CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage



Abstract
1. Introduction
2. Results
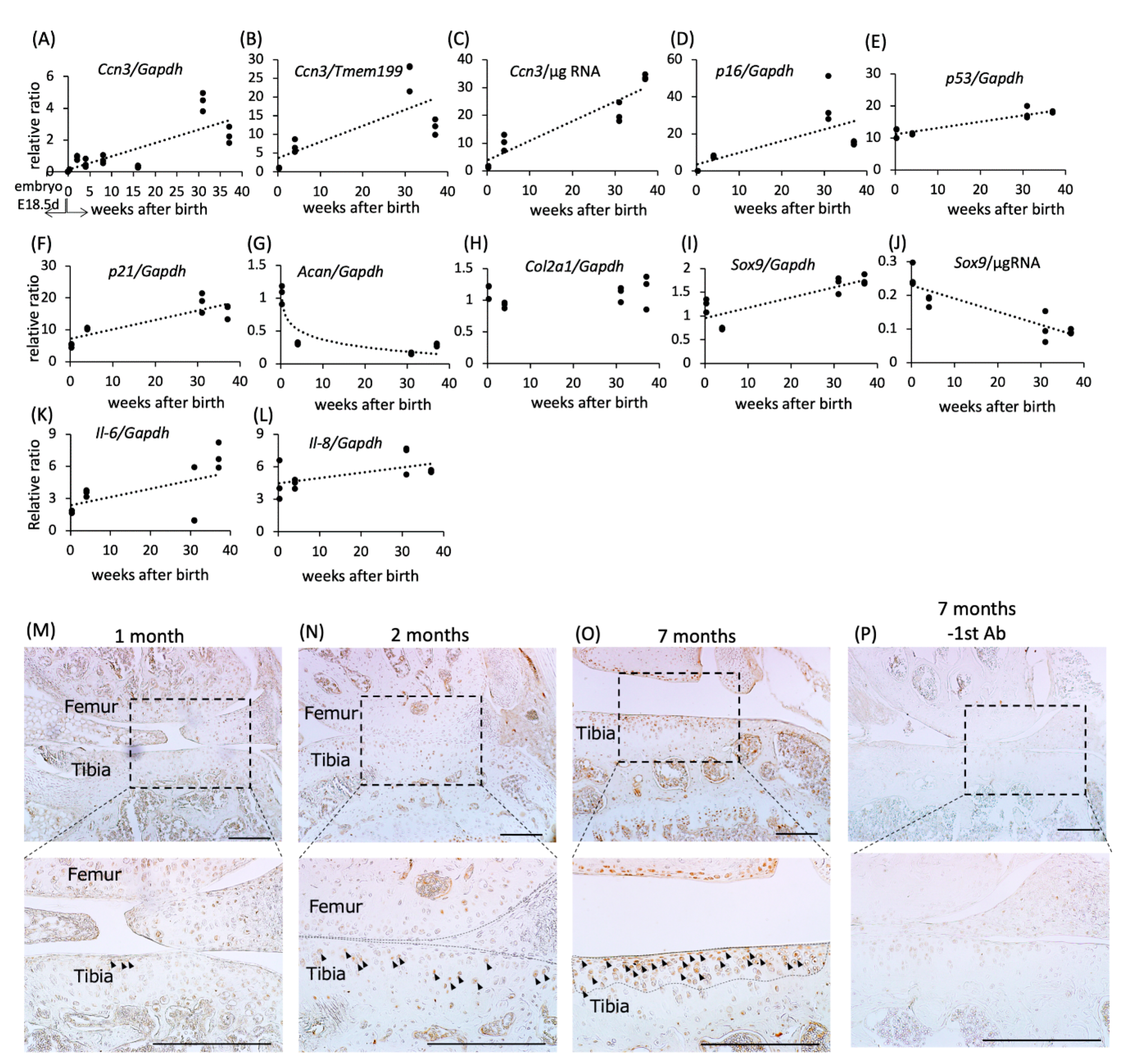
2.1. Increased Expression and Accumulation of CCN3 in Mouse Articular Cartilage with Aging
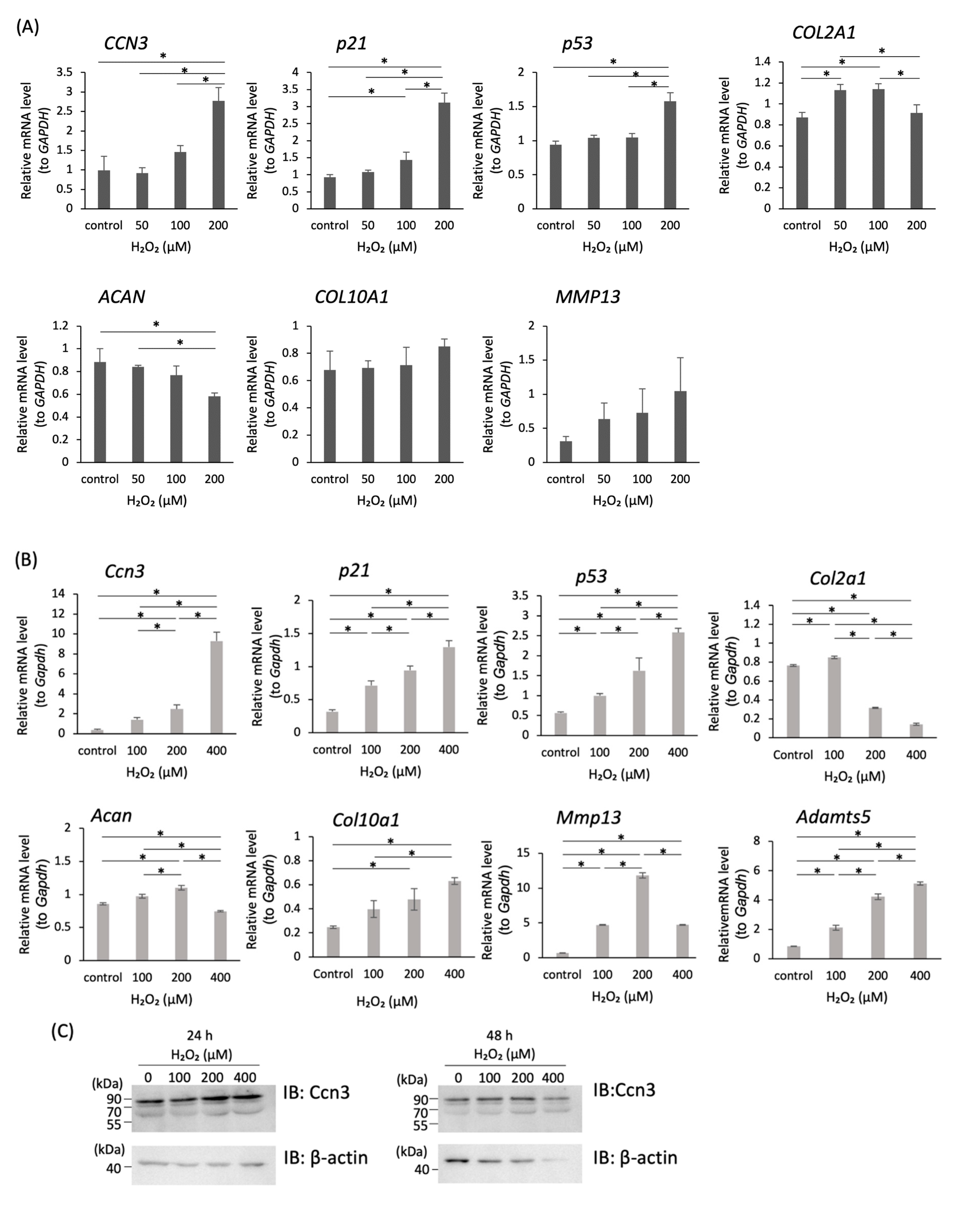
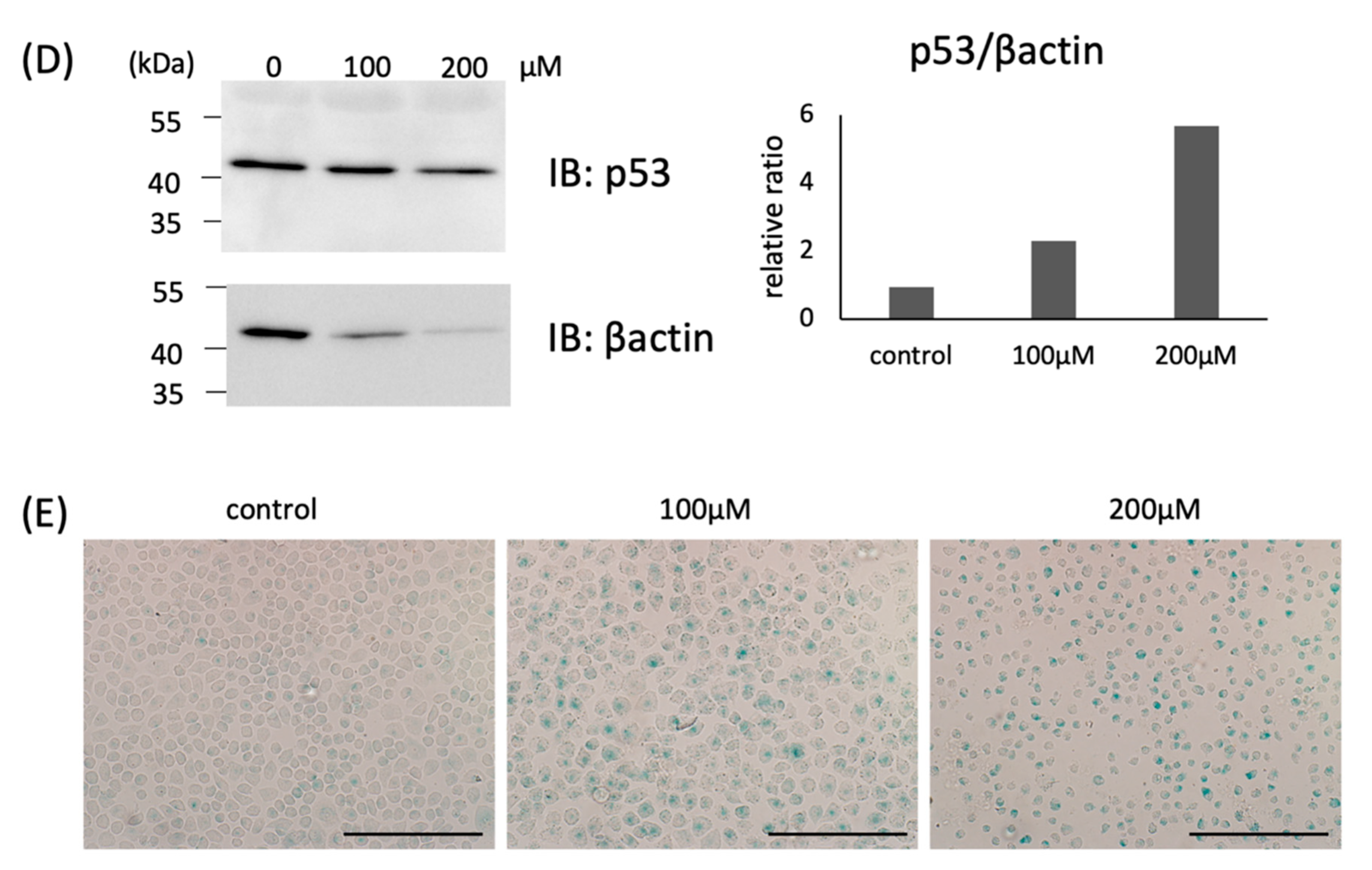
2.2. Increased Expression of CCN3 in H2O2-Treated Chondrocytes
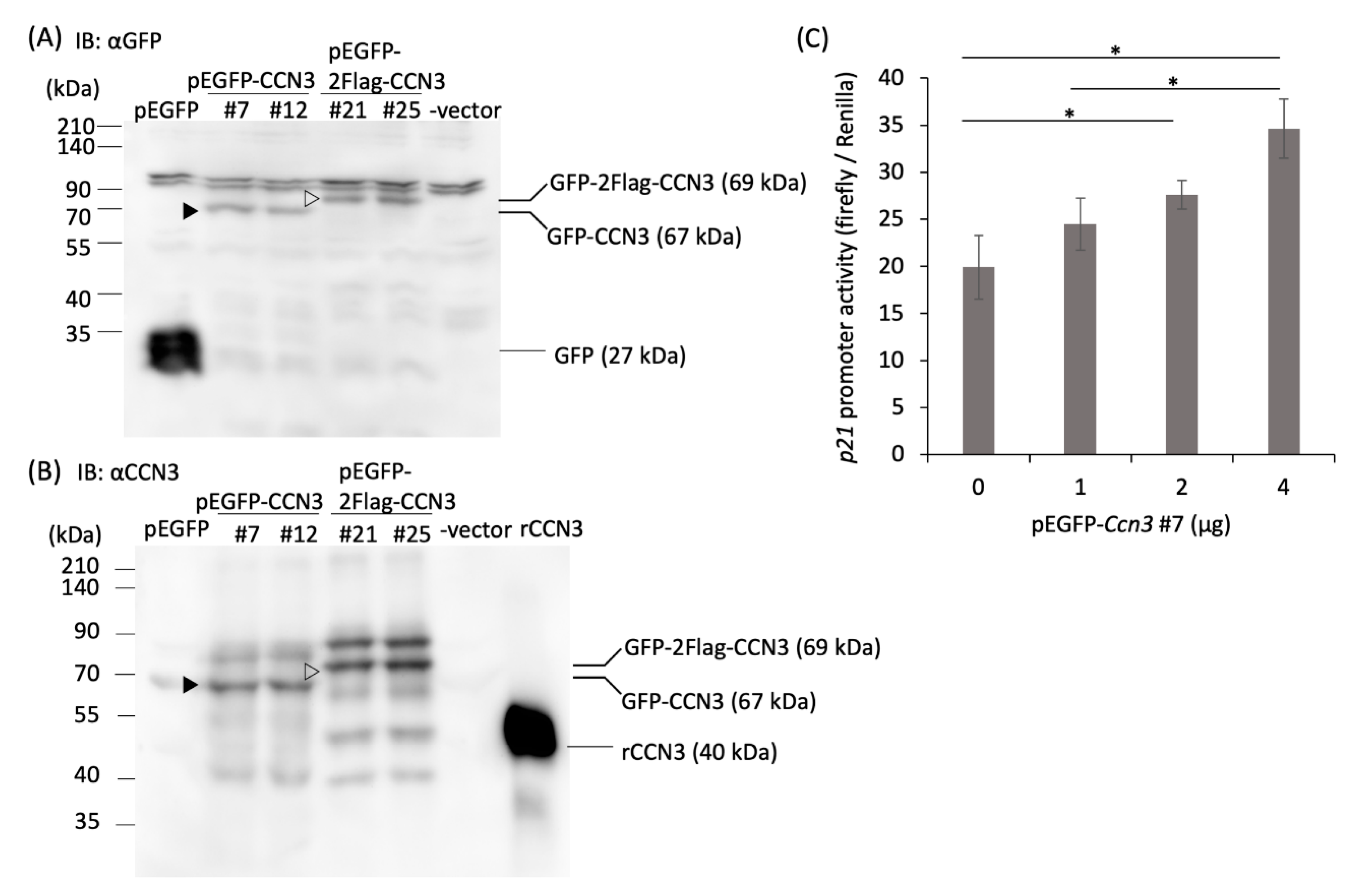
2.3. Overexpression of CCN3 Stimulates p21 Promoter in RCS Cells
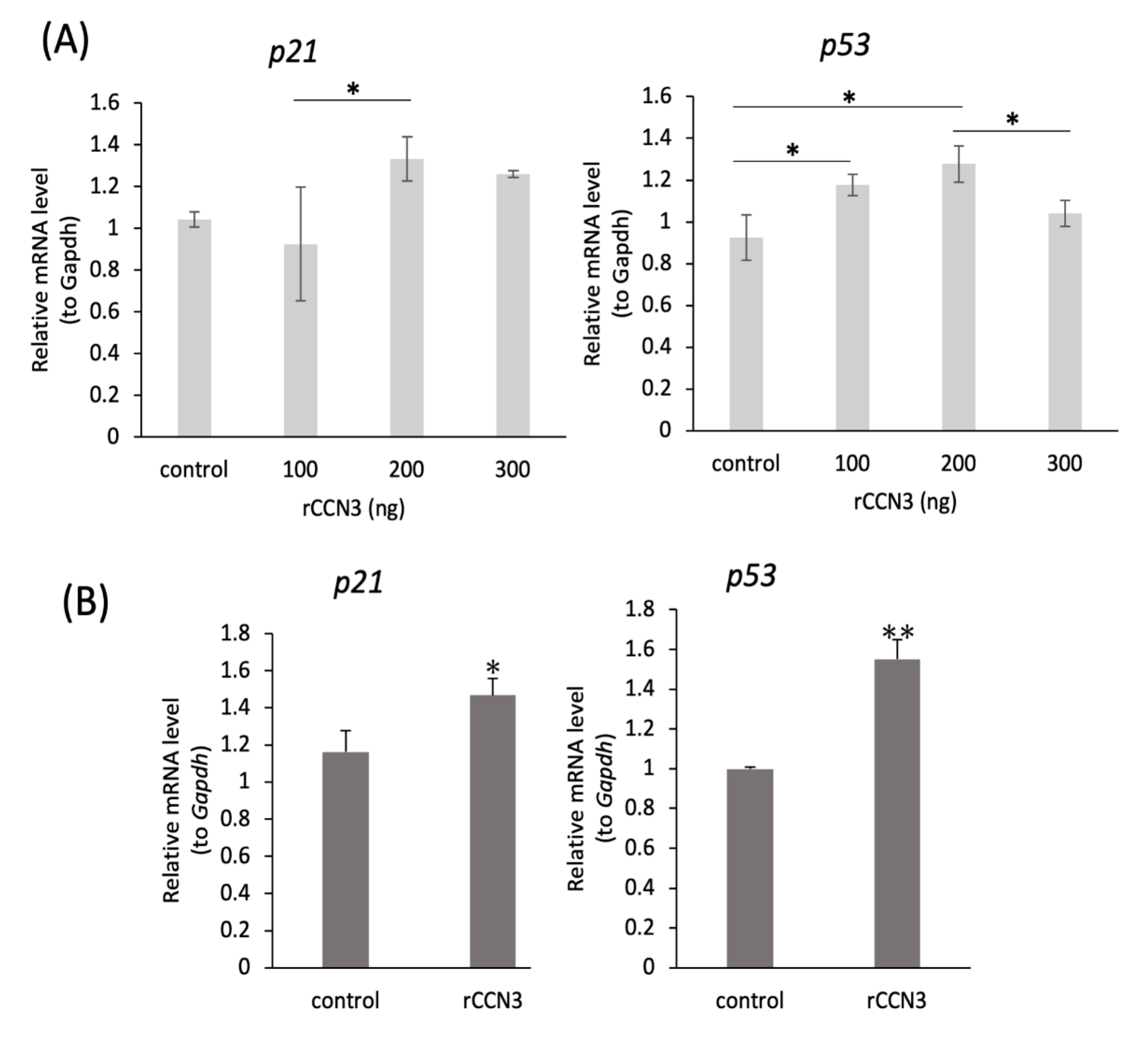
2.4. Treatment with CCN3 Stimulates p21, p53 mRNA Expression in Mouse Primary Chondrocytes and RCS Cells
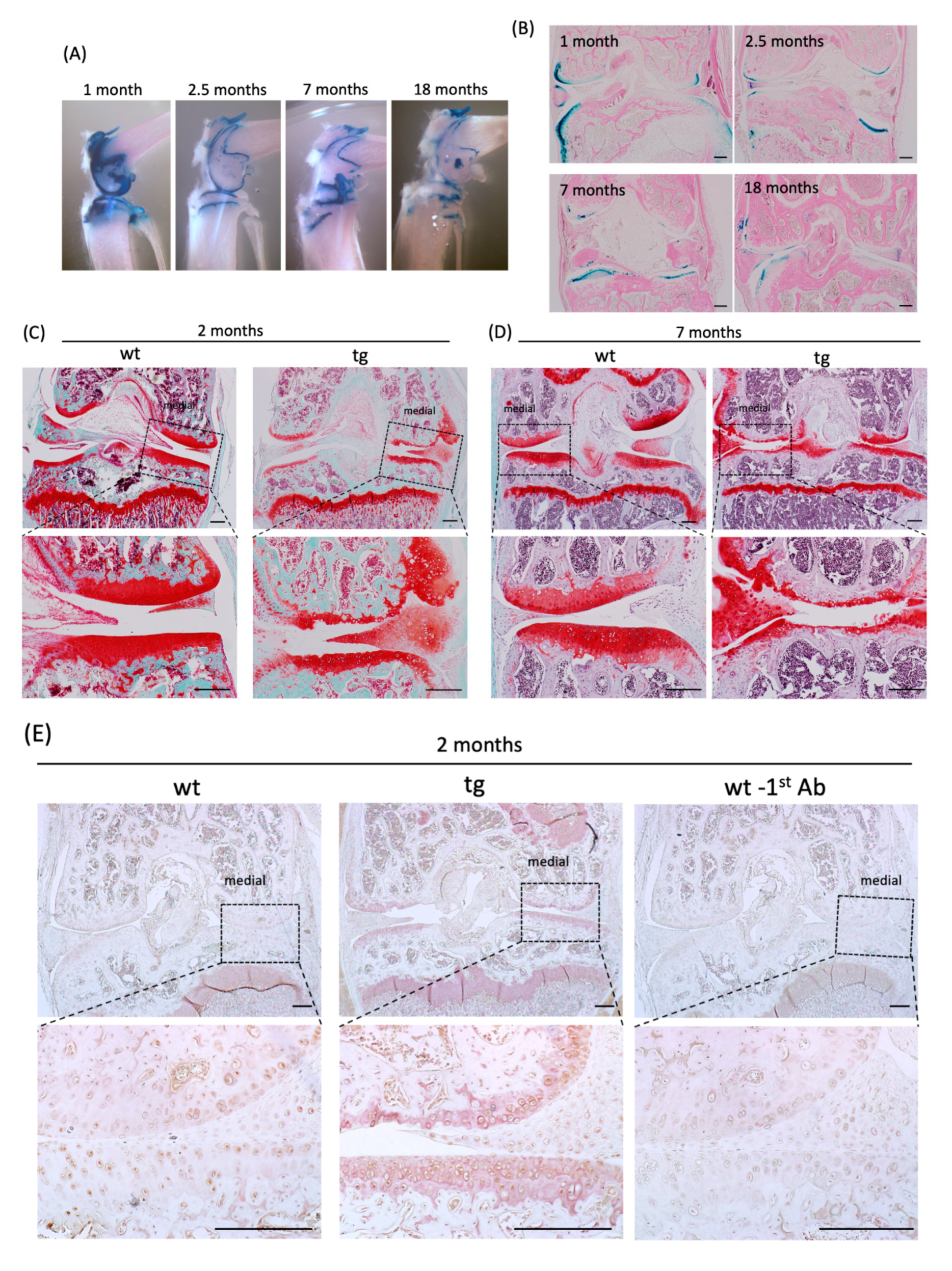
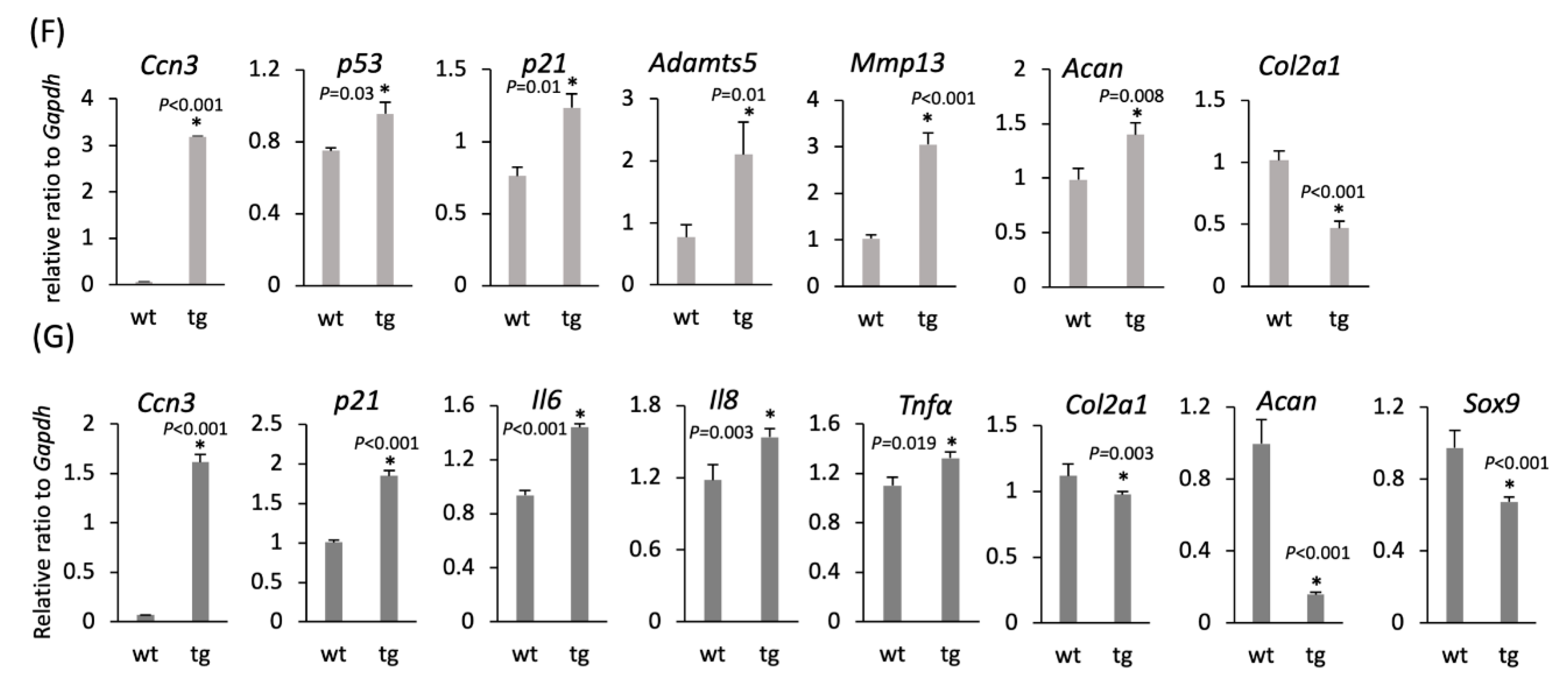
2.5. Cartilage-Specific CCN3 Overexpression Induced Degradative Changes in Articular Knee Joints and Rib Chondrocytes In Vitro
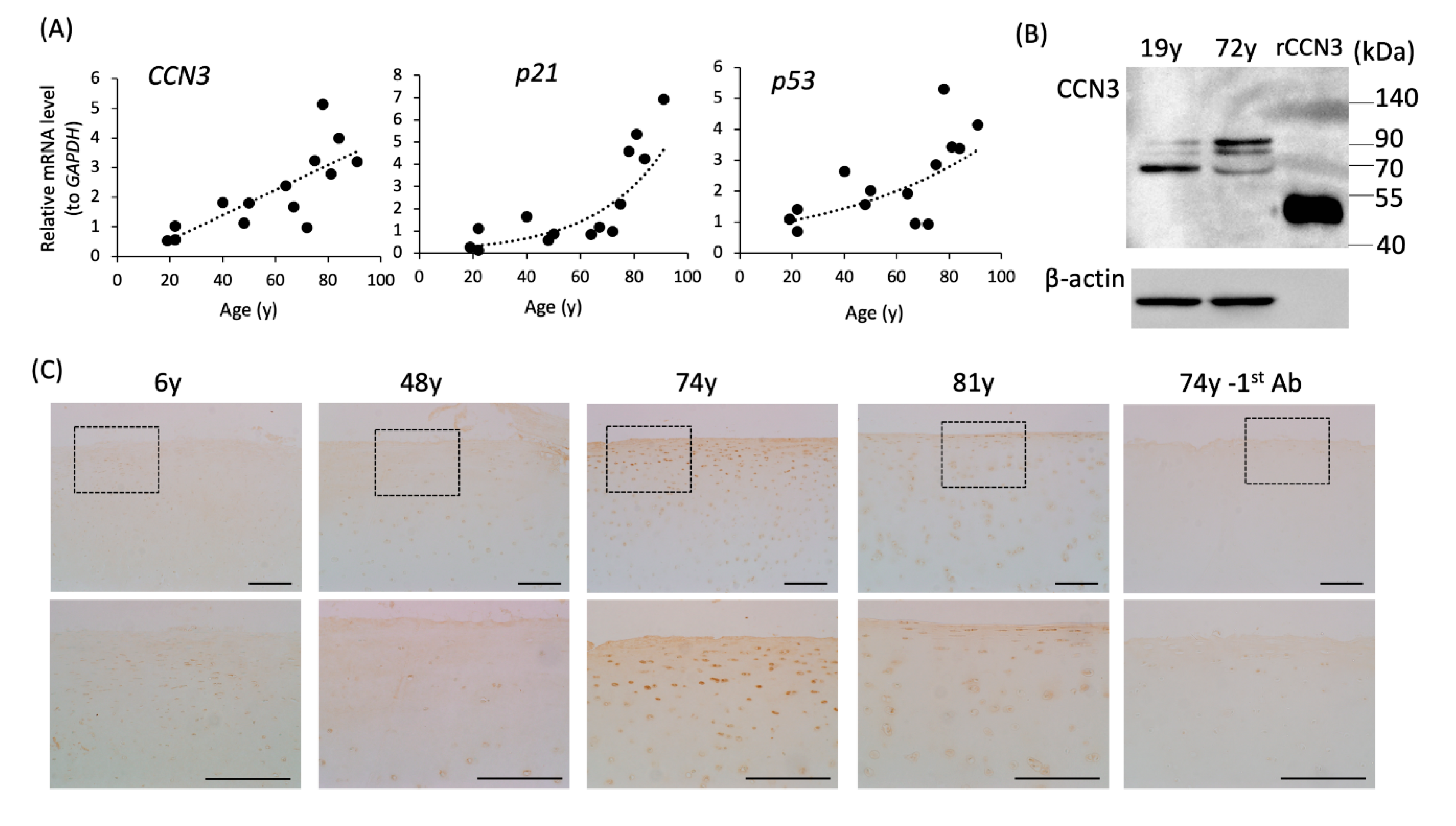
2.6. Elevation of CCN3, p53, and p21 mRNA Expression in Human Articular Cartilage with Age
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Cultures
5.2. Isolation of Total RNA
5.3. Real-Time Polymerase Chain Reaction
5.4. Histology
5.5. Immunohistochemistry
5.6. Recombinant Mouse CCN3 and CCN3 Antibody
5.7. Treatment with H₂O₂
5.8. Western Blots
5.9. Plasmids
5.10. p21 Promoter Reporter Gene Assay
5.11. Generation of Transgenic Mice
5.12. X-Gal Staining
5.13. Safranin-O Staining
5.14. Statistic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hermann, W.; Lambova, S.; Ladner, U.M. Current Treatment Options for Osteoarthritis. Curr. Rheumatol. Rev. 2018, 14, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Ashford, S.; Williard, J. Osteoarthritis: A review. Nurse Pract. 