The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Validation and Characterization of PADI2 Knockout U937 Cells
2.2. Effects of PADI2 Knockout on Proinflammatory Cytokines Expression and Secretion
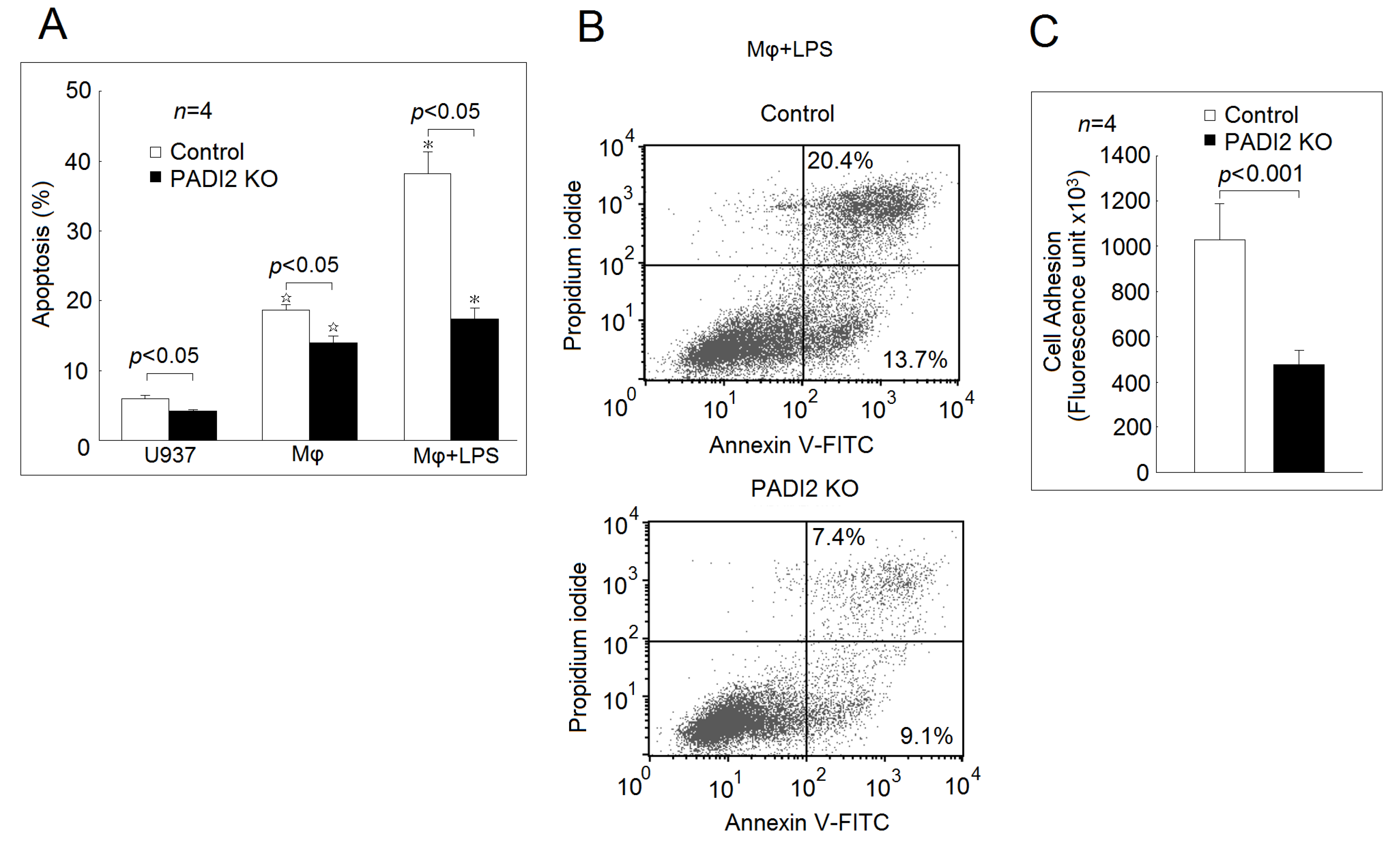
2.3. Effects of PADI2 Knockout on Cell Apoptosis and Adhesion
2.4. Effecst of PADI2 Knockout on the Protein Expression of Caspases
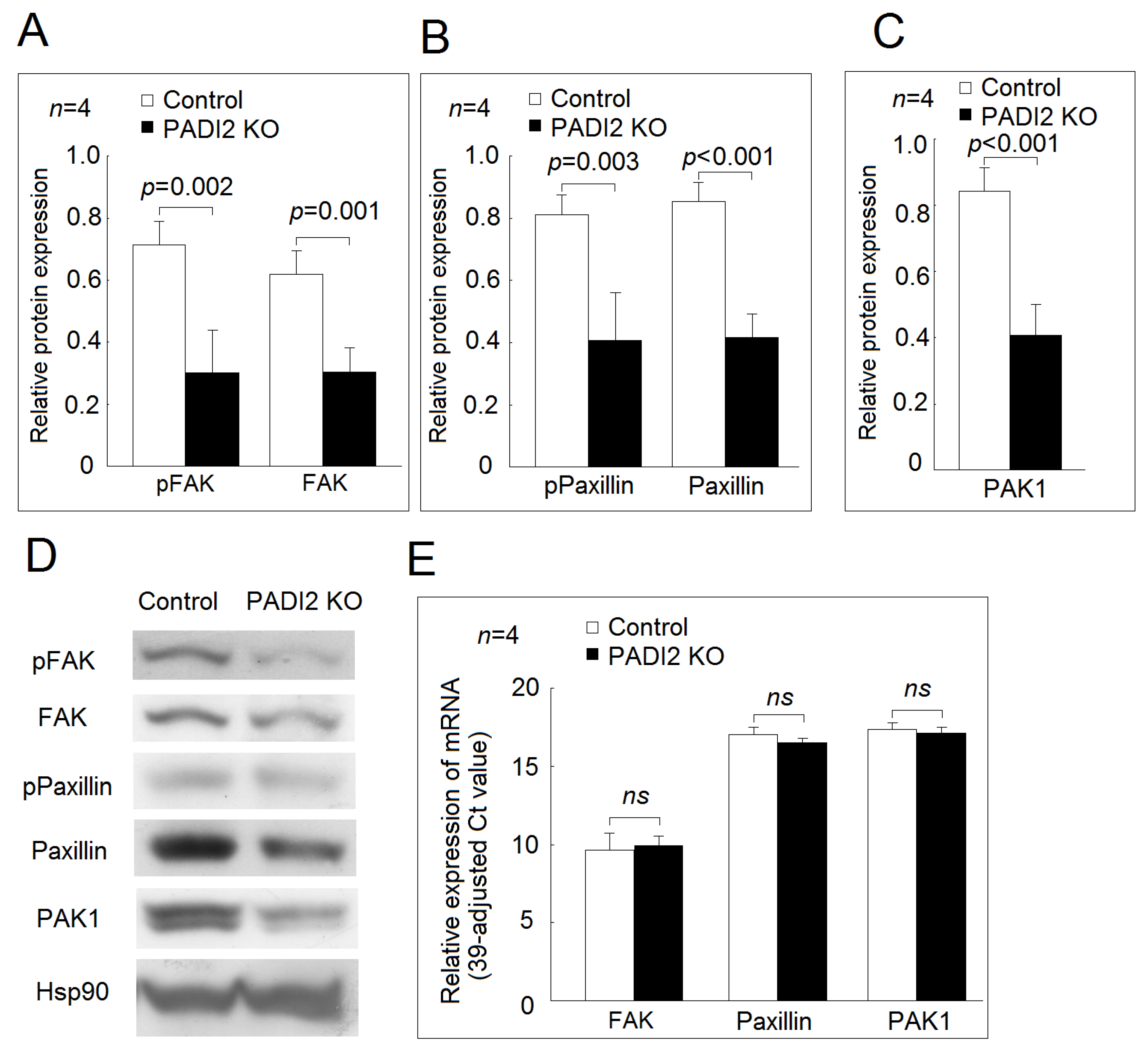
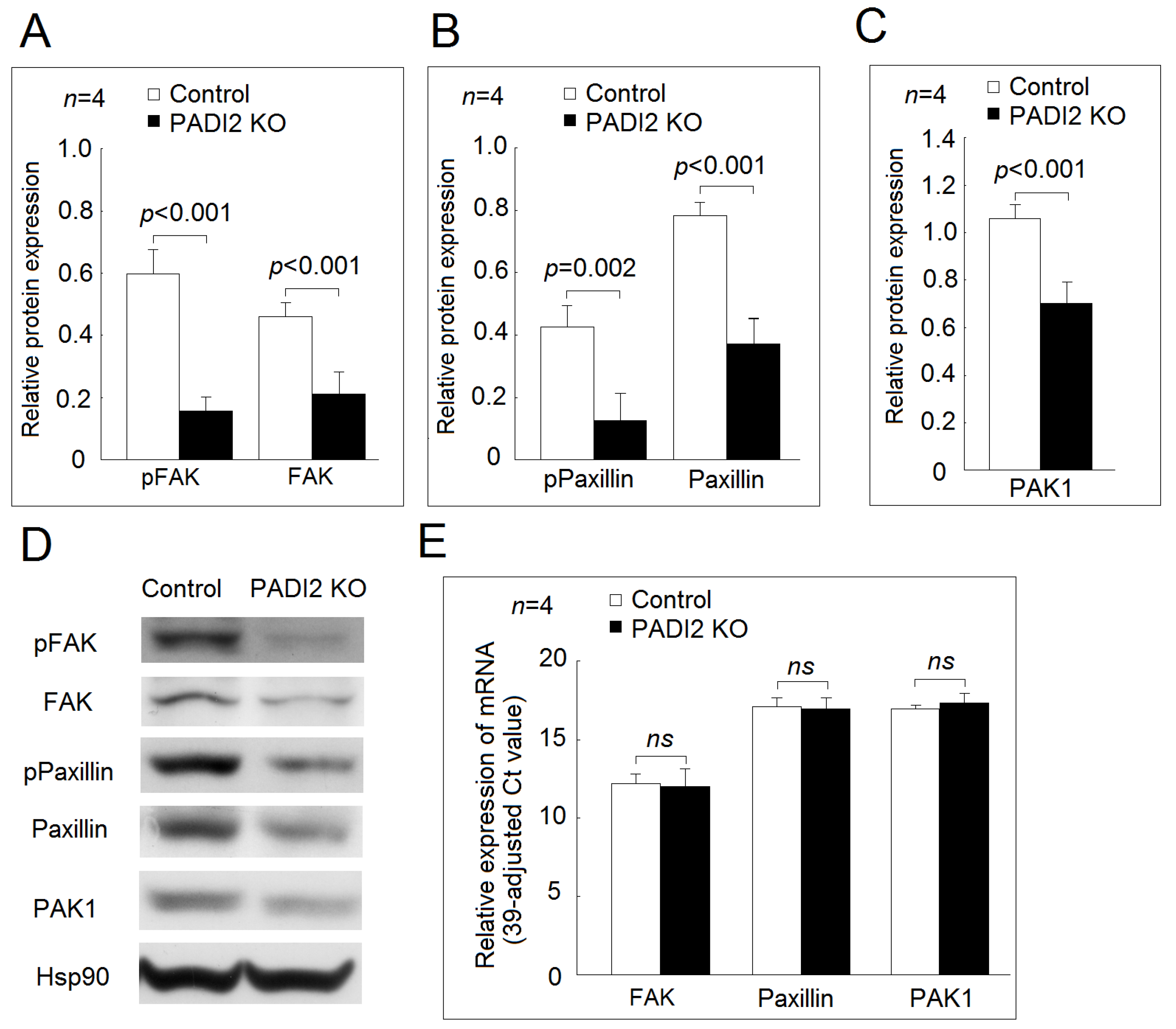
2.5. Effecst of PADI2 Knockout on the Protein and mRNA Expression of Adhesion-Related Genes
3. Discussion
4. Material and Methods
4.1. Purification of Anticitrullinated Protein Antibodies (ACPA) from Pooled ACPA(+) Sera in Patients with RA
4.2. Preparation of PADI2 Knockout U937 Cells
4.3. ELISA
4.4. Flow Cytometry Analysis
4.5. Cell Adhesion Assay
4.6. Preparation of the Cell Nuclear Protein Extract
4.7. Immunoprecipitation of Citrullinated Protein
4.8. Western Blotting Analysis
4.9. Measurement of Cytokine Expression Levels by RT-PCR
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PADI | peptidylarginine deiminases |
| IL | interleukin |
| TNF-α | tumor necrosis factor-alpha |
| ACPAs | anticitrullinated protein antibodies |
| PMA | phorbol 12-myristate 13-acetate |
| LPS | lipopolysaccharides |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| Cas9 | CRISPR-associated protein-9 nuclease |
| RT-PCR | real-time reverse transcription-polymerase chain reaction |
| ELISA | enzyme-linked immunosorbent assay |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| PAK1 | p21-activated kinase 1 |
