Effects of Ketamine on Rodent Fear Memory

 ,
,
Abstract
:1. Introduction
2. Effects of Ketamine on Fear Memory
2.1. Route of Ketamine Administration
2.2. Ketamine Dosage
2.3. Timing of Ketamine Administration
2.3.1. Pre-Fear Conditioning
2.3.2. Immediately Post-Fear Conditioning
2.3.3. Delayed Post-Fear Conditioning
2.4. Ketamine and Stress
2.5. Individual Variability
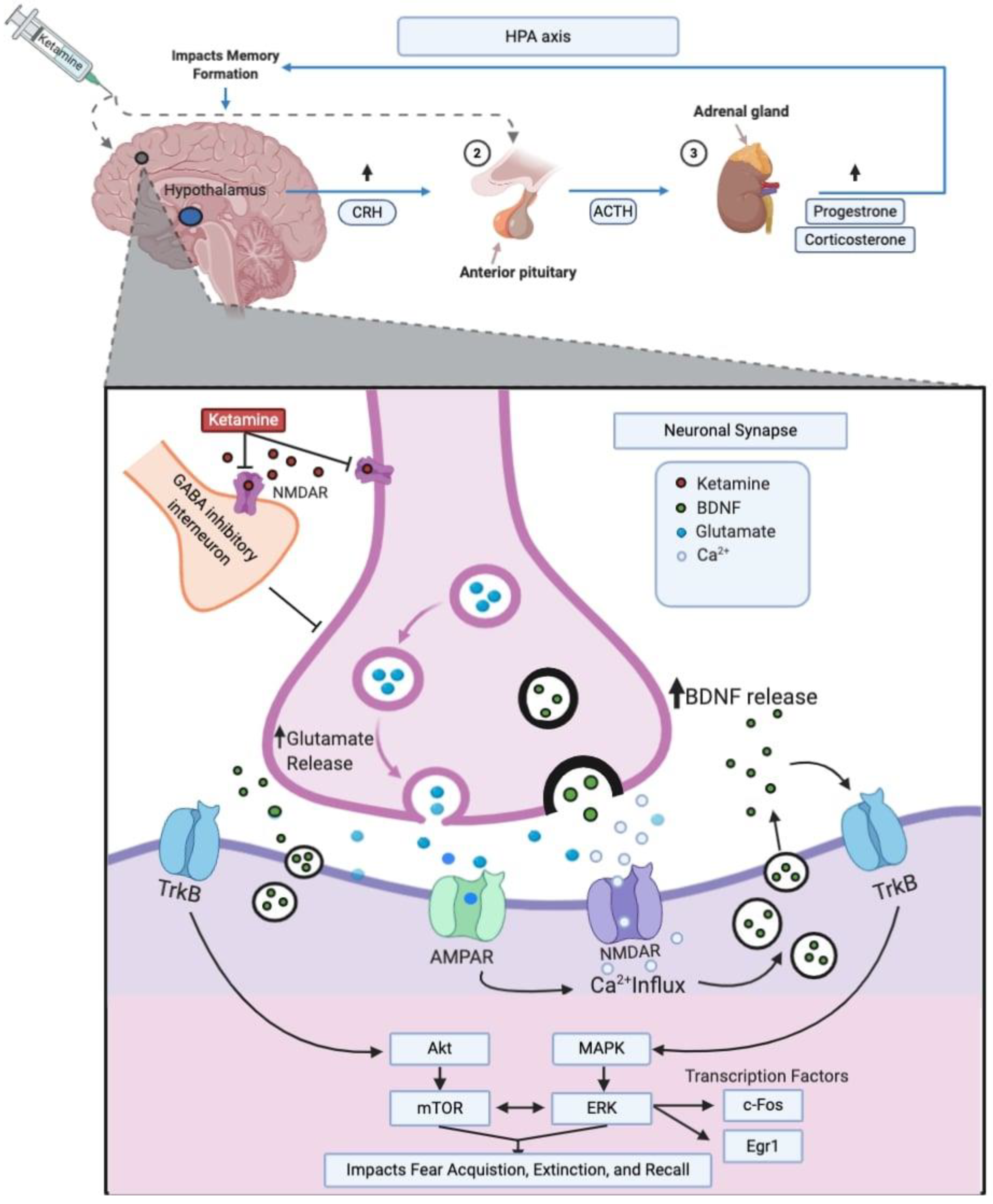
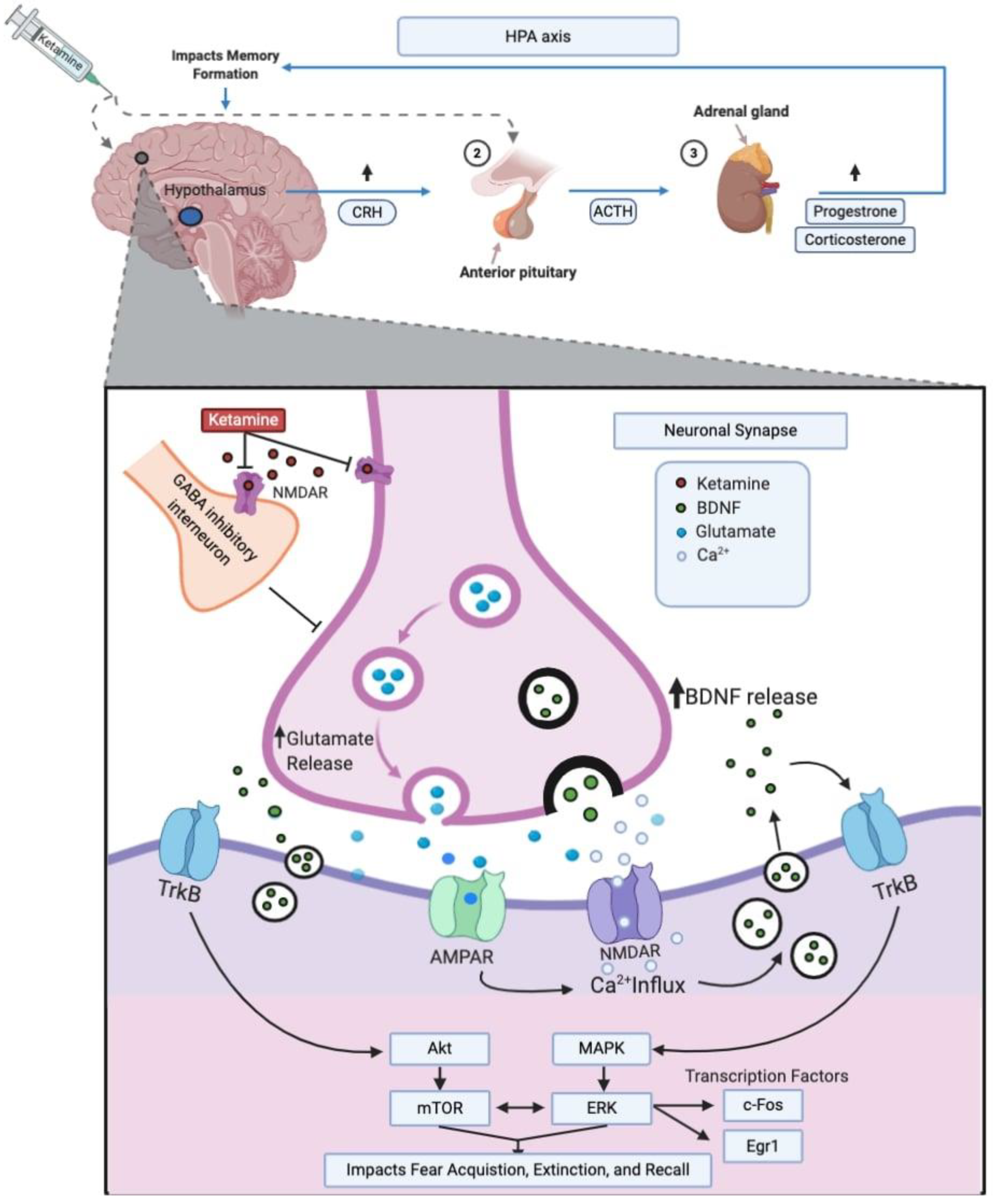
3. Molecular Mechanisms
3.1. Glutamatergic Signaling
3.2. BDNF
3.3. mTOR and ERK
3.4. Transcription Factors
3.5. Brain Glucose Metabolism
3.6. Stress Hormones and HPA Axis
4. Conclusions
5. Future Directions
Disclaimer
Funding
Acknowledgments
Conflicts of Interest
References
- Morris, C.; Perris, A.; Klein, J.; Mahoney, P. Anaesthesia in haemodynamically compromised emergency patients: Does ketamine represent the best choice of induction agent? Anaesthesia 2009, 64, 532–539. [Google Scholar] [CrossRef]
- Wedmore, I.S.; Butler, F.K., Jr. Battlefield Analgesia in Tactical Combat Casualty Care. Wilderness Environ. Med. 2017, 28, S109–S116. [Google Scholar] [CrossRef] [Green Version]
- Juven-Wetzler, A.; Cohen, H.; Kaplan, Z.; Kohen, A.; Porat, O.; Zohar, J. Immediate ketamine treatment does not prevent posttraumatic stress responses in an animal model for PTSD. Eur. Neuropsychopharmacol. 2014, 24, 469–479. [Google Scholar] [CrossRef]
- Honsberger, M.J.; Taylor, J.R.; Corlett, P.R. Memories reactivated under ketamine are subsequently stronger: A potential pre-clinical behavioral model of psychosis. Schizophr. Res. 2015, 164, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Saur, L.; Neves, L.T.; Greggio, S.; Venturin, G.T.; Jeckel, C.M.M.; Costa Da Costa, J.; Bertoldi, K.; Schallenberger, B.; Siqueira, I.R.; Mestriner, R.G.; et al. Ketamine promotes increased freezing behavior in rats with experimental PTSD without changing brain glucose metabolism or BDNF. Neurosci. Lett. 2017, 658, 6–11. [Google Scholar] [CrossRef]
- Clifton, N.E.; Thomas, K.L.; Hall, J. The effect of ketamine on the consolidation and extinction of contextual fear memory. J. Psychopharmacol. 2018, 32, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ju, W.; Zhang, H.; Sun, L. Effect of Ketamine on LTP and NMDAR EPSC in Hippocampus of the Chronic Social Defeat Stress Mice Model of Depression. Front. Behav. Neurosci. 2018, 12, 229. [Google Scholar] [CrossRef] [Green Version]
