Identifying Stabilin-1 and Stabilin-2 Double Knockouts in Reproduction and Placentation: A Descriptive Study


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Stab1 and Stab2 mRNA Expression in Mouse Placenta
2.2. Viability of Stab1/2 dKO Female Mice during Pregnancy
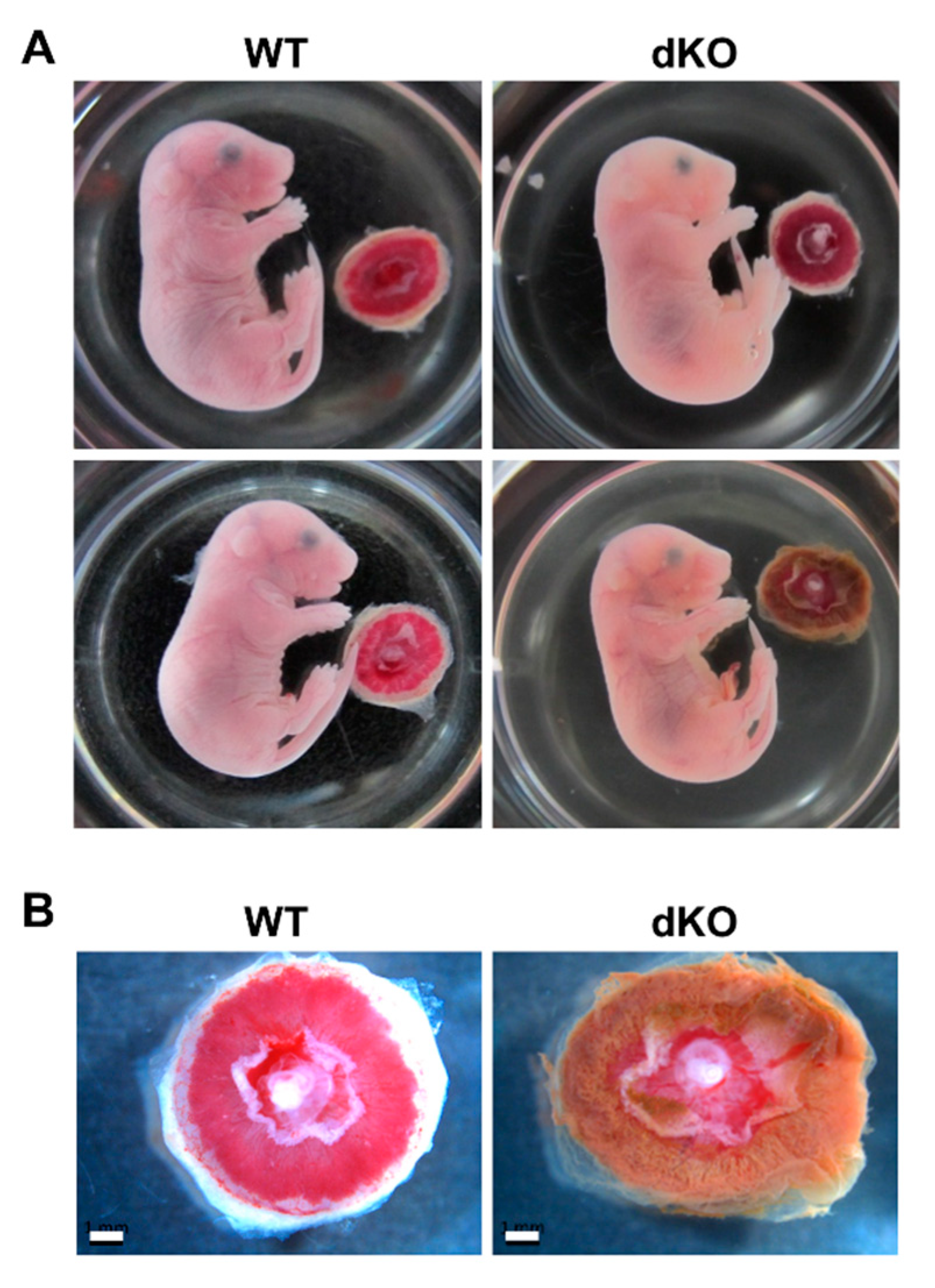
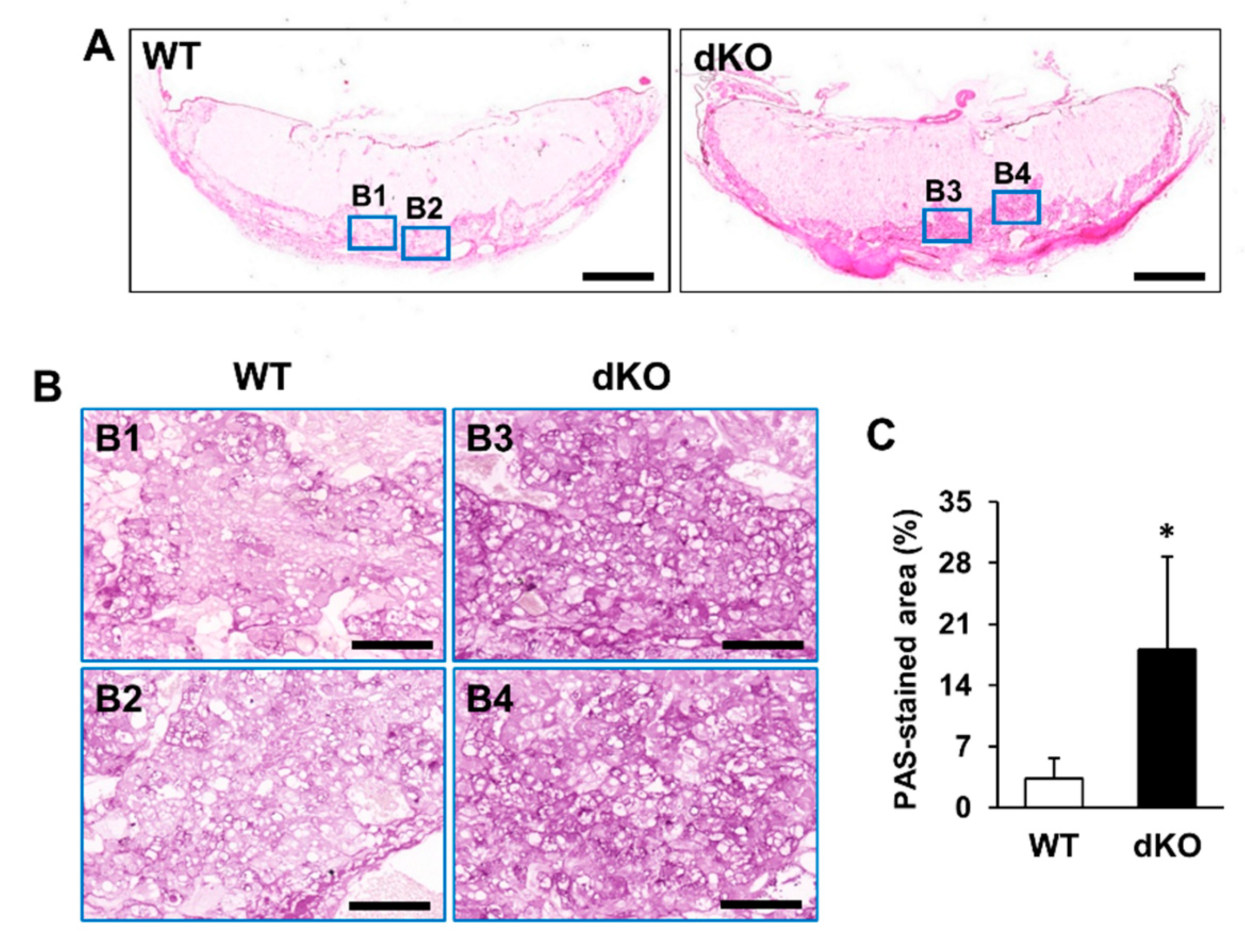
2.3. Phenotypic and Histological Defects of Placentas from Stab1/2 dKO Females
3. Discussion
4. Materials and Methods
4.1. RNA Extraction, RT-PCR, and qPCR
4.2. Animals and Sampling
4.3. Histological Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kzhyshkowska, J. Multifunctional receptor stabilin-1 in homeostasis and disease. Sci. World J. 2010, 10, 2039–2053. [Google Scholar] [CrossRef] [PubMed]
- Politz, O.; Gratchev, A.; McCourt, P.A.; Schledzewski, K.; Guillot, P.; Johansson, S.; Svineng, G.; Franke, P.; Kannicht, C.; Kzhyshkowska, J.; et al. Stabilin-1 and -2 constitute a novel family of fasciclin-like hyaluronan receptor homologues. Biochem. J. 2002, 362, 155–164. [Google Scholar]
- Palani, S.; Maksimow, M.; Miiluniemi, M.; Auvinen, K.; Jalkanen, S.; Salmi, M. Stabilin-1/CLEVER-1, a type 2 macrophage marker, is an adhesion and scavenging molecule on human placental macrophages. Eur. J. Immunol. 2011, 41, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Hirose, Y.; Saijou, E.; Sugano, Y.; Takeshita, F.; Nishimura, S.; Nonaka, H.; Chen, Y.R.; Sekine, K.; Kido, T.; Nakamura, T.; et al. Inhibition of Stabilin-2 elevates circulating hyaluronic acid levels and prevents tumor metastasis. Proc. Natl. Acad. Sci. USA 2012, 109, 4263–4268. