Pemafibrate Protects Against Retinal Dysfunction in a Murine Model of Diabetic Retinopathy

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
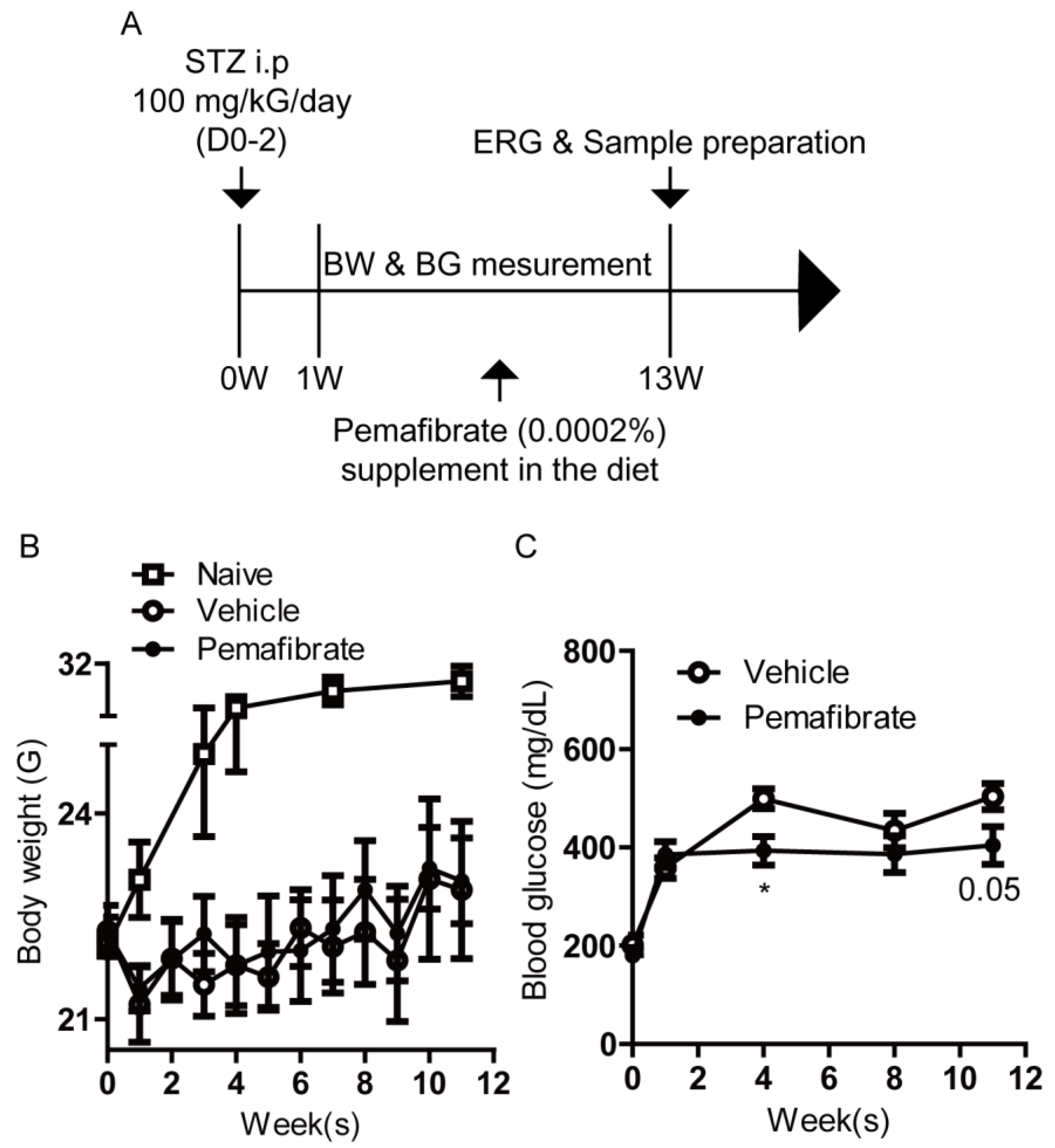
2.1. Pemafibrate Improves Diabetic Condition in an STZ-Induced Diabetic Mouse Model
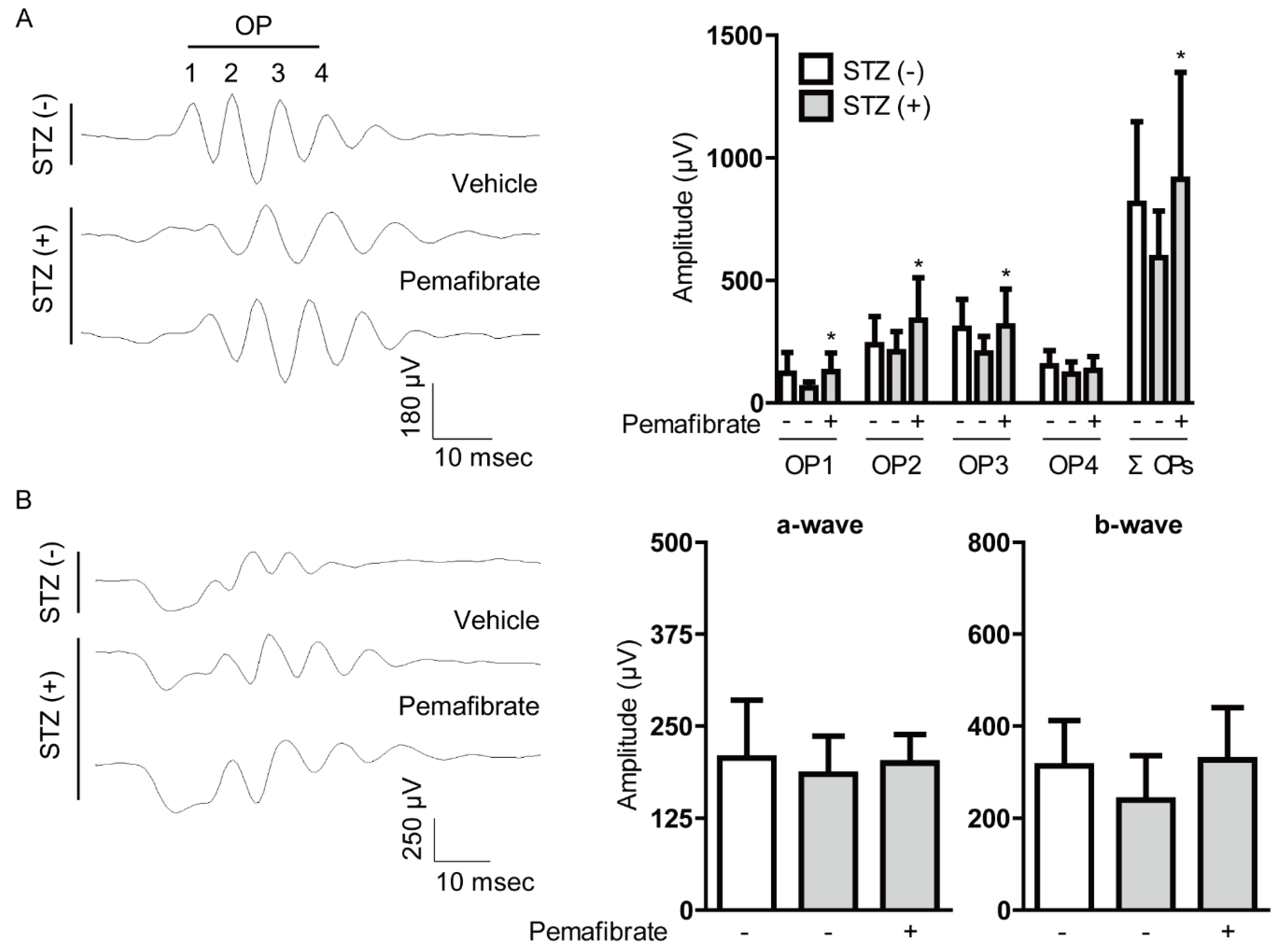
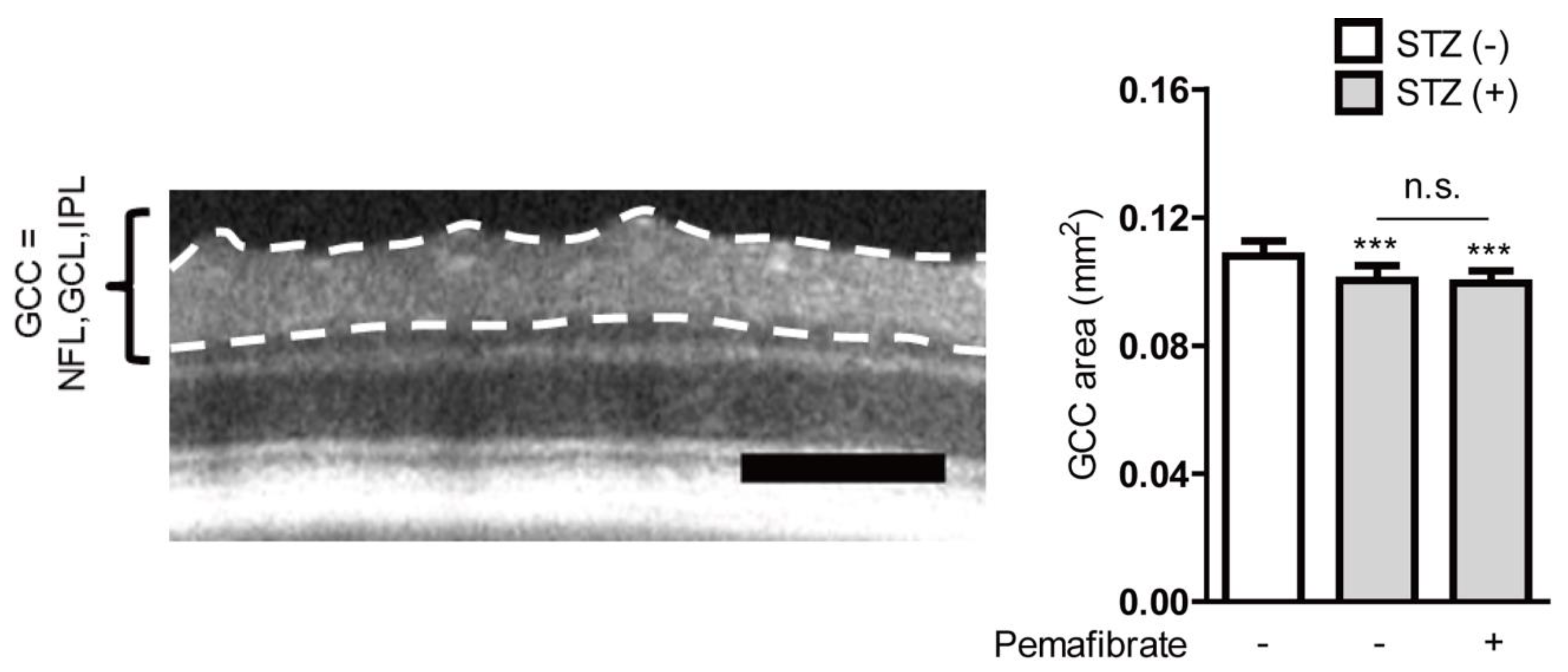
2.2. Pemafibrate Suppresses Visual Dysfunction in an STZ-Induced Diabetic Mouse Model
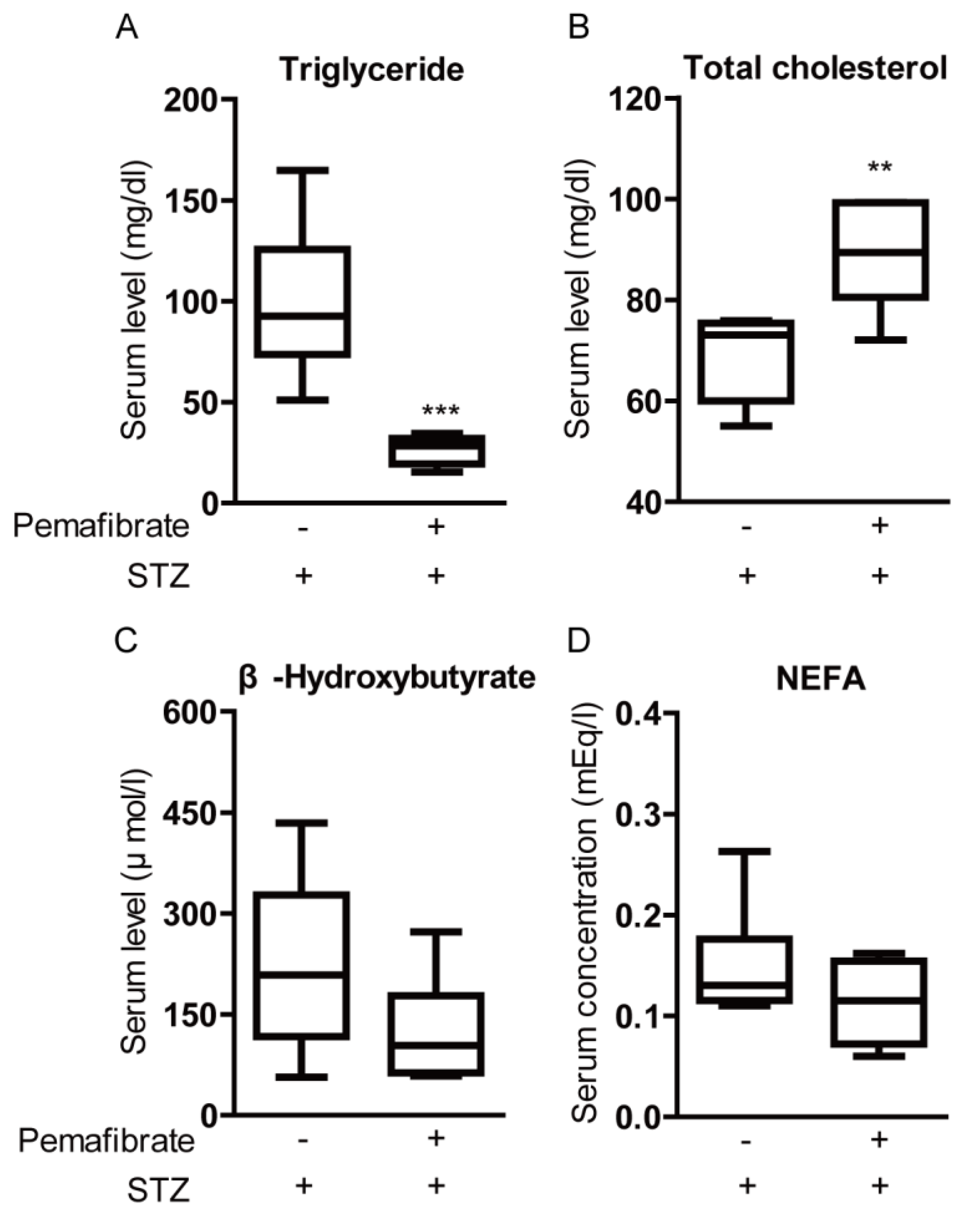
2.3. Pemafibrate Improves Lipid Metabolism in an STZ-Induced Diabetic Mouse Model
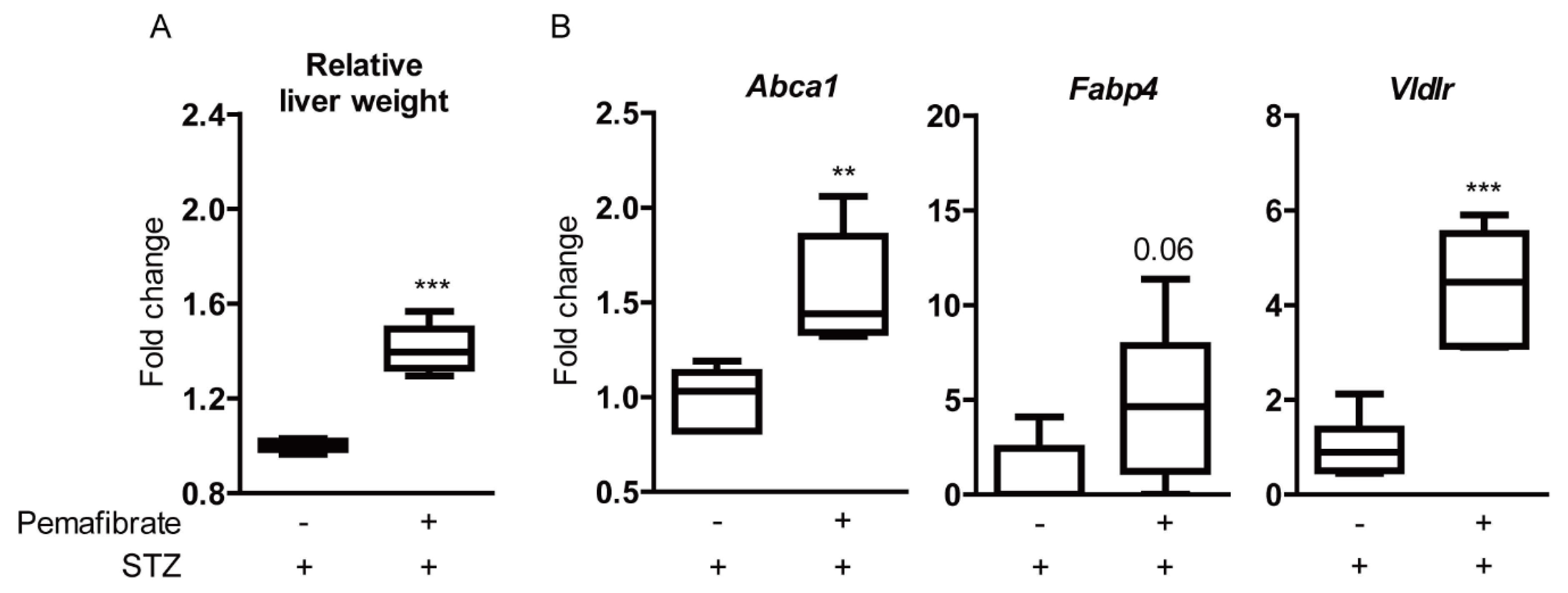
2.4. Pemafibrate Boosts Liver Function and Promotes Gene Expression of PPARα Downstream in an STZ-Induced Diabetic Mouse Model
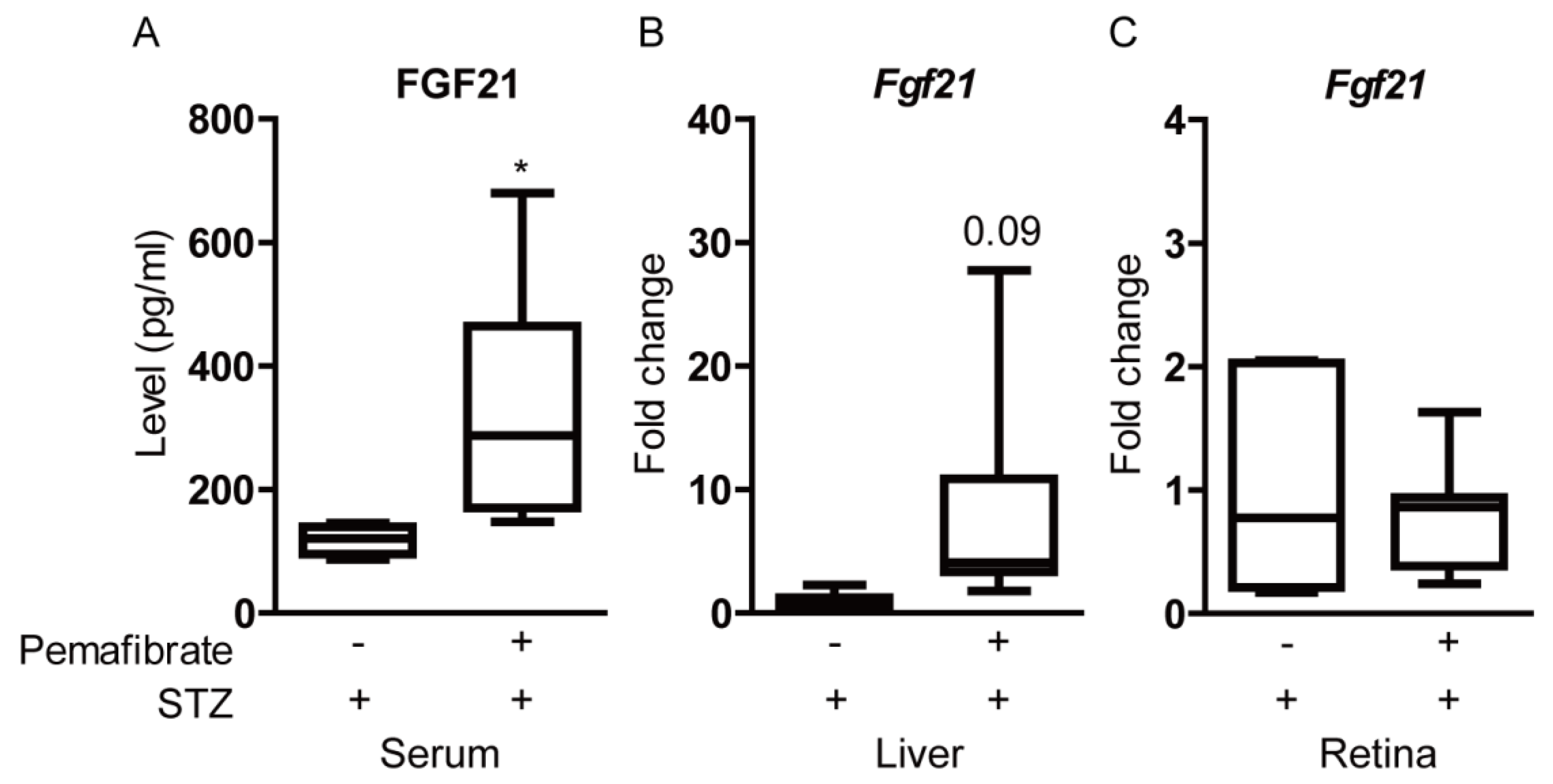
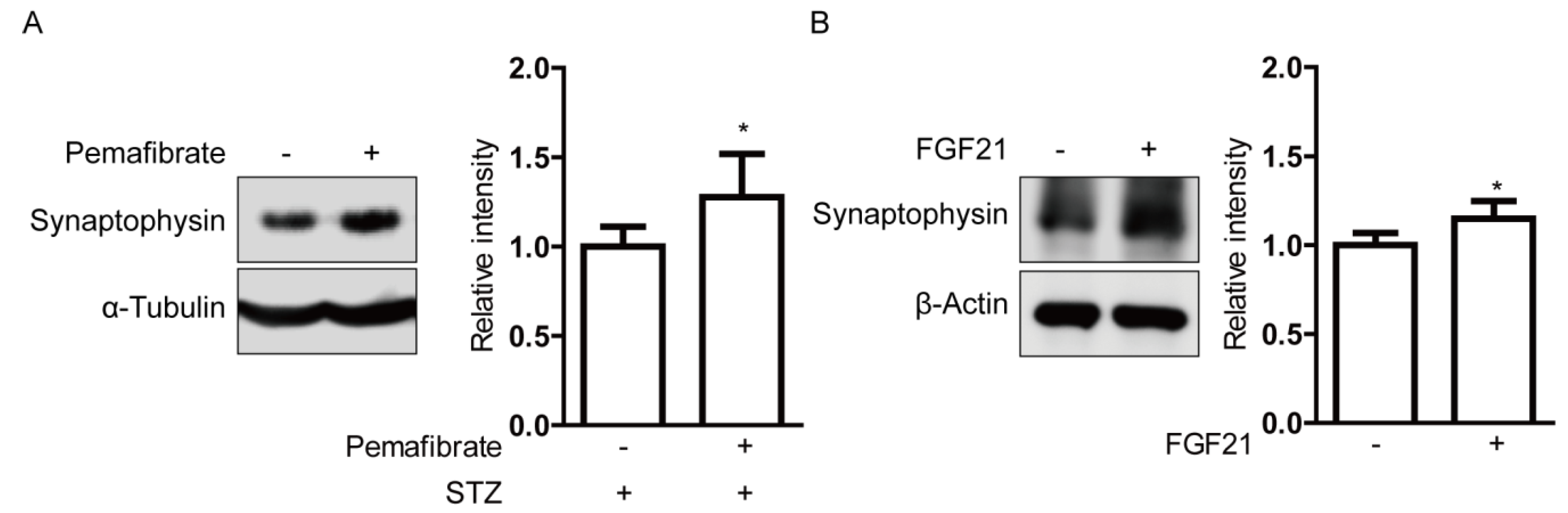
2.5. Pemafibrate Increases Serum FGF21 Level and Retinal Synaptophysin Expression in an STZ-Induced Diabetic Mouse Model
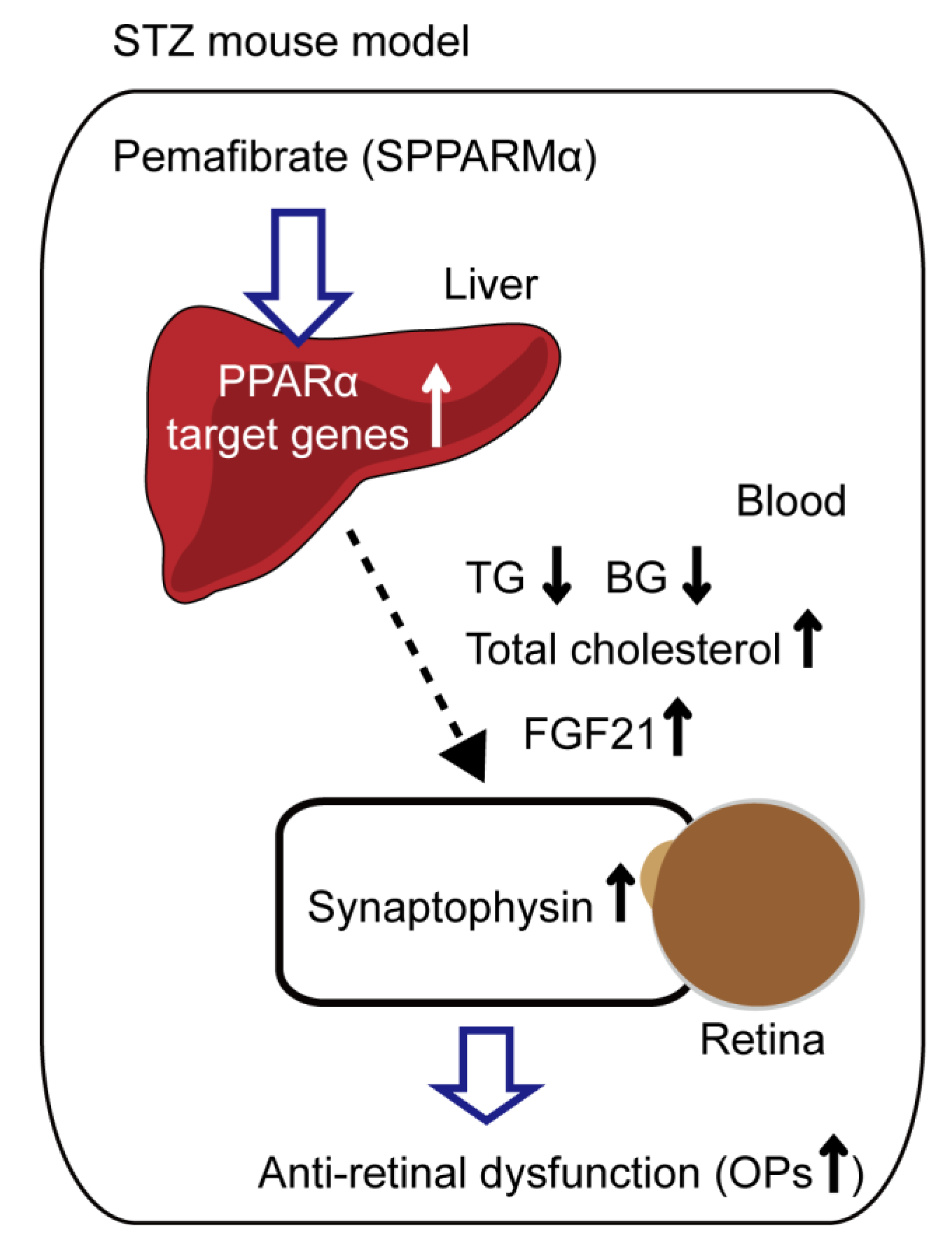
3. Discussion
4. Materials and Methods
4.1. Animal
4.2. Cell Line
4.3. STZ-Induced DR Model and Administration of Pemafibrate
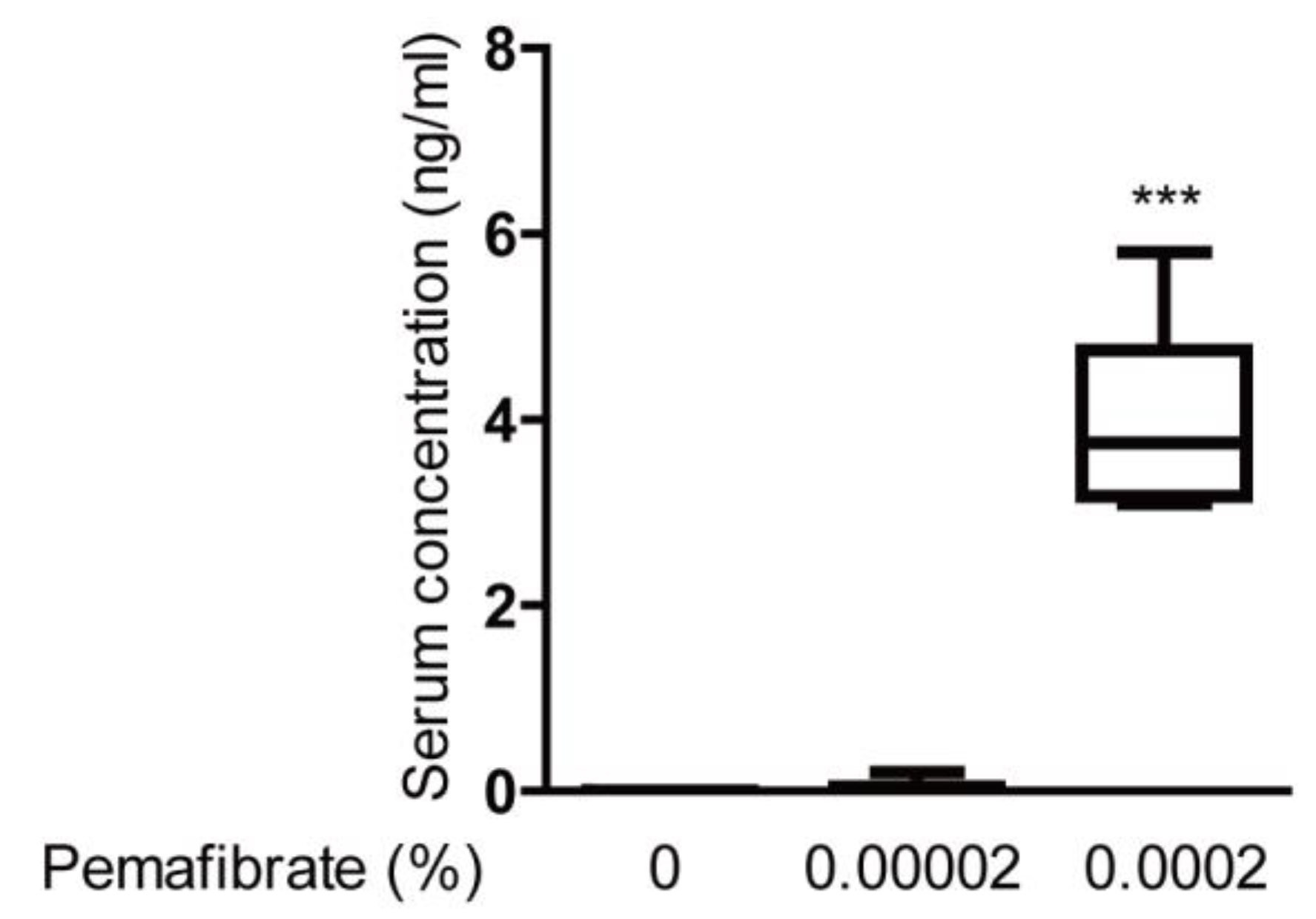
4.4. Pemafibrate Detection
4.5. Electroretinography (ERG)
4.6. Metabolic Measurements
4.7. Quantitative PCR
4.8. Western Blotting
4.9. Optical Coherence Tomography (OCT)
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DR | Diabetic retinopathy |
| ERG | Electroretinography |
| FGF21 | Fibroblast growth factor 21 |
| NEFA | Non-esterified fatty acids |
| OCT | Optical coherence tomography |
| OP | Oscillatory potential |
| PPARα | Peroxisome proliferator-activated receptor alpha |
| PC12D cells | Pheochromocytoma cells |
| STZ | Streptozotocin |
| SPPARMα | Selective peroxisome proliferator-activated receptor alpha modulator |
| VEGF | Vascular endothelial growth factor |
Appendix A

Appendix B

References
- Zheng, Y.; He, M.; Congdon, N. The worldwide epidemic of diabetic retinopathy. Indian J. Ophtalmol. 2012, 60, 428–431. [Google Scholar] [CrossRef]
- Fogli, S.; Del Re, M.; Rofi, E.; Posarelli, C.; Figus, M.; Danesi, R. Clinical pharmacology of intravitreal anti-VEGF drugs. Eye 2018, 32, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Pescosolido, N.; Barbato, A.; Stefanucci, A.; Buomprisco, G. Role of Electrophysiology in the Early Diagnosis and Follow-Up of Diabetic Retinopathy. J. Diabetes Res. 2015, 2015, 319692. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Masuda, D.; Matsuzawa, Y. Pemafibrate, a New Selective PPARα Modulator: Drug Concept and Its Clinical Applications for Dyslipidemia and Metabolic Diseases. Curr. Atheroscler. Rep. 2020, 22, 5. [Google Scholar] [CrossRef] [PubMed]
