Artificial Light at Night Increases Recruitment of New Neurons and Differentially Affects Various Brain Regions in Female Zebra Finches

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. ALAN Increases New Neuronal Recruitment in the MSt, HC, and NC
2.2. ALAN Increases New Neuronal Recruitment in Sub-Regions of MSt at Different Rates
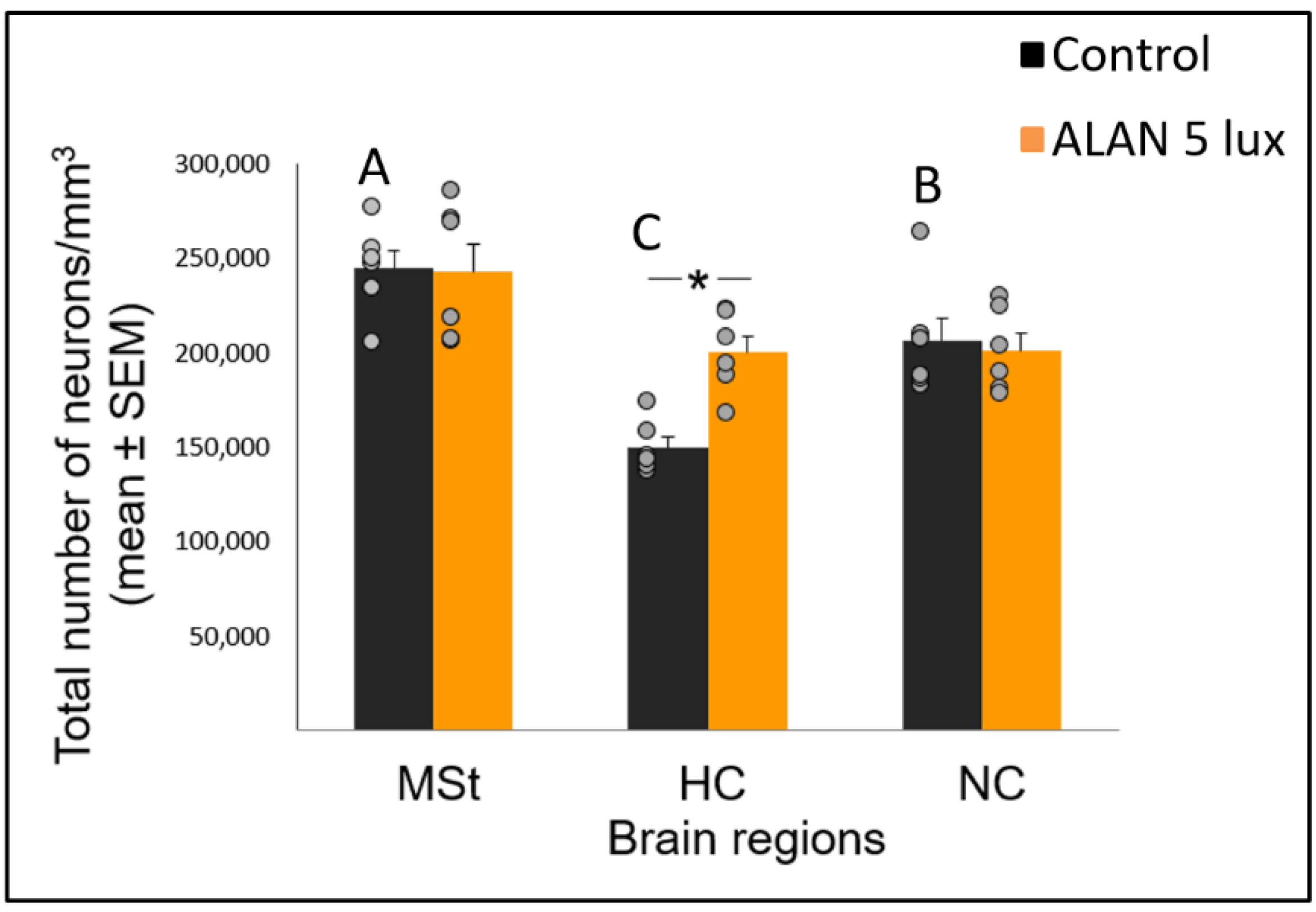
2.3. ALAN Increases Total Neuronal Densities in the HC
2.4. ALAN Reduces Nocturnal Melatonin Levels
2.5. ALAN Does Not Affect Body Mass
3. Discussion
3.1. ALAN Increases New Neuronal Recruitment in the MSt, HC, and NC
3.2. A Possible Temporal Differential Effect of ALAN on Total Neuronal Densities in Various Brain Regions
3.3. Possible Relation between Melatonin and Neuronal Recruitment in Birds
3.4. Conclusions and Future Directions
4. Materials and Methods
4.1. Experimental Design
4.2. Melatonin Levels in Plasma
4.3. BrdU Administration, Histology, and Immunohistochemistry
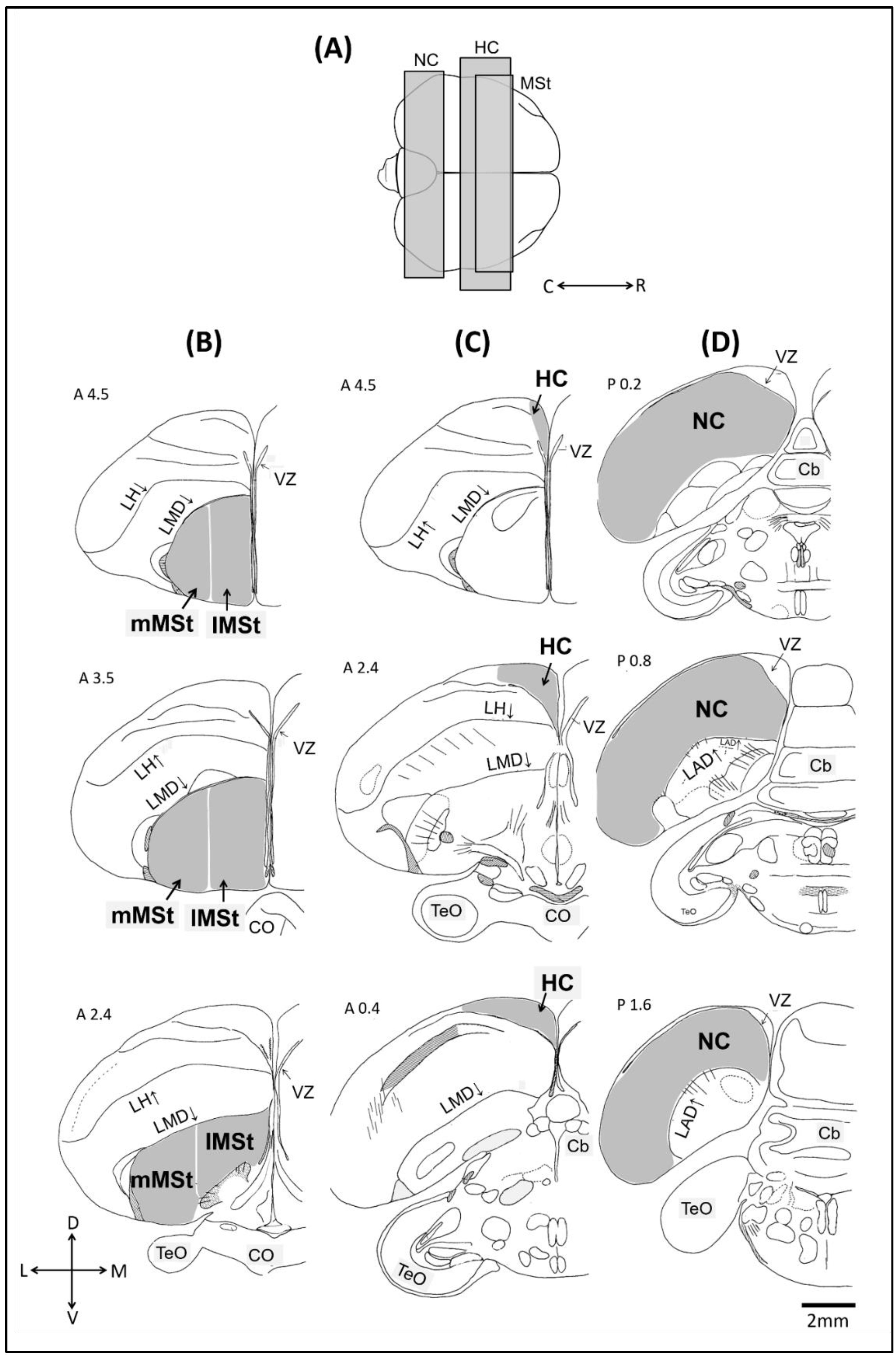
4.4. Mapping and Quantification
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALAN | Artificial light at night |
| HC | Hippocampus |
| MSt | Medial striatum |
| NC | Nidopallium caudale |
| NCL | Lateral caudal nidopallium |
| VZ | Ventricular zone |
| MEL | Melatonin |
| SE | Standard Error |
| lMSt | Lateral medial striatum |
| mMSt | Medial medial striatum |
| BrdU | Bromodeoxyuridine/5-bromo-2′-deoxyuridine |
| LD | Light-dark |
| IgG | Immunoglobulin G |
| CFL | Compact fluorescent lamp |
| DAPI | 4′,6-diamidino-2-phenylindole |
References
- Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light pollution, circadian photoreception, and melatonin in vertebrates. Sustainability 2019, 11, 6400. [Google Scholar] [CrossRef]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef]
- Taufique, S.T.; Prabhat, A.; Kumar, V. Illuminated night alters hippocampal gene expressions and induces depressive-like responses in diurnal corvids. Eur. J. Neurosci. 2018, 48, 3005–3018. [Google Scholar] [CrossRef]
- Taufique, S.T.; Prabhat, A.; Kumar, V. Light at night affects hippocampal and nidopallial cytoarchitecture: Implication for impairment of brain function in diurnal corvids. J. Exp. Zool. Part A 2019, 331, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, T.H.; Goldsmith, T.H. Optimization, constraint and history in the evolution of eyes. Q. Rev. Biol. 1990, 65, 281–322. [Google Scholar] [CrossRef]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The biological impacts of artificial light at night: The research challenge. Philos. Trans. R. Soc. B 2015, 370, 20140133. [Google Scholar] [CrossRef]
