The Wnt Signalling Pathway: A Tailored Target in Cancer

Abstract
1. Introduction
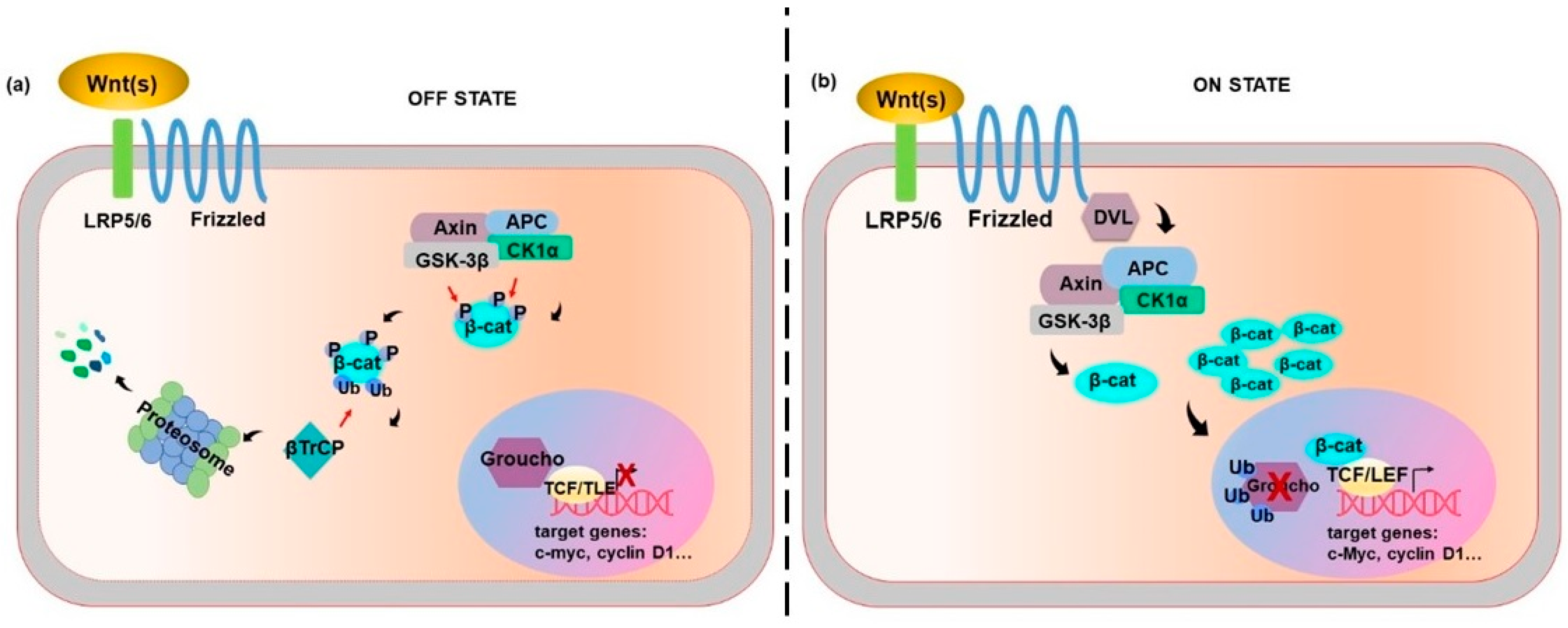
2. Wnt Canonical Pathway: β-Catenin Dependent
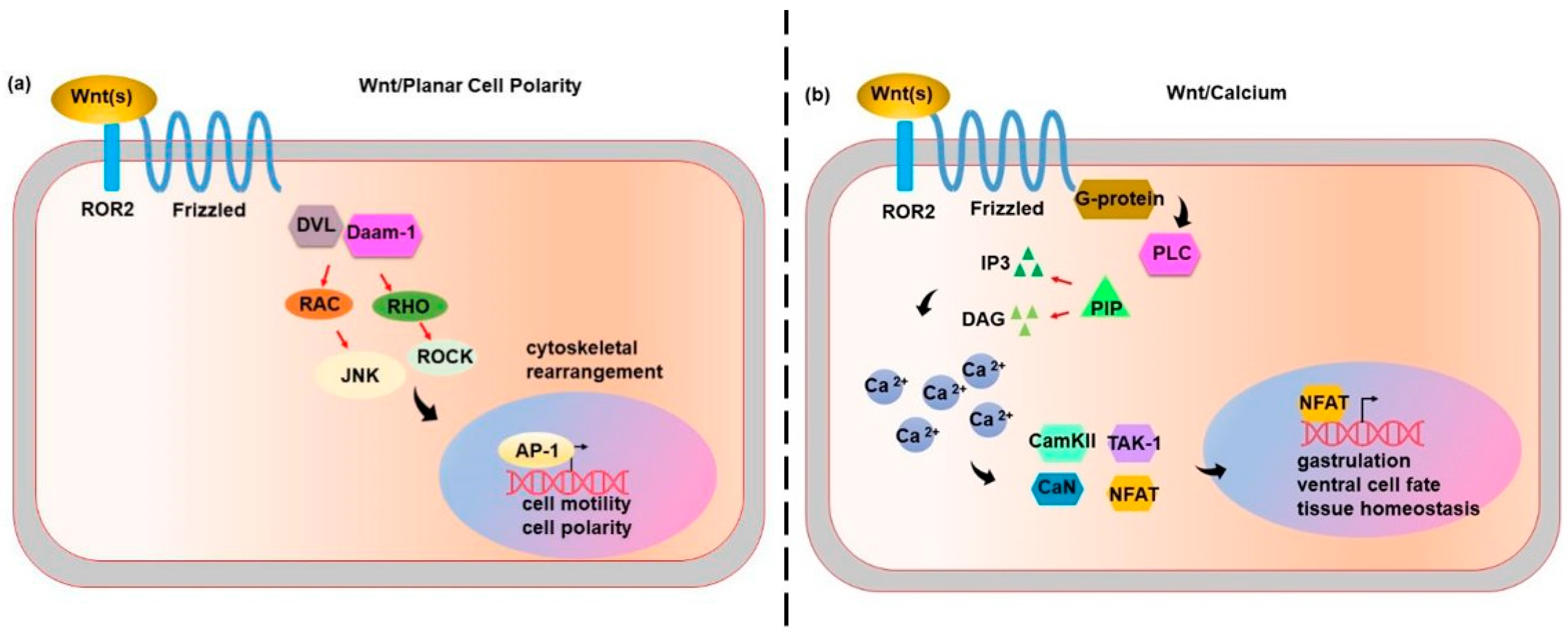
3. Wnt Non-Canonical Pathways: Wnt/Planar Cell Polarity (PCP) and Wnt/Calcium
4. Colorectal Cancer
5. Canonical Wnt/β-Catenin Pathway and CRC
6. Non-Canonical Wnt Pathway and CRC
7. Breast Cancer
8. Canonical Wnt Pathway and BC
9. Wnt Non-Canonical Pathway and BC
10. Triple-Negative Breast Cancer
11. Ovarian Cancer
12. Canonical Wnt Pathway and OC
13. Non-Canonical Wnt Pathway and OC
14. Wnt Pathway and Other Cancers
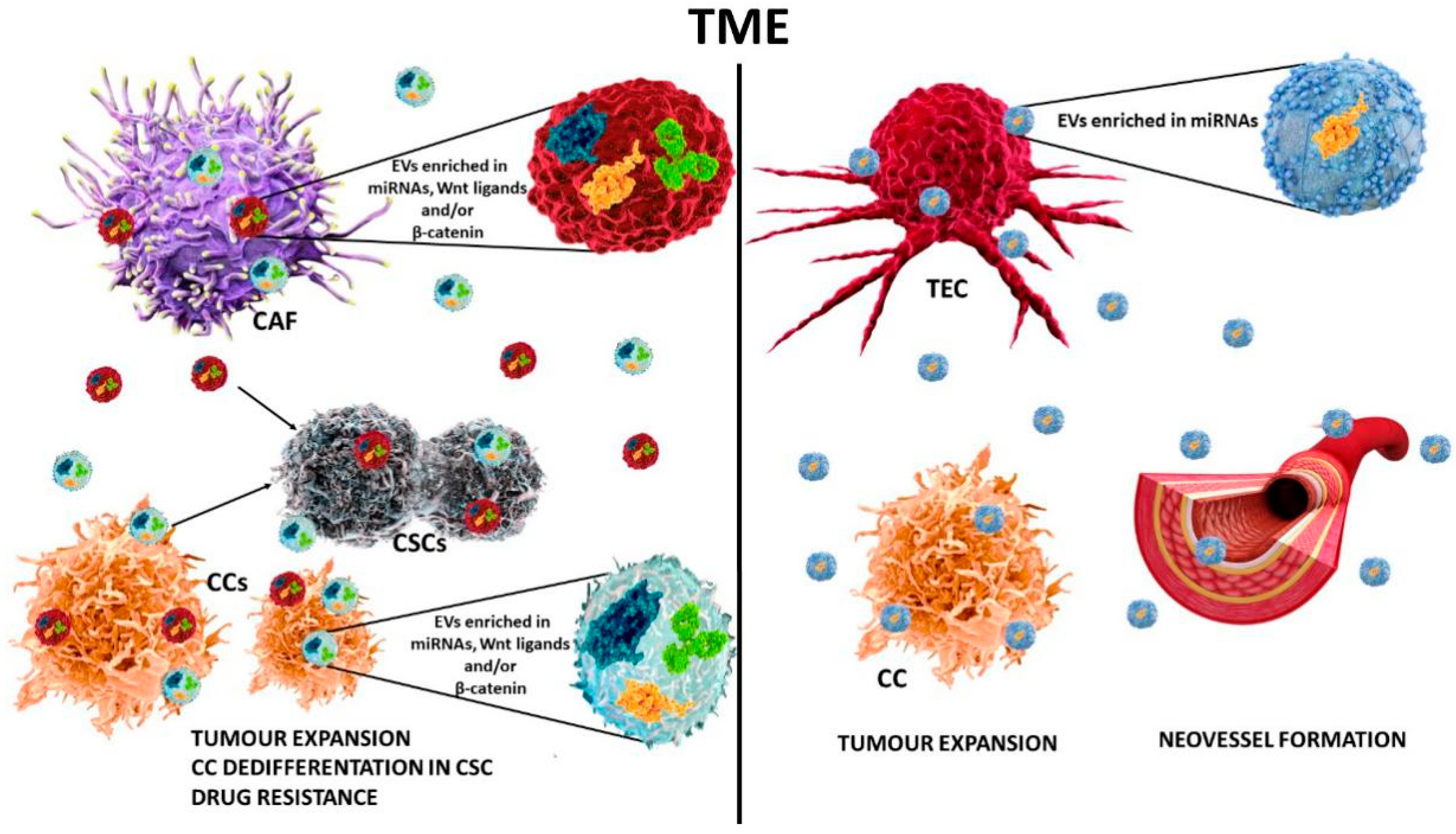
15. Extracellular Vesicles and the Wnt Pathway
16. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jackstadt, R.; Hodder, M.C.; Sansom, O.J. WNT and β-Catenin in Cancer: Genes and Therapy. Annu. Rev. Cancer Biol. 2020, 4, 177–196. [Google Scholar] [CrossRef]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Duchartre, Y.; Kim, Y.M.; Kahn, M. The Wnt signaling pathway in cancer. Crit. Rev. Oncol. Hematol. 2016, 99, 141–149. [Google Scholar] [CrossRef]
- Thrasivoulou, C.; Millar, M.; Ahmed, A. Activation of intracellular calcium by multiple Wnt ligands and translocation of β-catenin into the nucleus: A convergent model of Wnt/Ca2+and Wnt/β-catenin pathways. J. Biol. Chem. 2013, 288, 35651–35659. [Google Scholar] [CrossRef]
- Florian, M.C.; Nattamai, K.J.; Dörr, K.; Marka, G.; Überle, B.; Vas, V.; Eckl, C.; Andrä, I.; Schiemann, M.; Oostendorp, R.A.J.; et al. A canonical to non-canonical Wnt signalling switch in haematopoietic stem-cell ageing. Nature 2013, 503, 392–396. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-Catenin Signaling: Components, Mechanisms, and Diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Gajos-Michniewicz, A.; Czyz, M. WNT Signaling in Melanoma. Int. J. Mol. Sci. 2020, 21, 4852. [Google Scholar] [CrossRef]
