No Role of Osteocytic Osteolysis in the Development and Recovery of the Bone Phenotype Induced by Severe Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Strong Correlations of Serum PTH, ALP, and Serum Calcium Levels with Bone Matrix Mineralization, but Not with Osteocyte Lacunar Size
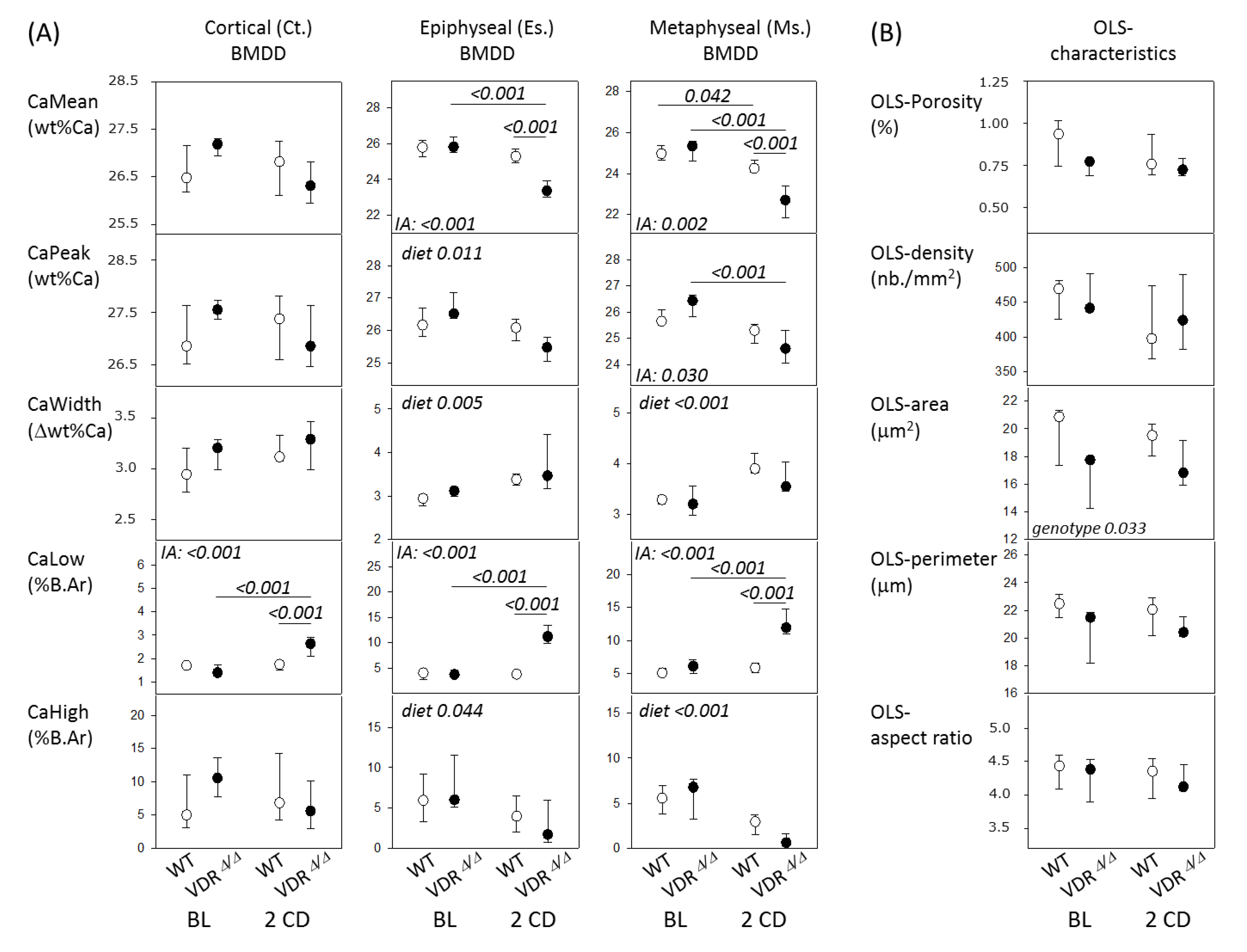
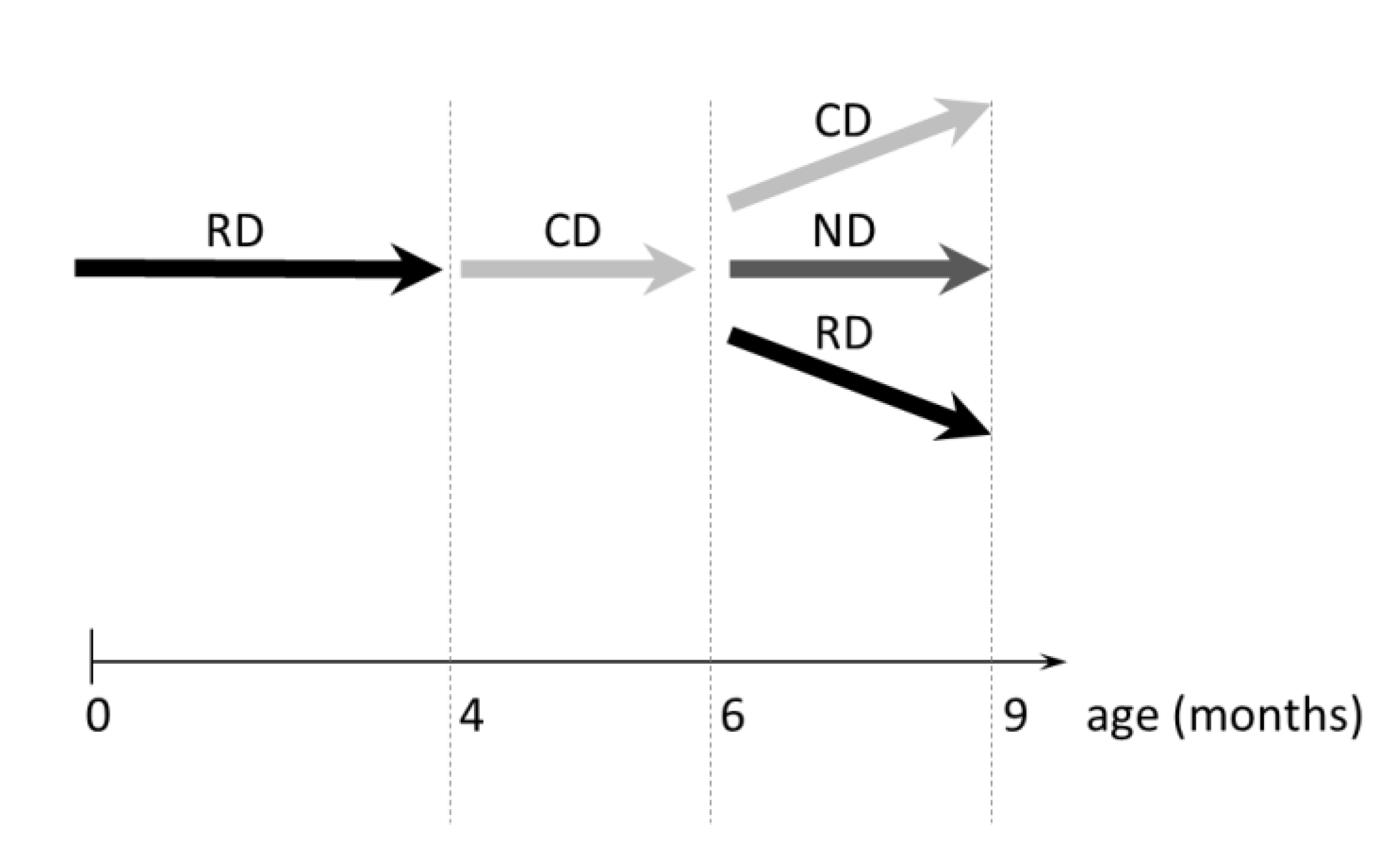
2.2. Bone Phenotype of VDRΔ/Δ Mice Is Similar to That of WT at Baseline but Significantly Different after 2 Months CD