2014, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hou, A.; Chen, P.; Tang, H.; Meng, H.; Cheng, X.; Wang, Y.; Zhang, Y.; Peng, J. Cellular senescence in osteoarthritis and anti-aging strategies. Mech. Ageing Dev. 2018, 175, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Livshits, G.; Zhai, G.; Hart, D.J.; Kato, B.S.; Wang, H.; Williams, F.M.; Spector, T.D. Interleukin-6 is a significant predictor of radiographic knee osteoarthritis: The Chingford study. Arthritis Rheum. 2009, 60, 2037–2045. [Google Scholar] [CrossRef]
- Philipot, D.; Guérit, D.; Platano, D.; Chuchana, P.; Olivotto, E.; Espinoza, F.; Dorandeu, A.; Pers, Y.M.; Piette, J.; Borzì, R.M.; et al. p16INK4a and its regulator miR-24 link senescence and chondrocyte terminal differentiation-associated matrix remodeling in osteoarthritis. Arthritis Res. Ther. 2014, 16, R58. [Google Scholar] [CrossRef]
- Ashraf, S.; Cha, B.-H.; Kim, J.-S.; Ahn, J.; Han, I.; Park, H.; Lee, S.-H. Regulation of senescence associated signaling mechanisms in chondrocytes for cartilage tissue regeneration. Osteoarthr. Cartil. 2016, 24, 196–205. [Google Scholar] [CrossRef]
- Tsuchida, A.I.; Beekhuizen, M.; Hart, M.C.; Radstake, T.R.; Dhert, W.J.A.; Saris, D.B.F.; Van Osch, G.J.V.M.; Creemers, L. Cytokine profiles in the joint depend on pathology, but are different between synovial fluid, cartilage tissue and cultured chondrocytes. Arthritis Res. 2014, 16, 441. [Google Scholar] [CrossRef]
- Hui, W.; Young, D.A.; Rowan, A.D.; Xu, X.; Cawston, T.E.; Proctor, C.J. Oxidative changes and signalling pathways are pivotal in initiating age-related changes in articular cartilage. Ann. Rheum. Dis. 2015, 75, 449–458. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, X.; Lotz, M.; Terkeltaub, R.; Liu-Bryan, R. Mitochondrial Biogenesis Is Impaired in Osteoarthritis Chondrocytes but Reversible via Peroxisome Proliferator-Activated Receptor γ Coactivator 1α. Arthritis Rheumatol. 2015, 67, 2141–2153. [Google Scholar] [CrossRef]