- Wang, Y.; Nathans, J. Tissue/planar cell polarity in vertebrates: New insights and new questions. Development 2007, 134, 647–658. [Google Scholar] [CrossRef]
- Saneyoshi, T.; Kume, S.; Amsaki, Y.; Mikoshiba, K. The wnt/calcium pathway activates nf-at and promotes ventral cell fate in xenopus embryos. Nature 2002, 417, 295–299. [Google Scholar] [CrossRef]
- Tao, Q.; Yokota, C.; Puck, H.; Kofron, M.; Birsoy, B.; Yan, D.; Asashima, M.; Wylie, C.C.; Lin, X.; Heasman, J. Maternal Wnt11 activates the canonical Wnt signaling pathway required for axis formation in Xenopus embryos. Cell 2005, 120, 857–871. [Google Scholar] [CrossRef]
- Steinhart, Z.; Angers, S. Wnt signaling in development and tissue homeostasis. Development 2018, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Bhandari, A.; Woodhouse, M.; Gupta, S. Colorectal cancer is a leading cause of cancer incidence and mortality among adults younger than 50 years in the USA: A SEER-based analysis with comparison to other young-onset cancers. J. Investig. Med. 2017, 65, 311–315. [Google Scholar] [CrossRef]
- Dienstmann, R.; Wang, X.; de Reyni, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; Bot, B.M.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Wong, C.C.; Zhang, J.; Dong, Y.; Li, X.; Kang, W.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. RNF6 promotes colorectal cancer by activating the Wnt/b-catenin pathway via ubiquitination of TLE3. Cancer Res. 2018, 78, 1958–1971. [Google Scholar] [CrossRef]
- Study of the Role of Leucine-rich Repeat-Containing g-protein Coupled Receptor 5 (LGR5) and WNT Pathway in Colon Cancer. Egypt. J. Biochem. Mol. Biol. Available online: https://www.ajol.info/index.php/ejbmb/article/view/191481 (accessed on 11 September 2020).
