A Lipidomic Signature Complements Stemness Features Acquisition in Liver Cancer Cells

 ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
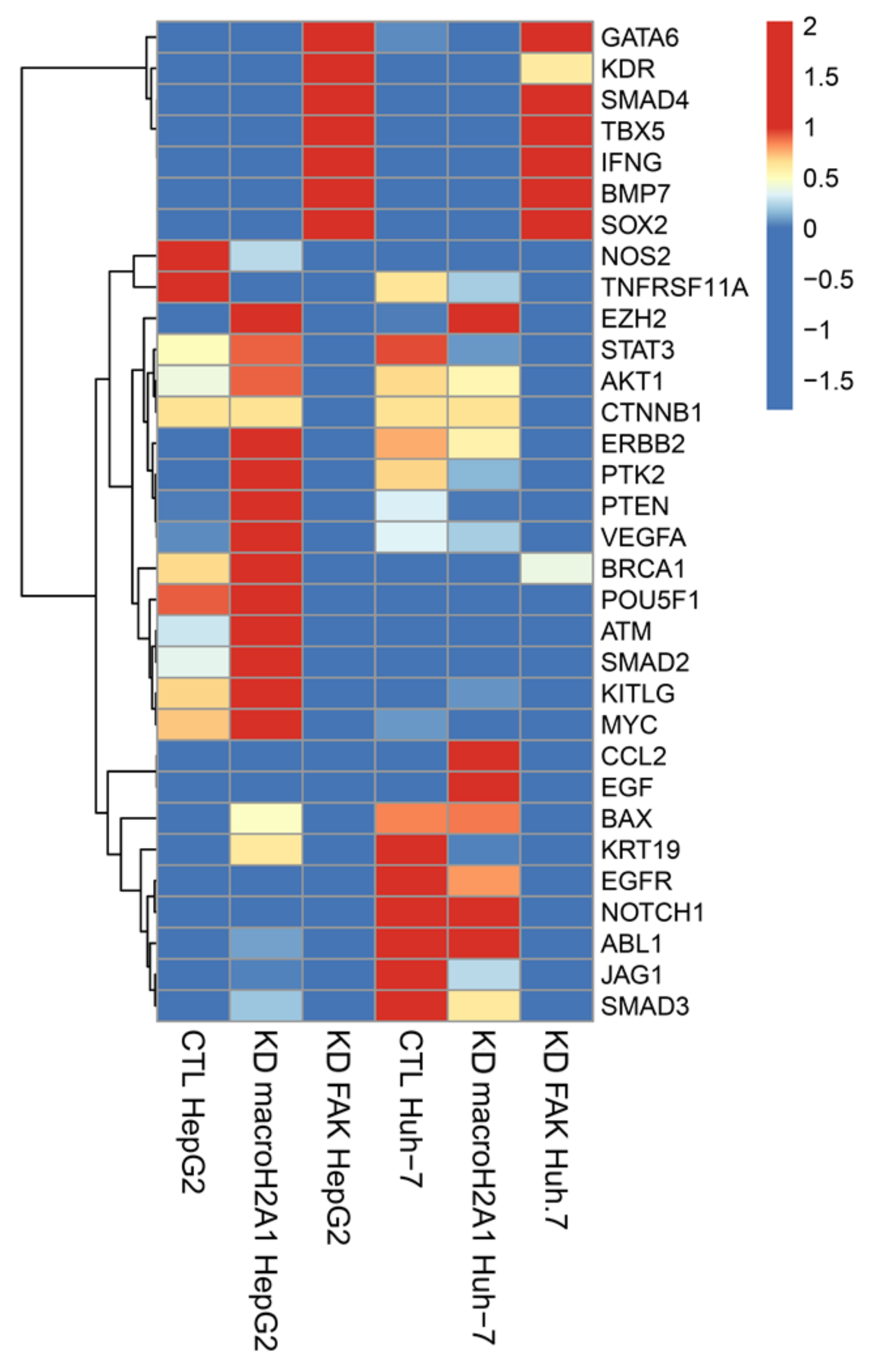
2.1. Transcriptomic Changes in HepG2 and Huh-7 Cells, upon Knockdown (KD) of macroH2A1 (CSC-like) or KD of FAK (CSC Inhibition)
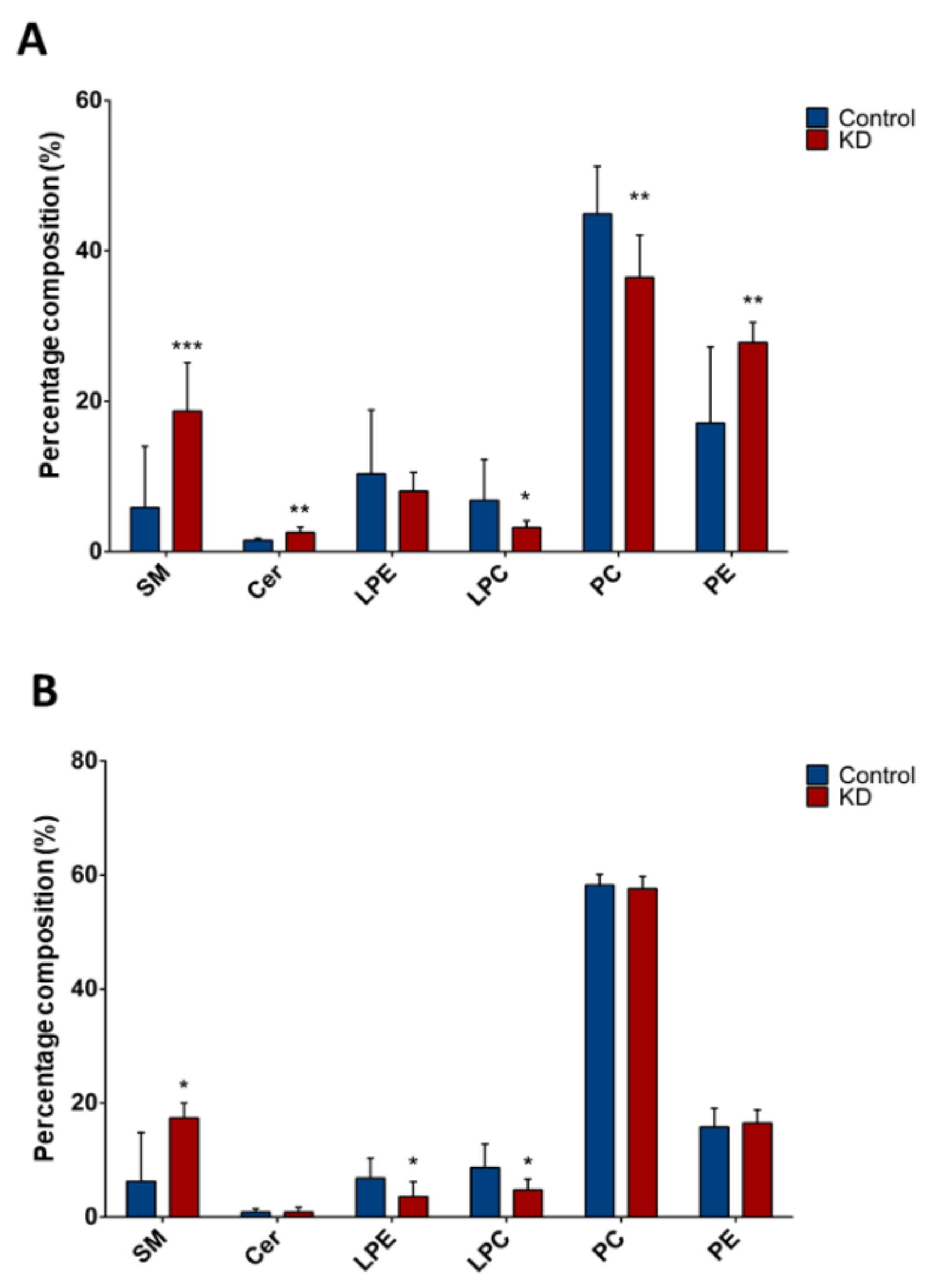
2.2. Lipid Class Composition in Huh-7 and HepG2 Cells Depleted for macroH2A1 or FAK
2.3. Changes in Specific Lipid Species in Huh-7 and HepG2 Cells Depleted for macroH2A1 or FAK
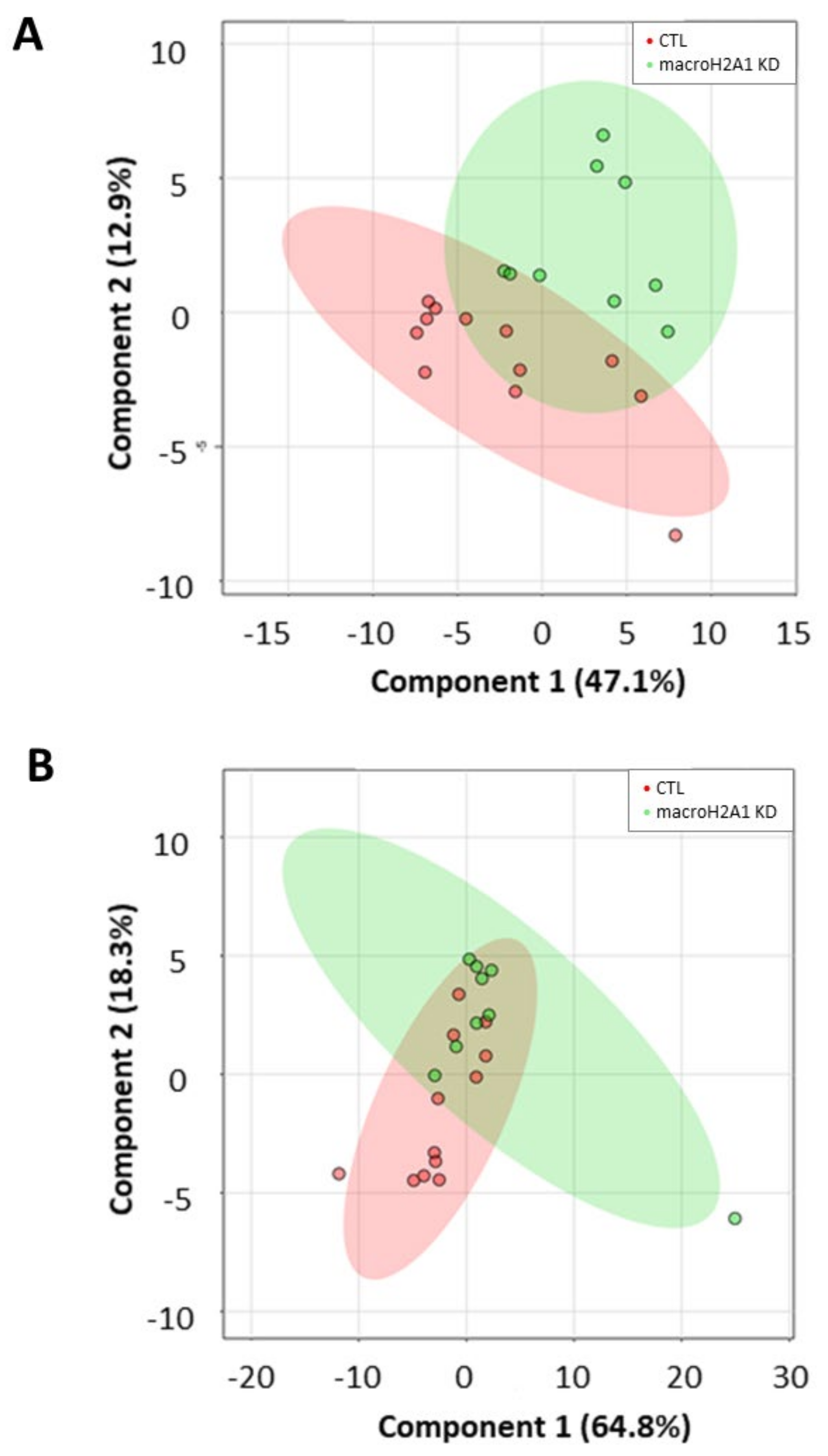
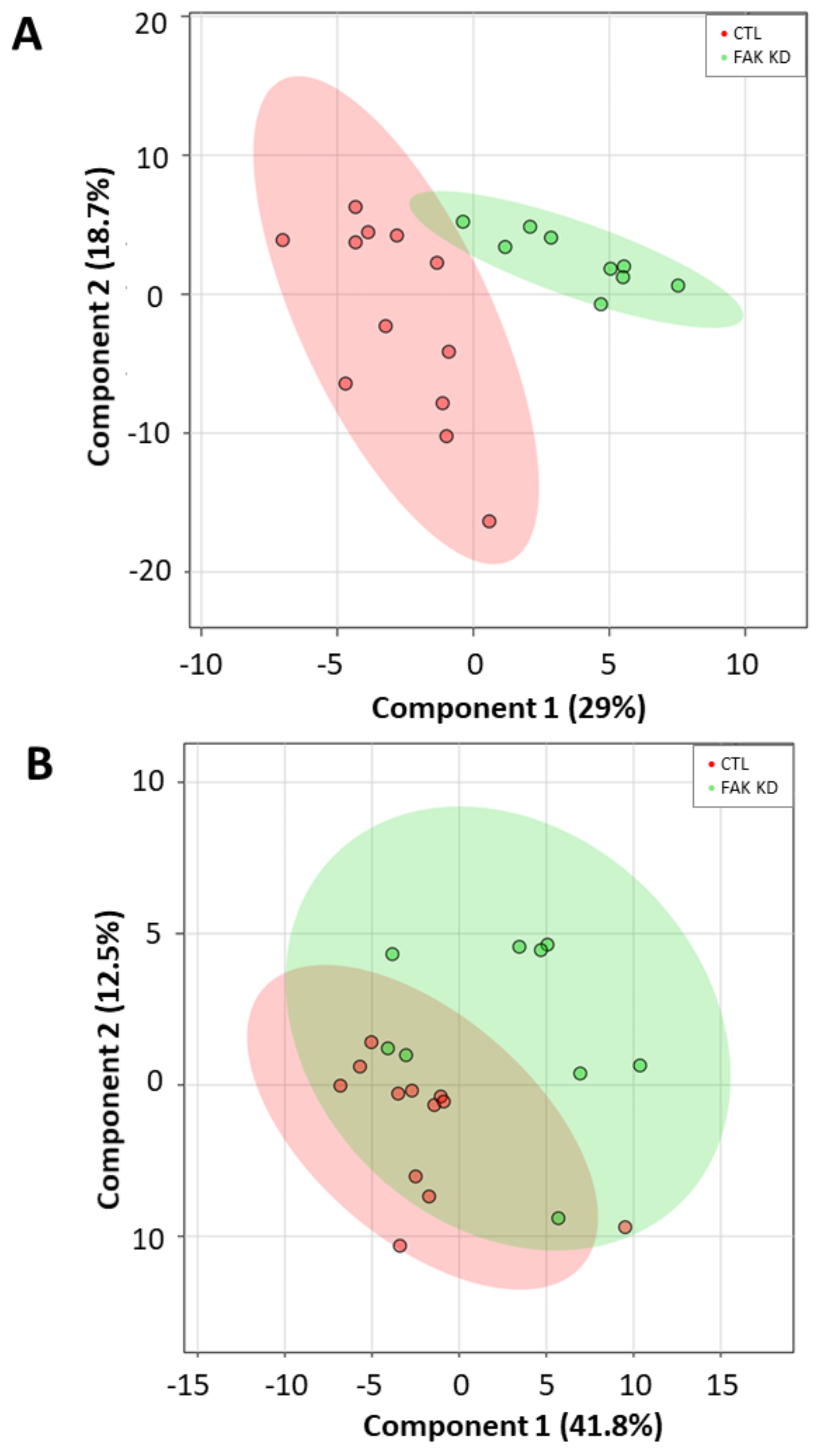
2.4. Lipid Marker Signature Discriminates Huh-7 Cells Depleted for macroH2A1 or FAK
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Extraction of Lipids from HCC Cell Pellets
4.3. Liquid Chromatography/Mass Spectrometry (LC/MS) Analysis
4.4. Transcriptomic Analyses
4.5. Open Array
4.6. Lipid Quantification Assays
4.7. Bioinformatic Analyses
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gores, G.J. Decade in review-hepatocellular carcinoma: HCC-subtypes, stratification and sorafenib. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 645–647. [Google Scholar] [CrossRef]
- Oikawa, T. Cancer Stem cells and their cellular origins in primary liver and biliary tract cancers. Hepatology 2016, 64, 645–651. [Google Scholar] [CrossRef]
- Mazzoccoli, G.; Miele, L.; Oben, J.; Grieco, A.; Vinciguerra, M. Biology, Epidemiology, Clinical Aspects of Hepatocellular Carcinoma and the Role of Sorafenib. Curr. Drug Targets 2016, 17, 783–799. [Google Scholar]
