H.; Anderluh, G.; Borges, H.L.; Moura-Neto, V.; Gabilan, N.H. Potentiation of anticancer-drug cytotoxicity by sea anemone pore-forming proteins in human glioblastoma cells. Anticancer Drugs 2008, 19, 517–525. [Google Scholar] [CrossRef]
- Bakrač, B.; Kladnik, A.; Maček, P.; McHaffie, G.; Werner, A.; Lakey, J.H.; Anderluh, G. A toxin-based probe reveals cytoplasmic exposure of golgi sphingomyelin. J. Biol. Chem. 2010, 285, 22186–22195. [Google Scholar] [CrossRef]
- Skocaj, M.; Bakrač, B.; Krizaj, I.; Maček, P.; Anderluh, G.; Sepcic, K. The sensing of membrane microdomains based on pore-forming toxins. Curr. Med. Chem. 2012, 20, 491–501. [Google Scholar]
- Makino, A.; Abe, M.; Murate, M.; Inaba, T.; Yilmaz, N.; Hullin-Matsuda, F.; Kishimoto, T.; Schieber, N.L.; Taguchi, T.; Arai, H.; et al. Visualization of the heterogeneous membrane distribution of sphingomyelin associated with cytokinesis, cell polarity, and sphingolipidosis. FASEB J. 2015, 29, 477–493. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-de-Torre, E.; Palacios-Ortega, J.; Slotte, J.P.; Gavilanes, J.G.; Martínez-del-Pozo, Á.; García-Linares, S. Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus. Int. J. Mol. Sci. 2020, 21, 8915. https://doi.org/10.3390/ijms21238915
Rivera-de-Torre E, Palacios-Ortega J, Slotte JP, Gavilanes JG, Martínez-del-Pozo Á, García-Linares S. Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus. International Journal of Molecular Sciences. 2020; 21(23):8915. https://doi.org/10.3390/ijms21238915
Chicago/Turabian StyleRivera-de-Torre, Esperanza, Juan Palacios-Ortega, J. Peter Slotte, José G. Gavilanes, Álvaro Martínez-del-Pozo, and Sara García-Linares. 2020. "Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus" International Journal of Molecular Sciences 21, no. 23: 8915. https://doi.org/10.3390/ijms21238915
APA StyleRivera-de-Torre, E., Palacios-Ortega, J., Slotte, J. P., Gavilanes, J. G., Martínez-del-Pozo, Á., & García-Linares, S. (2020). Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus. International Journal of Molecular Sciences, 21(23), 8915. https://doi.org/10.3390/ijms21238915