- Yang, L.; Xie, X.; Tang, H.; Kong, Y.; Xie, X.; Chen, J.; Song, C.; Liu, X.; Ye, F.; Li, N.; et al. LGR5 promotes breast cancer progression and maintains stem-like cells through activation of wnt/β-catenin signaling. Stem Cells 2015, 33, 2913–2924. [Google Scholar] [CrossRef]
- Yu, D.H.; Zhang, X.; Wang, H.; Zhang, L.; Chen, H.; Hu, M.; Dong, Z.; Zhu, G.; Qian, Z.; Fan, J.; et al. The essential role of TNIK gene amplification in gastric cancer growth. Oncogenesis 2014, 3. [Google Scholar] [CrossRef]
- Masuda, M.; Uno, Y.; Ohbayashi, N.; Ohata, H.; Mimata, A.; Kukimoto-Niino, M.; Moriyama, H.; Kashimoto, S.; Inoue, T.; Goto, N.; et al. TNIK inhibition abrogates colorectal cancer stemness. Nat. Commun. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Cho, Y.H.; Ro, E.J.; Yoon, J.S.; Kwak, D.K.; Cho, J.; Kang, D.W.; Lee, H.Y.; Choi, K.Y. Small molecule-induced simultaneous destabilization of β-catenin and RAS is an effective molecular strategy to suppress stemness of colorectal cancer cells. Cell Commun. Signal. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, L.; Zhang, C.; Yuan, Y. MiR-144-3p inhibits cell proliferation of colorectal cancer cells by targeting BCL6 via inhibition of Wnt/β-catenin signaling. Cell. Mol. Biol. Lett. 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Polo, J.M.; Dell’Oso, T.; Ranuncolo, S.M.; Cerchietti, L.; Beck, D.; Da Silva, G.F.; Prive, G.G.; Licht, J.D.; Melnick, A. Specific peptide interference reveals BCL6 transcriptional and oncogenic mechanisms in B-cell lymphoma cells. Nat. Med. 2004, 10, 1329–1335. [Google Scholar] [CrossRef]
- Shaffer, A.L.; Yu, X.; He, Y.; Boldrick, J.; Chan, E.P.; Staudt, L.M. BCL-6 represses genes that function in lymphocyte differentiation, inflammation, and cell cycle control. Immunity 2000, 13, 199–212. [Google Scholar] [CrossRef]
- Qi, S.; Song, Y.; Peng, Y.; Wang, H.; Long, H.; Yu, X.; Li, Z.; Fang, L.; Wu, A.; Luo, W.; et al. ZEB2 mediates multiple pathways regulating cell proliferation, migration, invasion, and apoptosis in glioma. PLoS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ng, V.H.; Hang, B.I.; Sawyer, L.M.; Neitzel, L.R.; Crispi, E.E.; Rose, K.L.; Popay, T.M.; Zhong, A.; Lee, L.A.; Tansey, W.P.; et al. Phosphorylation of XIAP at threonine 180 controls its activity in Wnt signaling. J. Cell Sci. 2018. [Google Scholar] [CrossRef]
- Wels, C.; Joshi, S.; Koefinger, P.; Bergler, H. Transcriptional Activation of ZEB1 by Slug Leads to Cooperative Regulation of the EMT like Phenotype in Melanoma. J. Invest. Dermatol. 2011, 131, 1877–1885. [Google Scholar] [CrossRef]
- Zhang, M.; Miao, F.; Huang, R.; Liu, W.; Zhao, Y.; Jiao, T.; Lu, Y.; Wu, F.; Wang, X.; Wang, H.; et al. RHBDD1 promotes colorectal cancer metastasis through the Wnt signaling pathway and its downstream target ZEB1. J. Exp. Clin. Cancer Res. 2018, 37, 1–14. [Google Scholar] [CrossRef]
- Deng, M.; Chen, Z.; Tan, J.; Liu, H. Down-regulation of SLC35C1 induces colon cancer through over-activating Wnt pathway. J. Cell. Mol. Med. 2020, 24, 3079–3090. [Google Scholar] [CrossRef]
- Xu, C.; Tian, G.; Jiang, C.; Xue, H.; Kuerbanjiang, M.; Sun, L.; Gu, L.; Zhou, H.; Liu, Y.; Zhang, Z.; et al. NPTX2 promotes colorectal cancer growth and liver metastasis by the activation of the canonical Wnt/β-catenin pathway via FZD6. Cell Death Dis. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yuan, H.; Wang, J.; Wu, Y.; Xiao, Y.; Mao, S. KLHL22 regulates the EMT and proliferation in colorectal cancer cells in part via the Wnt/β-catenin signaling pathway. Cancer Manag. Res. 2020, 12, 3981–3993. [Google Scholar] [CrossRef]
- Liu, Q.; Song, J.; Pan, Y.; Shi, D.; Yang, C.; Wang, S.; Xiong, B. Wnt5a/caMKII/ERK/CCL2 axis is required for tumor-associated macrophages to promote colorectal cancer progression. Int. J. Biol. Sci. 2020, 16, 1023–1034. [Google Scholar] [CrossRef]
- Yue, Z.; Yuan, Z.; Zeng, L.; Wang, Y.; Lai, L.; Li, J.; Sun, P.; Xue, X.; Qi, J.; Yang, Z.; et al. LGR4 modulates breast cancer initiation, metastasis, and cancer stem cells. FASEB J. 2018, 32, 2422–2437. [Google Scholar] [CrossRef]
- Ruffner, H.; Lle Sprunger, J.; Charlat, O.; Leighton-Davies, J.; Grosshans, B.; Salathe, A.; Zietzling, S.; Rie Beck, V.; Therier, M.; Isken, A.; et al. R-Spondin Potentiates Wnt/b-Catenin Signaling through Orphan Receptors LGR4 and LGR5. PLoS ONE 2012. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, L.; Jiang, J.; Yu, S.; Zhou, Q. Suppression of the proliferation and invasion of breast cancer cells by ST7L occurs through inhibition of activation of Wnt/GSK-3β/β-catenin signalling. Clin. Exp. Pharmacol. Physiol. 2020, 47, 119–126. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, Y.; Li, Q. TMED3 promotes proliferation and migration in breast cancer cells by activating Wnt/β-catenin signaling. Onco. Targets. Ther. 2020, 13, 5819–5830. [Google Scholar] [CrossRef]
- Prasad, C.P.; Södergren, K.; Andersson, T. Reduced production and uptake of lactate are essential for the ability of WNT5A signaling to inhibit breast cancer cell migration and invasion. Oncotarget 2017, 8, 71471–71488. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jeon, H.M.; Ju, M.K.; Kim, C.H.; Yoon, G.; Han, S.I.; Park, H.G.; Kang, H.S. Wnt/snail signaling regulates cytochrome c oxidase and glucose metabolism. Cancer Res. 2012, 72, 3607–3617. [Google Scholar] [CrossRef] [PubMed]
- Leris, A.C.A.; Roberts, T.R.; Jiang, W.G.; Newbold, R.F.; Mokbel, K. WNT5A expression in human breast cancer. Anticancer Res. 2005, 25, 731–734. [Google Scholar]
- Jönsson, M.; Dejmek, J.; Bendahl, P.O.; Andersson, T. Loss of Wnt-5a protein is associated with early relapse in invasive ductal breast carcinomas. Cancer Res. 2002, 62, 409–416. [Google Scholar] [PubMed]
- Kobayashi, Y.; Kadoya, T.; Amioka, A.; Hanaki, H.; Sasada, S.; Masumoto, N.; Yamamoto, H.; Arihiro, K.; Kikuchi, A.; Okada, M. Wnt5a-induced cell migration is associated with the aggressiveness of estrogen receptor-positive breast cancer. Oncotarget 2018, 9, 20979–20992. [Google Scholar] [CrossRef]
- Shen, H.; Yan, W.; Yuan, J.; Wang, Z.; Wang, C. Nek2B activates the wnt pathway and promotes triple-negative breast cancer chemothezrapy-resistance by stabilizing β-catenin. J. Exp. Clin. Cancer Res. 2019, 38, 1–17. [Google Scholar] [CrossRef]