- Kusunoki, M.; Natsume, Y.; Oshida, Y.; Miyata, T. 615-P: A Novel Lipid-Lowering Drug Pemafibrate Enhances Effects of Statins in Serum Lipids in Japanese Patients with Type 2 Diabetes. Diabetes 2019, 68. [Google Scholar] [CrossRef]
- Raza-Iqbal, S.; Tanaka, T.; Anai, M.; Inagaki, T.; Matsumura, Y.; Ikeda, K.; Taguchi, A.; Gonzalez, F.J.; Sakai, J.; Kodama, T. Transcriptome Analysis of K-877 (a Novel Selective PPARα Modulator (SPPARMα))-Regulated Genes in Primary Human Hepatocytes and the Mouse Liver. J. Atheroscler. Thromb. 2015, 22, 754–772. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Ozawa, N.; Miwa, Y.; Ishida, A.; Ohta, M.; Tsubota, K.; Kurihara, T. Pemafibrate Prevents Retinal Pathological Neovascularization by Increasing FGF21 Level in a Murine Oxygen-Induced Retinopathy Model. Int. J. Mol. Sci. 2019, 20, 5878. [Google Scholar] [CrossRef]
- Tezze, C.; Romanello, V.; Sandri, M. FGF21 as Modulator of Metabolism in Health and Disease. Front. Physiol. 2019, 10, 419. [Google Scholar] [CrossRef]
- Zibar, K.; Blaslov, K.; Bulum, T.; Ćuća, J.K.; Smirčić-Duvnjak, L. Basal and postprandial change in serum fibroblast growth factor-21 concentration in type 1 diabetic mellitus and in healthy controls. Endocrine 2015, 48, 848–855. [Google Scholar] [CrossRef]
- Xiao, Y.; Xu, A.; Law, L.S.; Chen, C.; Li, H.; Li, X.; Yang, L.; Liu, S.; Zhou, Z.; Lam, K.S. Distinct changes in serum fibroblast growth factor 21 levels in different subtypes of diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E54–E58. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, C.; Xin, Y.; Huang, Z.; Tan, Y.; Huang, Y.; Wang, Y.; Feng, W.; Li, X.; Li, W.; et al. Protective effect of FGF21 on type 1 diabetes-induced testicular apoptotic cell death probably via both mitochondrial- and endoplasmic reticulum stress-dependent pathways in the mouse model. Toxicol. Lett. 2013, 219, 65–76. [Google Scholar] [CrossRef]
- Zhang, C.; Shao, M.; Yang, H.; Chen, L.; Yu, L.; Cong, W.; Tian, H.; Zhang, F.; Cheng, P.; Jin, L.; et al. Attenuation of hyperlipidemia- and diabetes-induced early-stage apoptosis and late-stage renal dysfunction via administration of fibroblast growth factor-21 is associated with suppression of renal inflammation. PLoS ONE 2013, 8, e82275. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, J.; Guo, W.; Li, F.; Sun, W.; Chen, J.; Zhang, C.; Lu, X.; Tan, Y.; Feng, W.; et al. Up-regulation of Nrf2 is involved in FGF21-mediated fenofibrate protection against type 1 diabetic nephropathy. Free Radic. Biol. Med. 2016, 93, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Gong, Y.; Liegl, R.; Wang, Z.; Liu, C.H.; Meng, S.S.; Burnim, S.B.; Saba, N.J.; Fredrick, T.W.; Morss, P.C.; et al. FGF21 Administration Suppresses Retinal and Choroidal Neovascularization in Mice. Cell Rep. 2017, 18, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Wang, Z.; Liu, C.H.; Gong, Y.; Cakir, B.; Liegl, R.; Sun, Y.; Meng, S.S.; Burnim, S.B.; Arellano, I.; et al. Fibroblast Growth Factor 21 Protects Photoreceptor Function in Type 1 Diabetic Mice. Diabetes 2018, 67, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Fu, Z.; Wang, Z.; Cakir, B.; Cho, S.S.; Britton, W.; Sun, Y.; Hellström, A.; Talukdar, S.; Smith, L.E.H. Long-Acting FGF21 Inhibits Retinal Vascular Leakage in In Vivo and In Vitro Models. Int. J. Mol. Sci. 2020, 21, 1188. [Google Scholar] [CrossRef]
- Staiger, H.; Keuper, M.; Berti, L.; Hrabe de Angelis, M.; Häring, H.U. Fibroblast Growth Factor 21-Metabolic Role in Mice and Men. Endocr. Rev. 2017, 38, 468–488. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70. [Google Scholar] [CrossRef]
- Shirao, Y.; Kawasaki, K. Electrical responses from diabetic retina. Prog. Retin. Eye Res. 1998, 17, 59–76. [Google Scholar] [CrossRef]
- Kizawa, J.; Machida, S.; Kobayashi, T.; Gotoh, Y.; Kurosaka, D. Changes of Oscillatory Potentials and Photopic Negative Response in Patients with Early Diabetic Retinopathy. Jpn. J. Ophtalmol. 2006, 50, 367–373. [Google Scholar] [CrossRef]
- Hancock, H.A.; Kraft, T.W. Oscillatory potential analysis and ERGs of normal and diabetic rats. Investig. Ophtalmol. Vis. Sci. 2004, 45, 1002–1008. [Google Scholar] [CrossRef]
- Li, Q.; Zemel, E.; Miller, B.; Perlman, I. Early retinal damage in experimental diabetes: Electroretinographical and morphological observations. Exp. Eye Res. 2002, 74, 615–625. [Google Scholar] [CrossRef]
- Sergeys, J.; Etienne, I.; Van Hove, I.; Lefevere, E.; Stalmans, I.; Feyen, J.H.M.; Moons, L.; Van Bergen, T. Longitudinal In Vivo Characterization of the Streptozotocin-Induced Diabetic Mouse Model: Focus on Early Inner Retinal Responses. Investig. Ophtalmol. Vis. Sci. 2019, 60, 807–822. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, H.W.; Verbraak, F.D.; Kok, P.H.B.; Stehouwer, M.; Garvin, M.K.; Sonka, M.; DeVries, J.H.; Schlingemann, R.O.; Abràmoff, M.D. Early neurodegeneration in the retina of type 2 diabetic patients. Investig. Ophtalmol. Vis. Sci. 2012, 53, 2715–2719. [Google Scholar] [CrossRef]