- Blanco, F.J.; Rego, I.; Ruiz-Romero, C.; Blanco, F.J.; Rego-Perez, I. The role of mitochondria in osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 161–169. [Google Scholar] [CrossRef]
- Ulgherait, M.; Rana, A.; Rera, M.; Graniel, J.; Walker, D.W. AMPK Modulates Tissue and Organismal Aging in a Non-Cell-Autonomous Manner. Cell Rep. 2014, 8, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Caramés, B.; Olmer, M.; Kiosses, W.B.; Lotz, M.K. The relationship of autophagy defects to cartilage damage during joint aging in a mouse model. Arthritis Rheumatol. 2015, 67, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Caramés, B.; Taniguchi, N.; Otsuki, S.; Blanco, F.J.; Lotz, M.K. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum. 2010, 62, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, N.; DeGroot, J.; Ben Zaken, C.; Braun-Benjamin, O.; Maroudas, A.; Bank, R.A.; Mizrahi, J.; Schalkwijk, C.G.; Thorpe, S.R.; Baynes, J.W.; et al. Crosslinking by advanced glycation end products increases the stiffness of the collagen network in human articular cartilage: A possible mechanism through which age is a risk factor for osteoarthritis. Arthritis Rheum. 2002, 46, 114–123. [Google Scholar] [CrossRef]
- Holbourn, K.P.; Acharya, K.R.; Perbal, B. The CCN family of proteins: Structure–function relationships. Trends Biochem. Sci. 2008, 33, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, M.; Takigawa, M. The CCN Proteins: An Overview. Adv. Struct. Saf. Stud. 2016, 1489, 1–8. [Google Scholar] [CrossRef]