| FAK | focal adhesion kinase |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| HRP | horseradish peroxidase |
References
- Wang, S.; Wang, Y. Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim. Biophys. Acta BBA Bioenergy 2013, 1829, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.; Al Ghamdi, K.A.; Khan, R.H.; Uversky, V.; Redwan, E.M. An interplay of structure and intrinsic disorder in the functionality of peptidylarginine deiminases, A family of key autoimmunity-related enzymes. Cell. Mol. Life Sci. 2019, 76, 4635–4662. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-C.; Yu, H.-C. The roles of anti-citrullinated protein antibodies in the immunopathogenesis of rheumatoid arthritis. Tzu Chi Med. J. 2019, 31, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Curran, A.M.; Naik, P.; Giles, J.T.; Darrah, E. PAD enzymes in rheumatoid arthritis: Pathogenic effectors and autoimmune targets. Nat. Rev. Rheumatol. 2020, 16, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Beato, M.; Sharma, P. Peptidyl Arginine Deiminase 2 (PADI2)-Mediated Arginine Citrullination Modulates Transcription in Cancer. Int. J. Mol. Sci. 2020, 21, 1351. [Google Scholar] [CrossRef]
- Yuzhalin, A.E.; Gordon-Weeks, A.N.; Tognoli, M.L.; Jones, K.; Markelc, B.; Konietzny, R.; Fischer, R.; Muth, A.; O’Neill, E.; Thompson, P.R.; et al. Colorectal cancer liver metastatic growth depends on PAD4-driven citrullination of the extracellular matrix. Nat. Commun. 2018, 9, 4783. [Google Scholar] [CrossRef]
