Mechanisms of Interactions between Bile Acids and Plant Compounds—A Review




Abstract
1. Introduction
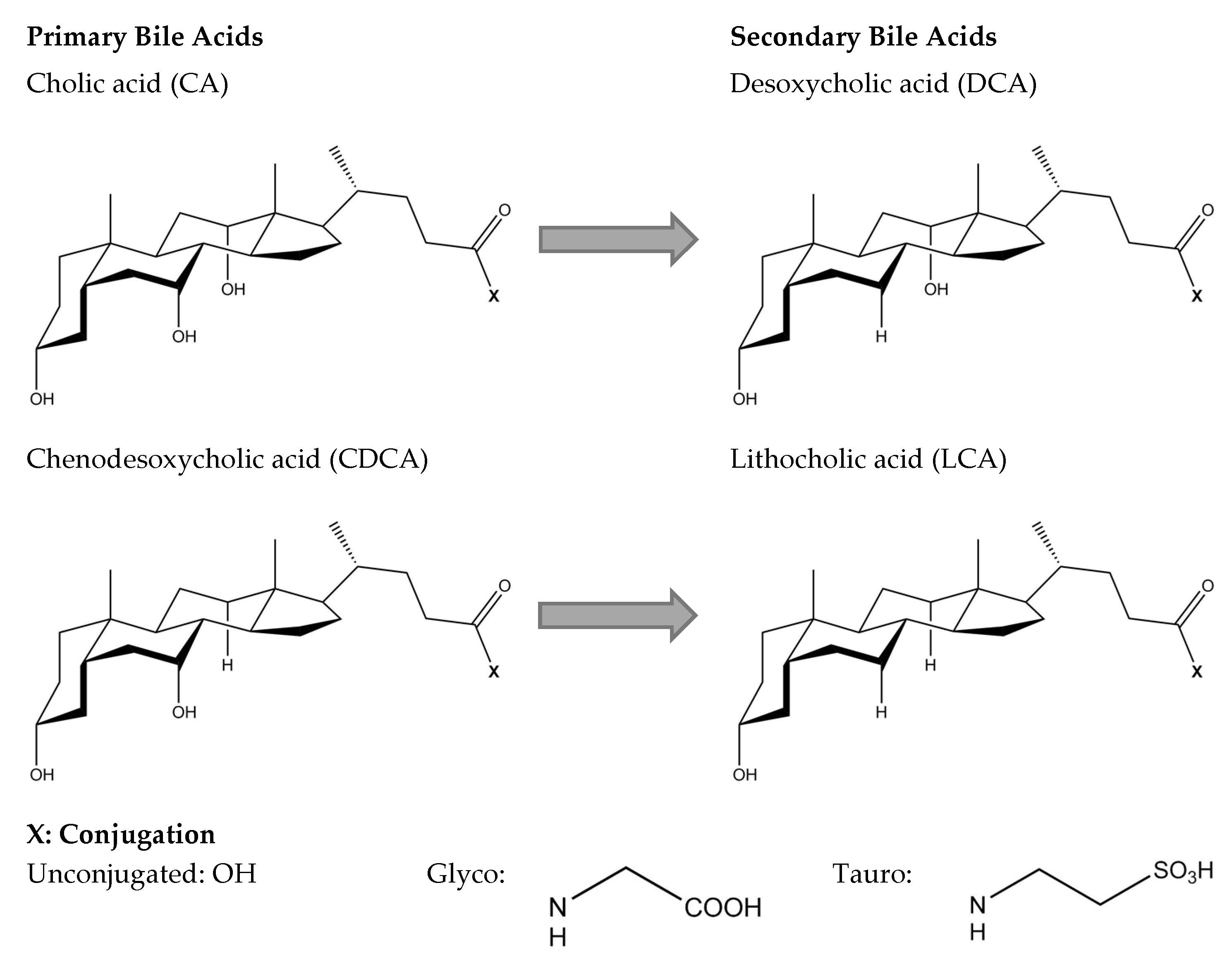
2. Bile Acid Metabolism and Chemistry
3. Principals and Mechanisms of Bile Acid Interactions
3.1. Bile Acid Interactions Related to Viscosity
3.2. Bile Acid Interactions on the Molecular Level
4. Analysis of Bile Acid Interactions
4.1. In Vivo Approaches to Study Bile Acid Interactions
4.2. In Vitro and Ex Vivo Approaches to Study Bile Acid Interactions
5. Bile Acid Interactions as Related to Plant Tissues
5.1. Interactions between Bile Acids and Dietary Fibres
5.2. Interactions between Bile Acids and Proteins
5.3. Interactions between Bile Acids and Phytochemicals
6. Bile Acid Interactions and Influences on Health
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CA | cholic acid |
| CDCA | chenodesoxycholic acid |
| CMC | critical micelle concentration |
| CYP7A1 | cholesterol 7α-hydroxylase |
| DCA | desoxycholic acid |
| EGCG | (−)-epigallocatechin-3-gallate |
| FXR | farnesoid X receptor |
| GLP-1 | glucagon-like peptide-1 |
| LCA | lithocholic acid |
| NMR | nuclear magnetic resonance |
References
- Di Ciaula, A.; Garruti, G.; Lunardi Baccetto, R.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.; Portincasa, P. Bile Acid Physiology. Ann. Hepatol. 2017, 16, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroen. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Metrani, R.; Shivanagoudra, S.R.; Jayaprakasha, G.K.; Patil, B.S. Review on Bile Acids: Effects of the Gut Microbiome, Interactions with Dietary Fiber, and Alterations in the Bioaccessibility of Bioactive Compounds. J. Agric. Food Chem. 2019, 67. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chiang, J.Y.L. Bile Acid Signaling in Metabolic Disease and Drug Therapy. Pharmacol. Rev. 2014, 66, 948–983. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef]
- Wolever, T.M.; Tosh, S.M.; Gibbs, A.L.; Brand-Miller, J.; Duncan, A.M.; Hart, V.; Lamarche, B.; Thomson, B.A.; Duss, R.; Wood, P.J. Physicochemical properties of oat β-glucan influence its ability to reduce serum LDL cholesterol in humans: A randomized clinical trial. Am. J. Clin. Nutr. 2010, 92, 723–732. [Google Scholar] [CrossRef]
- Kim, H.J.; White, P.J. In Vitro Bile-Acid Binding and Fermentation of High, Medium, and Low Molecular Weight β-Glucan. J. Agric. Food Chem. 2010, 58, 628–634. [Google Scholar] [CrossRef]
- Sayar, S.; Jannink, J.-L.; White, P.J. In Vitro Bile Acid Binding of Flours from Oat Lines Varying in Percentage and Molecular Weight Distribution of β-Glucan. J. Agric. Food Chem. 2005, 53, 8797–8803. [Google Scholar] [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Bader-Mittermaier, S.; Haller, D.; Eisner, P. Differentiation of adsorptive and viscous effects of dietary fibres on bile acid release by means of in vitro digestion and dialysis. Int. J. Mol. Sci. 2018, 19, 2193. [Google Scholar] [CrossRef]
- Macierzanka, A.; Torcello-Gómez, A.; Jungnickel, C.; Maldonado-Valderrama, J. Bile salts in digestion and transport of lipids. Adv. Colloid Interface Sci. 2019, 274, 102045. [Google Scholar] [CrossRef] [PubMed]
- Ellegard, L.; Andersson, H. Oat bran rapidly increases bile acid excretion and bile acid synthesis: An ileostomy study. Eur. J. Clin. Nutr. 2007, 61, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Lia, A.; Hallmans, G.; Sandberg, A.S.; Sundberg, B.; Aman, P.; Andersson, H. Oat beta-glucan increases bile acid excretion and a fiber-rich barley fraction increases cholesterol excretion in ileostomy subjects. Am. J. Clin. Nutr. 1995, 62, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Kahlon, T.S.; Smith, G.E. In vitro binding of bile acids by bananas, peaches, pineapple, grapes, pears, apricots and nectarines. Food Chem. 2007, 101, 1046–1051. [Google Scholar] [CrossRef]
