Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Nanoparticle Characterization
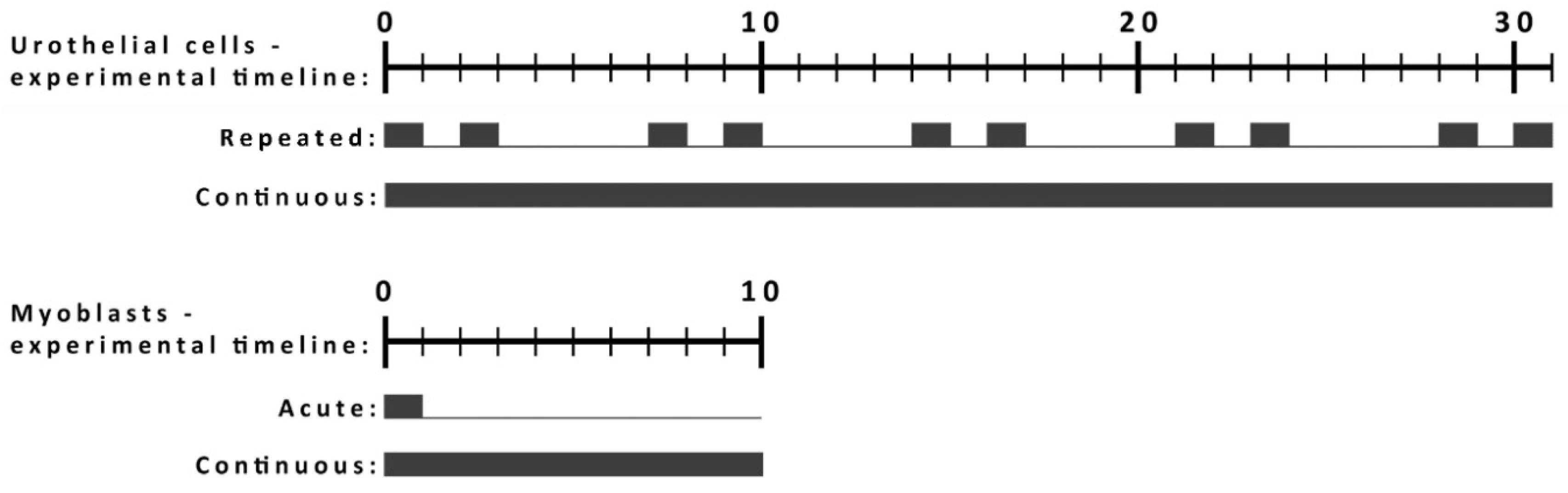
2.2. The Characteristics of L6 and NPU Cells and Exposure Protocols for the Long-Term Studies
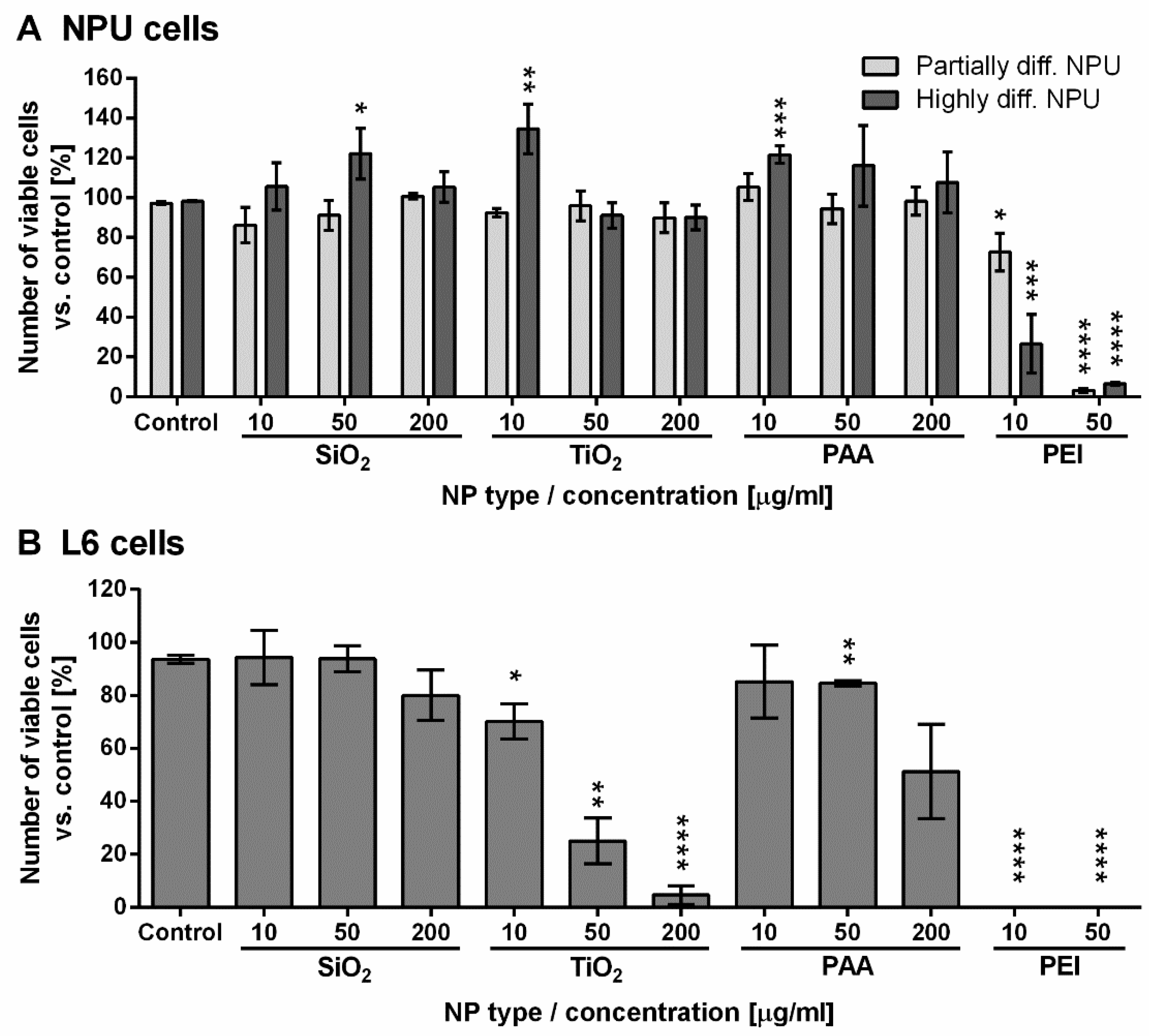
2.3. Short-Term Assessment of NPU and L6 Cell Viability for 96 h Exposure to NPs
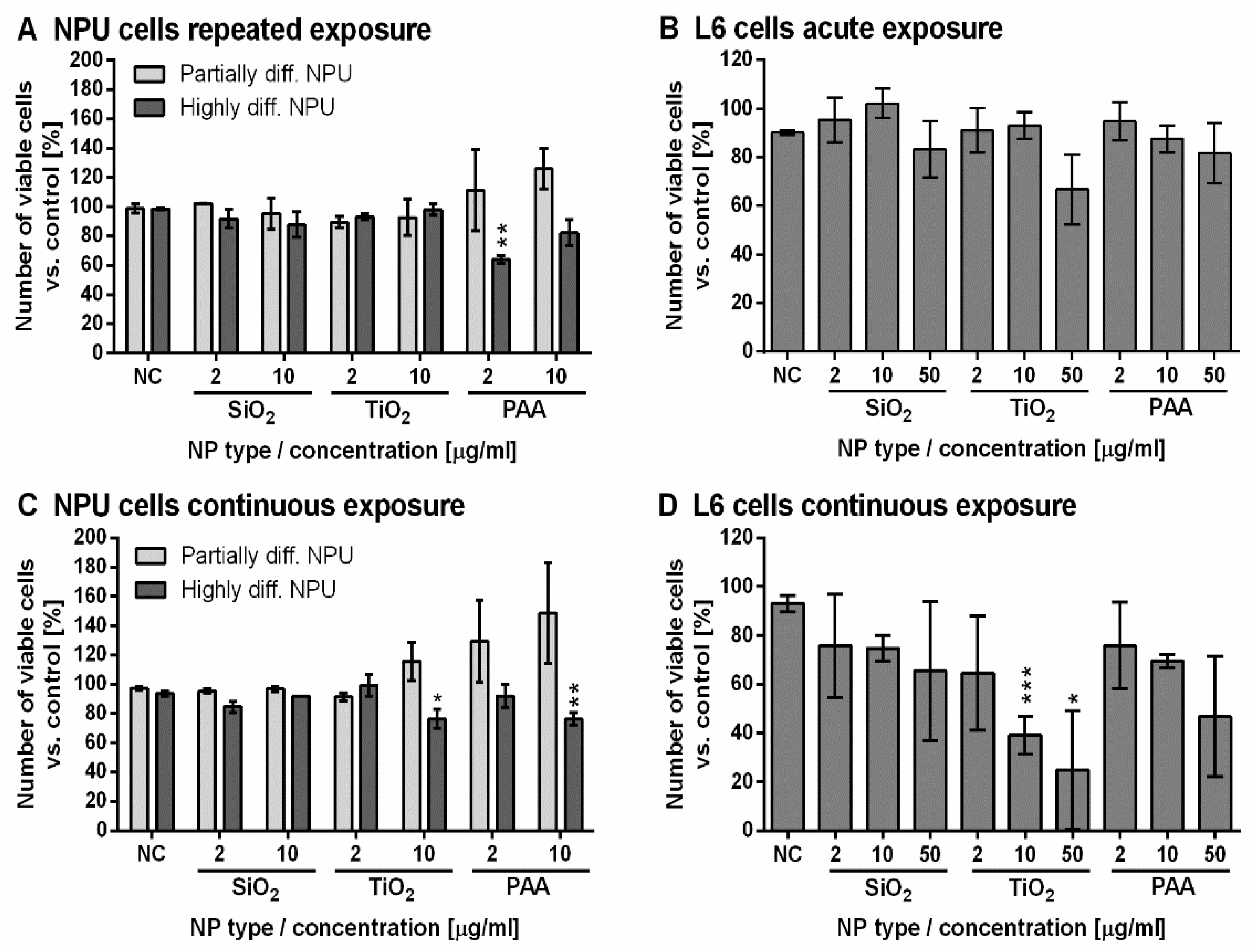
2.4. Long-Term Assessment of NPU and L6 Cell Viability for Exposure to NPs
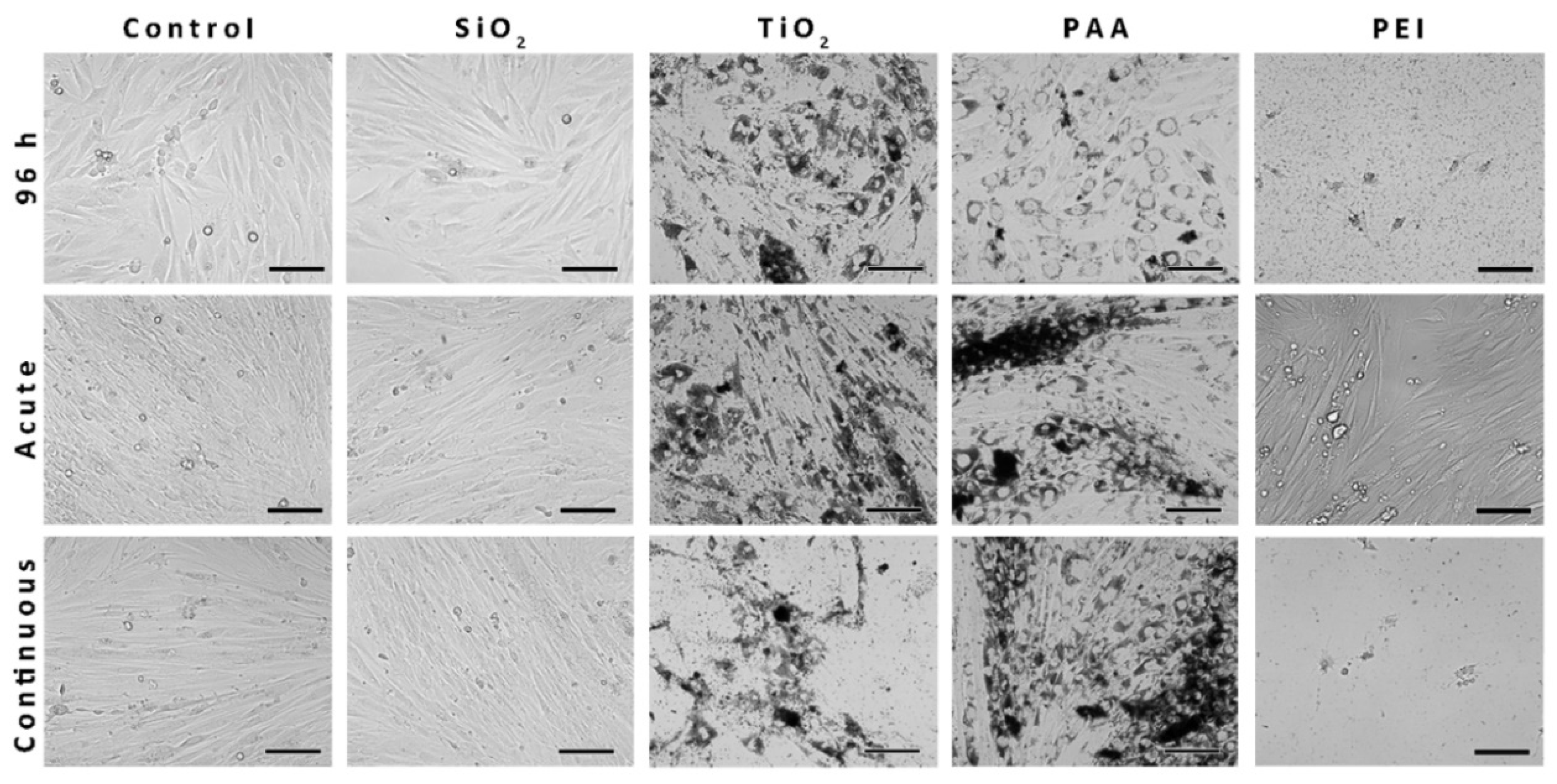
2.5. The Morphology of the L6 Cells Treated with NPs and Analyzed with Phase-Contrast Microscopy
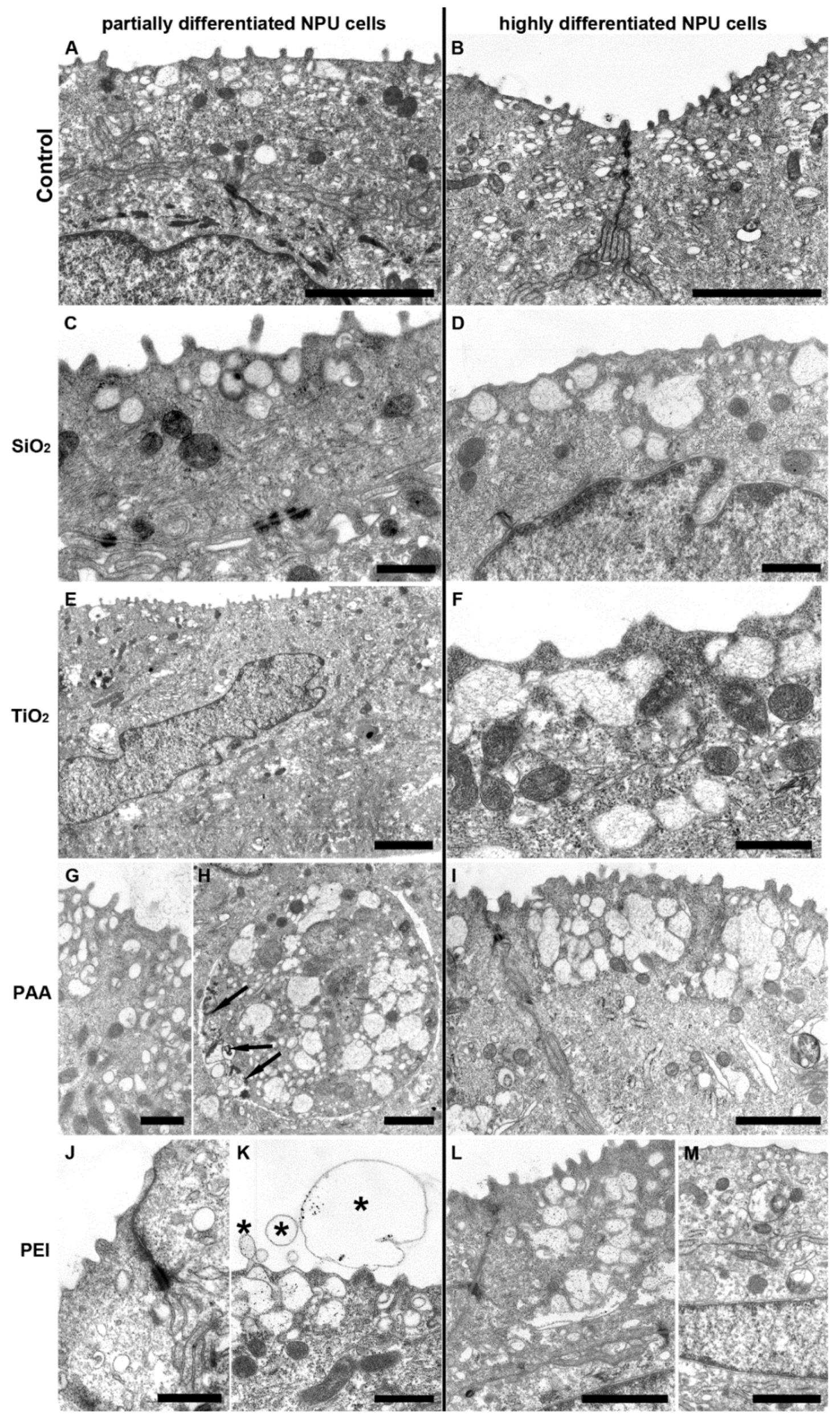
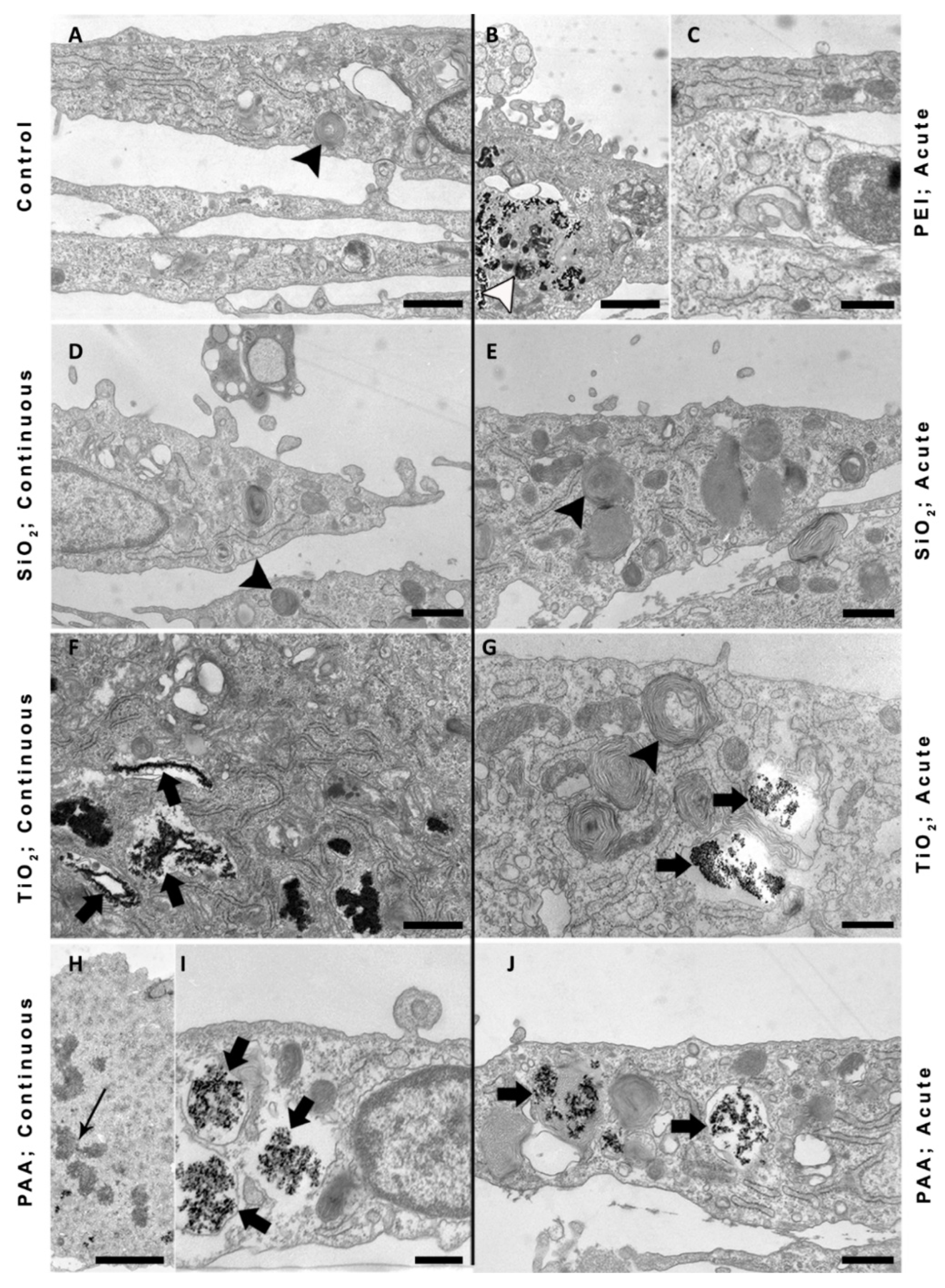
2.6. TEM Images of NPU Cells and L6 cells after Prolonged Exposure to NPs
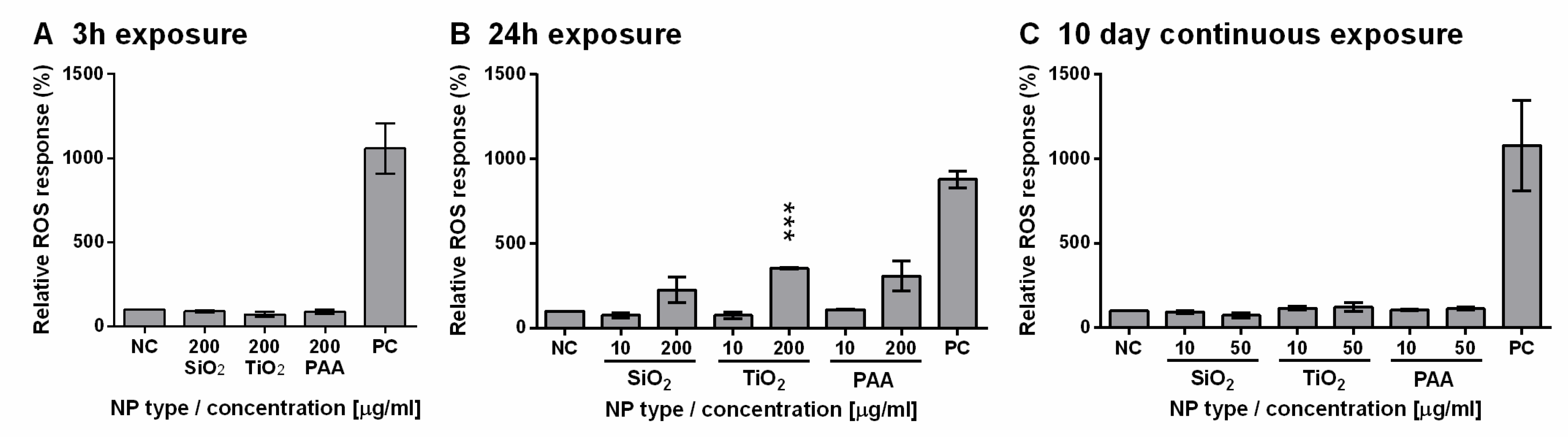
2.7. TiO2, PAA, and SiO2 NPs Induced ROS Formation in L6 Cells in a Dose-Dependent Manner
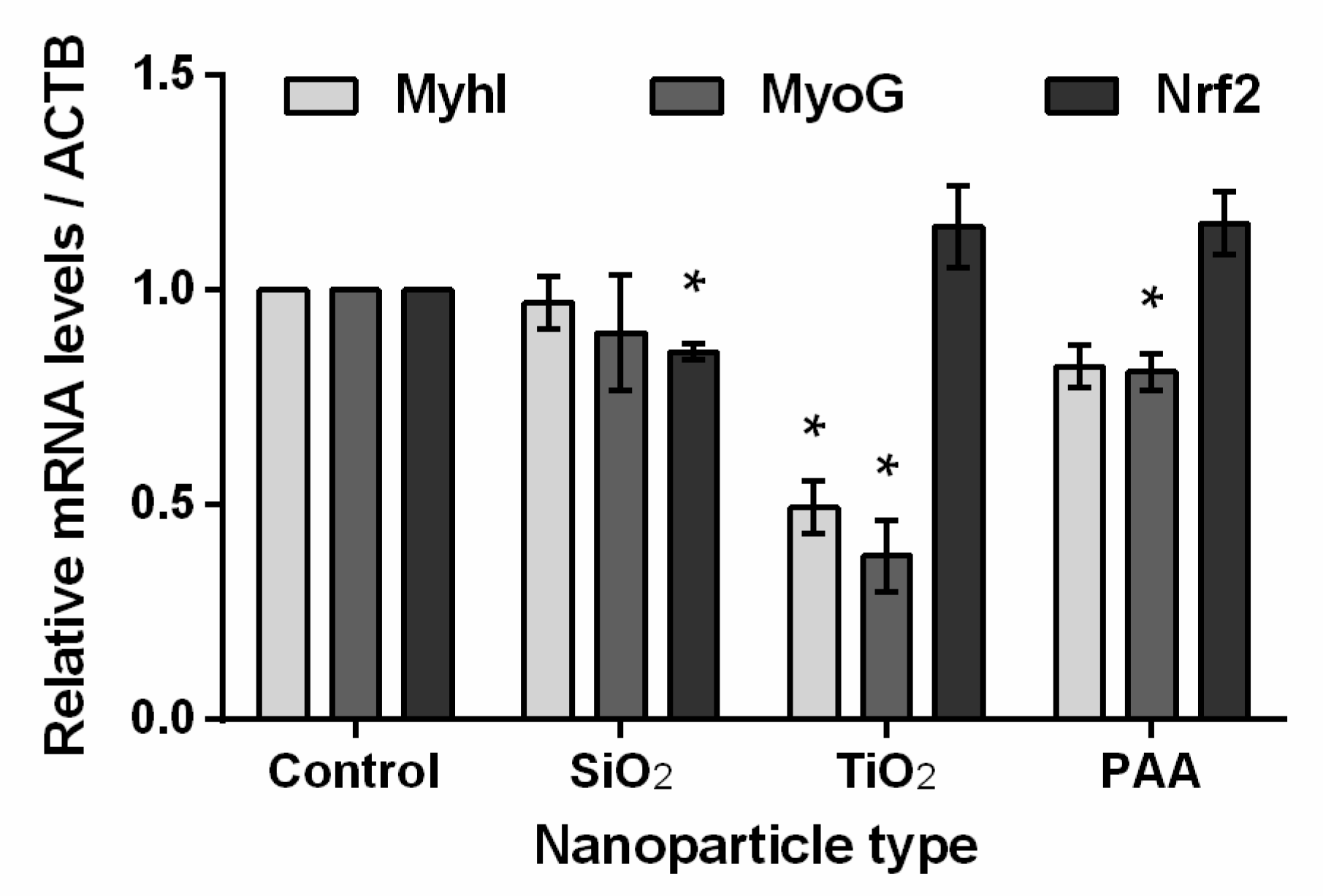
2.8. The Influence of TiO2, PAA, and SiO2 NPs on L6 Cells Differentiation and Oxidative Stress Biomarkers
3. Discussion
4. Materials and Methods
4.1. Nanoparticles Synthesis
4.2. Nanoparticles Characterization
4.3. Cell Models and Cell Culturing
4.4. Short-Term and Long-Term Exposure Protocols
4.5. Cell Viability Assays
4.6. Transmission Electron Microscopy
4.7. Measurements of the Intracellular ROS Levels in Rat L6 Cells
4.8. Quantitative PCR
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NPs | nanoparticles |
| Co–ferrite | cobalt–ferrite |
| PAA | polyacrylic acid |
| PEI | polyethylenimine |
| PI | propidium iodide |
| ROS | reactive oxygen species |
| qPCR | quantitative polymerase chain reaction |
| Nrf2 | nuclear factor (erythroid-derived 2)-like 2 |
| MyoG | myogenin |
| MyhI | myosin heavy chain I |
| NPU | normal porcine urothelial |
References
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; Von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed]
- Kamaly, N.; Xiao, Z.; Valencia, P.M.; Radovic-Moreno, A.F.; Farokhzad, O.C. Targeted polymeric therapeutic nanoparticles: Design, development and clinical translation. Chem. Soc. Rev. 2012, 41, 2971–3010. [Google Scholar] [CrossRef] [PubMed]
- Bregar, V.B.; Pavlin, M. Stability of nanoparticle suspensions in different biologically relevant media. Dig. J. Nanomater. Biostruct. 2012, 4, 1389–1400. [Google Scholar]