- Puvirajesinghe, T.M.; Bertucci, F.; Jain, A.; Scerbo, P.; Belotti, E.; Audebert, S.; Sebbagh, M.; Lopez, M.; Brech, A.; Finetti, P.; et al. Identification of p62/SQSTM1 as a component of non-canonical Wnt VANGL2-JNK signalling in breast cancer. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Yu, L.; Wang, C.; Pan, F.; Liu, Y.; Ren, X.; Zeng, H.; Shi, Y. HePTP promotes migration and invasion in triple-negative breast cancer cells via activation of Wnt/β-catenin signaling. Biomed. Pharmacother. 2019, 118. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.; Lv, Z.-D.; Xia, J.; Jin, L.-Y.; Yang, Z.-C. DLC-3 suppresses cellular proliferation, migration, and invasion in triple-negative breast cancer by the Wnt/β-catenin pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 1224–1232. [Google Scholar] [PubMed]
- Chen, M.W.; Yang, S.T.; Chien, M.H.; Hua, K.T.; Wu, C.J.; Hsiao, S.M.; Lin, H.; Hsiao, M.; Su, J.L.; Wei, L.H. The STAT3-miRNA-92-Wnt signaling pathway regulates spheroid formation and malignant progression in ovarian cancer. Cancer Res. 2017, 77, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Liu, A.; Zhu, J.; Lei, F.; Wu, S.; Zhang, X.; Ye, L.; Cao, L.; He, S. MiR-1207 overexpression promotes cancer stem cell-like traits in ovarian cancer by activating the Wnt/β-catenin signaling pathway. Oncotarget 2015, 6, 28882–28894. [Google Scholar] [CrossRef][Green Version]
- Ruan, X.; Liu, A.; Zhong, M.; Wei, J.; Zhang, W.; Rong, Y.; Liu, W.; Li, M.; Qing, X.; Chen, G.; et al. Silencing LGR6 Attenuates Stemness and Chemoresistance via Inhibiting Wnt/β-Catenin Signaling in Ovarian Cancer. Mol. Ther. Oncolytics 2019, 14, 94–106. [Google Scholar] [CrossRef]
- Park, S.; Cui, J.; Yu, W.; Wu, L.; Carmon, K.S.; Liu, Q.J. Differential activities and mechanisms of the four r-spondins in potentiating wnt/-catenin signaling. J. Biol. Chem. 2018, 293, 9759–9769. [Google Scholar] [CrossRef]
- Nguyen, V.H.L.; Hough, R.; Bernaudo, S.; Peng, C. Wnt/β-catenin signalling in ovarian cancer: Insights into its hyperactivation and function in tumorigenesis. J. Ovarian Res. 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Hou, R.; Jiang, L.; Yang, Z.; Wang, S.; Liu, Q. Rab14 is overexpressed in ovarian cancers and promotes ovarian cancer proliferation through Wnt pathway. Tumor Biol. 2016, 37, 16005–16013. [Google Scholar] [CrossRef]
- Asad, M.; Wong, M.K.; Tan, T.Z.; Choolani, M.; Low, J.; Mori, S.; Virshup, D.; Thiery, J.P.; Huang, R.Y.J. FZD7 drives in vitro aggressiveness in stem-A subtype of ovarian cancer via regulation of non-canonical wnt/PCP pathway. Cell Death Dis. 2014, 5, e1346. [Google Scholar] [CrossRef]
- Sun, L.; Wang, D.; Li, X.; Zhang, L.; Zhang, H.; Zhang, Y. Extracellular matrix protein ITGBL1 promotes ovarian cancer cell migration and adhesion through Wnt/PCP signaling and FAK/SRC pathway. Biomed. Pharmacother. 2016, 81, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhou, L.; Zhan, S.J.; Cheng, L.J.; Li, R.N.; Wang, H.; Liu, B.; Linghu, H. ALPL regulates the aggressive potential of high grade serous ovarian cancer cells via a non-canonical WNT pathway. Biochem. Biophys. Res. Commun. 2019, 513, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Muralidhar, S.; Filia, A.; Nsengimana, J.; Pozniak, J.; O’Shea, S.J.; Diaz, J.M.; Harland, M.; Randerson-Moor, J.A.; Reichrath, J.; Laye, J.P.; et al. Vitamin D–VDR signaling inhibits Wnt/b-catenin–mediated melanoma progression and promotes antitumor immunity. Cancer Res. 2019, 79, 5986–5998. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.S.; Jeong, W.J.; Park, J.; Kim, T.I.; Min, D.S.; Choi, K.Y. Role of oncogenic K-Ras in cancer stem cell activation by aberrant wnt/β-catenin signaling. J. Natl. Cancer Inst. 2014, 106, 1–10. [Google Scholar] [CrossRef]
- Lenz, H.J.; Kahn, M. Safely targeting cancer stem cells via selective catenin coactivator antagonism. Cancer Sci. 2014, 105, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Cha, P.H.; Cho, Y.H.; Lee, S.K.; Lee, J.; Jeong, W.J.; Moon, B.S.; Yun, J.H.; Yang, J.S.; Choi, S.; Yoon, J.; et al. Small-molecule binding of the axin RGS domain promotes β-catenin and Ras degradation. Nat. Chem. Biol. 2016, 12, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and cancer: Key paradigms in molecular therapy (Review). Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Dutta, A. MicroRNAs in Cancer Contents. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Liu, W.Y.; Yang, Z.; Sun, Q.; Yang, X.; Hu, Y.; Xie, H.; Gao, H.J.; Guo, L.M.; Yi, J.Y.; Liu, M.; et al. miR-377-3p drives malignancy characteristics via upregulating GSK-3β expression and activating NF-κB pathway in hCRC cells. J. Cell. Biochem. 2018, 119, 2124–2134. [Google Scholar] [CrossRef]
- Huang, L.; Liu, Z.; Hu, J.; Luo, Z.; Zhang, C.; Wang, L.; Wang, Z. MiR-377-3p suppresses colorectal cancer through negative regulation on Wnt/β-catenin signaling by targeting XIAP and ZEB2. Pharmacol. Res. 2020, 156, 104774. [Google Scholar] [CrossRef]
- Lv, S.; Zhang, J.; He, Y.; Liu, Q.; Wang, Z.; Liu, B.; Shi, L.; Wu, Y. MicroRNA-520e targets AEG-1 to suppress the proliferation and invasion of colorectal cancer cells through Wnt/GSK-3β/β-catenin signalling. Clin. Exp. Pharmacol. Physiol. 2020, 47, 158–167. [Google Scholar] [CrossRef]
- Hu, G.; Wei, Y.; Kang, Y. The multifaceted role of MTDH/AEG-1 in cancer progression. Clin. Cancer Res. 2009, 15, 5615–5620. [Google Scholar] [CrossRef] [PubMed]
- You, F.; Li, J.; Zhang, P.; Zhang, H.; Cao, X. MiR106a promotes the growth of transplanted breast cancer and decreases the sensitivity of transplanted tumors to cisplatin. Cancer Manag. Res. 2020, 12, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Lin, X.; Bu, J.; Lin, Z.; Chen, Y.; Qiu, Y.; Mo, H.; Tang, Y.; Fang, W.; Wu, Z. Timeless-Stimulated miR-5188-FOXO1/β-Catenin-c-Jun Feedback Loop Promotes Stemness via Ubiquitination of β-Catenin in Breast Cancer. Mol. Ther. 2019, 28, 313–327. [Google Scholar] [CrossRef]
- Han, L.; Yan, Y.; Zhao, L.; Liu, Y.; Lv, X.; Zhang, L.; Zhao, Y.; Zhao, H.; He, M.; Wei, M. LncRNA HOTTIP facilitates the stemness of breast cancer via regulation of miR-148a-3p/WNT1 pathway. J. Cell. Mol. Med. 2020, 24, 6242–6252. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, P.; Zhang, H. MiR-6838-5p suppresses cell metastasis and the EMT process in triple-negative breast cancer by targeting WNT3A to inhibit the Wnt pathway. J. Gene Med. 2019, 21, 1–8. [Google Scholar] [CrossRef]