- Calzavara, M.B.; Medrano, W.A.; Levin, R.; Kameda, S.R.; Andersen, M.L.; Tufik, S.; Silva, R.H.; Frussa-Filho, R.; Abilio, V.C. Neuroleptic drugs revert the contextual fear conditioning deficit presented by spontaneously hypertensive rats: A potential animal model of emotional context processing in schizophrenia? Schizophr. Bull. 2009, 35, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.M.; Heaney, C.F.; Sabbagh, J.J.; Murtishaw, A.S.; Magcalas, C.M.; Kinney, J.W. Deficits in emotional learning and memory in an animal model of schizophrenia. Behav. Brain Res. 2012, 233, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, W.; Erisir, A.; Morozov, A. Observation of Distressed Conspecific as a Model of Emotional Trauma Generates Silent Synapses in the Prefrontal-Amygdala Pathway and Enhances Fear Learning, but Ketamine Abolishes those Effects. Neuropsychopharmacology 2015, 40, 2536–2545. [Google Scholar] [CrossRef] [PubMed]
- Duclot, F.; Perez-Taboada, I.; Wright, K.N.; Kabbaj, M. Prediction of individual differences in fear response by novelty seeking, and disruption of contextual fear memory reconsolidation by ketamine. Neuropharmacology 2016, 109, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulyk, B.D. Antidepressant-Like Effects of Ketamine on Fear Conditioning and Extinction. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2017. [Google Scholar]
- Girgenti, M.J.; Ghosal, S.; LoPresto, D.; Taylor, J.R.; Duman, R.S. Ketamine accelerates fear extinction via mTORC1 signaling. Neurobiol. Dis. 2017, 100, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, J.C.; LaGamma, C.T.; Lim, S.C.; Tsitsiklis, M.; Neria, Y.; Brachman, R.A.; Denny, C.A. Prophylactic Ketamine Attenuates Learned Fear. Neuropsychopharmacology 2017, 42, 1577–1589. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.C.; Hill, C.; Mastrodonato, A.; LaGamma, C.T.; Kitayev, A.; Brachman, R.A.; Narain, N.R.; Kiebish, M.A.; Denny, C.A. Prophylactic ketamine alters nucleotide and neurotransmitter metabolism in brain and plasma following stress. Neuropsychopharmacology 2018, 43, 1813–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrodonato, A.; Martinez, R.; Pavlova, I.P.; LaGamma, C.T.; Brachman, R.A.; Robison, A.J.; Denny, C.A. Ventral CA3 Activation Mediates Prophylactic Ketamine Efficacy Against Stress-Induced Depressive-like Behavior. Biol. Psychiatry 2018, 84, 846–856. [Google Scholar] [CrossRef]
- Asim, M.; Hao, B.; Yang, Y.H.; Fan, B.F.; Xue, L.; Shi, Y.W.; Wang, X.G.; Zhao, H. Ketamine Alleviates Fear Generalization Through GluN2B-BDNF Signaling in Mice. Neurosci. Bull. 2020, 36, 153–164. [Google Scholar] [CrossRef]