- Dongowski, G. Interactions between dietary fibre-rich preparations and glycoconjugated bile acids in vitro. Food Chem. 2007, 104, 390–397. [Google Scholar] [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Eisner, P. Characterisation of the molecular interactions between primary bile acids and fractionated lupin cotyledons (Lupinus angustifolius L.). Food Chem. 2020, 323, 126780. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef]
- Shin, D.-J.; Wang, L. Bile Acid-Activated Receptors: A Review on FXR and Other Nuclear Receptors. In Bile Acids and Their Receptors; Fiorucci, S., Distrutti, E., Eds.; Springer: Cham, Switzerland, 2019; pp. 51–72. [Google Scholar]
- Russell, D.W.; Setchell, K.D. Bile acid biosynthesis. Biochemistry 1992, 31, 4737–4749. [Google Scholar] [CrossRef]
- Parker, R.; Rigby, N.M.; Ridout, M.J.; Gunning, A.P.; Wilde, P.J. The adsorption–desorption behaviour and structure function relationships of bile salts. Soft Matter 2014, 10, 6457–6466. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Maitra, U. Chemistry and biology of bile acids. Curr. Sci. 2004, 87, 1666–1683. [Google Scholar]
- Vlahcevic, R.Z.; Heuman, M.D.; Hylemon, B.P. Physiology and pathophysiology of enterohepatic circulation of bile acids. In Hepatology: A Textbook of Liver Desease, 3rd ed.; Zakim, D., Boyer, D.T., Eds.; Saunders: Philadelphia, PA, USA, 1996; pp. 376–417. [Google Scholar]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Hanafi, N.; Mohamed, A.; Sheikh Abdul Kadir, S.; Othman, M. Overview of Bile Acids Signaling and Perspective on the Signal of Ursodeoxycholic Acid, the Most Hydrophilic Bile Acid, in the Heart. Biomolecules 2018, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Torcello-Gómez, A.; Fernández Fraguas, C.; Ridout, M.J.; Woodward, N.C.; Wilde, P.J.; Foster, T.J. Effect of substituent pattern and molecular weight of cellulose ethers on interactions with different bile salts. Food Funct. 2015, 6, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Bosaeus, I.; Carlsson, N.G.; Sandberg, A.S.; Andersson, H. Effect of wheat bran and pectin on bile acid and cholesterol excretion in ileostomy patients. Hum. Nutr.-Clin. Nutr. 1986, 40, 429–440. [Google Scholar] [PubMed]
- Thandapilly, S.J.; Ndou, S.P.; Wang, Y.; Nyachoti, C.M.; Ames, N.P. Barley beta-glucan increases fecal bile acid excretion and short chain fatty acid levels in mildly hypercholesterolemic individuals. Food Funct. 2018, 9, 3092–3096. [Google Scholar] [CrossRef] [PubMed]
- Iaccarino, N.; Khakimov, B.; Mikkelsen, M.S.; Nielsen, T.S.; Jensen, M.G.; Randazzo, A.; Engelsen, S.B. Structurally different mixed linkage beta-glucan supplements differentially increase secondary bile acid excretion in hypercholesterolaemic rat faeces. Food Funct. 2020, 11, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Marasca, E.; Boulos, S.; Nystrom, L. Bile acid-retention by native and modified oat and barley beta-glucan. Carbohydr. Polym. 2020, 236, 116034. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, H.T.; Nielsen, M.S.; Christensen, N.J.; Christensen, U.; La Cour, T.V.; Motawia, M.S.; Jespersen, B.P.M.; Engelsen, S.B.; Møller, B.L. Molecular Interactions between Barley and Oat β-Glucans and Phenolic Derivatives. J. Agric. Food Chem. 2009, 57, 2056–2064. [Google Scholar] [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Eglmeier, J.; Haller, D.; Eisner, P. In vitro interactions of dietary fibre enriched food ingredients with primary and secondary bile acids. Nutrients 2019, 11, 1424. [Google Scholar] [CrossRef]