- Wu, R.; Zhao, B.; Ren, X.; Wu, S.; Liu, M.; Wang, Z.; Liu, W. Mir-27a-3p targeting gsk3β promotes triple-negative breast cancer proliferation and migration through wnt/β-catenin pathway. Cancer Manag. Res. 2020, 12, 6241–6249. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.; Shan, Y.; Bernaudo, S.; Peng, C. miR-590-3p Targets Cyclin G2 and FOXO3 to Promote Ovarian Cancer Cell Proliferation, Invasion, and Spheroid Formation. Int. J. Mol. Sci. 2019, 20, 1810. [Google Scholar] [CrossRef]
- Salem, M.; O’brien, J.A.; Bernaudo, S.; Shawer, H.; Ye, G.; Brki, J.; Amleh, A.; Vanderhyden, B.C.; Refky, B.; Yang, B.B.; et al. miR-590-3p Promotes Ovarian Cancer Growth and Metastasis via a Novel FOXA2-Versican Pathway. Cancer Res. 2018. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, W.; Huang, Y.; Wang, Z.; Jiang, T.; Wang, L. MiR-1180 from bone marrow MSCs promotes cell proliferation and glycolysis in ovarian cancer cells via SFRP1/Wnt pathway. Cancer Cell Int. 2019, 19, 66. [Google Scholar] [CrossRef]
- Situ, J.; Zhang, H.; Jin, Z.; Li, K.; Mao, Y.; Huang, W. MicroRNA-939 directly targets HDGF to inhibit the aggressiveness of prostate cancer via deactivation of the WNT/β-catenin pathway. Onco. Targets. Ther. 2020, 13, 4257–4270. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Min, Z.; Linhong, M.; Tao, R.; Yan, L.; Song, H. Hypoxia-induced exosomes promote hepatocellular carcinoma proliferation and metastasis via miR-1273f transfer. Exp. Cell Res. 2019, 385. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wei, K.; Hu, L.Q.; Zhou, C.R.; Lu, Z.B.; Zhan, G.S.; Pan, X.L.; Pan, C.F.; Wang, J.; Wen, W.; et al. Exosome-mediated transfer of miR-1260b promotes cell invasion through Wnt/β–catenin signaling pathway in lung adenocarcinoma. J. Cell. Physiol. 2020, 235, 6843–6853. [Google Scholar] [CrossRef]
- Lombardo, G.; Gili, M.; Grange, C.; Cavallari, C.; Dentelli, P.; Togliatto, G.; Taverna, D.; Camussi, G.; Brizzi, M.F. IL-3R-alpha blockade inhibits tumor endothelial cell-derived extracellular vesicle (EV)-mediated vessel formation by targeting the β-catenin pathway. Oncogene 2018, 37, 1175–1191. [Google Scholar] [CrossRef] [PubMed]
- Vymetalkova, V.; Vodicka, P.; Vodenkova, S.; Alonso, S.; Schneider-Stock, R. DNA methylation and chromatin modifiers in colorectal cancer. Mol. Aspects Med. 2019, 69, 73–92. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Mashima, T.; Mizutani, A.; Sato, A.; Aoyama, A.; Gong, B.; Yoshida, H.; Muramatsu, Y.; Nakata, K.; Matsuura, M.; et al. APC mutations as a potential biomarker for sensitivity to tankyrase inhibitors in colorectal cancer. Mol. Cancer Ther. 2017, 16, 752–762. [Google Scholar] [CrossRef]
- Bruschi, M.; Garnier, L.; Cleroux, E.; Giordano, A.; Dumas, M.; Bardet, A.F.; Kergrohen, T.; Quesada, S.; Cesses, P.; Weber, M.; et al. Loss of Apc Rapidly Impairs DNA Methylation Programs and Cell Fate Decisions in Lgr5+ Intestinal Stem Cells. Cancer Res. 2020, 80. [Google Scholar] [CrossRef]
- Proto, M.C.; Fiore, D.; Piscopo, C.; Franceschelli, S.; Bizzarro, V.; Laezza, C.; Lauro, G.; Feoli, A.; Tosco, A.; Bifulco, G.; et al. Inhibition of Wnt/β-Catenin pathway and Histone acetyltransferase activity by Rimonabant: A therapeutic target for colon cancer. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Liu, Y.; Rubin, B.; Bodine, P.V.N.; Billiard, J. Wnt5a induces homodimerization and activation of Ror2 receptor tyrosine kinase. J. Cell. Biochem. 2008, 105, 497–502. [Google Scholar] [CrossRef]
- Lee, J.M.; Kim, I.S.; Kim, H.; Lee, J.S.; Kim, K.; Yim, H.Y.; Jeong, J.; Kim, J.H.; Kim, J.Y.; Lee, H.; et al. RORα Attenuates Wnt/β-Catenin Signaling by PKCα-Dependent Phosphorylation in Colon Cancer. Mol. Cell 2010, 37, 183–195. [Google Scholar] [CrossRef]
- Voloshanenko, O.; Schwartz, U.; Kranz, D.; Rauscher, B.; Linnebacher, M.; Augustin, I.; Boutros, M. β-catenin-independent regulation of Wnt target genes by RoR2 and ATF2/ATF4 in colon cancer cells. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, M.; Ueno, K.; Hazama, S.; Okada, T.; Sakai, K.; Suehiro, Y.; Okayama, N.; Hirata, H.; Oka, M.; Imai, K.; et al. Possible involvement of Wnt11 in colorectal cancer progression. Mol. Carcinog. 2013, 52, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Gorroño-Etxebarria, I.; Aguirre, U.; Sanchez, S.; González, N.; Escobar, A.; Zabalza, I.; Quintana, J.M.; dM Vivanco, M.; Waxman, J.; Kypta, R.M. Wnt-11 as a potential prognostic biomarker and therapeutic target in colorectal cancer. Cancers (Basel) 2019, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vitale, I.; Manic, G.; Coussens, L.M.; Kroemer, G.; Galluzzi, L. Macrophages and Metabolism in the Tumor Microenvironment. Cell Metab. 2019, 30, 36–50. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Du, X.L.; Fox, E.E.; Lai, D. Competing Causes of Death for Women With Breast Cancer and Change Over Time From 1975 to 2003 NIH Public Access. Am. J. Clin. Oncol. 2008, 31, 105–116. [Google Scholar] [CrossRef]
- Yu, Q.C.; Verheyen, E.M.; Zeng, Y.A. Mammary development and breast cancer: A Wnt perspective. Cancers 2016, 8, 65. [Google Scholar] [CrossRef]
- Katoh, M. Canonical and non-canonical WNT signaling in cancer stem cells and their niches: Cellular heterogeneity, omics reprogramming, targeted therapy and tumor plasticity (Review). Int. J. Oncol. 2017, 51, 1357–1369. [Google Scholar] [CrossRef]
- Khramtsov, A.I.; Khramtsova, G.F.; Tretiakova, M.; Huo, D.; Olopade, O.I.; Goss, K.H. Wnt/β-catenin pathway activation is enriched in basal-like breast cancers and predicts poor outcome. Am. J. Pathol. 2010, 176, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, R.T.; Leong, A.S.Y.; Vääräniemi, J.P.; Fernando, S.S.E.; Eskelinen, S.M. Immunolocalization of the fodrin, E-cadherin, and β-catenin adhesion complex in infiltrating ductal carcinoma of the breast—Comparison with an in vitro model. J. Pathol. 1999, 187, 416–423. [Google Scholar] [CrossRef]
- Lacroix-Triki, M.; Geyer, F.C.; Lambros, M.B.; Savage, K.; Ellis, I.O.; Lee, A.H.S.; Reis-Filho, J.S. β-Catenin/Wnt signalling pathway in fibromatosis, metaplastic carcinomas and phyllodes tumours of the breast. Mod. Pathol. 2010, 23, 1438–1448. [Google Scholar] [CrossRef]