- Leask, A.; Abraham, D.J. All in the CCN family: Essential matricellular signaling modulators emerge from the bunker. J. Cell Sci. 2006, 119, 4803–4810. [Google Scholar] [CrossRef] [PubMed]
- Kawaki, H.; Kubota, S.; Suzuki, A.; Lazar, N.; Yamada, T.; Matsumura, T.; Ohgawara, T.; Maeda, T.; Perbal, B.; Lyons, K.M.; et al. Cooperative Regulation of Chondrocyte Differentiation by CCN2 and CCN3 Shown by a Comprehensive Analysis of the CCN Family Proteins in Cartilage. J. Bone Miner. Res. 2008, 23, 1751–1764. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. Human housekeeping genes, revisited. Trends Genet. 2013, 29, 569–574. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Rubingh, R.; DeMaria, M. Identification of stable senescence-associated reference genes. Aging Cell 2019, 18, e12911. [Google Scholar] [CrossRef]
- Chambers, M.G.; Kuffner, T.; Cowan, S.K.; Cheah, K.S.E.; Mason, R.M. Expression of collagen and aggrecan genes in normal and osteoarthritic murine knee joints. Osteoarthr. Cartil. 2002, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F.; Olex, A.L.; McNulty, M.A.; Carlson, C.S.; Callahan, M.F.; Ferguson, C.M.; Chou, J.; Leng, X.; Fetrow, J.S. Microarray analysis reveals age-related differences in gene expression during the development of osteoarthritis in mice. Arthritis Rheum. 2012, 3, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Durik, M.; Baker, D.J.; Van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef]
- Muñoz-Espín, D.; Serrano, M. Cellular senescence: From physiology to pathology. Nat. Rev. Mol. Cell Biol. 2014, 15, 482–496. [Google Scholar] [CrossRef]