- Miller, C.C.; Walker, J. The Stokes-Einstein law for diffusion in solution. Proc. R. Soc. Lond. Ser. A Contain. Pap. Math. Phys. Character 1924, 106, 724–749. [Google Scholar] [CrossRef]
- Madenci, D.; Egelhaaf, S.U. Self-assembly in aqueous bile salt solutions. Curr. Opin. Colloid Interface Sci. 2010, 15, 109–115. [Google Scholar] [CrossRef]
- Heuman, D.M. Quantitative estimation of the hydrophilic-hydrophobic balance of mixed bile salt solutions. J. Lipid Res. 1989, 30, 719–730. [Google Scholar] [PubMed]
- Camire, M.E.; Zhao, J.; Violette, D.A. In vitro binding of bile acids by extruded potato peels. J. Agric. Food Chem. 1993, 41, 2391–2394. [Google Scholar] [CrossRef]
- Huth, M.; Dongowski, G.; Gebhardt, E.; Flamme, W. Functional Properties of Dietary Fibre Enriched Extrudates from Barley. J. Cereal Sci. 2000, 32, 115–128. [Google Scholar] [CrossRef]
- Drzikova, B.; Dongowski, G.; Gebhardt, E.; Habel, A. The composition of dietary fibre-rich extrudates from oat affects bile acid binding and fermentation in vitro. Food Chem. 2005, 90, 181–192. [Google Scholar] [CrossRef]
- Dziedzic, K.; Gorecka, D.; Szwengiel, A.; Smoczynska, P.; Czaczyk, K.; Komolka, P. Binding of bile acids by pastry products containing bioactive substances during in vitro digestion. Food Funct. 2015, 6, 1011–1020. [Google Scholar] [CrossRef]
- Zacherl, C.; Eisner, P.; Engel, K.-H. In vitro model to correlate viscosity and bile acid-binding capacity of digested water-soluble and insoluble dietary fibres. Food Chem. 2011, 126, 423–428. [Google Scholar] [CrossRef]
- Araki, Y.; Andoh, A.; Fujiyama, Y.; Kanauchi, O.; Takenaka, K.; Higuchi, A.; Bamba, T. Germinated Barley Foodstuff Exhibits Different Adsorption Properties for Hydrophilic versus Hydrophobic Bile Acids. Digestion 2001, 64, 248–254. [Google Scholar] [CrossRef]
- Bae, I.Y.; Kim, S.M.; Lee, S.; Lee, H.G. Effect of enzymatic hydrolysis on cholesterol-lowering activity of oat β-glucan. New Biotechnol. 2010, 27, 85–88. [Google Scholar] [CrossRef]
- Grundy, M.; Fardet, A.; Tosh, S.; Richa, G.T.; Wilde, P. Processing of oat: The impact on oat’s cholesterol lowering effect. Food Funct. 2018, 9, 1328–1343. [Google Scholar] [CrossRef]
- Gunness, P.; Michiels, J.; Vanhaecke, L.; De Smet, S.; Kravchuk, O.; Van de Meene, A.; Gidley, M.J. Reduction in circulating bile acid and restricted diffusion across the intestinal epithelium are associated with a decrease in blood cholesterol in the presence of oat β-glucan. FASEB J. 2016, 30, 4227–4238. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Williams, B.A.; Gerrits, W.J.J.; Bird, A.R.; Kravchuk, O.; Gidley, M.J. Circulating triglycerides and bile acids are reduced by a soluble wheat arabinoxylan via modulation of bile concentration and lipid digestion rates in a pig model. Mol. Nutr. Food Res. 2016, 60, 642–651. [Google Scholar] [CrossRef]
- Thakare, R.; Alamoudi, J.A.; Gautam, N.; Rodrigues, A.D.; Alnouti, Y. Species differences in bile acids I. Plasma and urine bile acid composition. J. Appl. Toxicol. 2018, 38, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dawson, P.A. Animal models to study bile acid metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Flanagan, B.M.; Shelat, K.; Gilbert, R.G.; Gidley, M.J. Kinetic analysis of bile salt passage across a dialysis membrane in the presence of cereal soluble dietary fibre polymers. Food Chem. 2012, 134, 2007–2013. [Google Scholar] [CrossRef]