- Samarzija, I.; Sini, P.; Schlange, T.; MacDonald, G.; Hynes, N.E. Wnt3a regulates proliferation and migration of HUVEC via canonical and non-canonical Wnt signaling pathways. Biochem. Biophys. Res. Commun. 2009, 386, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Reintsch, W.; Steinbeisser, H. Xenopus frizzled 7 can act in canonical and non-canonical Wnt signaling pathways: Implications on early patterning and morphogenesis. Mech. Dev. 2000, 92, 227–237. [Google Scholar] [CrossRef]
- Michaelson, J.S.; Leder, P. β-catenin is a downstream effector of Wnt-mediated tumorigenesis in the mammary gland. Oncogene 2001, 20, 5093–5099. [Google Scholar] [CrossRef][Green Version]
- Klopocki, E.; Kristiansen, G.; Wild, P.J.; Klaman, I.; Castanos-Velez, E.; Singer, G.; Stöhr, R.; Simon, R.; Sauter, G.; Leibiger, H.; et al. Loss of SFRP1 is associated with breast cancer progression and poor prognosis in early stage tumors. Int. J. Oncol. 2004, 25, 641–649. [Google Scholar] [CrossRef]
- Woodward, W.A.; Chen, M.S.; Behbod, F.; Alfaro, M.P.; Buchholz, T.A.; Rosen, J.M. WNT/β-catenin mediates radiation resistance of mouse mammary progenitor cells. Proc. Natl. Acad. Sci. USA 2007, 104, 618–623. [Google Scholar] [CrossRef]
- Zhu, X.; Feng, J.; Fu, W.; Shu, X.; Wan, X.; Liu, J. Effects of cisplatin on the proliferation, invasion and apoptosis of breast cancer cells following β-catenin silencing. Int. J. Mol. Med. 2020, 45, 1838–1850. [Google Scholar] [CrossRef]
- Zhou, J.; Su, C.M.; Chen, H.A.; Du, S.; Li, C.W.; Wu, H.; Tsai, S.H.; Yeh, Y.T. Cryptanshinone inhibits the glycolysis and inhibits cell migration through PKM2/β-catenin axis in breast cancer. Onco. Targets. Ther. 2020, 13, 8629–8639. [Google Scholar] [CrossRef] [PubMed]
- Mikels, A.J.; Nusse, R. Purified Wnt5a protein activates or inhibits β-catenin-TCF signaling depending on receptor context. PLoS Biol. 2006, 4, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.P.; Bradley, A.; McMahon, A.P.; Jones, S. A Wnt5a pathway underlies outgrowth of multiple structures in the vertebrate embryo. Development 1999, 126, 1211–1223. [Google Scholar] [PubMed]
- Säfholm, A.; Tuomela, J.; Rosenkvist, J.; Dejmek, J.; Härkönen, P.; Andersson, T. The wnt-5a-derived hexapeptide Foxy-5 inhibits breast cancer metastasis in vivo by targeting cell motility. Clin. Cancer Res. 2008, 14, 6556–6563. [Google Scholar] [CrossRef] [PubMed]
- Borcherding, N.; Kusner, D.; Kolb, R.; Xie, Q.; Li, W.; Velez, G.; Askeland, R.; Weigel, R.J.; Zhang, W.; City, I. Paracrine WNT5A signaling inhibits expansion of tumor-initiating cells. Cancer Res. 2015, 75, 1972–1982. [Google Scholar] [CrossRef] [PubMed]
- Roarty, K.; Baxley, S.E.; Crowley, M.R.; Frost, A.R.; Serra, R. Loss of TGF-β or Wnt5a results in an increase in Wnt/β-catenin activity and redirects mammary tumour phenotype. Breast Cancer Res. 2009, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bergenfelz, C.; Medrek, C.; Ekström, E.; Jirström, K.; Janols, H.; Wullt, M.; Bredberg, A.; Leandersson, K. Wnt5a Induces a Tolerogenic Phenotype of Macrophages in Sepsis and Breast Cancer Patients. J. Immunol. 2012, 188, 5448–5458. [Google Scholar] [CrossRef]
- Bergenfelz, C.; Janols, H.; Wullt, M.; Jirström, K.; Bredberg, A.; Leandersson, K. Wnt5a Inhibits Human Monocyte-Derived Myeloid Dendritic Cell Generation. Scand. J. Immunol. 2013, 78, 194–204. [Google Scholar] [CrossRef]
- Fernandez-Cobo, M.; Zammarchi, F.; Mandeli, J.; Holland, J.F.; Pogo, B.G.T. Expression of Wnt5A and Wnt10B in non-immortalized breast cancer cells. Oncol. Rep. 2007, 17, 903–907. [Google Scholar] [CrossRef]
- Prasad, C.P.; Chaurasiya, S.K.; Guilmain, W.; Andersson, T. WNT5A signaling impairs breast cancer cell migration and invasion via mechanisms independent of the epithelial-mesenchymal transition. J. Exp. Clin. Cancer Res. 2016, 35, 1–15. [Google Scholar] [CrossRef]
- Bauer, M.; Bénard, J.; Gaasterland, T.; Willert, K.; Cappellen, D. WNT5A encodes two isoforms with distinct functions in cancers. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Abramson, V.G.; Lehmann, B.D.; Ballinger, T.J.; Pietenpol, J.A. Subtyping of triple-negative breast cancer: Implications for therapy. Cancer 2015, 121, 8–16. [Google Scholar] [CrossRef]
- Marra, A.; Viale, G.; Curigliano, G. Recent advances in triple negative breast cancer: The immunotherapy era. BMC Med. 2019, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef]
- Geyer, F.C.; Pareja, F.; Weigelt, B.; Rakha, E.; Ellis, I.O.; Schnitt, S.J.; Reis-Filho, J.S. The Spectrum of Triple-Negative Breast Disease: High- and Low-Grade Lesions. Am. J. Pathol. 2017, 187, 2139–2151. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Hou, R. Tetrandrine Reverses Paclitaxel Resistance in Human Ovarian Cancer via Inducing Apoptosis, Cell Cycle Arrest Through β-Catenin Pathway. Onco. Targets. Ther. 2020, 13, 3631–3639. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, A.B.; Joseph, P.; Kovalenko, O.; Singh, S.; Armstrong, A.; Redline, R.; Resnick, K.; Zanotti, K.; Waggoner, S.; Di Feo, A. Critical role of Wnt/ß-catenin signaling in driving epithelial ovarian cancer platinum resistance. Oncotarget 2015, 6, 23720–23734. [Google Scholar] [CrossRef] [PubMed]
- Sauriol, A.; Simeone, K.; Portelance, L.; Meunier, L.; Leclerc-Desaulniers, K.; De Ladurantaye, M.; Chergui, M.; Kendall-Dupont, J.; Rahimi, K.; Carmona, E.; et al. Modeling the diversity of epithelial ovarian cancer through ten novel well characterized cell lines covering multiple subtypes of the disease. Cancers 2020, 12, 2222. [Google Scholar] [CrossRef]
- Jeong, J.W.; Lee, H.S.; Franco, H.L.; Broaddus, R.R.; Taketo, M.M.; Tsai, S.Y.; Lydon, J.P.; DeMayo, F.J. Β-Catenin Mediates Glandular Formation and Dysregulation of Β-Catenin Induces Hyperplasia Formation in the Murine Uterus. Oncogene 2009, 28, 31–40. [Google Scholar] [CrossRef]
- Ryland, G.L.; Bearfoot, J.L.; Doyle, M.A.; Boyle, S.E.; Choong, D.Y.H.; Rowley, S.M.; Tothill, R.W.; Gorringe, K.L.; Campbell, I.G. MicroRNA Genes and Their Target 3′-Untranslated Regions Are Infrequently Somatically Mutated in Ovarian Cancers. PLoS ONE 2012, 7, e35805. [Google Scholar] [CrossRef]