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Lee, E.H.; Park, S.Y.; Choi, H.; Koh, J.T.; Park, E.K.; Kim, I.S.; Kim, J.E. Ablation of Stabilin-1 enhances bone-resorbing activity in osteoclast in vitro. Calcif. Tissue Int. 2019, 105, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Tamura, Y.; Adachi, H.; Osuga, J.; Yahagi, N.; Sekiya, M.; Okazaki, H.; Tomita, S.; Iizuka, Y.; Shimano, H.; Nagai, R.; et al. FEEL-1 and FEEL-2 are endocytic receptors for advanced glycation end products. J. Biol. Chem. 2003, 278, 12613–12617. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, M.; Schledzewski, K.; Hansen, B.; Goerdt, S. Expression of stabilin-2, a novel fasciclin-like hyaluronan receptor protein, in murine sinusoidal endothelia, avascular tissues, and at solid/liquid interfaces. Histochem. Cell Biol. 2003, 120, 361–369. [Google Scholar] [CrossRef]
- Lee, W.; Park, S.Y.; Yoo, Y.; Kim, S.Y.; Kim, J.E.; Kim, S.W.; Seo, Y.K.; Park, E.K.; Kim, I.S.; Bae, J.S. Macrophagic Stabilin-1 restored disruption of vascular integrity caused by sepsis. Thromb. Haemost. 2018, 118, 1776–1789. [Google Scholar] [CrossRef] [PubMed]
- Kzhyshkowska, J.; Gratchev, A.; Schmuttermaier, C.; Brundiers, H.; Krusell, L.; Mamidi, S.; Zhang, J.; Workman, G.; Sage, E.H.; Anderle, C.; et al. Alternatively activated macrophages regulate extracellular levels of the hormone placental lactogen via receptor-mediated uptake and transcytosis. J. Immunol. 2008, 180, 3028–3037. [Google Scholar] [CrossRef] [PubMed]
- Schledzewski, K.; Géraud, C.; Arnold, B.; Wang, S.; Gröne, H.J.; Kempf, T.; Wollert, K.C.; Straub, B.K.; Schirmacher, P.; Demory, A.; et al. Deficiency of liver sinusoidal scavenger receptors stabilin-1 and -2 mice causes glomerulofibrotic nephropathy via impaired hepatic clearance of noxious blood factors. J. Clin. Investig. 