- Li, X.; Jiao, W.; Zhang, W.; Xu, Y.; Cao, J.; Jiang, W. Characterizing the Interactions of Dietary Condensed Tannins with Bile Salts. J. Agric. Food Chem. 2019, 67, 9543–9550. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, E.; Bayramoglu, B. Characterization of the self-assembly and size dependent structural properties of dietary mixed micelles by molecular dynamics simulations. Biophys. Chem. 2019, 248, 16–27. [Google Scholar] [CrossRef]
- Drzikova, B.; Dongowski, G.; Gebhardt, E. Dietary fibre-rich oat-based products affect serum lipids, microbiota, formation of short-chain fatty acids and steroids in rats. Br. J. Nutr. 2005, 94, 1012–1025. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Chow, F.I. In Vitro Binding of Bile Acids by Rice Bran, Oat Bran, Wheat Bran, and Corn Bran. Cereal Chem. 2000, 77, 518–521. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Westerhout, J.; Wortelboer, H.; Verhoeckx, K. Ussing Chamber. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., Lopez-Exposito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 263–273. [Google Scholar]
- Mackie, A.; Rigby, N.; Harvey, P.; Bajka, B. Increasing dietary oat fibre decreases the permeability of intestinal mucus. J. Funct. Foods 2016, 26, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Flanagan, B.M.; Gidley, M.J. Molecular interactions between cereal soluble dietary fibre polymers and a model bile salt deduced from 13C NMR titration. J. Cereal Sci. 2010, 52, 444–449. [Google Scholar] [CrossRef]
- Ogawa, K.; Hirose, S.; Nagaoka, S.; Yanase, E. Interaction between Tea Polyphenols and Bile Acid Inhibits Micellar Cholesterol Solubility. J. Agric. Food Chem. 2016, 64, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Pigliacelli, C.; Belton, P.; Wilde, P.; Qi, S. Probing the molecular interactions between pharmaceutical polymeric carriers and bile salts in simulated gastrointestinal fluids using NMR spectroscopy. J. Colloid Interface Sci. 2019, 551, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Nishizawa, M.; Inoue, N.; Hosoya, T.; Yoshida, M.; Ukawa, Y.; Sagesaka, Y.M.; Doi, T.; Nakayama, T.; Kumazawa, S.; et al. Epigallocatechin Gallate Decreases the Micellar Solubility of Cholesterol via Specific Interaction with Phosphatidylcholine. J. Agric. Food Chem. 2014, 62, 2881–2890. [Google Scholar] [CrossRef]
- Gunness, P.; Flanagan, B.M.; Mata, J.P.; Gilbert, E.P.; Gidley, M.J. Molecular interactions of a model bile salt and porcine bile with (1,3:1,4)-β-glucans and arabinoxylans probed by 13C NMR and SAXS. Food Chem. 2016, 197, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Naumann, S.; Schweiggert-Weisz, U.; Martin, A.; Schuster, M.; Eisner, P. Effects of extrusion processing on the physiochemical and functional properties of lupin kernel fibre. Food Hydrocoll. 2021, 111, 106222. [Google Scholar] [CrossRef]
- Fuller, S.; Beck, E.; Salman, H.; Tapsell, L. New Horizons for the Study of Dietary Fiber and Health: A Review. Plant Foods Hum. Nutr. 2016, 71, 1–12. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; de Boeck, G.; Becker, K. Dietary Roles of Non-Starch Polysachharides in Human Nutrition: A Review. Crit. Rev. Food Sci. Nutr. 2012, 52, 899–935. [Google Scholar] [CrossRef]
- van Bennekum, A.M.; Nguyen, D.V.; Schulthess, G.; Hauser, H.; Phillips, M.C. Mechanisms of cholesterol-lowering effects of dietary insoluble fibres: Relationships with intestinal and hepatic cholesterol parameters. Br. J. Nutr. 2005, 94, 331–337. [Google Scholar] [CrossRef]