- Ali, F.; Gerhard, D.M.; Sweasy, K.; Pothula, S.; Pittenger, C.; Duman, R.S.; Kwan, A.C. Ketamine disinhibits dendrites and enhances calcium signals in prefrontal dendritic spines. Nat. Commun. 2020, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.K.; Luna, V.M.; LaGamma, C.T.; Xu, X.; Deng, S.X.; Suckow, R.F.; Cooper, T.B.; Shah, A.; Brachman, R.A.; Mendez-David, I.; et al. Sex-specific neurobiological actions of prophylactic (R,S)-ketamine, (2R,6R)-hydroxynorketamine, and (2S,6S)-hydroxynorketamine. Neuropsychopharmacology 2020, 45, 1545–1556. [Google Scholar] [CrossRef]
- Radford, K.D.; Spencer, H.F.; Zhang, M.; Berman, R.Y.; Girasek, Q.L.; Choi, K.H. Association between intravenous ketamine-induced stress hormone levels and long-term fear memory renewal in Sprague-Dawley rats. Behav. Brain Res. 2020, 378, 112259. [Google Scholar] [CrossRef]
- Babar, E.; Ozgunen, T.; Melik, E.; Polat, S.; Akman, H. Effects of ketamine on different types of anxiety/fear and related memory in rats with lesions of the median raphe nucleus. Eur. J. Pharmacol 2001, 431, 315–320. [Google Scholar] [CrossRef]
- Radford, K.D.; Park, T.Y.; Jaiswal, S.; Pan, H.; Knutsen, A.; Zhang, M.; Driscoll, M.; Osborne-Smith, L.A.; Dardzinski, B.J.; Choi, K.H. Enhanced fear memories and brain glucose metabolism ((18)F-FDG-PET) following sub-anesthetic intravenous ketamine infusion in Sprague-Dawley rats. Transl. Psychiatry 2018, 8, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marietta, M.P.; White, P.F.; Pudwill, C.R.; Way, W.L.; Trevor, A.J. Biodisposition of ketamine in the rat: Self-induction of metabolism. J. Pharmacol. Exp. Ther. 1976, 196, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Radford, K.D.; Park, T.Y.; Lee, B.H.; Moran, S.; Osborne, L.A.; Choi, K.H. Dose-response characteristics of intravenous ketamine on dissociative stereotypy, locomotion, sensorimotor gating, and nociception in male Sprague-Dawley rats. Pharmacol. Biochem. Behav. 2017, 153, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrier, N.; Kabbaj, M. Sex differences in the antidepressant-like effects of ketamine. Neuropharmacology 2013, 70, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Saito, Y.; Yanagawa, Y.; Otani, S.; Hiraide, S.; Shimamura, K.; Matsumoto, M.; Togashi, H. Early postnatal stress alters extracellular signal-regulated kinase signaling in the corticolimbic system modulating emotional circuitry in adult rats. Eur. J. Neurosci. 2012, 35, 135–145. [Google Scholar] [CrossRef]