- Barghout, S.H.; Zepeda, N.; Xu, Z.; Steed, H.; Lee, C.H.; Fu, Y. Elevated β-catenin activity contributes to carboplatin resistance in A2780cp ovarian cancer cells. Biochem. Biophys. Res. Commun. 2015, 468, 173–178. [Google Scholar] [CrossRef]
- Shin, W.S.; Gim, J.; Won, S.; Lee, S.T. Biphasic regulation of tumorigenesis by PTK7 expression level in esophageal squamous cell carcinoma. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Berger, H.; Breuer, M.; Peradziryi, H.; Podleschny, M.; Jacob, R.; Borchers, A. PTK7 localization and protein stability is affected by canonical Wnt ligands. J. Cell Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, G.; Yin, Y.; Wang, J.; Wang, H.; Wei, W.; Guo, Q.; Ma, H.; Shi, Q.; Zhou, X.; et al. PTK7 protein is decreased in epithelial ovarian carcinomas with poor prognosis. Int. J. Clin. Exp. Pathol. 2014, 7, 7881–7889. [Google Scholar] [PubMed]
- Golubkov, V.S.; Chekanov, A.V.; Cieplak, P.; Aleshin, A.E.; Chernov, A.V.; Zhu, W.; Radichev, I.A.; Zhang, D.; Dong, P.D.; Strongin, A.Y. The Wnt/planar cell polarity protein-tyrosine kinase-7 (PTK7) is a highly efficient proteolytic target of membrane type-1 matrix metalloproteinase: Implications in cancer and embryogenesis. J. Biol. Chem. 2010, 285, 35740–35749. [Google Scholar] [CrossRef]
- Zhao, J.B.; Xue, J.F.; Zhang, W.Z.; Ren, Y.L.; Yan, D.M. Long noncoding RNA FGD5-AS1 promotes glioma cell proliferation, migration and invasion by regulating wnt/β-catenin pathway. Cancer Manag. Res. 2020, 12, 6187–6193. [Google Scholar] [CrossRef]
- Nomura, M.; Rainusso, N.; Lee, Y.C.; Dawson, B.; Coarfa, C.; Han, R.; Larson, J.L.; Shuck, R.; Kurenbekova, L.; Yustein, J.T. Tegavivint and the β-Catenin/ALDH Axis in Chemotherapy-Resistant and Metastatic Osteosarcoma. J. Natl. Cancer Inst. 2019, 111, 1216–1227. [Google Scholar] [CrossRef]
- Chen, T.; Lei, S.; Zeng, Z.; Zhang, J.; Xue, Y.; Sun, Y.; Lan, J.; Xu, S.; Mao, D.; Guo, B. Linc00261 inhibits metastasis and the WNT signaling pathway of pancreatic cancer by regulating a miR-552-5p/FOXO3 axis. Oncol. Rep. 2020, 43, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.S.; De Beer, M.A.; Giepmans, B.N.G.; Zuhorn, I.S. Endocytosis of Extracellular Vesicles and Release of Their Cargo from Endosomes. ACS Nano 2020, 14, 4444–4455. [Google Scholar] [CrossRef]
- Kalra, H.; Gangoda, L.; Fonseka, P.; Chitti, S.V.; Liem, M.; Keerthikumar, S.; Samuel, M.; Boukouris, S.; Al Saffar, H.; Collins, C.; et al. Extracellular vesicles containing oncogenic mutant β-catenin activate Wnt signalling pathway in the recipient cells. J. Extracell. Vesicles 2019, 8. [Google Scholar] [CrossRef]
- Dovrat, S.; Caspi, M.; Zilberberg, A.; Lahav, L.; Firsow, A.; Gur, H.; Rosin-Arbesfeld, R. 14-3-3 and β-catenin are secreted on extracellular vesicles to activate the oncogenic Wnt pathway. Mol. Oncol. 2014, 8, 894–911. [Google Scholar] [CrossRef]
- Hu, Y.B.; Yan, C.; Mu, L.; Mi, Y.L.; Zhao, H.; Hu, H.; Li, X.L.; Tao, D.D.; Wu, Y.Q.; Gong, J.P.; et al. Exosomal Wnt-induced dedifferentiation of colorectal cancer cells contributes to chemotherapy resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef]
- Szvicsek, Z.; Oszvald, Á.; Szabó, L.; Sándor, G.O.; Kelemen, A.; Soós, A.Á.; Pálóczi, K.; Harsányi, L.; Tölgyes, T.; Dede, K.; et al. Extracellular vesicle release from intestinal organoids is modulated by Apc mutation and other colorectal cancer progression factors. Cell. Mol. Life Sci. 2019, 76, 2463–2476. [Google Scholar] [CrossRef]
- Yamashita, T.; Budhu, A.; Forgues, M.; Xin, W.W. Activation of hepatic stem cell marker EpCAM by Wnt-β-catenin signaling in hepatocellular carcinoma. Cancer Res. 2007, 67, 10831–10839. [Google Scholar] [CrossRef]
- Ishiguro, K.; Yan, I.K.; Lewis-Tuffin, L.; Patel, T. Targeting Liver Cancer Stem Cells Using Engineered Biological Nanoparticles for the Treatment of Hepatocellular Cancer. Hepatol. Commun. 2020, 4, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Faict, S.; Muller, J.; De Veirman, K.; De Bruyne, E.; Maes, K.; Vrancken, L.; Heusschen, R.; De Raeve, H.; Schots, R.; Vanderkerken, K.; et al. Exosomes play a role in multiple myeloma bone disease and tumor development by targeting osteoclasts and osteoblasts. Blood Cancer J. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Wu, A.T.H.; Bamodu, O.A.; Yadav, V.K.; Chao, T.Y.; Tzeng, Y.M.; Mukhopadhyay, D.; Hsiao, M.; Lee, J.C. Ovatodiolide suppresses oral cancer malignancy by down-regulating exosomal mir-21/STAT3/β-catenin cargo and preventing oncogenic transformation of normal gingival fibroblasts. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Yamamoto, H.; Kishida, S.; Kishida, M.; Awada, C.; Takao, T.; Kikuchi, A. Wnt5b-associated exosomes promote cancer cell migration and proliferation. Cancer Sci. 2017, 108, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.W.; Cao, C.H.; Han, K.; Zhao, Y.X.; Cai, M.Y.; Xiang, Z.C.; Zhang, J.X.; Chen, J.W.; Zhong, L.P.; Huang, Y.; et al. APC-activated long noncoding RNA inhibits colorectal carcinoma pathogenesis through reduction of exosome production. J. Clin. Investig. 2019, 129, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, T.; Grange, C.; Cavallari, C.; Navarro-Tableros, V.; Lombardo, G.; Rosso, A.; Cedrino, M.; Pomatto, M.A.C.; Koni, M.; Veneziano, F.; et al. Targeting IL-3Rα on tumor-derived endothelial cells blunts metastatic spread of triple-negative breast cancer via extracellular vesicle reprogramming. Oncogenesis 2020, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Kamerkar, S.; Lebleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef]
- Matsuda, A.; Ishiguro, K.; Yan, I.K.; Patel, T. Extracellular Vesicle-Based Therapeutic Targeting of β-Catenin to Modulate Anticancer Immune Responses in Hepatocellular Cancer. Hepatol. Commun. 2019, 3, 525–541. [Google Scholar] [CrossRef]
- Cavallari, C.; Camussi, G.; Brizzi, M.F. Extracellular Vesicles in the Tumour Microenvironment: Eclectic Supervisors. Int. J. Mol. Sci. 2020, 21, 6768. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Lam, E.W.F.; Sun, Y. Extracellular vesicles in the tumor microenvironment: Old stories, but new tales. Mol. Cancer 2019, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lucchetti, D.; Tenore, C.R.; Colella, F.; Sgambato, A. Extracellular Vesicles and Cancer: A Focus on Metabolism, Cytokines, and Immunity. Cancers 2020, 12, 171. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein | Related Cancers | Expression Level | Pathway Interaction | Impact on Tumour | Ref. |