- Ul Ain, H.B.; Saeed, F.; Ahmed, A.; Khan, M.A.; Niaz, B.; Tufail, T. Improving the physicochemical properties of partially enhanced soluble dietary fiber through innovative techniques: A coherent review. J. Food Process. Preserv. 2019, 43, 12. [Google Scholar] [CrossRef]
- Cookson, F.B.; Altschul, R.; Fedor-off, S. The effects of alfalfa on serum cholesterol and in modifying or preventing cholesterol-induced athero-sclerosis in rabbits. J. Atheroscler. Res. 1967, 7, 69–81. [Google Scholar] [CrossRef]
- Ghaffarzadegan, T.; Marungruang, N.; Fåk, F.; Nyman, M. Molecular Properties of Guar Gum and Pectin Modify Cecal Bile Acids, Microbiota, and Plasma Lipopolysaccharide-Binding Protein in Rats. PLoS ONE 2016, 11, e0157427. [Google Scholar] [CrossRef] [PubMed]
- Dongowski, G. Influence of pectin structure on the interaction with bile acids under in vitro conditions. Z. Lebensm. Unters. Forsch. 1995, 201, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Dongowski, G. Effect of pH on the in vitro interactions between bile acids and pectin. Z. Lebensm. Unters. Forsch. 1997, 205, 185–192. [Google Scholar] [CrossRef]
- Pfeffer, P.E.; Doner, L.W.; Hoagland, P.D.; McDonald, G.G. Molecular interactions with dietary fiber components. Investigation of the possible association of pectin and bile acids. J. Agric. Food Chem. 1981, 29, 455–461. [Google Scholar] [CrossRef]
- Lopez-Pena, C.; Arroyo-Maya, I.J.; McClements, D.J. Interaction of a bile salt (sodium taurocholate) with cationic (ε-polylysine) and anionic (pectin) biopolymers under simulated gastrointestinal conditions. Food Hydrocoll. 2019, 87, 352–359. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Woodruff, C.L. In Vitro Binding of Bile Acids by Rice Bran, Oat Bran, Barley and β-Glucan Enriched Barley. Cereal Chem. 2003, 80, 260–263. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, C.; Ou, S. In vitro binding capacities of three dietary fibers and their mixture for four toxic elements, cholesterol, and bile acid. J. Hazard. Mater. 2011, 186, 236–239. [Google Scholar] [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Haller, D.; Eisner, P. Retention of primary bile acids by lupin cell wall polysaccharides under in vitro digestion conditions. Nutrients 2019, 11, 2117. [Google Scholar] [CrossRef]
- Weickert, M.O.; Hattersley, J.G.; Kyrou, I.; Arafat, A.M.; Rudovich, N.; Roden, M.; Nowotny, P.; von Loeffelholz, C.; Matysik, S.; Schmitz, G.; et al. Effects of supplemented isoenergetic diets varying in cereal fiber and protein content on the bile acid metabolic signature and relation to insulin resistance. Nutr. Diabetes 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Weickert, M.O.; Roden, M.; Isken, F.; Hoffmann, D.; Nowotny, P.; Osterhoff, M.; Blaut, M.; Alpert, C.; Gögebakan, Ö.; Bumke-Vogt, C.; et al. Effects of supplemented isoenergetic diets differing in cereal fiber and protein content on insulin sensitivity in overweight humans. Am. J. Clin. Nutr. 2011, 94, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, M.A.; Hamilton, D. Studies on the adsorption of bile salts to non-absorbed components of diet. Biochim. Biophys. Acta Mol. Basis Dis. 1968, 152, 165–173. [Google Scholar] [CrossRef]
- Gallaher, D.; Schneeman, B.O. Intestinal interaction of bile acids, phospholipids, dietary fibers, and cholestyramine. Am. J. Physiol. Gastrointest. Liver Physiol. 1986, 250, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Górecka, D.; Korczak, J.; Balcerowski, E.; Decyk, K. Sorption of bile acids and cholesterol by dietary fiber of carrots, cabbage and apples. Electron. J. Pol. Agric. Univ. 2002, 5. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.459.4177&rep=rep1&type=pdf (accessed on 17 August 2020).