- Miracle, A.D.; Brace, M.F.; Huyck, K.D.; Singler, S.A.; Wellman, C.L. Chronic stress impairs recall of extinction of conditioned fear. Neurobiol. Learn. Mem. 2006, 85, 213–218. [Google Scholar] [CrossRef]
- Izquierdo, A.; Wellman, C.L.; Holmes, A. Brief uncontrollable stress causes dendritic retraction in infralimbic cortex and resistance to fear extinction in mice. J. Neurosci. 2006, 26, 5733–5738. [Google Scholar] [CrossRef] [Green Version]
- Brachman, R.A.; McGowan, J.C.; Perusini, J.N.; Lim, S.C.; Pham, T.H.; Faye, C.; Gardier, A.M.; Mendez-David, I.; David, D.J.; Hen, R.; et al. Ketamine as a Prophylactic Against Stress-Induced Depressive-like Behavior. Biol. Psychiatry 2016, 79, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Sleigh, J.; Harvey, M.; Voss, L.; Denny, B. Ketamine—More mechanisms of action than just NMDA blockade. Trends Anaesth. Crit. Care 2014, 4, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrova, L.R.; Phillips, A.G.; Wang, Y.T. Antidepressant effects of ketamine and the roles of AMPA glutamate receptors and other mechanisms beyond NMDA receptor antagonism. J. Psychiatry Neurosci. 2017, 42, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Bliss, T.V.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Luscher, C.; Malenka, R.C. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 2012, 4, a005710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, E.; Tang, Y.P.; Rampon, C.; Tsien, J.Z. NMDA receptor-dependent synaptic reinforcement as a crucial process for memory consolidation. Science 2000, 290, 1170–1174. [Google Scholar] [CrossRef] [Green Version]
- Davis, M. NMDA receptors and fear extinction: Implications for cognitive behavioral therapy. Dialogues Clin. Neurosci. 2011, 13, 463–474. [Google Scholar]
- Zanos, P.; Gould, T.D. Mechanisms of ketamine action as an antidepressant. Mol. Psychiatry 2018, 23, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.; Gardier, A.M. Fast-acting antidepressant activity of ketamine: Highlights on brain serotonin, glutamate, and GABA neurotransmission in preclinical studies. Pharmacol. Ther. 2019, 199, 58–90. [Google Scholar] [CrossRef]