- Lai, N.-S.; Yu, H.-C.; Tung, C.-H.; Huang, K.-Y.; Huang, H.-B.; Lu, M.-C. Increased peptidylarginine deiminases expression during the macrophage differentiation and participated inflammatory responses. Arthritis Res. 2019, 21, 108. [Google Scholar] [CrossRef]
- Bawadekar, M.; Shim, D.; Johnson, C.J.; Warner, T.F.; Rebernick, R.; Damgaard, D.; Nielsen, C.H.; Pruijn, G.J.M.; Nett, J.E.; Shelef, M.A. Peptidylarginine deiminase 2 is required for tumor necrosis factor alpha-induced citrullination and arthritis, but not neutrophil extracellular trap formation. J. Autoimmun. 2017, 80, 39–47. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.; Lin, C.-Y.; Gootenberg, J.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Zhang, F. High-Throughput functional genomics using CRISPR–Cas9. Nat. Rev. Genet. 2015, 16, 299–311. [Google Scholar] [CrossRef]
- Sun, B.; Dwivedi, N.; Bechtel, T.J.; Paulsen, J.L.; Muth, A.; Bawadekar, M.; Li, G.; Thompson, P.R.; Shelef, M.A.; Schiffer, C.A.; et al. Citrullination of NF-κB p65 promotes its nuclear localization and TLR-induced expression of IL-1β and TNFα. Sci. Immunol. 2017, 2, eaal3062. [Google Scholar] [CrossRef] [PubMed]
- Stefanelli, V.L.; Choudhury, S.; Hu, P.; Liu, Y.; Schwenzer, A.; Yeh, C.-R.; Chambers, D.M.; Pesson, K.; Li, W.; Segura, T.; et al. Citrullination of fibronectin alters integrin clustering and focal adhesion stability promoting stromal cell invasion. Matrix Boil. 2019, 82, 86–104. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Llambi, F. Cell death signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef] [PubMed]