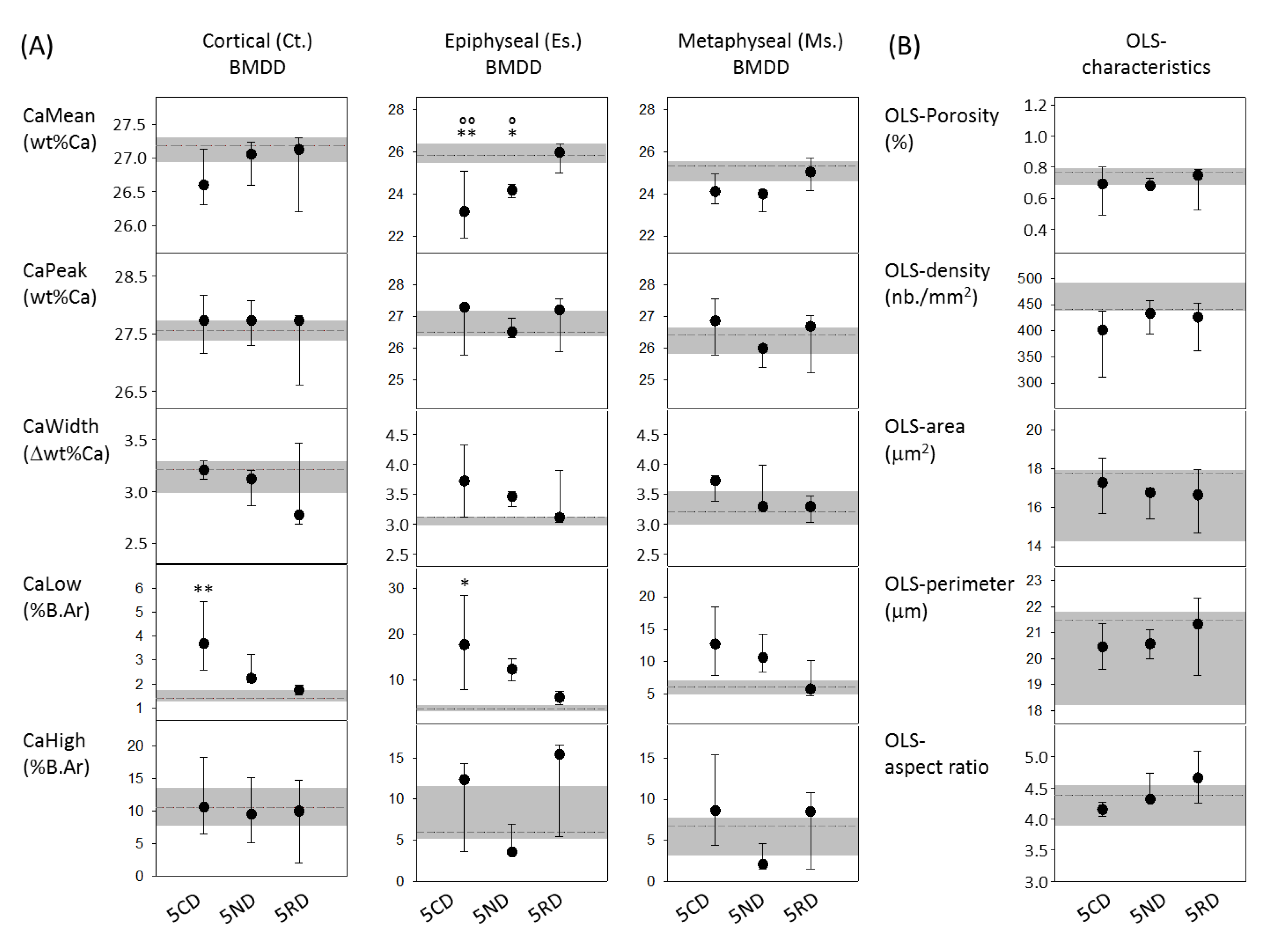
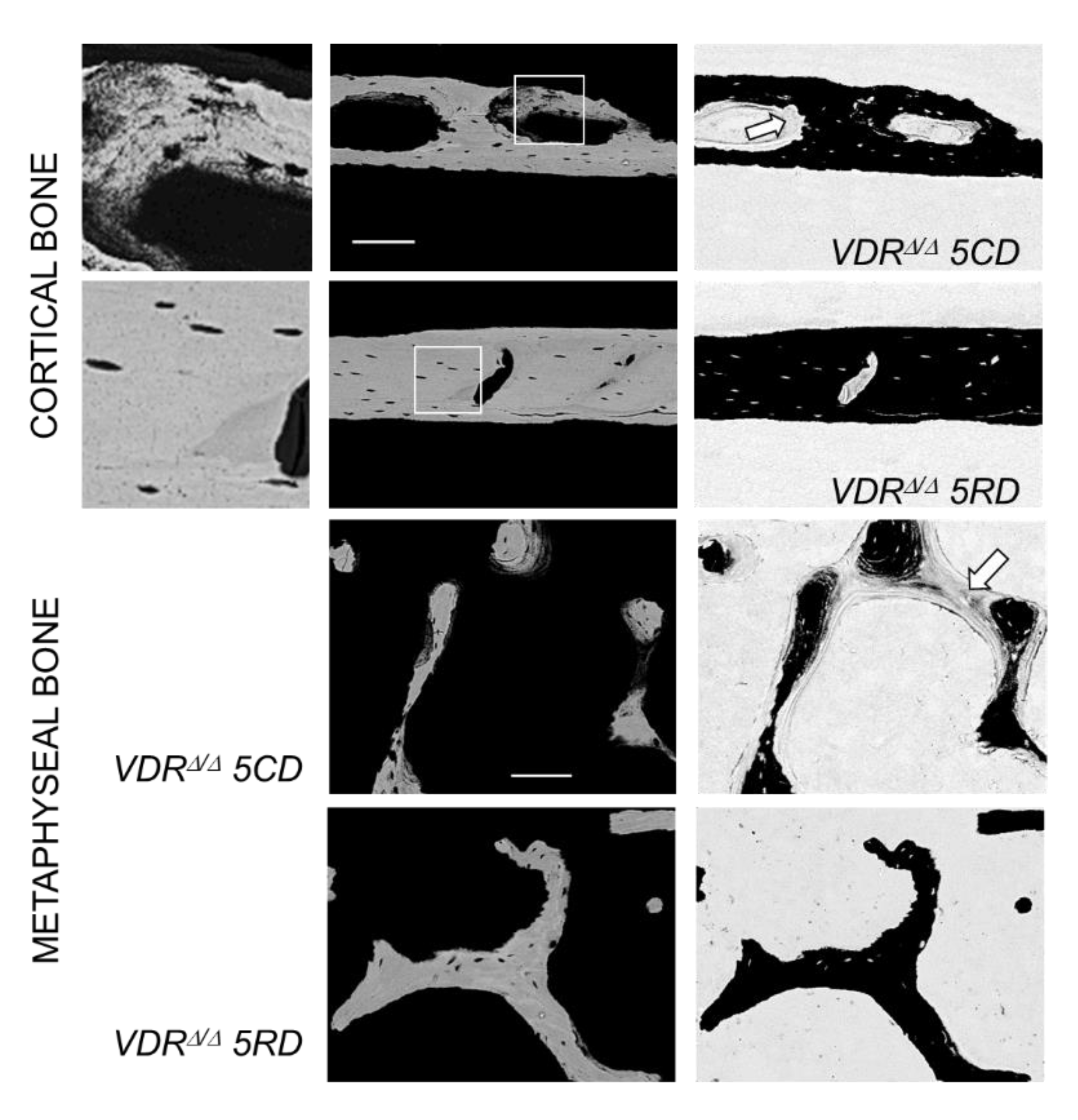
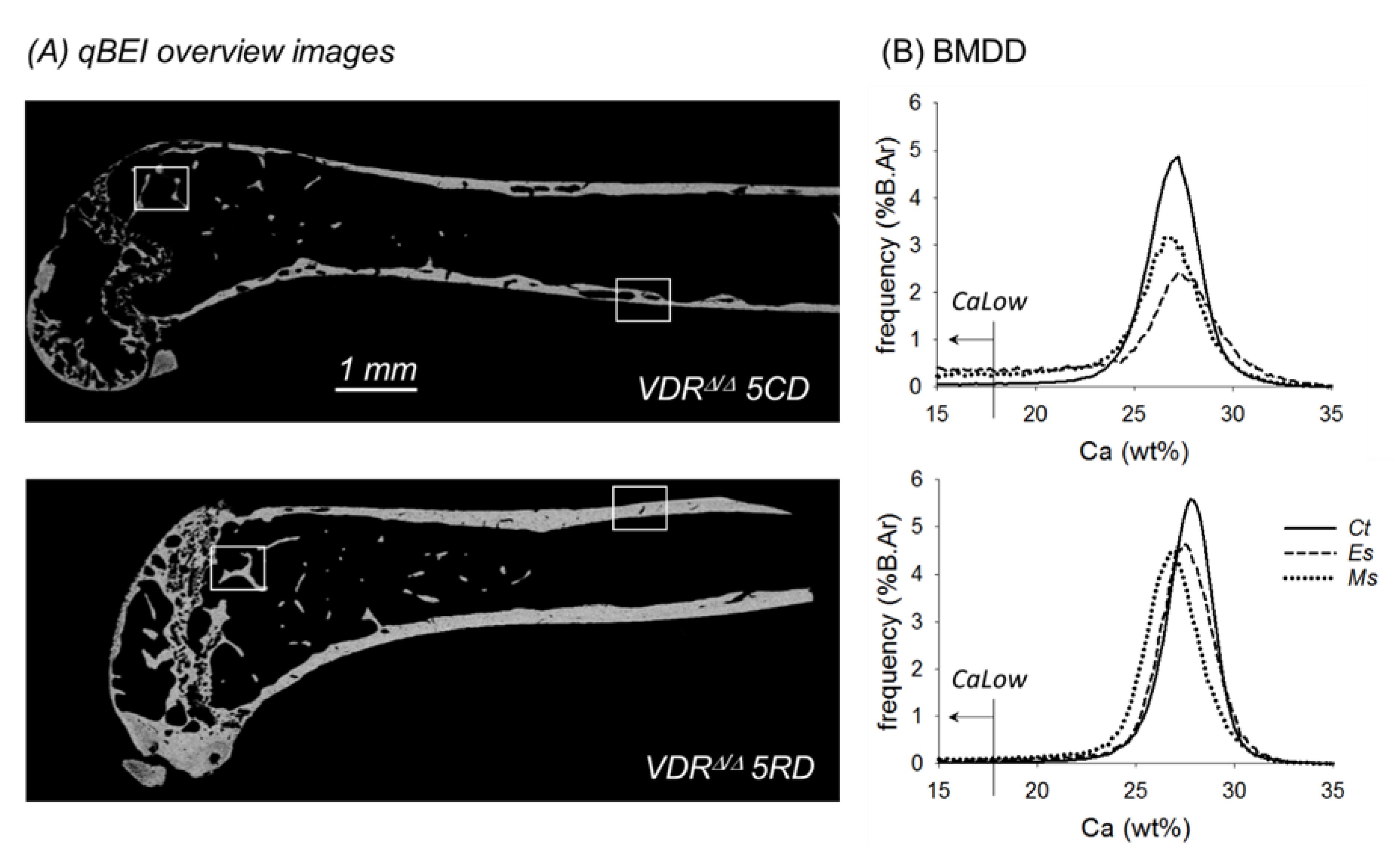
2.3. From the 9-Month-Old VDRΔ/Δ Animals, Only Those on RD Have Similar BMDD Compared to Baseline
3. Discussion
4. Materials and Methods
4.1. Bone Samples
4.2. Quantitative Backscattered Electron Imaging (qBEI)
4.3. Analysis of Structural Morphometric Parameters of Mineralized Bone Tissue
4.4. Osteocyte Lacunae Sections (OLS) Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| VDR | vitamin D receptor |