- Duman, R.S.; Li, N.; Liu, R.J.; Duric, V.; Aghajanian, G. Signaling pathways underlying the rapid antidepressant actions of ketamine. Neuropharmacology 2012, 62, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Browne, C.A.; Lucki, I. Antidepressant effects of ketamine: Mechanisms underlying fast-acting novel antidepressants. Front. Pharmacol. 2013, 4, 161. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Radford, K.D.; Driscoll, M.; Purnomo, S.; Kim, J.; Choi, K.H. Effects of subanesthetic intravenous ketamine infusion on neuroplasticity-related proteins in the prefrontal cortex, amygdala, and hippocampus of Sprague-Dawley rats. IBRO Rep. 2019, 6, 87–94. [Google Scholar] [CrossRef]
- Nguyen, L.; Lucke-Wold, B.P.; Logsdon, A.F.; Scandinaro, A.L.; Huber, J.D.; Matsumoto, R.R. Behavioral and biochemical effects of ketamine and dextromethorphan relative to its antidepressant-like effects in Swiss Webster mice. Neuroreport 2016, 27, 1004–1011. [Google Scholar] [CrossRef] [Green Version]
- Carriere, A.; Romeo, Y.; Acosta-Jaquez, H.A.; Moreau, J.; Bonneil, E.; Thibault, P.; Fingar, D.C.; Roux, P.P. ERK1/2 phosphorylate Raptor to promote Ras-dependent activation of mTOR complex 1 (mTORC1). J. Biol. Chem. 2011, 286, 567–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.H.; Chiang, Y.W.; Liang, K.C.; Thompson, R.F.; Liu, I.Y. Extra-cellular signal-regulated kinase 1/2 (ERK1/2) activated in the hippocampal CA1 neurons is critical for retrieval of auditory trace fear memory. Brain Res. 2010, 1326, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Schafe, G.E.; Atkins, C.M.; Swank, M.W.; Bauer, E.P.; Sweatt, J.D.; LeDoux, J.E. Activation of ERK/MAP kinase in the amygdala is required for memory consolidation of pavlovian fear conditioning. J. Neurosci. 2000, 20, 8177–8187. [Google Scholar] [CrossRef] [PubMed]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Lee, B.; Liu, R.J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.Y.; Aghajanian, G.; Duman, R.S. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 2010, 329, 959–964. [Google Scholar] [CrossRef] [Green Version]
- Donahue, R.J.; Muschamp, J.W.; Russo, S.J.; Nestler, E.J.; Carlezon, W.A., Jr. Effects of striatal DeltaFosB overexpression and ketamine on social defeat stress-induced anhedonia in mice. Biol. Psychiatry 2014, 76, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.L.; Everitt, B.J.; Thomas, K.L. Independent cellular processes for hippocampal memory consolidation and reconsolidation. Science 2004, 304, 839–843. [Google Scholar] [CrossRef]