- Kritchevsky, D.; Story, J.A. Comparison of the Binding of Various Bile Acids and Bile Salts in Vitro by Several Types of Fiber. J. Nutr. 1976, 106, 1292–1294. [Google Scholar] [CrossRef]
- Sayar, S.; Jannink, J.-L.; White, P.J. In Vitro Bile Acid Binding Activity within Flour Fractions from Oat Lines with Typical and High β-Glucan Amounts. J. Agric. Food Chem. 2006, 54, 5142–5148. [Google Scholar] [CrossRef][Green Version]
- American Association of Cereal Chemists. The definition of dietary fiber (report of the dietary fiber definition committee to the board of directors of the American Association of Cereal Chemists). Cereal Foods World 2001, 48, 112–126. [Google Scholar]
- Funk, C.; Grabber, J.H.; Steinhart, H.; Bunzel, M. Artificial Lignification of Maize Cell Walls Does Not Affect In Vitro Bile Acid Adsorption. Cereal Chem. 2008, 85, 14–18. [Google Scholar] [CrossRef]
- Elhardallou, S.B. The bile acids binding of the fibre-rich fractions of three starchy legumes. Plant Foods Hum. Nutr. 1992, 42, 207–218. [Google Scholar] [CrossRef]
- Kahlon, T.S.; Woodruff, C.L. In vitro binding of bile acids by soy protein, pinto beans, black beans and wheat gluten. Food Chem. 2002, 79, 425–429. [Google Scholar] [CrossRef]
- Yoshie-Stark, Y.; Wäsche, A. In vitro binding of bile acids by lupin protein isolates and their hydrolysates. Food Chem. 2004, 88, 179–184. [Google Scholar] [CrossRef]
- Barbana, C.; Boucher, A.C.; Boye, J.I. In vitro binding of bile salts by lentil flours, lentil protein concentrates and lentil protein hydrolysates. Food Res. 2011, 44, 174–180. [Google Scholar] [CrossRef]
- Higaki, N.; Sato, K.; Suda, H.; Suzuka, T.; Komori, T.; Saeki, T.; Nakamura, Y.; Ohtsuki, K.; Iwami, K.; Kanamoto, R. Evidence for the existence of a soybean resistant protein that captures bile acid and stimulates its fecal excretion. Biosci. Biotechnol. Biochem. 2006, 70, 2844–2852. [Google Scholar] [CrossRef]
- Bosaeus, I.; Sandström, B.; Andersson, H. Bile acid and cholesterol excretion in human beings given soya-bean-and meat-protein-based diets: A study in ileostomy subjects. Br. J. Nutr. 1988, 59, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fan, M. Interaction behaviors and structural characteristics of zein/NaTC nanoparticles. RSC Adv. 2019, 9, 5748–5755. [Google Scholar] [CrossRef]
- McDougall, G.J.; Allwood, J.W.; Pereira-Caro, G.; Brown, E.M.; Ternan, N.; Verrall, S.; Stewart, D.; Lawther, R.; O’Connor, G.; Rowland, I.; et al. Nontargeted LC-MS(n) Profiling of Compounds in Ileal Fluids That Decrease after Raspberry Intake Identifies Consistent Alterations in Bile Acid Composition. J. Nat. Prod. 2016, 79, 2606–2615. [Google Scholar] [CrossRef]
- Fotschki, B.; Juskiewicz, J.; Jurgonski, A.; Rigby, N.; Sojka, M.; Kolodziejczyk, K.; Mackie, A.; Zdunczyk, Z. Raspberry pomace alters cecal microbial activity and reduces secondary bile acids in rats fed a high-fat diet. J. Nutr. Biochem. 2017, 46, 13–20. [Google Scholar] [CrossRef]
- Huang, J.; Feng, S.; Liu, A.; Dai, Z.; Wang, H.; Reuhl, K.; Lu, W.; Yang, C.S. Green Tea Polyphenol EGCG Alleviates Metabolic Abnormality and Fatty Liver by Decreasing Bile Acid and Lipid Absorption in Mice. Mol. Nutr. Food Res. 2018, 62, 1700696. [Google Scholar] [CrossRef]
- Sembries, S.; Dongowski, G.; Mehrländer, K.; Will, F.; Dietrich, H. Physiological Effects of Extraction Juices from Apple, Grape, and Red Beet Pomaces in Rats. J. Agric. Food Chem. 2006, 54, 10269–10280. [Google Scholar] [CrossRef]
- Hosoyamada, Y.; Yamada, M. Effects of Dietary Fish Oil and Apple Polyphenol on the Concentration Serum Lipids and Excretion of Fecal Bile Acids in Rats. J. Nutr. Sci. Vitaminol. 2017, 63, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Quifer-Rada, P.; Choy, Y.Y.; Calvert, C.C.; Waterhouse, A.L.; Lamuela-Raventos, R.M. Use of metabolomics and lipidomics to evaluate the hypocholestreolemic effect of Proanthocyanidins from grape seed in a pig model. Mol. Nutr. Food Res. 2016, 60, 2219–2227. [Google Scholar] [CrossRef] [PubMed]
- Olivero-David, R.; Ruiz-Roso, M.B.; Caporaso, N.; Perez-Olleros, L.; De Las Heras, N.; Lahera, V.; Ruiz-Roso, B. In vivo bioavailability of polyphenols from grape by-product extracts, and effect on lipemia of normocholesterolemic Wistar rats. J. Sci. Food Agric. 2018, 98, 5581–5590. [Google Scholar] [CrossRef] [PubMed]