- Aigner, T.; Dudhia, J. Phenotypic modulation of chondrocytes as a potential therapeutic target in osteoarthritis: A hypothesis. Ann. Rheum. Dis. 1997, 56, 287–291. [Google Scholar] [CrossRef]
- Troeberg, L.; Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta. 2012, 1824, 133–145. [Google Scholar] [CrossRef]
- Kamekura, S.; Hoshi, K.; Shimoaka, T.; Chung, U.; Chikuda, H.; Yamada, T.; Uchida, M.; Ogata, N.; Seichi, A.; Nakamura, K.; et al. Osteoarthritis development in novel experimental mouse models induced by knee joint instability. Osteoarthr. Cartil. 2005, 13, 632–641. [Google Scholar] [CrossRef]
- Shimoyama, T.; Hiraoka, S.; Takemoto, M.; Koshizaka, M.; Tokuyama, H.; Tokuyama, T.; Watanabe, A.; Fujimoto, M.; Kawamura, H.; Sato, S.; et al. CCN3 Inhibits Neointimal Hyperplasia Through Modulation of Smooth Muscle Cell Growth and Migration. Arter. Thromb. Vasc. Biol. 2010, 30, 675–682. [Google Scholar] [CrossRef]
- Heath, E.; Tahri, D.; Andermarcher, E.; Schofield, P.N.; Fleming, S.; Boulter, C.A. Abnormal skeletal and cardiac development, cardiomyopathy, muscle atrophy and cataracts in mice with a targeted disruption of the Nov (Ccn3) gene. BMC Dev. Biol. 2008, 8, 18. [Google Scholar] [CrossRef]
- Roddy, K.; Boulter, C. Targeted mutation of NOV/CCN3 in mice disrupts joint homeostasis and causes osteoarthritis-like disease. Osteoarthr. Cartil. 2015, 23, 607–615. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suresh, S.; McCallum, L.; Crawford, L.J.; Lu, W.H.; Sharpe, D.J.; Irvine, A.E. The matricellular protein CCN3 regulates NOTCH1 signalling in chronic myeloid leukaemia. J. Pathol. 2013, 231, 378–387. [Google Scholar] [CrossRef] [PubMed][Green Version]