- Barsegyan, A.; Mackenzie, S.M.; Kurose, B.D.; McGaugh, J.L.; Roozendaal, B. Glucocorticoids in the prefrontal cortex enhance memory consolidation and impair working memory by a common neural mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 16655–16660. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, T.W.; Lovallo, W.R. Enhanced memory for emotional material following stress-level cortisol treatment in humans. Psychoneuroendocrinology 2001, 26, 307–317. [Google Scholar] [CrossRef]
- Barros, L.A.; Tufik, S.; Andersen, M.L. The role of progesterone in memory: An overview of three decades. Neurosci. Biobehav. Rev. 2015, 49, 193–204. [Google Scholar] [CrossRef]
- de Quervain, D.; Schwabe, L.; Roozendaal, B. Stress, glucocorticoids and memory: Implications for treating fear-related disorders. Nat. Rev. Neurosci. 2017, 18, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Hergovich, N.; Singer, E.; Agneter, E.; Eichler, H.G.; Graselli, U.; Simhandl, C.; Jilma, B. Comparison of the effects of ketamine and memantine on prolactin and cortisol release in men. a randomized, double-blind, placebo-controlled trial. Neuropsychopharmacology 2001, 24, 590–593. [Google Scholar] [CrossRef] [Green Version]
- Krystal, J.H.; D’Souza, D.C.; Karper, L.P.; Bennett, A.; Abi-Dargham, A.; Abi-Saab, D.; Cassello, K.; Bowers, M.B., Jr.; Vegso, S.; Heninger, G.R.; et al. Interactive effects of subanesthetic ketamine and haloperidol in healthy humans. Psychopharmacology 1999, 145, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Khalili-Mahani, N.; Martini, C.H.; Olofsen, E.; Dahan, A.; Niesters, M. Effect of subanaesthetic ketamine on plasma and saliva cortisol secretion. Br. J. Anaesth. 2015, 115, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Radford, C.K.D.; Park, T.Y.; Osborne-Smith, L.; Choi, K.H. Effects of Subanesthetic Intravenous Ketamine Infusion on Corticosterone and Brain-Derived Neurotrophic Factor in the Plasma of Male Sprague-Dawley Rats. AANA J. 2018, 86, 393–400. [Google Scholar] [PubMed]
- Deak, T.; Kudinova, A.; Lovelock, D.F.; Gibb, B.E.; Hennessy, M.B. A multispecies approach for understanding neuroimmune mechanisms of stress. Dialogues Clin. Neurosci. 2017, 19, 37–53. [Google Scholar]

{kind=link}
| Study | Sex, Species, Strain, Age | Dose, Route, Timing | Behavioral Results | Molecular Results |
|---|---|---|---|---|
| Ketamine Increased Fear Memory | ||||
| Juven-Wetzler et al., 2014 [3] | Male, rats, S-D | 0.5, 5, 15 mg/kg IP 1 h after predator-scent stress | Increased fear memory | Attenuated increase in CORT after predator-scent stress exposure |