|---|---|---|---|---|---|
| RNF6 | CRC | Upregulated | β-catenin | cell growth cell cycle progression EMT metastasis | [15] |
| LGR5 | CRC, BC | Upregulated | β-catenin | proliferation migration | [16,17] |
| TNIK | Gastric | Upregulated | β-catenin | cell growth | [18,19] |
| KYA1797K | CRC | Upregulated | β-catenin | tumour growth stem cell features | [20] |
| BCL6 | CRC | Upregulated | β-catenin | cellular proliferation tumour development tumour progression | [21,22,23] |
| ZEB2 and ZEB1 | CRC | Upregulated | β-catenin | tumour progression invasion | [17,24,25,26] |
| XIAP | CRC | Upregulated | β-catenin | proliferation chemoresistance | [24,25] |
| RHBDD1 | CRC | Upregulated | β-catenin | metastasis stemness EMT migration invasiveness | [27] |
| SLC35C1 | CRC | Downregulated | β-catenin | cell proliferation cell progression | [28] |
| NPTX2 | CRC | Upregulated | β-catenin | tumour stages lymphatic invasion metastasis | [29] |
| KLHL22 | CRC | Downregulated | β-catenin | invasion migration | [30] |
| CCL2 | CRC | Upregulated | Non-canonical | progression | [31] |
| LGR4 | BC | Upregulated | β-catenin | tumorigenesis metastasis CSC maintenance | [17,26,32,33] |
| ST7L | BC | Downregulated | GSK-3β | proliferation invasion | [34] |
| TMED | BC | Upregulated | β-catenin | cell cycle progression colony formation migration | [35] |
| Wnt5a | BC | Downregulated | β-catenin | migration lactate production invasion | [36,37] |
| Wnt5a | BC | Downregulated | β-catenin -cyclin D1 -TGF-β | cell proliferation aggressiveness | [38,39] |
| Wnt5a | BC | Upregulated | ALCAM | vessel invasion tumour size migration | [40] |
| Nek2B | TNBC | Upregulated | β-catenin | chemoresistance | [41] |
| VANGL2 | TNBC | Upregulated | p62/SQSTM1 (PCP) | migration anchorage-dependent and independent cell proliferation | [42] |
| HePTP | TNBC | Upregulated | -GSK-3β β-catenin | metastasis | [43] |
| DLC-3 | TNBC | Downregulated | β-catenin | proliferation colony formation migration invasion | [44] |
| DKK1 | OC | Downregulated | β-catenin | stemness | [45] |
| SFRP1 | OC | Downregulated | β-catenin | cell growth stem-like phenotype | [46] |
| AXIN2 | OC | Downregulated | β-catenin | stem-like phenotype | [46] |
| LGR6 | OC | Upregulated | β-catenin | stemness chemoresistance | [47,48] |
| RAB14 | OC | Upregulated | β-catenin | proliferation chemoresistance invasion | [49,50] |
| FZD7 | OC | Upregulated | Non-canonical | EMT cell cycle progression migration | [51] |
| ITGBL1 | OC | Upregulated | Non-canonical | migration adhesion | [52] |
| ALPL | OC | Upregulated | Non-canonical | EMT migration invasion | [53] |
| VDR | Melanoma | Upregulated | β-catenin | tumour growth immune response | [54] |
| miRNA | Related Cancer | Expression Level | Impact on Tumour | Ref. |
|---|---|---|---|---|
| miR-144-3p | CRC | Downregulated | cell proliferation | [21,22,23] |
| miR-377-3p | CRC | Upregulated | cell expansion EMT repression of apoptosis | [60] |
| miR-377-3p | CRC | Downregulated | proliferation migration chemoresistance | [61] |
| miR-520e | CRC | Downregulated | cell proliferation colony formation invasion | [62,63] |
| miR106a | BC | Upregulated | cell growth cisplatin sensitivity | [64] |
| miR-5188 | BC | Upregulated | tumour cell proliferation metastasis formation EMT chemoresistance | [65] |
| miR-148a | BC | Downregulated | cell migration invasion | [66] |
| miR-6838-5p | BC | Downregulated | cell invasion migration EMT | [42,67] |
| miR-27a-3p | BC | Upregulated | proliferation migration. | [68] |
| miR-1207 | OC | Upregulated | tumorigenicity stem cell-like traits stemness | [46] |
| miR-590-3p | OC | Upregulated | cell growth migration, invasion | [69,70] |
| miR-1180 | OC | Upregulated | cell proliferation glycolysis | [71] |
| miR-939 | PCa | Downregulated | tumour stage metastasis | [72] |
| miR-92a-3p | CRC EVs | Upregulated | cancer progression stemness EMT drug resistance | [73] |
| miR-1273f | HCC EVs | Upregulation | cell proliferation migration invasiveness EMT | [74] |
| miR-1260b | LAC EVs | Upregulation | cell invasion metastasis | [75] |
| miR-214-3p | TEC EVs | Upregulation | neovessel formation | [76] |
| miR-24-3p | TEC EVs | Downregulation | neovessel formation | [76] |
| EV Cargo | EV Source | Target Cells | Related Cancers | Expression Level | Pathway Interaction | Impact on Tumour Cells | Ref. |
|---|---|---|---|---|---|---|---|
| Mutant β-catenin in EVs | LIM1215 | RKO | CRC | Upregulated | β-catenin | migration, metastasis tumour growth | [129] |
| 14-3-3ζ in EVs | HEK293T | COS-7, SW480 | CRC | Upregulated | β-catenin GSK-3β DVL2 | survival migration | [130] |
| Wnt ligands in EVs | CAFs | CRC | CRC | Upregulated | β-catenin | dedifferentiation drug resistance colony formation | [131,132] |
| β-catenin in EVs | milk | HCC | HCC | Silenced | β-catenin | proliferation tumour growth | [133,134] |
| DKK-1 in EVs | MM | MM | MM | Upregulated | β-catenin | osteoclast activity osteoblast differentiation | [135] |
| EVs | OSCC | OSCC | OSCC | Upregulated | β-catenin | metastasis stemness chemoresistance | [136] |
| Wnt5b in EVs | Caco-2 and PANC-1 | A549 | Lung cancer | Upregulated | β-catenin dependent and independent pathways | proliferation migration | [137] |
| EVs | CAFs | BC | BC | Upregulated | Wnt-PCP | cell growth and motility | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koni, M.; Pinnarò, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. https://doi.org/10.3390/ijms21207697
Koni M, Pinnarò V, Brizzi MF. The Wnt Signalling Pathway: A Tailored Target in Cancer. International Journal of Molecular Sciences. 2020; 21(20):7697. https://doi.org/10.3390/ijms21207697
Chicago/Turabian StyleKoni, Malvina, Veronica Pinnarò, and Maria Felice Brizzi. 2020. "The Wnt Signalling Pathway: A Tailored Target in Cancer" International Journal of Molecular Sciences 21, no. 20: 7697. https://doi.org/10.3390/ijms21207697
APA StyleKoni, M., Pinnarò, V., & Brizzi, M. F. (2020). The Wnt Signalling Pathway: A Tailored Target in Cancer. International Journal of Molecular Sciences, 21(20), 7697. https://doi.org/10.3390/ijms21207697