- Strojan, K.; Leonardi, A.; Bregar, V.B.; Križaj, I.; Svete, J.; Pavlin, M. Dispersion of nanoparticles in different media importantly determines the composition of their protein corona. PLoS ONE 2017, 12, e0169552. [Google Scholar] [CrossRef]
- Singh, N.; Manshian, B.; Jenkins, G.J.; Griffiths, S.M.; Williams, P.M.; Maffeis, T.G.; Wright, C.J.; Doak, S.H. NanoGenotoxicology: The DNA damaging potential of engineered nanomaterials. Biomaterials 2009, 30, 3891–3914. [Google Scholar] [CrossRef]
- Fröhlich, E. Cellular targets and mechanisms in the cytotoxic action of non-biodegradable engineered nanoparticles. Curr. Drug Metab. 2013, 14, 976–988. [Google Scholar] [CrossRef]
- Zoroddu, M.A.; Medici, S.; Ledda, A.; Nurchi, V.M.; Lachowicz, J.I.; Peana, M. Toxicity of nanoparticles. Curr. Med. Chem. 2014, 21, 3837–3853. [Google Scholar] [CrossRef]
- Lojk, J.; Strojan, K.; Mis, K.; Bregar, B.V.; Hafner-Bratkovič, I.; Bizjak, M.; Pirkmajer, S.; Pavlin, M. Cell stress response to two different types of polymer coated cobalt ferrite nanoparticles. Toxicol. Lett. 2017, 270, 108–118. [Google Scholar] [CrossRef]
- Peeters, P.M.; Eurlings, I.M.J.; Perkins, T.N.; Wouters, E.F.M.; Schins, R.P.F.; Borm, P.; Drommer, W.; Reynaert, N.L.; Albrecht, C. Silica-induced NLRP3 inflammasome activation in vitro and in rat lungs. Part. Fibre Toxicol. 2014, 11, 58. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, L.; Chen, Q.; Chen, C. Cytotoxic potential of silver nanoparticles. Yonsei Med. J. 2014, 55, 283–291. [Google Scholar] [CrossRef]
- Elgrabli, D.; Beaudouin, R.; Jbilou, N.; Floriani, M.; Péry, A.; Rogerieux, F.; Lacroix, G. Biodistribution and clearance of TiO2 nanoparticles in rats after intravenous injection. PLoS ONE 2015, 10, e0124490. [Google Scholar] [CrossRef]
- Almeida, J.P.M.; Chen, A.L.; Foster, A.; Drezek, R. In vivobiodistribution of nanoparticles. Nanomedicine 2011, 6, 815–835. [Google Scholar] [CrossRef] [PubMed]
- Borak, B.; Biernat, P.; Prescha, A.; Baszczuk, A.; Pluta, J. In vivo study on the biodistribution of silica particles in the bodies of rats. Adv. Clin. Exp. Med. 2012, 21, 13–18. [Google Scholar] [PubMed]
- Jain, T.K.; Reddy, M.K.; Morales, M.A.; Leslie-Pelecky, D.L.; Labhasetwar, V. Biodistribution, clearance, and biocompatibility of iron oxide magnetic nanoparticles in rats. Mol. Pharm. 2008, 5, 316–327. [Google Scholar] [CrossRef]