| Honsberger et al., 2015 [4] | Male, rats, S-D, adult | 10 mg/kg IP, 15 min before or just after reactivation | Pre-reactivation ketamine increased fear memory | Effect nullified by GluN2B antagonist ifenprodil |
| Saur et al., 2017 [5] | Male, rats, Wistar, 12 w | 10 mg/kg IP 1 day before situational reminder | Increased fear memory | No change in brain glucose metabolism or BDNF levels |
| Clifton et al., 2018 [6] | Male, rats, Lister Hooded, adult | 8, 25 mg/kg IP 30 min before or just after CFC and extinction | Pre-extinction ketamine impaired fear extinction consolidation | - |
| Yang et al., 2018 [7] | Male, mice, C57BL/6 J, 8 w | 5 mg/kg IP 1 d before CFC | Increased contextual fear memory | Partially restored hippocampal GluN2B subunit density, LTP induction, NMDAR EPSC amplitude in depressed mice |
| Ketamine Decreased Fear Memory | ||||
| Calzavara et al., 2009 [8] | Male, rats, Wistar, 5 months | 10 mg/kg IP 15 min before CFC | Reduced fear memory | - |
| Bolton et al., 2012 [9] | Male, rats, S-D, adult | 8 mg/kg SC 30 min before trace or delay FC | Reduced trace fear memory No effect on delay fear memory | Increased GABAB1b in amygdala of trace conditioned animals receiving ketamine |
| Ito et al., 2015 [10] | Male, mice, 129SvEv/C57BL/6N, P60–75 | 10 mg/kg IP after observational fear | Prevented increased passive avoidance fear memory | Increased AMPAR/NMDAR current ratio, and reduced silent synapses in IL-PFC–BLA pathway |
| Duclot et al., 2016 [11] | Male, rats, S-D, 8 w | 10, 20 mg/kg IP after reactivation | Impaired reconsolidation | 10 mg/kg decreased Egr1 mRNA, increased BDNF mRNA |
| Kulyk et al., 2017 [12] | Male, rats, Long-Evans, adult | 10 mg/kg IP 60 min pre-FC or pre-extinction, just after FC 15 mg/kg IP 60 min pre-extinction | Post-FC and pre-extinction 10 mg/kg ketamine decreased fear recall 15 mg/kg nullified CORT-induced increase in fear memory | |
| Girgenti et al., 2017 [13] | Male, rats, S-D, 7–9 w | 10 mg/kg IP 24 h after FC | Enhanced extinction and reduced return of fear memory | Increased p-ERK and p-AKT Effect nullified by mTOR antagonist rapamycin |
| McGowan et al., 2017 [14] | Male, mice, 129S6/SvEvTac, 8 w | 30 mg/kg IP 1 week, 1 month, 1 day, 1 h before CFC | Ketamine 1 week before CFC reduced fear memory | - |
| McGowan et al., 2018 [15] | Male, mice, 129S6/SvEvTac, 7 w | 30 mg/kg IP 1 w before CFC | Reduced fear memory | Altered nucleotide and amino-acid-derived neurotransmitter metabolism |
| Mastrodonato et al., 2018 [16] | Male, mice, 129S6/SvEvTac, ArcCreERT2 x ChR2–EYFP | 30 mg/kg IP 1 w before CFC | Reduced fear memory | Increased ΔFosB levels; ΔFosB transcription antagonist blocked ketamine effect |