- Dawson, A.; King, V.M.; Bentley, G.E.; Ball, G.F. Photoperiodic control of seasonality in birds. J. Biol. Rhythm. 2001, 16, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Moaraf, S.; Vistoropsky, Y.; Pozner, T.; Heiblum, R.; Okuliarová, M.; Zeman, M.; Barnea, A. Artificial light at night affects brain plasticity and melatonin in birds. Neurosci. Lett. 2020, 716, 134639. [Google Scholar] [CrossRef] [PubMed]
- Kirn, J.R.; Schwabl, H. Photoperiod regulation of neuron death in the adult canary. J. Neurobiol. 1997, 33, 223–231. [Google Scholar] [CrossRef]
- Scharff, C.; Kirn, J.R.; Grossman, M.; Macklis, J.D.; Nottebohm, F. Targeted neuronal death affects neuronal replacement and vocal behavior in adult songbirds. Neuron 2000, 25, 481–492. [Google Scholar] [CrossRef]
- Paton, J.A.; Nottebohm, F. Neurons generated in the adult brain are recruited into functional circuits. Science 1984, 225, 1046–1048. [Google Scholar] [CrossRef] [PubMed]
- Vates, G.E.; Broome, B.M.; Mello, C.V.; Nottebohm, F. Auditory pathways of caudal telencephalon and their relation to the song system of adult male zebra finches (Taenopygia guttata). J. Comp. Neurol. 1996, 366, 613–642. [Google Scholar] [CrossRef]
- Mello, C.V.; Clayton, D.F. Song-induced ZENK gene expression in auditory pathways of songbird brain and its relation to the song control system. J. Neurosc. 1994, 14, 6652–6666. [Google Scholar] [CrossRef]
- Mello, C.V.; Vates, E.; Okuhata, S.; Nottebohm, F. Descending auditory pathways in the adult male zebra finch (Taeniopygia guttata). J. Comp. Neurol. 1998, 395, 137–160. [Google Scholar] [CrossRef]
- Kuenzel, W.J.; Medina, L.; Csillag, A.; Perkel, D.J.; Reiner, A. The avian subpallium: New insights into structural and functional subdivisions occupying the lateral subpallial wall and their embryological origins. Brain Res. 2011, 1424, 67–101. [Google Scholar] [CrossRef]
- Watanabe, S. Effects of lobus parolfactorius lesions on repeated acquisition of spatial discrimination in pigeons. Brain Behav. Evol. 2001, 58, 333–342. [Google Scholar] [CrossRef]
- Matsushima, T.; Izawa, E.I.; Aoki, N.; Yanagihara, S. The mind through chick eyes: Memory, cognition and anticipation. Zool. Sci. 2003, 20, 395–408. [Google Scholar] [CrossRef]