- Heringa, M.B.; Peters, R.J.B.; Bleys, R.L.A.W.; Van Der Lee, M.K.; Tromp, P.C.; Van Kesteren, P.C.E.; Van Eijkeren, J.C.H.; Undas, A.K.; Oomen, A.G.; Bouwmeester, H. Detection of titanium particles in human liver and spleen and possible health implications. Part. Fibre Toxicol. 2018, 15, 15. [Google Scholar] [CrossRef]
- Annangi, B.; Bach, J.; Vales, G.; Rubio, L.; Marcos, R.; Hernández, A. Long-term exposures to low doses of cobalt nanoparticles induce cell transformation enhanced by oxidative damage. Nanotoxicology 2014, 9, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Annangi, B.; Rubio, L.; Alaraby, M.; Bach, J.; Marcos, R.; Hernández, A. Acute and long-term in vitro effects of zinc oxide nanoparticles. Arch. Toxicol. 2015, 90, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Armand, L.; Tarantini, A.; Beal, D.; Biola-Clier, M.; Bobyk, L.; Sorieul, S.; Pernet-Gallay, K.; Marie-Desvergne, C.; Lynch, I.; Herlin-Boime, N.; et al. Long-term exposure of A549 cells to titanium dioxide nanoparticles induces DNA damage and sensitizes cells towards genotoxic agents. Nanotoxicology 2016, 10, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Brun, E.; Carrière, M.; Mabondzo, A. In vitro evidence of dysregulation of blood–brain barrier function after acute and repeated/long-term exposure to TiO2 nanoparticles. Biomaterials 2012, 33, 886–896. [Google Scholar] [CrossRef]
- Chen, N.; Song, Z.M.; Tang, H.; Xi, W.S.; Cao, A.; Liu, Y.; Wang, H. Toxicological effects of caco-2 cells following short-term and long-term exposure to Ag nanoparticles. Int. J. Mol. Sci. 2016, 17, 974. [Google Scholar] [CrossRef]
- Coccini, T.; Manzo, L.; Bellotti, V.; De Simone, U. Assessment of cellular responses after short- and long-term exposure to silver nanoparticles in human neuroblastoma (SH-SY5Y) and astrocytoma (D384) cells. Sci. World J. 2014, 2014, 259765. [Google Scholar] [CrossRef]
- Coccini, T.; Grandi, S.; Lonati, D.; Locatelli, C.; De Simone, U. Comparative cellular toxicity of titanium dioxide nanoparticles on human astrocyte and neuronal cells after acute and prolonged exposure. NeuroToxicology 2015, 48, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Comfort, K.K.; Braydich-Stolle, L.K.; Maurer, E.I.; Hussain, S.M. Less is more: Long-termin vitroexposure to low levels of silver nanoparticles provides new insights for nanomaterial evaluation. ACS Nano 2014, 8, 3260–3271. [Google Scholar] [CrossRef] [PubMed]