- Yamashita, T.; Wang, X.W. Cancer stem cells in the development of liver cancer. J. Clin. Investig. 2013, 123, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Nio, K.; Yamashita, T.; Kaneko, S. The evolving concept of liver cancer stem cells. Mol. Cancer 2017, 16, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guan, M.; Lin, Y.; Cui, X.; Zhang, Y.; Zhao, Z.; Zhu, J. Aberrant Lipid Metabolism in Hepatocellular Carcinoma Revealed by Liver Lipidomics. Int. J. Mol. Sci. 2017, 18, 2550. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, J.; Huang, C.; Li, N.; Zou, L.; Chia, S.E.; Chen, S.; Yu, K.; Ling, Q.; Cheng, Q.; et al. Comparison of hepatic and serum lipid signatures in hepatocellular carcinoma patients leads to the discovery of diagnostic and prognostic biomarkers. Oncotarget 2018, 9, 5032–5043. [Google Scholar] [CrossRef]
- Muir, K.; Hazim, A.; He, Y.; Peyressatre, M.; Kim, D.Y.; Song, X.; Beretta, L. Proteomic and lipidomic signatures of lipid metabolism in NASH-associated hepatocellular carcinoma. Cancer Res. 2013, 73, 4722–4731. [Google Scholar] [CrossRef]
- Lin, L.; Ding, Y.; Wang, Y.; Wang, Z.; Yin, X.; Yan, G.; Zhang, L.; Yang, P.; Shen, H. Functional lipidomics: Palmitic acid impairs hepatocellular carcinoma development by modulating membrane fluidity and glucose metabolism. Hepatology 2017, 66, 432–448. [Google Scholar] [CrossRef]
- Vinciguerra, M.; Carrozzino, F.; Peyrou, M.; Carlone, S.; Montesano, R.; Benelli, R.; Foti, M. Unsaturated fatty acids promote hepatoma proliferation and progression through downregulation of the tumor suppressor PTEN. J. Hepatol. 2009, 50, 1132–1141. [Google Scholar] [CrossRef]
- Vinciguerra, M.; Sgroi, A.; Veyrat-Durebex, C.; Rubbia-Brandt, L.; Buhler, L.H.; Foti, M. Unsaturated fatty acids inhibit the expression of tumor suppressor phosphatase and tensin homolog (PTEN) via microRNA-21 up-regulation in hepatocytes. Hepatology 2009, 49, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, M.; Veyrat-Durebex, C.; Moukil, M.A.; Rubbia-Brandt, L.; Rohner-Jeanrenaud, F.; Foti, M. PTEN