- Reiner, A.; Perkel, D.J.; Bruce, L.L.; Butler, A.; Csillag, A.; Kuenzel, W.; Medina, L.; Paxinos, G.; Shimizu, T.; Striedter, G.; et al. The avian brain nomenclature forum: Terminology for a new century in comparative neuroanatomy. J. Comp. Neurol. 2004, 473, E1. [Google Scholar] [CrossRef]
- Sherry, D.F.; Vaccarino, A.L.; Buckenham, K.; Herz, R.S. The hippocampal complex of food-storing birds. Brain Behav. Evol. 1989, 34, 308–317. [Google Scholar] [CrossRef]
- Shettleworth, S.J. Spatial memory in food-storing birds. Philos. Trans. R. Soc. B 1990, 329, 143–151. [Google Scholar] [CrossRef]
- Smulders, T.V. The avian hippocampal formation and the stress response. Brain Behav. Evol. 2017, 90, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.; Samplonius, J.M.; Schlicht, E.; Valcu, M.; Kempenaers, B. Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behav. Ecol. 2014, 25, 1037–1047. [Google Scholar] [CrossRef]
- De Jong, M.; Jening, L.; Ouyang, J.Q.; van Oers, K.; Spoelstra, K.; Visser, M.E. Dose-dependent responses of avian daily rhythms to artificial light at night. Physiol. Behav. 2016, 155, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kirn, J.; Loughlin, B.O.; Kasparian, S.; Nottebohm, F. Cell death and neuronal recruitment in the high vocal center of adult male canaries are temporally related to changes in song. Proc. Natl. Acad. Sci. USA 1994, 91, 7844–7848. [Google Scholar] [CrossRef]
- Jarvis, E.D.; Güntürkün, O.; Bruce, L.; Csillag, A.; Karten, H.; Kuenzel, W.; Medina, L.; Paxinos, G.; Perkel, D.J.; Shimizu, T.; et al. Avian brains and a new understanding of vertebrate brain evolution. Nat. Rev. Neurosci. 2005, 6, 151–159. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Bowman, R.; Donnelly, R. (Eds.) Avian Ecology and Conservation in an Urbanizing World; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Medina, L.; Reiner, A. Distribution of choline acetyltransferase immunoreactivity in the pigeon brain. J. Comp. Neurol. 1994, 342, 497–537. [Google Scholar] [CrossRef]
- Reiner, A.; Karle, E.J.; Anderson, K.D.; Medina, L. Catecholaminergic perikarya and fivers in the avian nervous system. In Phylogeny and Development of Catecholaminergic Systems in the CNS of Vertebrates; Smeets, W.J., Reiner, A., Eds.; Cambridge University: Cambridge, UK, 1994; pp. 135–181. [Google Scholar]
- Metzger, M.; Toledo, C.; Braun, K. Serotonergic innervation of the telencephalon in the domestic chick. Brain Res. Bull. 1996, 52, 163–174. [Google Scholar] [CrossRef]