- De Marzi, L.; Monaco, A.; De Lapuente, J.; Ramos, D.; Borras, M.; Di Gioacchino, M.; Santucci, S.; Poma, A. Cytotoxicity and genotoxicity of ceria nanoparticles on different cell lines in vitro. Int. J. Mol. Sci. 2013, 14, 3065–3077. [Google Scholar] [CrossRef] [PubMed]
- De Simone, U.; Manzo, L.; Profumo, A.; Coccini, T. In Vitro Toxicity Evaluation of Engineered Cadmium-Coated Silica Nanoparticles on Human Pulmonary Cells. Hindawi Publishig J. Toxicol. 2013, 931785. [Google Scholar] [CrossRef]
- Huang, S.; Chueh, P.J.; Lin, Y.W.; Shih, T.S.; Chuang, S.M. Disturbed mitotic progression and genome segregation are involved in cell transformation mediated by nano-TiO2 long-term exposure. Toxicol. Appl. Pharmacol. 2009, 241, 182–194. [Google Scholar] [CrossRef]
- Jacobsen, N.R.; Saber, A.T.; White, P.A.; Møller, P.; Pojana, G.; Vogel, U.; Loft, S.; Gingerich, J.; Soper, L.; Douglas, G.R.; et al. Increased mutant frequency by carbon black, but not quartz, in thelacZ andcII transgenes of muta™ mouse lung epithelial cells. Environ. Mol. Mutagen. 2007, 48, 451–461. [Google Scholar] [CrossRef]
- Kocbek, P.; Teskač, K.; Kreft, M.E.; Kristl, J. Toxicological aspects of long-term treatment of keratinocytes with ZnO and TiO2 nanoparticles. Small 2010, 6, 1908–1917. [Google Scholar] [CrossRef]
- McCracken, C.; Zane, A.; Knight, D.A.; Dutta, P.; Waldman, W.J. Minimal intestinal epithelial cell toxicity in response to short- and long-term food-relevant inorganic nanoparticle exposure. Chem. Res. Toxicol. 2013, 26, 1514–1525. [Google Scholar] [CrossRef]
- Medina-Reyes, E.I.; Déciga-Alcaraz, A.; Freyre-Fonseca, V.; Buenrostro, N.L.D.; Flores-Flores, J.O.; Gutierrez-Lopez, G.F.; Sánchez-Pérez, Y.; García-Cuellar, C.M.; Pedraza-Chaverri, J.; Chirino, Y.I. Titanium dioxide nanoparticles induce an adaptive inflammatory response and invasion and proliferation of lung epithelial cells in chorioallantoic membrane. Environ. Res. 2015, 136, 424–434. [Google Scholar] [CrossRef]
- Mrakovcic, M.; Absenger, M.; Riedl, R.; Smole, C.; Roblegg, E.; Fröhlich, L.F.; Fröhlich, E. Assessment of long-term effects of nanoparticles in a microcarrier cell culture system. PLoS ONE 2013, 8, e56791. [Google Scholar] [CrossRef]
- Mrakovcic, M.; Meindl, C.; Roblegg, E.; Fröhlich, E. Reaction of monocytes to polystyrene and silica nanoparticles in short-term and long-term exposures. Toxicol. Res. 2014, 3, 86–97. [Google Scholar] [CrossRef]