down-regulation by unsaturated fatty acids triggers hepatic steatosis via an NF-kappaBp65/mTOR-dependent mechanism. Gastroenterology 2008, 134, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Christen, S.; Parik, S.; Broekaert, D.; Yoshinaga, K.; Talebi, A.; Dehairs, J.; Escalona-Noguero, C.; Schmieder, R.; Cornfield, T.; et al. Evidence for an alternative fatty acid desaturation pathway increasing cancer plasticity. Nature 2019, 566, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Li, J.; Chen, S.; Cai, J.; Ban, Y.; Peng, Q.; Zhou, Y.; Zeng, Z.; Peng, S.; Li, X.; et al. Emerging role of lipid metabolism alterations in Cancer stem cells. J. Exp. Clin. Cancer Res. 2018, 37, 118. [Google Scholar] [CrossRef]
- Mancini, R.; Noto, A.; Pisanu, M.E.; De Vitis, C.; Maugeri-Sacca, M.; Ciliberto, G. Metabolic features of cancer stem cells: The emerging role of lipid metabolism. Oncogene 2018, 37, 2367–2378. [Google Scholar] [CrossRef]
- Tirinato, L.; Pagliari, F.; Limongi, T.; Marini, M.; Falqui, A.; Seco, J.; Candeloro, P.; Liberale, C.; Di Fabrizio, E. An Overview of Lipid Droplets in Cancer and Cancer Stem Cells. Stem Cells Int. 2017, 2017, 1656053. [Google Scholar] [CrossRef]
- Lo Re, O.; Douet, J.; Buschbeck, M.; Fusilli, C.; Pazienza, V.; Panebianco, C.; Castracani, C.C.; Mazza, T.; Li Volti, G.; Vinciguerra, M. Histone variant macroH2A1 rewires carbohydrate and lipid metabolism of hepatocellular carcinoma cells towards cancer stem cells. Epigenetics 2018, 13, 829–845. [Google Scholar] [CrossRef]
- Lo Re, O.; Fusilli, C.; Rappa, F.; Van Haele, M.; Douet, J.; Pindjakova, J.; Rocha, S.W.; Pata, I.; Valcikova, B.; Uldrijan, S.; et al. Induction of cancer cell stemness by depletion of macrohistone H2A1 in hepatocellular carcinoma. Hepatology 2018, 67, 636–650. [Google Scholar] [CrossRef]