- Reiner, M.A.; Medina, L.; Veenman, C.L. Structural and functional evolution of the basal ganglia in vertebrates. Brain Res. Rev. 1998, 28, 235–285. [Google Scholar] [CrossRef]
- Veenman, C.L.; Wild, J.M.; Reiner, A. Organization of the avian “corticostriatal” projection system: A retrograde and anterograde pathway tracing study in pigeons. J. Comp. Neurol. 1995, 354, 87–126. [Google Scholar] [CrossRef]
- Wild, J.M. Descending projections of the songbird nucleus robustus archistriatalis. J. Comp. Neurol. 1993, 338, 225–241. [Google Scholar] [CrossRef]
- Barnea, A.; Mishal, A.; Nottebohm, F. Social and spatial changes induce multiple survival regimes for new neurons in two regions of the adult brain: An anatomical representation of time? Behav. Brain Res. 2006, 167, 63–74. [Google Scholar] [CrossRef]
- Adar, E.; Nottebohm, F.; Barnea, A. The relationship between nature of social change, age, and position of new neurons and their survival in adult zebra finch brain. J. Neurosci. Res. 2008, 28, 5394–5400. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barnea, A.; Pravosudov, V. Birds as a model to study adult neurogenesis: Bridging evolutionary, comparative and neuroethological approaches. Eur. J. Neurosci. 2011, 34, 884–907. [Google Scholar] [CrossRef] [PubMed]
- Heldmaier, G.; Werner, D. Environmental signal processing and adaptation. In Environmental Signal Processing and Adaptation; Springer: Berlin/Heidelberg, Germany, 2003; pp. 1–8. [Google Scholar]
- Maldonado, K.E.; Cavieres, G.; Veloso, C.; Canals, M.; Sabat, P. Physiological responses in rufous-collared sparrows to thermal acclimation and seasonal acclimatization. J. Comp. Physiol. B 2009, 179, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Balthazart, J.; Boseret, G.; Konkle, A.T.; Hurley, L.L.; Ball, G.F. Doublecortin as a marker of adult neuroplasticity in the canary song control nucleus HVC. Eur. J. Neurosci. 2008, 27, 801–817. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, A.; Fujioka, T.; Tsuruta, R.; Izumi, T.; Kasaoka, S.; Maekawa, T. Effects of a constant light environment on hippocampal neurogenesis and memory in mice. Neurosci. Lett. 2011, 488, 41–44. [Google Scholar] [CrossRef]
- Li, D.; Ma, S.; Guo, D.; Cheng, T.; Li, H.; Tian, Y.; Li, J.; Guan, F.; Yang, B.; Wang, J. Environmental circadian disruption worsens neurologic impairment and inhibits hippocampal neurogenesis in adult rats after traumatic brain injury. Cell. Mol. Neurobiol. 2016, 36, 1045–1055. [Google Scholar] [CrossRef]