- Quignard, S.; Mosser, G.; Boissière, M.; Coradin, T. Long-term fate of silica nanoparticles interacting with human dermal fibroblasts. Biomaterials 2012, 33, 4431–4442. [Google Scholar] [CrossRef] [PubMed]
- Thurnherr, T.; Brandenberger, C.; Fischer, K.; Diener, L.; Manser, P.; Maeder-Althaus, X.; Kaiser, J.P.; Krug, H.F.; Rothen-Rutishauser, B.; Wick, P. A comparison of acute and long-term effects of industrial multiwalled carbon nanotubes on human lung and immune cells in vitro. Toxicol. Lett. 2011, 200, 176–186. [Google Scholar] [CrossRef]
- Tsai, S.W.; Liaw, J.W.; Kao, Y.C.; Huang, M.Y.; Lee, C.Y.; Rau, L.R.; Huang, C.Y.; Wei, K.C.; Ye, T.C. Internalized gold nanoparticles do not affect the osteogenesis and apoptosis of MG63 osteoblast-like Cells: A quantitative, in vitro study. PLoS ONE 2013, 8, e76545. [Google Scholar] [CrossRef]
- Vales, G.; Rubió, L.; Marcos, R. Long-term exposures to low doses of titanium dioxide nanoparticles induce cell transformation, but not genotoxic damage in BEAS-2B cells. Nanotoxicology 2014, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luanpitpong, S.; Castranova, V.; Tse, W.; Lu, Y.; Pongrakhananon, V.; Rojanasakul, Y. Carbon nanotubes induce malignant transformation and tumorigenesis of human lung epithelial cells. Nano Lett. 2011, 11, 2796–2803. [Google Scholar] [CrossRef]
- Wang, S.; Hunter, L.A.; Arslan, Z.; Wilkerson, M.G.; Wickliffe, J.K. Chronic exposure to nanosized, anatase titanium dioxide is not cyto- or genotoxic to Chinese hamster ovary cells. Environ. Mol. Mutagen. 2011, 52, 614–622. [Google Scholar] [CrossRef]
- Kreft, M.E.; Hudoklin, S.; Jezernik, K.; Romih, R. Formation and maintenance of blood–urine barrier in urothelium. Protoplasma 2010, 246, 3–14. [Google Scholar] [CrossRef]
- Tratnjek, L.; Romih, R.; Kreft, M.E. Differentiation-dependent rearrangements of actin filaments and microtubules hinder apical endocytosis in urothelial cells. Histochem. Cell Biol. 2017, 148, 143–156. [Google Scholar] [CrossRef]
- Višnjar, T.; Kreft, M.E. Air–liquid and liquid–liquid interfaces influence the formation of the urothelial permeability barrier in vitro. Vitr. Cell. Dev. Biol. Anim. 2013, 49, 196–204. [Google Scholar] [CrossRef]