- Lo Re, O.; Mazza, T.; Giallongo, S.; Sanna, P.; Rappa, F.; Vinh Luong, T.; Li Volti, G.; Drovakova, A.; Roskams, T.; Van Haele, M.; et al. Loss of histone macroH2A1 in hepatocellular carcinoma cells promotes paracrine-mediated chemoresistance and CD4(+)CD25(+)FoxP3(+) regulatory T cells activation. Theranostics 2020, 10, 910–924. [Google Scholar] [CrossRef]
- Visweswaran, M.; Arfuso, F.; Warrier, S.; Dharmarajan, A. Aberrant lipid metabolism as an emerging therapeutic strategy to target cancer stem cells. Stem Cells 2020, 38, 6–14. [Google Scholar] [CrossRef]
- Gnani, D.; Romito, I.; Artuso, S.; Chierici, M.; De Stefanis, C.; Panera, N.; Crudele, A.; Ceccarelli, S.; Carcarino, E.; D’Oria, V.; et al. Focal adhesion kinase depletion reduces human hepatocellular carcinoma growth by repressing enhancer of zeste homolog 2. Cell Death Differ. 2017, 24, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Duan, J.; Wang, L.; Xiao, S.; Li, L.; Yan, X.; Yao, W.; Wu, L.; Zhang, S.; Zhang, Y.; et al. PTK2 promotes cancer stem cell traits in hepatocellular carcinoma by activating Wnt/beta-catenin signaling. Cancer Lett. 2019, 450, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Condello, S.; Thomes-Pepin, J.; Ma, X.; Xia, Y.; Hurley, T.D.; Matei, D.; Cheng, J.X. Lipid Desaturation Is a Metabolic Marker and Therapeutic Target of Ovarian Cancer Stem Cells. Cell Stem Cell 2017, 20, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y. Therapeutic targeting of lipid synthesis metabolism for selective elimination of cancer stem cells. Arch. Pharmacal Res. 2019, 42, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.K.Y.; Kweon, S.M.; Chi, F.; Hwang, E.; Kabe, Y.; Higashiyama, R.; Qin, L.; Yan, R.; Wu, R.P.; Lai, K.; et