- van Dijk, H.W.; Kok, P.H.; Garvin, M.; Sonka, M.; Devries, J.H.; Michels, R.P.; van Velthoven, M.E.; Schlingemann, R.O.; Verbraak, F.D.; Abràmoff, M.D. Selective loss of inner retinal layer thickness in type 1 diabetic patients with minimal diabetic retinopathy. Investig. Ophtalmol. Vis. Sci. 2009, 50, 3404–3409. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Asahiyama, M.; Tanaka, T.; Yamamoto, S.; Murakami, K.; Kamiya, W.; Matsumura, Y.; Osawa, T.; Anai, M.; Fruchart, J.-C.; et al. Pemafibrate, a selective PPARα modulator, prevents non-alcoholic steatohepatitis development without reducing the hepatic triglyceride content. Sci. Rep. 2020, 10, 7818. [Google Scholar] [CrossRef] [PubMed]
- Peraza, M.A.; Burdick, A.D.; Marin, H.E.; Gonzalez, F.J.; Peters, J.M. The toxicology of ligands for peroxisome proliferator-activated receptors (PPAR). Toxicol. Sci. 2006, 90, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Wachtmeister, L. Oscillatory potentials in the retina: What do they reveal. Prog. Retin. Eye Res. 1998, 17, 485–521. [Google Scholar] [CrossRef]
- Heynen, H.; Wachtmeister, L.; van Norren, D. Origin of the oscillatory potentials in the primate retina. Vis. Res. 1985, 25, 1365–1373. [Google Scholar] [CrossRef]
- Yonemura, D.; Kawasaki, K. New approaches to ophthalmic electrodiagnosis by retinal oscillatory potential, drug-induced responses from retinal pigment epithelium and cone potential. Doc. Ophtalmol. 1979, 48, 163–222. [Google Scholar] [CrossRef]
- Araki, E.; Yamashita, S.; Arai, H.; Yokote, K.; Satoh, J.; Inoguchi, T.; Nakamura, J.; Maegawa, H.; Yoshioka, N.; Tanizawa, Y.; et al. Effects of Pemafibrate, a Novel Selective PPARα Modulator, on Lipid and Glucose Metabolism in Patients With Type 2 Diabetes and Hypertriglyceridemia: A Randomized, Double-Blind, Placebo-Controlled, Phase 3 Trial. Diabetes Care 2018, 41, 538. [Google Scholar] [CrossRef] [PubMed]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress-A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Nakagawa, Y.; Oishi, A.; Han, S.-I.; Wang, Y.; Kumagai, K.; Ohno, H.; Mizunoe, Y.; Iwasaki, H.; Sekiya, M.; et al. The Peroxisome Proliferator-Activated Receptor α (PPARα) Agonist Pemafibrate Protects against Diet-Induced Obesity in Mice. Int. J. Mol. Sci. 2018, 19, 2148. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Kessoku, T.; Ogawa, Y.; Tomeno, W.; Imajo, K.; Fujita, K.; Yoneda, M.; Takizawa, T.; Saito, S.; Nagashima, Y.; et al. Pemafibrate, a novel selective peroxisome proliferator-activated receptor alpha modulator, improves the pathogenesis in a rodent model of nonalcoholic steatohepatitis. Sci. Rep. 2017, 7, 42477. [Google Scholar] [CrossRef]
- Gross, B.; Pawlak, M.; Lefebvre, P.; Staels, B. PPARs in obesity-induced T2DM, dyslipidaemia and NAFLD. Nat. Rev. Endocrinol. 2017, 13, 36–49. [Google Scholar] [CrossRef]
- Kersten, S. Integrated physiology and systems biology of PPARα. Mol. Metab. 2014, 3, 354–371. [Google Scholar] [CrossRef]
- Fruchart, J.C. Peroxisome proliferator-activated receptor-alpha (PPARalpha): At the crossroads of obesity, diabetes and cardiovascular disease. Atherosclerosis 2009, 205, 1–8. [Google Scholar] [CrossRef]
- Stone, N.J.; Robinson, J.G.; Lichtenstein, A.H.; Bairey Merz, C.N.; Blum, C.B.; Eckel, R.H.; Goldberg, A.C.; Gordon, D.; Levy, D.; Lloyd-Jones, D.M.; et al. 2013 ACC/AHA Guideline on the Treatment of Blood Cholesterol to Reduce Atherosclerotic Cardiovascular Risk in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Am. Coll. Cardiol. 2014, 63, 2889–2934. [Google Scholar] [CrossRef]
- Bell, D.S.; Al Badarin, F.; O’Keefe, J.H., Jr. Therapies for diabetic dyslipidaemia. Diabetes Obes. Metab. 2011, 13, 313–325. [Google Scholar] [CrossRef]
- Araki, E.; Yamashita, S.; Arai, H.; Yokote, K.; Satoh, J.; Inoguchi, T.; Nakamura, J.; Maegawa, H.; Yoshioka, N.; Tanizawa, Y.; et al. Efficacy and safety of pemafibrate in people with type 2 diabetes and elevated triglyceride levels: 52-week data from the PROVIDE study. Diabetes Obes. Metab. 2019, 21, 1737–1744. [Google Scholar] [CrossRef]
- Wong-Riley, M.T.T. Energy metabolism of the visual system. Eye Brain 2010, 2, 99–116. [Google Scholar] [CrossRef]
- Cecilia, O.-M.; José Alberto, C.-G.; José, N.-P.; Ernesto Germán, C.-M.; Ana Karen, L.-C.; Luis Miguel, R.-P.; Ricardo Raúl, R.-R.; Adolfo Daniel, R.-C. Oxidative Stress as the Main Target in Diabetic Retinopathy Pathophysiology. J. Diabetes Res. 2019, 2019, 8562408. [Google Scholar] [CrossRef] [PubMed]
- Zhan, S.S.; Beyreuther, K.; Schmitt, H.P. Quantitative assessment of the synaptophysin immuno-reactivity of the cortical neuropil in various neurodegenerative disorders with dementia. Dementia 1993, 4, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Calderone, J.B.; Lewis, G.P.; Linberg, K.A.; Fisher, S.K.; Jacobs, G.H. Cone Photoreceptor Recovery after Experimental Detachment and Reattachment: An Immunocytochemical, Morphological, and Electrophysiological Study. Investig. Opthalmol. Vis. Sci. 2003, 44, 416–425. [Google Scholar] [CrossRef]