2011, 121, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Akison, L.K.; Nitert, M.D.; Clifton, V.L.; Moritz, K.M.; Simmons, D.G. Review: Alterations in placental glycogen deposition in complicated pregnancies: Current preclinical and clinical evidence. Placenta 2017, 54, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Hazan, A.D.; Smith, S.D.; Jones, R.L.; Whittle, W.; Lye, S.J.; Dunk, C.E. Vascular-leukocyte interactions: Mechanisms of human decidual spiral artery remodeling in vitro. Am. J. Pathol. 2010, 177, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Cross, J.C. Development of the hemochorial maternal vascular spaces in the placenta through endothelial and vasculogenic mimicry. Dev. Biol. 2014, 387, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Rost, M.S.; Sumanas, S. Hyaluronic acid receptor Stabilin-2 regulates Erk phosphorylation and arterial-venous differentiation in zebrafish. PLoS ONE 2014, 9, e88614. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Conroy, N.; Burton, G.J.; Ferguson-Smith, A.C. Origin and characteristics of glycogen cells in the developing murine placenta. Dev. Dyn. 2006, 235, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- Oyelese, Y.; Ananth, C.V. Placental abruption. Obstet. Gynecol. 2006, 108, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Tikkanen, M. Etiology, clinical manifestations, and prediction of placental abruption. Acta Obstet. Gynecol. Scand. 2010, 89, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Bogdan, A.; Balassa, T.; Csabai, T.; Szekeres-Bartho, J. The decidua-the maternal bed embracing the embryo-maintains the pregnancy. Semin. Immunopathol. 2016, 38, 635–649. [Google Scholar] [CrossRef]
- Herington, J.L.; Bany, B.M. Effect of the conceptus on uterine natural killer cell numbers and function in the mouse uterus during decidualization. Biol. Reprod. 2007, 76, 579–588. [Google Scholar] [CrossRef]
- Simmons, D.G.; Fortier, A.L.; Cross, J.C. Diverse subtypes and developmental origins of trophoblast giant cells in the mouse placenta. Dev. Biol. 2007, 304, 567–578. [Google Scholar] [CrossRef]