al. Stearoyl-CoA Desaturase Promotes Liver Fibrosis and Tumor Development in Mice via a Wnt Positive-Signaling Loop by Stabilization of Low-Density Lipoprotein-Receptor-Related Proteins 5 and 6. Gastroenterology 2017, 152, 1477–1491. [Google Scholar] [CrossRef]
- Morita, Y.; Sakaguchi, T.; Ikegami, K.; Goto-Inoue, N.; Hayasaka, T.; Hang, V.T.; Tanaka, H.; Harada, T.; Shibasaki, Y.; Suzuki, A.; et al. Lysophosphatidylcholine acyltransferase 1 altered phospholipid composition and regulated hepatoma progression. J. Hepatol. 2013, 59, 292–299. [Google Scholar] [CrossRef]
- Skill, N.J.; Scott, R.E.; Wu, J.; Maluccio, M.A. Hepatocellular carcinoma associated lipid metabolism reprogramming. J. Surg. Res. 2011, 169, 51–56. [Google Scholar] [CrossRef]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Martinez, L.; Torres, S.; Baulies, A.; Alarcon-Vila, C.; Elena, M.; Fabrias, G.; Casas, J.; Caballeria, J.; Fernandez-Checa, J.C.; Garcia-Ruiz, C. Myristic acid potentiates palmitic acid-induced lipotoxicity and steatohepatitis associated with lipodystrophy by sustaning de novo ceramide synthesis. Oncotarget 2015, 6, 41479–41496. [Google Scholar] [CrossRef]
- Cotte, A.K.; Cottet, V.; Aires, V.; Mouillot, T.; Rizk, M.; Vinault, S.; Binquet, C.; de Barros, J.P.; Hillon, P.; Delmas, D. Phospholipid profiles and hepatocellular carcinoma risk and prognosis in cirrhotic patients. Oncotarget 2019, 10, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Krautbauer, S.; Meier, E.M.; Rein-Fischboeck, L.; Pohl, R.; Weiss, T.S.; Sigruener, A.; Aslanidis, C.; Liebisch, G.; Buechler, C. Ceramide and polyunsaturated phospholipids are strongly reduced in human hepatocellular carcinoma. Biochim. Biophys. Acta 2016, 1861, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Kimhofer, T.; Fye, H.; Taylor-Robinson, S.; Thursz, M.; Holmes, E. Proteomic and metabonomic biomarkers for hepatocellular carcinoma: A comprehensive review. Br. J. Cancer 2015, 112, 1141–1156. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, B.; Bhardwaj, G.; Lau, J.Y. Expression of p21 and p27 in hepatoma cell lines with different p53 gene profile. J. Hepatol. 1999, 31, 386. [Google Scholar] [CrossRef]
- Shiraha, H.; Yamamoto, K.; Namba, M. Human hepatocyte carcinogenesis (review). Int. J. Oncol. 2013, 42, 1133–1138. [Google Scholar] [CrossRef]
- Shetzer, Y.; Molchadsky, A.; Rotter, V. Oncogenic Mutant p53 Gain of Function Nourishes the Vicious Cycle of Tumor Development and Cancer Stem-Cell Formation. Cold Spring Harb. Perspect. Med. 2016, 6. [Google Scholar] [CrossRef]
- Ricchi, M.; Odoardi, M.R.; Carulli, L.; Anzivino, C.; Ballestri, S.; Pinetti, A.; Fantoni, L.I.; Marra, F.; Bertolotti, M.; Banni, S.; et al. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J. Gastroenterol. Hepatol. 2009, 24, 830–840. [Google Scholar] [CrossRef]
- Hishikawa, D.; Valentine, W.J.; Iizuka-Hishikawa, Y.; Shindou, H.; Shimizu, T. Metabolism and functions of docosahexaenoic acid-containing membrane glycerophospholipids. FEBS Lett. 2017, 591, 2730–2744. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Szymańska, E.; Saccenti, E.; Smilde, A.K.; Westerhuis, J.A. Double-check: Validation of diagnostic statistics for PLS-DA models in metabolomics studies. Metabolomics 2012, 8, 3–16. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Species | Fold Change | p-Value |
|---|---|---|
| Huh-7 Cells Depleted for macroH2A1 | ||
| LPC (14:0) | 2.3 | 0.013 |
| LPC (22:5) | 3.5 | 0.018 |
| PC (16:0/20:5) | 3.0 | 0.018 |
| PC (18:0/20:5) | 2.7 | 0.023 |