- Snyder, J.S.; Soumier, A.; Brewer, M.; Pickel, J.; Cameron, H.A. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 2011, 476, 458–461. [Google Scholar] [CrossRef]
- Gualtieri, F.; Armstrong, E.A.; Longmoor, G.K.; D’Eath, R.B.; Sandilands, V.; Boswell, T.; Smulders, T.V. Unpredictable chronic mild stress suppresses the incorporation of new neurons at the caudal pole of the chicken Hippocampal Formation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Brooks, D.S.; Cassone, V.M. Daily and circadian regulation of 2-[125I] iodomelatonin binding in the chick brain. Endocrinology 1992, 131, 1297–1304. [Google Scholar] [CrossRef]
- Hendel, R.C.; Turek, F.W. Suppression of locomotor activity in sparrows by treatment with melatonin. Physiol. Behav. 1987, 21, 275–278. [Google Scholar] [CrossRef]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Raap, T.; Pinxten, R.; Eens, M. Light pollution disrupts sleep in free-living animals. Sci. Rep. 2015, 5, 13557. [Google Scholar] [CrossRef]
- Mishra, I.; Knerr, R.M.; Stewart, A.A.; Payette, W.I.; Richter, M.M.; Ashley, N.T. Light at night disrupts diel patterns of cytokine gene expression and endocrine profiles in zebra finch (Taeniopygia guttata). Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Robert, K.A.; Lesku, J.A.; Partecke, J.; Chambers, B. Artificial light at night desynchronizes strictly seasonal reproduction in a wild mammal. Proc. R. Soc. B 2015, 282, 20151745. [Google Scholar] [CrossRef] [PubMed]
- Brüning, A.; Hölker, F.; Franke, S.; Preuer, T.; Kloas, W. Spotlight on fish: Light pollution affects circadian rhythms of European perch but does not cause stress. Sci. Total Environ. 2015, 511, 516–522. [Google Scholar] [CrossRef]
- Wren, M.A.; Dauchy, R.T.; Hanifin, J.P.; Jablonski, M.R.; Warfield, B.; Brainard, G.C.; Blask, D.E.; Hill, S.M.; Ooms, T.G.; Bohm, R.P., Jr. Effect of different spectral transmittances through tinted animal cages on circadian metabolism and physiology in Sprague-Dawley rats. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 44–51. [Google Scholar]
- Kilic, E.; Kilic, Ü.; Bacigaluppi, M.; Guo, Z.; Abdallah, N.B.; Wolfer, D.P.; Reiter, R.J.; Hermann, D.M.; Bassetti, C.L. Delayed melatonin administration promotes neuronal survival, neurogenesis and motor recovery, and attenuates hyperactivity and anxiety after mild focal cerebral ischemia in mice. J. Pineal. Res. 2008, 45, 142–148. [Google Scholar] [CrossRef]
- Pozner, T.; Vistoropsky, Y.; Moaraf, S.; Heiblum, R.; Barnea, A. Questioning Seasonality of Neuronal Plasticity in the Adult Avian Brain. Sci. Rep. 2018, 8, 11289. [Google Scholar] [CrossRef]