- Park, S.Y.; Yun, Y.; Lim, J.S.; Kim, M.J.; Kim, S.Y.; Kim, J.E.; Kim, I.S. Stabilin-2 modulates the efficiency of myoblast fusion during myogenic differentiation and muscle regeneration. Nat. Commun. 2016, 7, 10871. [Google Scholar] [CrossRef] [PubMed]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.; Perez-Garcia, V.; Hemberger, M. Regulation of placental development and its impact on fetal growth—new insights from mouse models. Front. Endocrinol. 2018, 9, 570. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Lee, E.-H.; Kim, E.N.; Son, W.-C.; Kim, Y.H.; Park, S.-Y.; Kim, I.-S.; Kim, J.-E. Identifying Stabilin-1 and Stabilin-2 Double Knockouts in Reproduction and Placentation: A Descriptive Study. Int. J. Mol. Sci. 2020, 21, 7235. https://doi.org/10.3390/ijms21197235
Kim S-Y, Lee E-H, Kim EN, Son W-C, Kim YH, Park S-Y, Kim I-S, Kim J-E. Identifying Stabilin-1 and Stabilin-2 Double Knockouts in Reproduction and Placentation: A Descriptive Study. International Journal of Molecular Sciences. 2020; 21(19):7235. https://doi.org/10.3390/ijms21197235
Chicago/Turabian StyleKim, Soon-Young, Eun-Hye Lee, Eun Na Kim, Woo-Chan Son, Yeo Hyang Kim, Seung-Yoon Park, In-San Kim, and Jung-Eun Kim. 2020. "Identifying Stabilin-1 and Stabilin-2 Double Knockouts in Reproduction and Placentation: A Descriptive Study" International Journal of Molecular Sciences 21, no. 19: 7235. https://doi.org/10.3390/ijms21197235
APA StyleKim, S.-Y., Lee, E.-H., Kim, E. N., Son, W.-C., Kim, Y. H., Park, S.-Y., Kim, I.-S., & Kim, J.-E. (2020). Identifying Stabilin-1 and Stabilin-2 Double Knockouts in Reproduction and Placentation: A Descriptive Study. International Journal of Molecular Sciences, 21(19), 7235. https://doi.org/10.3390/ijms21197235