| PTH | parathyroid hormone |
| PTHrP | parathyroid hormone related protein |
| VDRΔ/Δ | mutants with a non-functioning VDR |
| sHPT | secondary hyperparathyroidism |
| BMDD | bone mineralization density distribution |
| qBEI | quantitative backscattered electron imaging |
| OLS | osteocyte lacunae sections |
| RD | rescue diet |
| CD | challenge diet |
| ND | normal diet |
Appendix A

References
- Lieben, L.; Carmeliet, G.J. The delicate balance between vitamin D, calcium and bone homeostasis: Lessons learned from intestinal- and osteocyte-specific VDR null mice. Steroid Biochem. Mol. Biol. 2013, 136, 102–106. [Google Scholar] [CrossRef]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef]
- Erben, R.G.; Soegiarto, D.W.; Weber, K.; Zeitz, U.; Lieberherr, M.; Gniadecki, R.; Möller, G.; Adamski, J.; Balling, R. Deletion of deoxyribonucleic acid binding domain of the vitamin D receptor abrogates genomic and nongenomic functions of vitamin D. Mol. Endocrinol. 2002, 16, 1524–1537. [Google Scholar] [CrossRef]
- Fleet, J.C. The role of vitamin D in the endocrinology controlling calcium homeostasis. Mol. Cell. Endocrinol. 2017, 453, 36–45. [Google Scholar] [CrossRef]
- Christakos, S.; Li, S.; DeLa Cruz, J.; Verlinden, L.; Carmeliet, G. Vitamin D and Bone. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2019; Available online: https://doi-org.ez.srv.meduniwien.ac.at/10.1007/164_2019_338 (accessed on 10 October 2020).