- Zeman, M.; Gwinner, E. Ontogeny of the rhythmic melatonin production in a precocial and an altricial bird, the Japanese quail and the European starling. J. Comp. Physiol. A 1993, 172, 333–338. [Google Scholar] [CrossRef]
- Van’t Hof, T.J.; Gwinner, E. Development of post-hatching melatonin rhythm in zebra finches (Poephila guttata). Experientia 1996, 52, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Vistoropsky, Y.; Heiblum, R.; Smorodinsky, N.I.; Barnea, A. Active immunization against vasoactive intestinal polypeptide decreases neuronal recruitment and inhibits reproduction in zebra finches. J. Comp. Neurol. 2016, 524, 2516–2528. [Google Scholar] [CrossRef] [PubMed]
- Cattan, A.; Ayali, A.; Barnea, A. The cell birth marker BrdU does not affect recruitment of subsequent cell divisions in the adult avian brain. BioMed Res. Int. 2015, 2015, 126078. [Google Scholar] [CrossRef] [PubMed]
- Guillery, R.W.; Herrup, K. Quantification without pontification: Choosing a method for counting objects in sectioned tissues. J. Comp. Neurol. 1997, 386, 2–7. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moaraf, S.; Heiblum, R.; Vistoropsky, Y.; Okuliarová, M.; Zeman, M.; Barnea, A. Artificial Light at Night Increases Recruitment of New Neurons and Differentially Affects Various Brain Regions in Female Zebra Finches. Int. J. Mol. Sci. 2020, 21, 6140. https://doi.org/10.3390/ijms21176140
Moaraf S, Heiblum R, Vistoropsky Y, Okuliarová M, Zeman M, Barnea A. Artificial Light at Night Increases Recruitment of New Neurons and Differentially Affects Various Brain Regions in Female Zebra Finches. International Journal of Molecular Sciences. 2020; 21(17):6140. https://doi.org/10.3390/ijms21176140
Chicago/Turabian StyleMoaraf, Stan, Rachel Heiblum, Yulia Vistoropsky, Monika Okuliarová, Michal Zeman, and Anat Barnea. 2020. "Artificial Light at Night Increases Recruitment of New Neurons and Differentially Affects Various Brain Regions in Female Zebra Finches" International Journal of Molecular Sciences 21, no. 17: 6140. https://doi.org/10.3390/ijms21176140
APA StyleMoaraf, S., Heiblum, R., Vistoropsky, Y., Okuliarová, M., Zeman, M., & Barnea, A. (2020). Artificial Light at Night Increases Recruitment of New Neurons and Differentially Affects Various Brain Regions in Female Zebra Finches. International Journal of Molecular Sciences, 21(17), 6140. https://doi.org/10.3390/ijms21176140