- Visavadiya, N.P.; Narasimhacharya, A.V. Asparagus root regulates cholesterol metabolism and improves antioxidant status in hypercholesteremic rats. Evid.-Based Complement. Altern. Med. 2009, 6, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Inoue, N.; Shimizu-Ibuka, A.; Tadaishi, M.; Takita, T.; Arai, S.; Mura, K. Serum cholesterol reduction by feeding a high-cholesterol diet containing a lower-molecular-weight polyphenol fraction from peanut skin. Biosci. Biotechnol. Biochem. 2012, 76, 834–837. [Google Scholar] [CrossRef]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Granados, O.; Torre-Villalvazo, I.; Serna-Saldivar, S.O.; Torres, N.; Palacios-González, B.; Tovar, A.R. Flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats modulate lipid metabolism and biliary cholesterol secretion in C57BL/6 mice. Br. J. Nutr. 2014, 112, 886–899. [Google Scholar] [CrossRef]
- Zhao, D. Challenges associated with elucidating the mechanisms of the hypocholesterolaemic activity of saponins. J. Funct. Foods 2016, 23, 52–65. [Google Scholar] [CrossRef]
- Ikeda, I.; Yamahira, T.; Kato, M.; Ishikawa, A. Black-tea polyphenols decrease micellar solubility of cholesterol in vitro and intestinal absorption of cholesterol in rats. J. Agric. Food Chem. 2010, 58, 8591–8595. [Google Scholar] [CrossRef]
- Ngamukote, S.; Mäkynen, K.; Thilawech, T.; Adisakwattana, S. Cholesterol-lowering activity of the major polyphenols in grape seed. Molecules 2011, 16, 5054–5061. [Google Scholar] [CrossRef]
- Raederstorff, D.G.; Schlachter, M.F.; Elste, V.; Weber, P. Effect of EGCG on lipid absorption and plasma lipid levels in rats. J. Nutr. Biochem. 2003, 14, 326–332. [Google Scholar] [CrossRef]
- Shishikura, Y.; Khokhar, S.; Murray, B.S. Effects of Tea Polyphenols on Emulsification of Olive Oil in a Small Intestine Model System. J. Agric. Food Chem. 2006, 54, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.F.; Jayaprakasha, G.K.; Patil, B.S. In Vitro Bile Acid Binding Capacities of Red Leaf Lettuce and Cruciferous Vegetables. J. Agric. Food Chem. 2017, 65, 8054–8062. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.; Jayaprakasha, G.K.; Patil, B. In vitro digestion with bile acids enhances the bioaccessibility of kale polyphenols. Food Funct. 2018, 9, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Hamauzu, Y.; Suwannachot, J. Non-extractable polyphenols and in vitro bile acid-binding capacity of dried persimmon (Diospyros kaki) fruit. Food Chem. 2019, 293, 127–133. [Google Scholar] [CrossRef]
- Ushiroda, C.; Naito, Y.; Takagi, T.; Uchiyama, K.; Mizushima, K.; Higashimura, Y.; Yasukawa, Z.; Okubo, T.; Inoue, R.; Honda, A.; et al. Green tea polyphenol (epigallocatechin-3-gallate) improves gut dysbiosis and serum bile acids dysregulation in high-fat diet-fed mice. J. Clin. Biochem. Nutr. 2019, 65, 34–46. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, L.; Shen, C.L.; Wang, J.S. Green tea polyphenols modify gut-microbiota dependent metabolisms of energy, bile constituents and micronutrients in female Sprague-Dawley rats. J. Nutr. Biochem. 2018, 61, 68–81. [Google Scholar] [CrossRef]
- Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef]
- Chen, M.L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.D.; Zhang, Q.Y.; Mi, M.T. Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. mBio 2016, 7, e02210–e02215. [Google Scholar] [CrossRef]
- Lairon, D. Dietary Fibres and Dietary Lipids. In Advanced Dietary Fibre Technology; McCleary, B.V., Prosky, L., Eds.; Blackwell Science: Malden, MA, USA, 2001; pp. 177–185. [Google Scholar]
- Stamler, J.; Daviglus, M.L.; Garside, D.B.; Dyer, A.R.; Greenland, P.; Neaton, J.D. Relationship of baseline serum cholesterol levels in 3 large cohorts of younger men to long-term coronary, cardiovascular, and all-cause mortality and to longevity. JAMA 2000, 284, 311–318. [Google Scholar] [CrossRef]
- Parolini, C.; Manzini, S.; Busnelli, M.; Rigamonti, E.; Marchesi, M.; Diani, E.; Sirtori, C.R.; Chiesa, G. Effect of the combinations between pea proteins and soluble fibres on cholesterolaemia and cholesterol metabolism in rats. Br. J. Nutr. 2013, 110, 1394–1401. [Google Scholar] [CrossRef]