- Qing, H.; Ardeshirpour, L.; Pajevic, P.D.; Dusevich, V.; Jähn, K.; Kato, S.; Wysolmerski, J.; Bonewald, L.F. Demonstration of osteocytic perilacunar/canalicular remodeling in mice during lactation. J. Bone Miner. Res. 2012, 27, 1018–1029. [Google Scholar] [CrossRef] [Green Version]
- Wysolmerski, J.J. Osteocytic osteolysis: Time for a second look? Bonekey Rep. 2012, 1, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsourdi, E.; Jähn, K.; Rauner, M.; Busse, B.; Bonewald, L.F. Physiological and pathological osteocytic osteolysis. J. Musculoskelet. Neuronal Interact. 2018, 18, 292–303. [Google Scholar] [PubMed]
- Weber, K.; Bergow, C.; Hirmer, S.; Schüler, C.; Erben, R.G. Vitamin D-independent therapeutic effects of extracellular calcium in a mouse model of adult-onset secondary hyperparathyroidism. J. Bone Miner. Res. 2009, 24, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Pirro, A.E.; Amling, M.; Delling, G.; Baron, R.; Bronson, R.; Demay, M.B. Targeted ablation of the vitamin D receptor: An animal model of vitamin D-dependent rickets type II with alopecia. Proc. Natl. Acad. Sci. USA 1997, 94, 9831–9835. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, T.; Handa, Y.; Uematsu, Y.; Takeda, S.; Sekine, K.; Yoshihara, Y.; Kawakami, T.; Arioka, K.; Sato, H.; Uchiyama, Y.; et al. Mice lacking the vitamin D receptor exhibit impaired bone formation, uterine hypoplasia and growth retardation after weaning. Nat. Genet. 1997, 16, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Lieben, L.; Masuyama, R.; Torrekens, S.; Van Looveren, R.; Schrooten, J.; Baatsen, P.; Lafage-Proust, M.H.; Dresselaers, T.; Feng, J.Q.; Bonewald, L.F.; et al. Normocalcemia is maintained in mice under conditions of calcium malabsorption by vitamin D-induced inhibition of bone mineralization. J. Clin. Investig. 2012, 122, 1803–1815. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Amling, M.; Pirro, A.E.; Priemel, M.; Meuse, J.; Baron, R.; Delling, G.; Demay, M.B. Normalization of mineral ion homeostasis by dietary means prevents hyperparathyroidism, rickets, and osteomalacia, but not alopecia in vitamin D receptor-ablated mice. Endocrinology 1998, 139, 4391–4396. [Google Scholar] [CrossRef] [PubMed]
- Rolvien, T.; Krause, M.; Jeschke, A.; Yorgan, T.; Püschel, K.; Schinke, T.; Busse, B.; Demay, M.B.; Amling, M. Vitamin D regulates osteocyte survival and perilacunar remodeling in human and murine bone. Bone 2017, 103, 78–87. [Google Scholar] [CrossRef]
- Lanske, B.; Densmore, M.J.; Erben, R.G. Vitamin D endocrine system and osteocytes. Bonekey Rep. 2014, 3, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misof, B.M.; Blouin, S.; Roschger, P.; Werzowa, J.; Klaushofer, K.; Lehmann, G. Bone matrix mineralization and osteocyte lacunae characteristics in patients with chronic kidney disease—Mineral bone disorder (CKD-MBD). J. Musculoskelet. Neuronal Interact. 2019, 19, 196–206. [Google Scholar]
- Nawrot-Wawrzyniak, K.; Misof, B.M.; Roschger, P.; Pańczyk-Tomaszewska, M.; Ziółkowska, H.; Klaushofer, K.; Fratzl-Zelman, N. Changes in bone matrix mineralization after growth hormone treatment in children and adolescents with chronic kidney failure treated by dialysis: A paired biopsy study. Am. J. Kidney Dis. 2013, 61, 767–777. [Google Scholar] [CrossRef]