- He, S.; Sharpless, N.E. Senescence in Health and Disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Joliot, V.; Martinerie, C.; Dambrine, G.; Plassiart, G.; Brisac, M.; Crochet, J.; Perbal, B. Proviral rearrangements and overexpression of a new cellular gene (nov) in myeloblastosis-associated virus type 1-induced nephroblastomas. Mol. Cell. Biol. 1992, 12, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Yeger, H.; Martinerie, C.; Laurent, M.; Alami, J.; Schofield, P.N.; Perbal, B. novH: Differential expression in developing kidney and Wilm’s tumors. Am. J. Pathol. 1998, 152, 1563–1575. [Google Scholar]
- Yu, C.; Le, A.-T.; Yeger, H.; Perbal, B.; Alman, B.A. NOV (CCN3) regulation in the growth plate and CCN family member expression in cartilage neoplasia. J. Pathol. 2003, 201, 609–615. [Google Scholar] [CrossRef]
- Akashi, S.; Nishida, T.; El-Seoudi, A.; Takigawa, M.; Iida, S.; Kubota, S. Metabolic regulation of the CCN family genes by glycolysis in chondrocytes. J. Cell Commun. Signal. 2017, 12, 245–252. [Google Scholar] [CrossRef]
- Huang, X.; Ni, B.; Mao, Z.; Xi, Y.; Chu, X.; Zhang, R.; Ma, X.; You, H. NOV/CCN3 induces cartilage protection by inhibiting PI3K/AKT/mTOR pathway. J. Cell. Mol. Med. 2019, 23, 7525–7534. [Google Scholar] [CrossRef]
- Peidl, A. A friend in knee: CCN3 may inhibit osteoarthritis progression. J. Cell. Commun. Signal. 2018, 12, 489–490. [Google Scholar] [CrossRef]
- Hattori, T.; Müller, C.; Gebhard, S.; Bauer, E.; Pausch, F.; Schlund, B.; Bösl, M.R.; Hess, A.; Surmann-Schmitt, C.; Von Der Mark, H.; et al. SOX9 is a major negative regulator of cartilage vascularization, bone marrow formation and endochondral ossification. Development 2010, 137, 901–911. [Google Scholar] [CrossRef]