- Višnjar, T.; Kreft, M.E. The complete functional recovery of chitosan-treated biomimetic hyperplastic and normoplastic urothelial models. Histochem. Cell Biol. 2014, 143, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Višnjar, T.; Kocbek, P.; Kreft, M.E. Hyperplasia as a mechanism for rapid resealing urothelial injuries and maintaining high transepithelial resistance. Histochem. Cell Biol. 2012, 137, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed]
- Gandaglia, G.; Briganti, A.; Gontero, P.; Mondaini, N.; Novara, G.; Salonia, A.; Sciarra, A.; Montorsi, F. The role of chronic prostatic inflammation in the pathogenesis and progression of benign prostatic hyperplasia (BPH). BJU Int. 2013, 112, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Nakamura, M.; Sakamoto, W.; Yogo, T.; Miki, H.; Ozaki, S.; Abe, M.; Matsumoto, T.; Ishimura, K. Superparamagnetic nanoparticle clusters for cancer theranostics combining magnetic resonance imaging and hyperthermia treatment. Theranostics 2013, 3, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Raftis, J.; Miller, M.; Langrish, J.; Krystek, P.; Campbell, C.; Donaldon, K.; Cassee, F.; Newby, D.; Mills, N.L.; Duffin, R. Gold nanoparticles translocate from the lung into the blood in man and accumulate at sites of vascular inflammation in apolipoproteinE knockout mice. Eur. Respir. J. 2015, 46, PA4117. [Google Scholar] [CrossRef]
- Briolay, A.; Jaafar, R.; Némoz, G.; Bessueille, L. Myogenic differentiation and lipid-raft composition of L6 skeletal muscle cells are modulated by PUFAs. Biochim. Biophys. Acta 2013, 1828, 602–613. [Google Scholar] [CrossRef]
- Van Teunenbroek, T.; Baker, J.; Dijkzeul, A. Towards a more effective and efficient governance and regulation of nanomaterials. Part. Fibre Toxicol. 2017, 14, 54. [Google Scholar] [CrossRef]
- Iavicoli, I.; Leso, V.; Fontana, L.; Bergamaschi, A. Toxicological effects of titanium dioxide nanoparticles: A review of in vitro mammalian studies. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 481–508. [Google Scholar]
- Skocaj, M.; Filipic, M.; Petkovic, J.; Novak, S. Titanium dioxide in our everyday life; is it safe? Radiol. Oncol. 2011, 45, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Oberdörster, G. Safety assessment for nanotechnology and nanomedicine: Concepts of nanotoxicology. J. Intern. Med. 2010, 267, 89–105. [Google Scholar] [CrossRef]
- Report of the Workshop on Risk Assesment of Manufactured Nanomaterials in a Regulatory Context No. 21—ENV/JM/MONO(2010)10; OECD-WPMN: Washington, DC, USA, 2009.