- Roschger, P.; Dempster, D.W.; Zhou, H.; Paschalis, E.P.; Silverberg, S.J.; Shane, E.; Bilezikian, J.P.; Klaushofer, K. New observations on bone quality in mild primary hyperparathyroidism as determined by quantitative backscattered electron imaging. J. Bone Miner. Res. 2007, 22, 717–723. [Google Scholar] [CrossRef]
- Ryan, J.W.; Starczak, Y.; Tsangari, H.; Sawyer, R.K.; Davey, R.A.; Atkins, G.J.; Morris, H.A.; Anderson, P.H. Sex-related differences in the skeletal phenotype of aged vitamin D receptor global knockout mice. J. Steroid Biochem. Mol. Biol. 2016, 164, 361–368. [Google Scholar] [CrossRef]
- Streicher, C.; Zeitz, U.; Andrukhova, O.; Rupprecht, A.; Pohl, E.; Larsson, T.E.; Windisch, W.; Lanske, B.; Erben, R.G. Long-term Fgf23 deficiency does not influence aging, glucose homeostasis, or fat metabolism in mice with a nonfunctioning vitamin D receptor. Endocrinology 2012, 153, 1795–1805. [Google Scholar] [CrossRef]
- Ng, A.H.; Omelon, S.; Variola, F.; Allo, B.; Willett, T.L.; Alman, B.A.; Grynpas, M.D. Adynamic Bone Decreases Bone Toughness During Aging by Affecting Mineral and Matrix. J. Bone Miner. Res. 2016, 31, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Hofstaetter, J.G.; Misof, B.M.; Jones, D.C.; Zoehrer, R.; Blouin, S.; Schueler, C.; Paschalis, E.P.; Erben, R.G.; Weinkamer, R.; Klaushofer, K.; et al. Biomechanical and bone material properties of Schnurri-3 null mice. JBMR Plus 2019, 3, e10226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heveran, C.M.; Rauff, A.; King, K.B.; Carpenter, R.D.; Ferguson, V.L. A new open-source tool for measuring 3D osteocyte lacunar geometries from confocal laser scanning microscopy reveals age-related changes to lacunar size and shape in cortical mouse bone. Bone 2018, 110, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, K.; Hoshi, K.; Kawamoto, S.; Tanaka, M.; Ejiri, S.; Ozawa, H. Osteocytic osteolysis observed in rats to which parathyroid hormone was continuously administered. J. Bone Miner. Metab. 2004, 22, 524–529. [Google Scholar] [CrossRef]
- Fratzl-Zelman, N.; Gamsjaeger, S.; Blouin, S.; Kocijan, R.; Plasenzotti, P.; Rokidi, S.; Nawrot-Wawrzyniak, K.; Roetzer, K.; Uyanik, G.; Haeusler, G.; et al. Alterations of bone material properties in adult patients with X-linked hypophosphatemia (XLH). J. Struct. Biol. 2020, 211, 107556. [Google Scholar] [CrossRef] [PubMed]
- Nango, N.; Kubota, S.; Hasegawa, T.; Yashiro, W.; Momose, A.; Matsuo, K. Osteocyte-directed bone demineralization along canaliculi. Bone 2016, 84, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschger, P.; Fratzl, P.; Eschberger, J.; Klaushofer, K. Validation of quantitative backscattered electron imaging for the measurement of mineral density distribution in human bone biopsies. Bone 1998, 23, 319–326. [Google Scholar] [CrossRef]
- Roschger, P.; Paschalis, E.P.; Fratzl, P.; Klaushofer, K. Bone mineralization density distribution in health and disease. Bone 2008, 42, 456–466. [Google Scholar] [CrossRef]