| LPC (20:5) | 4.4 | 0.036 |
| LPC (22:4) | 2.8 | 0.058 |
| Cer (16:0) | 0.4 | 0.069 |
| LPE (20:0) | 0.4 | 0.070 |
| LPC (22:6) | 3.2 | 0.076 |
| HepG2 Cells Depleted for macroH2A1 | ||
| LPC (22:5) | 6.6 | 0.003 |
| PC (18:2/22:5) | 4.6 | 0.005 |
| LPC (20:3) | 4.2 | 0.009 |
| PC (18:2/20:5) | 5.9 | 0.010 |
| LPC (18:3) | 5.3 | 0.011 |
| LPC (22:4) | 4.9 | 0.020 |
| LPC (20:5) | 3.6 | 0.021 |
| PC (18:1/20:5) | 5.4 | 0.025 |
| PC (18:1/22:5) | 4.4 | 0.025 |
| PC (18:1/22:4) | 3.4 | 0.027 |
| PC (18:2/22:6) | 4.1 | 0.030 |
| LPC (20:1) | 3.1 | 0.036 |
| LPC (20:2) | 3.3 | 0.040 |
| LPC (18:2) | 3.6 | 0.052 |
| PC (18:0/20:5) | 3.0 | 0.060 |
| LPC (14:0) | 2.8 | 0.065 |
| LPC (18:1) | 3.2 | 0.066 |
| PC (16:0/20:5) | 3.0 | 0.067 |
| LPC (20:4) | 4.1 | 0.068 |
| LPC (16:1) | 2.9 | 0.071 |
| PE (18:1/22:4) | 2.6 | 0.074 |
| PC (16:0/22:4) | 2.1 | 0.082 |
| PC (16:1/18:2) | 2.6 | 0.084 |
| LPE (22:4) | 2.6 | 0.092 |
| Lipid Species | Fold Change | p-Value |
|---|---|---|
| Huh-7 Cells Depleted for FAK | ||
| LPC (16:0) | 2.5 | <0.001 |
| LPC (14:0) | 3.5 | <0.001 |
| LPE (14:0) | 2.2 | 0.002 |
| LPE (20:3) | 0.4 | 0.006 |
| PE (18:0/20:3) | 0.5 | 0.015 |
| LPC (22:0) | 2.4 | 0.018 |
| PE (18:0/18:1) | 0.4 | 0.026 |
| LPC (16:1) | 2.1 | 0.063 |
| Cer (16:0) | 0.2 | 0.065 |
| LPC (20:5) | 2.0 | 0.070 |
| PE (18:1/22:4) | 2.1 | 0.082 |
| LPC (20:1) | 2.4 | 0.094 |
| HepG2 Cells Depleted for FAK | ||
| Cer (24:1) | 2.7 | 0.070 |
| Cer (24:0) | 2.2 | 0.073 |
| PE (18:0/20:3) | 0.5 | 0.080 |
| LPE (20:0) | 0.5 | 0.084 |
| SM (18:0) | 2.0 | 0.087 |
| PE (16:0/20:3) | 0.5 | 0.087 |
| PE (16:0/14:0) | 0.4 | 0.093 |
| SM (16:0) | 2.0 | 0.095 |
| PC/PE | p-Value | PC/LPC | p-Value | UFA | p-Value | |
|---|---|---|---|---|---|---|
| CTL Huh-7 | 4 ± 0.30 | - | 8 ± 1.43 | - | 158 ± 12 | - |