- Risk Assessment Of Products Of Nanotechnologies; Scientific Committee on Emerging and Newly Identified Health Risks SCHENIR: Brussels, Belgium, 2009.
- The Appropriateness of Existing Methodologies to Assess the Potential Risks Associated with Engineered and Adventitious Products of Nanotechnologies; Scientific Committee on Emerging and Newly Identified Health Risks SCHENIR: Brussels, Belgium, 2006.
- Pavlin, M.; Lojk, J.; Bregar, V.B.; Rajh, M.; Mis, K.; Kreft, M.E.; Pirkmajer, S.; Veranič, P. Cell type-specific response to high intracellular loading of polyacrylic acid-coated magnetic nanoparticles. Int. J. Nanomed. 2015, 10, 1449–1462. [Google Scholar] [CrossRef]
- Shin, S.W.; Song, I.H.; Um, S.H. Role of physicochemical properties in nanoparticle toxicity. Nanomaterials 2015, 5, 1351–1365. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Symonds, P.; Murray, J.C.; Hunter, A.C.; Debska, G.; Szewczyk, A. A two-stage poly(ethylenimine)-mediated cytotoxicity: Implications for gene transfer/therapy. Mol. Ther. 2005, 11, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.H.; Bae, H.C.; Jang, Y.; Jeong, S.H.; Na Lee, H.; Ryu, W.I.; Yoo, M.G.; Kim, Y.R.; Kim, M.K.; Lee, J.K.; et al. Effect of the size and surface charge of silica nanoparticles on cutaneous toxicity. Mol. Cell. Toxicol. 2013, 9, 67–74. [Google Scholar] [CrossRef]
- Lojk, J.; Čibej, U.; Karlaš, D.; Šajn, L.; Pavlin, M. Comparison of two automatic cell-counting solutions for fluorescent microscopic images. J. Microsc. 2015, 260, 107–116. [Google Scholar] [CrossRef]
- Prijic, S.; Prosen, L.; Cemazar, M.; Scancar, J.; Romih, R.; Lavrenčak, J.; Bregar, V.B.; Coer, A.; Krzan, M.; Žnidaršič, A.; et al. Surface modified magnetic nanoparticles for immuno-gene therapy of murine mammary adenocarcinoma. Biomaterials 2012, 33, 4379–4391. [Google Scholar] [CrossRef]
- Strojan, K.; Lojk, J.; Bregar, V.B.; Veranič, P.; Pavlin, M. Glutathione reduces cytotoxicity of polyethyleneimine coated magnetic nanoparticles in CHO cells. Toxicol. In Vitro 2017, 41, 12–20. [Google Scholar] [CrossRef]
- Hotze, E.M.; Phenrat, T.; Lowry, G.V. Nanoparticle aggregation: Challenges to understanding transport and reactivity in the environment. J. Environ. Qual. 2010, 39, 1909–1924. [Google Scholar] [CrossRef]
- Khandelwal, P.; Abraham, S.N.; Apodaca, G. Cell biology and physiology of the uroepithelium. Am. J. Physiol. Physiol. 2009, 297, F1477–F1501. [Google Scholar] [CrossRef]
- Lasič, E.; Višnjar, T.; Kreft, M.E. Properties of the urothelium that establish the blood–urine barrier and their implications for drug delivery. In Reviews of Physiology, Biochemistry and Pharmacology; Springer: Cham, Switzerland, 2015; Volume 168, pp. 1–29. [Google Scholar] [CrossRef]
- Lojk, J.; Bregar, V.B.; Strojan, K.; Hudoklin, S.; Veranič, P.; Pavlin, M.; Kreft, M.E. Increased endocytosis of magnetic nanoparticles into cancerous urothelial cells versus normal urothelial cells. Histochem. Cell Biol. 2017, 149, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Pavlin, M.; Bregar, V.B.; Lojk, J.; Šuštar, V.; Veranič, P. Visualization of internalization of functionalized cobalt ferrite nanoparticles and their intracellular fate. Int. J. Nanomed. 2013, 8, 919–931. [Google Scholar] [CrossRef]
- Ambrosone, A.; Di Vettimo, M.R.S.; Malvindi, M.A.; Roopin, M.; Levy, O.; Marchesano, V.; Pompa, P.P.; Tortiglione, C.; Tino, A. Impact of amorphous SiO2 nanoparticles on a living organism: Morphological, behavioral, and molecular biology implications. Front. Bioeng. Biotechnol. 2014, 2, 37. [Google Scholar] [CrossRef] [PubMed]
- Gazzano, E.; Ghiazza, M.; Polimeni, M.; Bolis, V.; Fenoglio, I.; Attanasio, A.; Mazzucco, G.; Fubini, B.; Ghigo, D. Physicochemical determinants in the cellular responses to nanostructured amorphous silicas. Toxicol. Sci. 2012, 128, 158–170. [Google Scholar] [CrossRef]
- Rabolli, V.; Thomassen, L.C.J.; Princen, C.; Napierska, D.; Gonzalez, L.; Kirsch-Volders, M.; Hoet, P.H.; Huaux, F.; Kirschhock, C.E.A.; Martens, J.A.; et al. Influence of size, surface area and microporosity on thein vitrocytotoxic activity of amorphous silica nanoparticles in different cell types. Nanotoxicology 2010, 4, 307–318. [Google Scholar] [CrossRef]
- Chang, J.S.; Chang, K.L.B.; Hwang, D.F.; Kong, Z.L. In vitro cytotoxicitiy of silica nanoparticles at high concentrations strongly depends on the metabolic activity type of the cell line. Environ. Sci. Technol. 2007, 41, 2064–2068. [Google Scholar] [CrossRef]
- Chung, T.H.; Wu, S.H.; Yao, M.; Lu, C.W.; Lin, Y.S.; Hung, Y.; Mou, C.Y.; Chen, Y.C.; Huang, D.M. The effect of surface charge on the uptake and biological function of mesoporous silica nanoparticles in 3T3-L1 cells and human mesenchymal stem cells. Biomaterials 2007, 28, 2959–2966. [Google Scholar] [CrossRef]
- Imani, R.; Veranič, P.; Iglič, A.; Kreft, M.E.; Pazoki, M.; Hudoklin, S. Combined cytotoxic effect of UV-irradiation and TiO2 microbeads in normal urothelial cells, low-grade and high-grade urothelial cancer cells. Photochem. Photobiol. Sci. 2015, 14, 583–590. [Google Scholar] [CrossRef][Green Version]
- Nel, A.E.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef]
- Li, N.; Xia, T.; Nel, A.E. The role of oxidative stress in ambient particulate matter-induced lung diseases and its implications in the toxicity of engineered nanoparticles. Free. Radic. Biol. Med. 2008, 44, 1689–1699. [Google Scholar] [CrossRef] [PubMed]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of nanoparticle-induced oxidative stress and toxicity. BioMed Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef] [PubMed]