- Vossenaar, E.R.; Radstake, T.; Van Der Heijden, A.; Mansum, M.A.M.V.; Dieteren, C.; De Rooij, D.-J.; Barrera, P.; Zendman, A.; Van Venrooij, W.J. Expression and activity of citrullinating peptidylarginine deiminase enzymes in monocytes and macrophages. Ann. Rheum. Dis. 2004, 63, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Hojo-Nakashima, I.; Sato, R.; Nakashima, K.; Hagiwara, T.; Yamada, M. Dynamic Expression of Peptidylarginine Deiminase 2 in Human Monocytic Leukaemia THP-1 Cells during Macrophage Differentiation. J. Biochem. 2009, 146, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Darrah, E.; Rosen, A.; Giles, J.T.; Andrade, F. Peptidylarginine deiminase 2, 3 and 4 have distinct specificities against cellular substrates: Novel insights into autoantigen selection in rheumatoid arthritis. Ann. Rheum. Dis. 2011, 71, 92–98. [Google Scholar] [CrossRef]
- Lee, H.J.; Joo, M.; Abdolrasulnia, R.; Young, D.G.; Choi, I.; Ware, L.B. Peptidylarginine deiminase 2 suppresses inhibitory κB kinase activity in lipopolysaccharide-stimulated RAW 264.7 macrophages. J. Biol. Chem. 2010, 285, 39655–39662. [Google Scholar] [CrossRef]
- Mishra, N.; Schwerdtner, L.; Sams, K.; Mondal, S.; Ahmad, F.; Schmidt, R.E. Cutting edge: Protein arginine deiminase 2 and 4 regulate NLRP3 inflammasome-dependent IL-1beta maturation and ASC speck formation in macrophages. J. Immunol. 2019, 203, 795–800. [Google Scholar] [CrossRef]
- McInnes, I.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef]