- Roschger, P.; Matsuo, K.; Misof, B.M.; Tesch, W.; Jochum, W.; Wagner, E.F.; Fratzl, P.; Klaushofer, K. Normal mineralization and nanostructure of sclerotic bone in mice overexpressing Fra-one. Bone 2004, 34, 776–782. [Google Scholar] [CrossRef]
- Glatt, V.; Canalis, E.; Stadmeyer, L.; Bouxsein, M.L. Age-related changes in trabecular architecture differ in female and male C57BL/6J mice. J. Bone Miner. Res. 2007, 22, 1197–1207. [Google Scholar] [CrossRef]
- Blouin, S.; Fratzl-Zelman, N.; Glorieux, F.H.; Roschger, P.; Klaushofer, K.; Marini, J.C.; Rauch, F. Hypermineralization and High Osteocyte Lacunar Density in Osteogenesis Imperfecta Type V Bone Indicate Exuberant Primary Bone Formation. J. Bone Miner. Res. 2017, 32, 1884–1892. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studied Site | BMDD Parameter | PTH | ALP | Serum Ca |
|---|---|---|---|---|
| Cortical diaphysis | CaMean | ns | ns | ns |
| CaPeak | ns | ns | ns | |
| CaWidth | ns | ns | ns | |
| CaLow | 0.62 *** | 0.63 *** | −0.69 *** | |
| CaHigh | ns | ns | ns | |
| Epiphyseal spongiosa | CaMean | −0.72 *** | −0.73 *** | 0.63 *** |
| CaPeak | ns | ns | ns | |
| CaWidth | ns | ns | ns | |
| CaLow | 0.86 *** | 0.81 *** | −0.60 *** | |
| CaHigh | ns | ns | ns | |
| Metaphyseal spongiosa | CaMean | −0.63 *** | −0.60 *** | ns |
| CaPeak | ns | ns | ns | |
| CaWidth | ns | ns | ns | |
| CaLow | 0.82 *** | 0.77 *** | −0.66 *** | |
| CaHigh | ns | ns | ns |
| Studied Site | Bone Microstructure | WT BL (n = 5) | VDRΔ/Δ BL (n = 4) | WT 2CD (n = 6) | VDRΔ/Δ 2CD (n = 4) |
|---|---|---|---|---|---|
| Es | md.BV/TV (%) | 25.5 (19.6; 34.5) | 26.5 (19.2; 27.2) | 32.0 (25.0;35.0) | 9.7 ** (6.3; 19.0) |
| md.Tb.Th (μm) | 64 (61; 77) | 69 (60; 78) | 78 (68; 86) | 50 ** (47; 62) | |
| md.Tb.N (/mm) | 4.0 (2.9; 4.9) | 3.6 (3.1; 3.9) | 4.0 (3.8; 4.2) | 2.0 * (1.3; 3.0) | |
| Ms | md.BV/TV (%) | 11.5 (10.6; 18.3) | 9.6 (9.0; 15.9) | 11.1 (9.8; 13.6) | 7.1 (3.6; 12.0) |
| md.Tb.Th (μm) | 41 (37; 45) | 41 (37; 58) | 47 (41; 59) | 38 (28; 54) | |
| md.Tb.N (/mm) | 3.0 (2.8; 4.1) | 2.5 (2.2; 2.8) | 2.2 (1.9; 2.6) | 1.7 (1.2; 2.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misof, B.M.; Blouin, S.; Hofstaetter, J.G.; Roschger, P.; Zwerina, J.; Erben, R.G. No Role of Osteocytic Osteolysis in the Development and Recovery of the Bone Phenotype Induced by Severe Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice. Int. J. Mol. Sci. 2020, 21, 7989. https://doi.org/10.3390/ijms21217989
Misof BM, Blouin S, Hofstaetter JG, Roschger P, Zwerina J, Erben RG. No Role of Osteocytic Osteolysis in the Development and Recovery of the Bone Phenotype Induced by Severe Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice. International Journal of Molecular Sciences. 2020; 21(21):7989. https://doi.org/10.3390/ijms21217989
Chicago/Turabian StyleMisof, Barbara M., Stéphane Blouin, Jochen G. Hofstaetter, Paul Roschger, Jochen Zwerina, and Reinhold G. Erben. 2020. "No Role of Osteocytic Osteolysis in the Development and Recovery of the Bone Phenotype Induced by Severe Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice" International Journal of Molecular Sciences 21, no. 21: 7989. https://doi.org/10.3390/ijms21217989