- Tomita, N.; Hattori, T.; Itoh, S.; Aoyama, E.; Yao, M.; Yamashiro, T.; Takigawa, M. Cartilage-specific over-expression of CCN family member 2/connective tissue growth factor (CCN2/CTGF) stimulates insulin-like growth factor expression and bone growth. PLoS ONE 2013, 8, e59226. [Google Scholar] [CrossRef] [PubMed]
- King, K.B.; Kimura, J.H. The establishment and characterization of an immortal cell line with a stable chondrocytic phenotype. J. Cell. Biochem. 2003, 89, 992–1004. [Google Scholar] [CrossRef] [PubMed]
- Itahana, K.; Itahana, Y.; Dimri, G.P. Colorimetric detection of senescence-associated β galactosidase. Methods. Mol. Biol. 2013, 965, 143–156. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| Ccn3 | cagaccccaacaaccagact | acttctctccgttgcggtaa |
| p16 | gaactctttcggtcgtaccc | agttcgaatctgcaccgtagt |
| p21 | gaacatctcagggccgaaaa | tgcgcttggagtgatagaaatc |
| p53 | tcttatccgggtggaaggaaa | ggcgaaaagtctgcctgtctt |
| Aggrecan | gaggagagaactggagaag | ggcgatagtggaatacaa |
| Col2a1 | tggtggagcagcaagagcaa | cagtggacagtagacggaggaaa |
| Col10a1 | tgctgcctcaaataccctttct | tggcgtatgggatgaagtattg |
| Sox9 | atctgaagaaggagagcgag | tcagaagtctccagagcttg |
| Il-6 | ggagcccaccaagaacgata | tcccaagaaggcaactggat |
| Il-8 | aatttccaccggcaatgaag | cccgaattggaaagggaaat |
| Mmp13 | tcctcggagactggtaatgg | tgatgaaacctggacaagca |
| Adamts-5 | ggcatcattcatgtgacac | gcatcgtaggtctgtcctg |
| Tnfα | aggggattatggctcagggt | tgagtccttgatggtggtgc |
| Tmem199 | aaatggcgtcttccttgcttgc | atcactgcccgcgtgtttctt |
| Gapdh * | caatgaccccttcattgacc | gacaagcttcccgttctcag |
| Gene | Forward | Reverse |
|---|---|---|
| Ccn3 | tgaagtctctgactccagcatt | tggctttcagggatttcttg |
| p21 | caaagtatgccgtcgtctgttc | gaagtcaaagttccaccgttctc |
| p53 | tccgactataccactatccactaca | ggcacaaacacgaacctcaaa |