- Kurihara, T.; Ozawa, Y.; Shinoda, K.; Nagai, N.; Inoue, M.; Oike, Y.; Tsubota, K.; Ishida, S.; Okano, H. Neuroprotective effects of angiotensin II type 1 receptor (AT1R) blocker, telmisartan, via modulating AT1R and AT2R signaling in retinal inflammation. Investig. Opthalmol. Vis. Sci. 2006, 47, 5545–5552. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, T.; Ozawa, Y.; Nagai, N.; Shinoda, K.; Noda, K.; Imamura, Y.; Tsubota, K.; Okano, H.; Oike, Y.; Ishida, S. Angiotensin II type 1 receptor signaling contributes to synaptophysin degradation and neuronal dysfunction in the diabetic retina. Diabetes 2008, 57, 2191–2198. [Google Scholar] [CrossRef]
- Spiwoks-Becker, I.; Vollrath, L.; Seeliger, M.W.; Jaissle, G.; Eshkind, L.G.; Leube, R.E. Synaptic vesicle alterations in rod photoreceptors of synaptophysin-deficient mice. Neuroscience 2001, 107, 127–142. [Google Scholar] [CrossRef]
- Yang, X.; Hui, Q.; Yu, B.; Huang, Z.; Zhou, P.; Wang, P.; Wang, Z.; Pang, S.; Li, J.; Wang, H.; et al. Design and Evaluation of Lyophilized Fibroblast Growth Factor 21 and Its Protection against Ischemia Cerebral Injury. Bioconjugate Chem. 2018, 29, 287–295. [Google Scholar] [CrossRef]
- Taliyan, R.; Chandran, S.K.; Kakoty, V. Therapeutic Approaches to Alzheimer’s Type of Dementia: A Focus on FGF21 Mediated Neuroprotection. Curr. Pharm. Des. 2019, 25, 2555–2568. [Google Scholar] [CrossRef] [PubMed]
- Fon Tacer, K.; Bookout, A.L.; Ding, X.; Kurosu, H.; John, G.B.; Wang, L.; Goetz, R.; Mohammadi, M.; Kuro-o, M.; Mangelsdorf, D.J.; et al. Research Resource: Comprehensive Expression Atlas of the Fibroblast Growth Factor System in Adult Mouse. Mol. Endocrinol. 2010, 24, 2050–2064. [Google Scholar] [CrossRef]
- Sa-nguanmoo, P.; Chattipakorn, N.; Chattipakorn, S.C. Potential roles of fibroblast growth factor 21 in the brain. Metab. Brain Dis. 2016, 31, 239–248. [Google Scholar] [CrossRef]
- Hsuchou, H.; Pan, W.; Kastin, A.J. The fasting polypeptide FGF21 can enter brain from blood. Peptides 2007, 28, 2382–2386. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, J.; Liu, H.; Xiong, Y.; Zou, Y.; Huang, W.; Shao, M.; Wu, J.; Yu, L.; Wang, X.; et al. FGF21 Protects the Blood-Brain Barrier by Upregulating PPARγ via FGFR1/β-klotho after Traumatic Brain Injury. J. Neuotrauma. 2018, 35, 2091–2103. [Google Scholar] [CrossRef] [PubMed]