| CTL HepG2 | 2 ± 0.30 | - | 11 ± 2.71 | - | 174 ± 19 | - |
| macroH2A1 KD Huh-7 | 3 ± 0.20 | 0.01 | 13 ± 2.03 | 0.03 | 231 ± 21 | 0.01 |
| macroH2A1 KD HepG2 | 1 ± 0.11 | 0.01 | 15 ± 3.47 | 0.02 | 194 ± 10 | 0.09 |
| FAK KD Huh-7 | 2.5 ± 0.23 | 0.001 | 6.5 ± 0.83 | 0.6 | 161 ± 36 | - |
| FAK KD HepG2 | 2 ± 0.16 | 0.21 | 7 ± 0.74 | 0.06 | 169 ± 17 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas Serna, I.M.; Romito, I.; Maugeri, A.; Lo Re, O.; Giallongo, S.; Mazzoccoli, G.; Oben, J.A.; Li Volti, G.; Mazza, T.; Alisi, A.; et al. A Lipidomic Signature Complements Stemness Features Acquisition in Liver Cancer Cells. Int. J. Mol. Sci. 2020, 21, 8452. https://doi.org/10.3390/ijms21228452
Rivas Serna IM, Romito I, Maugeri A, Lo Re O, Giallongo S, Mazzoccoli G, Oben JA, Li Volti G, Mazza T, Alisi A, et al. A Lipidomic Signature Complements Stemness Features Acquisition in Liver Cancer Cells. International Journal of Molecular Sciences. 2020; 21(22):8452. https://doi.org/10.3390/ijms21228452
Chicago/Turabian StyleRivas Serna, Irma Magaly, Ilaria Romito, Andrea Maugeri, Oriana Lo Re, Sebastiano Giallongo, Gianluigi Mazzoccoli, Jude A. Oben, Giovanni Li Volti, Tommaso Mazza, Anna Alisi, and et al. 2020. "A Lipidomic Signature Complements Stemness Features Acquisition in Liver Cancer Cells" International Journal of Molecular Sciences 21, no. 22: 8452. https://doi.org/10.3390/ijms21228452
APA StyleRivas Serna, I. M., Romito, I., Maugeri, A., Lo Re, O., Giallongo, S., Mazzoccoli, G., Oben, J. A., Li Volti, G., Mazza, T., Alisi, A., & Vinciguerra, M. (2020). A Lipidomic Signature Complements Stemness Features Acquisition in Liver Cancer Cells. International Journal of Molecular Sciences, 21(22), 8452. https://doi.org/10.3390/ijms21228452