- Feng, K.; Lan, Y.; Zhu, X.; Li, J.; Chen, T.; Huang, Q.; Ho, C.T.; Chen, Y.; Cao, Y. Hepatic Lipidomics Analysis Reveals the Anti-obesity and Cholesterol-lowering Effects of Tangeretin in High-Fat Diet-Fed Rats. J. Agric. Food Chem. 2020, 68. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.A.; Luo, G.; Billin, A.N.; Bisi, J.; McNeill, Y.Y.; Kozarsky, K.F.; Donahee, M.; Wang, D.Y.; Mansfield, T.A.; Kliewer, S.A.; et al. Definition of a novel growth factor-dependent signal cascade for the suppression of bile acid biosynthesis. Genes Dev. 2003, 17, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of Bile Acids and Bile Acid Receptors in Metabolic Regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef]
- Kuipers, F.; Groen, A.K. FXR: The key to benefits in bariatric surgery? Nat. Med. 2014, 20, 337–338. [Google Scholar] [CrossRef] [PubMed]
- Calderon, G.; McRae, A.; Rievaj, J.; Davis, J.; Zandvakili, I.; Linker-Nord, S.; Burton, D.; Roberts, G.; Reimann, F.; Gedulin, B.; et al. Ileo-colonic delivery of conjugated bile acids improves glucose homeostasis via colonic GLP-1-producing enteroendocrine cells in human obesity and diabetes. EBioMedicine 2020, 55, 102759. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Ung, T.T.; Kim, N.H.; Jung, Y.D. Role of bile acids in colon carcinogenesis. World J. Clin. Cases 2018, 6, 577–588. [Google Scholar] [CrossRef]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Vasavan, T.; Ferraro, E.; Ibrahim, E.; Dixon, P.; Gorelik, J.; Williamson, C. Heart and bile acids—Clinical consequences of altered bile acid metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1345–1355. [Google Scholar] [CrossRef]
- Dosch, A.R.; Imagawa, D.K.; Jutric, Z. Bile Metabolism and Lithogenesis: An Update. Surg. Clin. N. Am. 2019, 99, 215–229. [Google Scholar] [CrossRef]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary bile acids and short chain fatty acids in the colon: A focus on colonic microbiome, cell proliferation, inflammation, and cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bile Acid | CMC (mM) 1 | Nagg 1 | Hydrophobicity 2 |
|---|---|---|---|
| Glycocholic acid (GCA) | 4 | 9 |  |
| Taurocholic acid (TCA) | 3–18 | 3–7 | |
| Glycochenodesoxycholic acid (GCDCA) | 1–2 | 15 | |
| Taurochenodesoxycholic acid (TCDCA) | 0.9–7 | 5–26 | |
| Glycodesoxycholic acid (GDCA) | 1–2 | 13–16 | |
| Taurodesoxycholic acid (TDCA) | 2–3 | 12–19 |
| Approach to Study Bile Acid Interactions | Details | Benefits | Limitations | Reference |
|---|---|---|---|---|
| Human studies | Bile acid analysis of faecal samples |
|
| [2,31] |
| Human ileostomy studies | Bile acid analysis of ileal contents |
|
| [12,26] |
| Animal models | Bile acid analysis of contents of intestinal sites or faecal samples |
|
| [6,43] |
| In vitro models based on centrifugation | Bile acid analysis in supernatant |
|
| [8,14] |
| In vitro models based on dialysis | Bile acid transport across a dialysis membrane |
|
| [10,47] |
| Structural in vitro techniques | Nuclear magnetic resonance, microcalorimetry, etc. |
|
| [5,48] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naumann, S.; Haller, D.; Eisner, P.; Schweiggert-Weisz, U. Mechanisms of Interactions between Bile Acids and Plant Compounds—A Review. Int. J. Mol. Sci. 2020, 21, 6495. https://doi.org/10.3390/ijms21186495
Naumann S, Haller D, Eisner P, Schweiggert-Weisz U. Mechanisms of Interactions between Bile Acids and Plant Compounds—A Review. International Journal of Molecular Sciences. 2020; 21(18):6495. https://doi.org/10.3390/ijms21186495
Chicago/Turabian StyleNaumann, Susanne, Dirk Haller, Peter Eisner, and Ute Schweiggert-Weisz. 2020. "Mechanisms of Interactions between Bile Acids and Plant Compounds—A Review" International Journal of Molecular Sciences 21, no. 18: 6495. https://doi.org/10.3390/ijms21186495
APA StyleNaumann, S., Haller, D., Eisner, P., & Schweiggert-Weisz, U. (2020). Mechanisms of Interactions between Bile Acids and Plant Compounds—A Review. International Journal of Molecular Sciences, 21(18), 6495. https://doi.org/10.3390/ijms21186495