| Asim et al., 2019 [17] | Male, mice, C57BL/6, 6–8 w | 30 mg/kg IP 1 h or 1 week before, just after, 22 h after FC | Ketamine 22 h after FC reduced fear generalization | Increased BDNF and lowered GluN2B protein expression in BLA and IL-PFC; BDNF and GluN2B antagonists restored fear generalization |
| Ali et al., 2020 [18] | Male, mice, C57BL/6J, 10 w | 10 mg/kg SC 15 min before trace and delay FC | Decreased trace fear memory No effect in delay fear memory | GluN2B knockdown in somatostatin-expressing (SST) interneurons abolished ketamine reduction of trace fear memory |
| Chen et al., 2020 [19] | Male and female, mice, 129S6/SvEv, 7 w | 30 mg/kg IP 1 w before CFC | Attenuated contextual fear memory in males | Blocked AMPAR current bursting |
| No Effects of Ketamine on Fear Memory | ||||
| Radford et al., 2020 [20] | Male, rats, S-D | 10 mg/kg IV after FC | No effect on long-term fear memory | Increased CORT and progesterone |
| Mixed Effects of Ketamine on Fear Memory | ||||
| Babar et al., 2001 [21] | Male, rats, Wistar | 3 mg/kg IP 15 min before testing 10 mg/kg IP 15 min before testing | 3 mg/kg decreased freezing 10 mg/kg increased escape, decreased avoidance and freezing | - |
| Radford et al., 2018 [22] | Male, rats, S-D | 2, 10, 20 mg/kg IV after FC 10 mg/kg IP after FC | 10 mg/kg IV increased fear memory 10 mg/kg IP decreased fear memory | Infusion after FC increased glucose metabolism in amygdala, hippocampus, hypothalamus, midbrain; decreased in cerebellum |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, K.H.; Berman, R.Y.; Zhang, M.; Spencer, H.F.; Radford, K.D. Effects of Ketamine on Rodent Fear Memory. Int. J. Mol. Sci. 2020, 21, 7173. https://doi.org/10.3390/ijms21197173
Choi KH, Berman RY, Zhang M, Spencer HF, Radford KD. Effects of Ketamine on Rodent Fear Memory. International Journal of Molecular Sciences. 2020; 21(19):7173. https://doi.org/10.3390/ijms21197173
Chicago/Turabian StyleChoi, Kwang H., Rina Y. Berman, Michael Zhang, Haley F. Spencer, and Kennett D. Radford. 2020. "Effects of Ketamine on Rodent Fear Memory" International Journal of Molecular Sciences 21, no. 19: 7173. https://doi.org/10.3390/ijms21197173
APA StyleChoi, K. H., Berman, R. Y., Zhang, M., Spencer, H. F., & Radford, K. D. (2020). Effects of Ketamine on Rodent Fear Memory. International Journal of Molecular Sciences, 21(19), 7173. https://doi.org/10.3390/ijms21197173