- Yoo, K.C.; Yoon, C.H.; Kwon, D.; Hyun, K.H.; Woo, S.J.; Kim, R.K.; Lim, E.J.; Suh, Y.; Kim, M.J.; Yoon, T.H.; et al. Titanium dioxide induces apoptotic cell death through reactive oxygen species-mediated Fas upregulation and Bax activation. Int. J. Nanomed. 2012, 7, 1203–1214. [Google Scholar] [CrossRef]
- Saquib, Q.; Al-Khedhairy, A.A.; Siddiqui, M.A.; Abou-Tarboush, F.M.; Azam, A.; Musarrat, J. Titanium dioxide nanoparticles induced cytotoxicity, oxidative stress and DNA damage in human amnion epithelial (WISH) cells. Toxicol. In Vitro 2012, 26, 351–361. [Google Scholar] [CrossRef]
- Ramkumar, K.M.; Manjula, C.; Gnanakumar, G.; Kanjwal, M.A.; Sekar, T.V.; Paulmurugan, R.; Rajaguru, P. Oxidative stress-mediated cytotoxicity and apoptosis induction by TiO2 nanofibers in HeLa cells. Eur. J. Pharm. Biopharm. 2012, 81, 324–333. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, J.; Xu, J.; Sun, L.; Chen, M.; Lan, M. Nano-SiO2 induces apoptosis via activation of p53 and Bax mediated by oxidative stress in human hepatic cell line. Toxicol. In Vitro 2010, 24, 751–758. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, J.; Chen, M.; Sun, L.; Lan, M. In vitro toxicity of silica nanoparticles in myocardial cells. Environ. Toxicol. Pharmacol. 2010, 29, 131–137. [Google Scholar] [CrossRef]
- Wang, F.; Gao, F.; Lan, M.; Yuan, H.; Huang, Y.; Liu, J. Oxidative stress contributes to silica nanoparticle-induced cytotoxicity in human embryonic kidney cells. Toxicol. In Vitro 2009, 23, 808–815. [Google Scholar] [CrossRef]
- Sun, J.; Chen, Q.; Xue, Y. Kupffer cell-mediated hepatic injury induced by silica nanoparticles in vitro and in vivo. Int. J. Nanomed. 2013, 8, 1129–1140. [Google Scholar] [CrossRef]
- Naqvi, S.; Samim, M.; Abdin, M.; Ahmed, F.J.; Maitra, A.; Prashant, C.; Dinda, A.K. Concentration-dependent toxicity of iron oxide nanoparticles mediated by increased oxidative stress. Int. J. Nanomed. 2010, 5, 983–989. [Google Scholar] [CrossRef]
- Keenan, C.R.; Goth-Goldstein, R.; Lucas, N.; Sedlak, D.L. Oxidative stress induced by zero-valent iron nanoparticles and Fe(II) in human bronchial epithelial cells. Environ. Sci. Technol. 2009, 43, 4555–4560. [Google Scholar] [CrossRef] [PubMed]
- Stern-Straeter, J.; Bonaterra, G.A.; Kassner, S.S.; Zügel, S.; Hormann, K.; Kinscherf, R.; Goessler, U.R. Characterization of human myoblast differentiation for tissue-engineering purposes by quantitative gene expression analysis. J. Tissue Eng. Regen. Med. 2011, 5, e197–e206. [Google Scholar] [CrossRef] [PubMed]
- Rivolta, I.; Panariti, A.; Miserocchi, G. The effect of nanoparticle uptake on cellular behavior: Disrupting or enabling functions? Nanotechnol. Sci. Appl. 2012, 5, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.W.; Zhang, J.; Tew, K.D. Oxidative stress, redox regulation and diseases of cellular differentiation. Biochim. Biophys. Acta 2015, 1850, 1607–1621. [Google Scholar] [CrossRef]
- Tourinho, F.A.; Franck, R. Aqueous ferrofluids based on manganese and cobalt ferrites. J. Mater. Sci. 1990, 25, 3249–3254. [Google Scholar] [CrossRef]
- Jerman, U.D.; Kolenc, M.; Steyer, A.; Veranič, P.; Poljšak-Prijatelj, M.; Kreft, M.E. A novel strain of porcine adenovirus detected in urinary bladder urothelial cell culture. Viruses 2014, 6, 2505–2518. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NP Type | Dispersion Media | Z-average [nm] | Number-Based Hydrodynamic Diameter [nm] | PDI | Zeta Potential [mV] |
|---|---|---|---|---|---|
| PAA | distilled water | 138 ± 48 | 64 ± 22 | 0.3± 0.1 | −56.3 ± 6 |
| MEM alpha + 2% FCS | 573 ± 333 | 62 ± 31 | 0.5 ± 0.0 | −25 ± 2 | |
| * UroM (+Ca2+ − SFBS) | 1637 ± 461 | 239 ± 59 | 0.4 ± 0.0 | −22.2 ± 1 | |
| PEI | distilled water | 136 ± 25 | 104 ± 49 | 0.2 ± 0.1 | 54.4 ± 4 |
| * MEM alpha + 2% FCS | 1352 ± 273 | 1075 ± 337 | 0.3 ± 0.1 | −3 ± 1 | |
| * UroM (+Ca2+ − SFBS) | 1308 ± 39 | 1071 ± 327 | 0.3 ± 0.1 | 5 ± 2 | |
| SiO2 ** | distilled water | 515 ± 206 | 238 ± 70 | .6 ± 0.2 | 2.4 ± 2 |
| MEM alpha + 2% FCS | 145 ± 22 | 47 ± 2 | 0.4 ± 0.1 | −9 ± 5 | |
| UroM (+Ca2+ − SFBS) | 2594 ± 303 | 88 ± 118 | 0.4 ± 0.1 | NA | |
| TiO2 | distilled water | 604 ± 300 | 201 ± 34 | 0.3 ± 0.1 | 28 ± 3 |
| * MEM alpha + 2% FCS | 697 ± 244 | 240 ± 68 | 0.3 ± 0.1 | −5.0 ± 5 | |
| * UroM (+Ca2+ − SFBS) | 1108 ± 559 | 293 ± 141 | 0.3 ± 0.0 | −8.6 ± 0.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skočaj, M.; Bizjak, M.; Strojan, K.; Lojk, J.; Erdani Kreft, M.; Miš, K.; Pirkmajer, S.; Bregar, V.B.; Veranič, P.; Pavlin, M. Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles. Int. J. Mol. Sci. 2020, 21, 7545. https://doi.org/10.3390/ijms21207545
Skočaj M, Bizjak M, Strojan K, Lojk J, Erdani Kreft M, Miš K, Pirkmajer S, Bregar VB, Veranič P, Pavlin M. Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles. International Journal of Molecular Sciences. 2020; 21(20):7545. https://doi.org/10.3390/ijms21207545
Chicago/Turabian StyleSkočaj, Matej, Maruša Bizjak, Klemen Strojan, Jasna Lojk, Mateja Erdani Kreft, Katarina Miš, Sergej Pirkmajer, Vladimir Boštjan Bregar, Peter Veranič, and Mojca Pavlin. 2020. "Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles" International Journal of Molecular Sciences 21, no. 20: 7545. https://doi.org/10.3390/ijms21207545
APA StyleSkočaj, M., Bizjak, M., Strojan, K., Lojk, J., Erdani Kreft, M., Miš, K., Pirkmajer, S., Bregar, V. B., Veranič, P., & Pavlin, M. (2020). Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles. International Journal of Molecular Sciences, 21(20), 7545. https://doi.org/10.3390/ijms21207545