- Owen, B.M.; Bookout, A.L.; Ding, X.; Lin, V.Y.; Atkin, S.D.; Gautron, L.; Kliewer, S.A.; Mangelsdorf, D.J. FGF21 contributes to neuroendocrine control of female reproduction. Nat. Med. 2013, 19, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Vaz, J. The Blood-Retinal Barrier in the Management of Retinal Disease: EURETINA Award Lecture. Ophthalmol. Int. J. Ophthalmol. Z. Augenheilkd. 2017, 237, 1–10. [Google Scholar] [CrossRef]
- Klaassen, I.; Van Noorden, C.J.; Schlingemann, R.O. Molecular basis of the inner blood-retinal barrier and its breakdown in diabetic macular edema and other pathological conditions. Prog. Retin. Eye Res. 2013, 34, 19–48. [Google Scholar] [CrossRef]
- Shiono, A.; Sasaki, H.; Sekine, R.; Abe, Y.; Matsumura, Y.; Inagaki, T.; Tanaka, T.; Kodama, T.; Aburatani, H.; Sakai, J.; et al. PPARα activation directly upregulates thrombomodulin in the diabetic retina. Sci. Rep. 2020, 10, 10837. [Google Scholar] [CrossRef]
- Ciudin, A.; Hernández, C.; Simó, R. Molecular Implications of the PPARs in the Diabetic Eye. PPAR Res. 2013, 2013, 686525. [Google Scholar] [CrossRef]
- Davidson, M.H.; Armani, A.; McKenney, J.M.; Jacobson, T.A. Safety Considerations with Fibrate Therapy. Am. J. Cardiol. 2007, 99, S3–S18. [Google Scholar] [CrossRef]
- Emami, F.; Hariri, A.; Matinfar, M.; Nematbakhsh, M. Fenofibrate-induced renal dysfunction, yes or no? J. Res. Med. Sci. 2020, 25, 39. [Google Scholar] [CrossRef]
- Fruchart, J.-C. Pemafibrate (K-877), a novel selective peroxisome proliferator-activated receptor alpha modulator for management of atherogenic dyslipidaemia. Cardiovasc. Diabetol. 2017, 16, 124. [Google Scholar] [CrossRef] [PubMed]
- Fruchart, J.C. Selective peroxisome proliferator-activated receptor α modulators (SPPARMα): The next generation of peroxisome proliferator-activated receptor α-agonists. Cardiovasc. Diabetol. 2013, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Abe, K.; Toma, T.; Nishikawa, M.; Ozawa, H.; Okuda, A.; Araki, T.; Oda, S.; Inoue, K.; Shibuya, K.; et al. Design and synthesis of highly potent and selective human peroxisome proliferator-activated receptor alpha agonists. Bioorg. Med. Chem. Lett. 2007, 17, 4689–4693. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, S.; Arai, H.; Yokote, K.; Araki, E.; Suganami, H.; Yamashita, S. Efficacy and safety of pemafibrate (K-877), a selective peroxisome proliferator-activated receptor α modulator, in patients with dyslipidemia: Results from a 24-week, randomized, double blind, active-controlled, phase 3 trial. J. Clin. Lipidol. 2018, 12, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Miyake, S.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Phase II enzyme induction by a carotenoid, lutein, in a PC12D neuronal cell line. Biochem. Biophys. Res. Commun. 2014, 446, 535–540. [Google Scholar] [CrossRef]
- Miwa, Y.; Hoshino, Y.; Shoda, C.; Jiang, X.; Tsubota, K.; Kurihara, T. Pharmacological HIF inhibition prevents retinal neovascularization with improved visual function in a murine oxygen-induced retinopathy model. Neurochem. Int. 2019, 128, 21–31. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Direction | Sequence (5’ → 3’) | Accession Number |
|---|---|---|---|
| β-actin | Forward | GGGAAATCGTGCGTGACA | NM_007393.5 |
| Reverse | CAAGAAGGAAGGCTGGAAAA | ||
| Abca1 | Forward | CGTTTCCGGGAAGTGTCCTA | NM_013454.3 |
| Reverse | GCTAGAGATGACAAGGAGGATGGA | ||
| Fabp4 | Forward | CCGCAGACGACAGGA | NM_024406.3 |
| Reverse | CTCATGCCCTTTCATAAACT | ||
| Fgf21 | Forward | AACAGCCATTCACTTTGCCTGAGC | NM_020013.4 |
| Reverse | GGCAGCTGGAATTGTGTTCTGACT | ||
| Vldlr | Forward | GAGCCCCTGAAGGAATGCC | NM_001161420.1 |
| Reverse | CCTATAACTAGGTCTTTGCAGATATGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomita, Y.; Lee, D.; Miwa, Y.; Jiang, X.; Ohta, M.; Tsubota, K.; Kurihara, T. Pemafibrate Protects Against Retinal Dysfunction in a Murine Model of Diabetic Retinopathy. Int. J. Mol. Sci. 2020, 21, 6243. https://doi.org/10.3390/ijms21176243
Tomita Y, Lee D, Miwa Y, Jiang X, Ohta M, Tsubota K, Kurihara T. Pemafibrate Protects Against Retinal Dysfunction in a Murine Model of Diabetic Retinopathy. International Journal of Molecular Sciences. 2020; 21(17):6243. https://doi.org/10.3390/ijms21176243
Chicago/Turabian StyleTomita, Yohei, Deokho Lee, Yukihiro Miwa, Xiaoyan Jiang, Masayuki Ohta, Kazuo Tsubota, and Toshihide Kurihara. 2020. "Pemafibrate Protects Against Retinal Dysfunction in a Murine Model of Diabetic Retinopathy" International Journal of Molecular Sciences 21, no. 17: 6243. https://doi.org/10.3390/ijms21176243
APA StyleTomita, Y., Lee, D., Miwa, Y., Jiang, X., Ohta, M., Tsubota, K., & Kurihara, T. (2020). Pemafibrate Protects Against Retinal Dysfunction in a Murine Model of Diabetic Retinopathy. International Journal of Molecular Sciences, 21(17), 6243. https://doi.org/10.3390/ijms21176243