| Aggrecan | agaatcaagtggagccgtgtt | ggggatggctggatagttgg |
| Col2a1 | ctcacgccttcccattgttg | gttgttttggggttgagggttt |
| Col10a1 | acctcccaccccattcca | acccactattgctgctcactc |
| Mmp13 | tgcggttcactttgaggaca | tcttcttctatgaggcgggga |
| Adamts-5 | tagaccctacagcaactccgt | cctccacacactccacactt |
| Gene | Forward | Reverse |
|---|---|---|
| CCN3 | ggagcgcgctataaaacctg | tcccctctcgcttttaccaa |
| p21 | tggggctgggagtagttgt | gctggaaggtgtttggggt |
| p53 | ggtcggtgggttggtagttt | gtgtgggatggggtgagattt |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuwahara, M.; Kadoya, K.; Kondo, S.; Fu, S.; Miyake, Y.; Ogo, A.; Ono, M.; Furumatsu, T.; Nakata, E.; Sasaki, T.; et al. CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage. Int. J. Mol. Sci. 2020, 21, 7556. https://doi.org/10.3390/ijms21207556
Kuwahara M, Kadoya K, Kondo S, Fu S, Miyake Y, Ogo A, Ono M, Furumatsu T, Nakata E, Sasaki T, et al. CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage. International Journal of Molecular Sciences. 2020; 21(20):7556. https://doi.org/10.3390/ijms21207556
Chicago/Turabian StyleKuwahara, Miho, Koichi Kadoya, Sei Kondo, Shanqi Fu, Yoshiko Miyake, Ayako Ogo, Mitsuaki Ono, Takayuki Furumatsu, Eiji Nakata, Takako Sasaki, and et al. 2020. "CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage" International Journal of Molecular Sciences 21, no. 20: 7556. https://doi.org/10.3390/ijms21207556
APA StyleKuwahara, M., Kadoya, K., Kondo, S., Fu, S., Miyake, Y., Ogo, A., Ono, M., Furumatsu, T., Nakata, E., Sasaki, T., Minagi, S., Takigawa, M., Kubota, S., & Hattori, T. (2020). CCN3 (NOV) Drives Degradative Changes in Aging Articular Cartilage. International Journal of Molecular Sciences, 21(20), 7556. https://doi.org/10.3390/ijms21207556