- Macdonald, I.J.; Liu, S.-C.; Su, C.-M.; Wang, Y.-H.; Tsai, C.-H.; Tang, C.-H. Implications of Angiogenesis Involvement in Arthritis. Int. J. Mol. Sci. 2018, 19, 2012. [Google Scholar] [CrossRef]
- Chang, H.-H.; Liu, G.-Y.; Dwivedi, N.; Sun, B.; Okamoto, Y.; Kinslow, J.D.; Deane, K.D.; Demoruelle, M.K.; Norris, J.M.; Thompson, P.R.; et al. A molecular signature of preclinical rheumatoid arthritis triggered by dysregulated PTPN22. JCI Insight 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.-Y.; Liao, Y.-F.; Chang, W.-H.; Liu, C.-C.; Hsieh, M.-C.; Hsu, P.-C.; Tsay, G.J.; Hung, H.-C. Overexpression of peptidylarginine deiminase IV features in apoptosis of haematopoietic cells. Apoptosis 2006, 11, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Osma-García, I.C.; Punzón, C.; Fresno, M.; Diaz-Munoz, M.D. Dose-Dependent effects of prostaglandin E2 in macrophage adhesion and migration. Eur. J. Immunol. 2015, 46, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Bokoch, G.M. Biology of the p21-Activated Kinases. Ann. Rev. Biochem. 2003, 72, 743–781. [Google Scholar] [CrossRef] [PubMed]
- Rajah, A.; Boudreau, C.G.; Ilie, A.; Wee, T.L.; Tang, K.; Borisov, A.Z. Paxillin S273 phosphorylation regulates adhesion dynamics and cell migration through a common protein complex with PAK1 and betaPIX. Sci. Rep. 2019, 9, 11430. [Google Scholar] [CrossRef]
- Kuzina, E.; Kudriaeva, A.A.; Glagoleva, I.S.; Gabibov, A.G.; Belogurov, A.A.; Knorre, V.D. Deimination of the myelin basic protein decelerates its proteasome-mediated metabolism. Dokl. Biochem. Biophys. 2016, 469, 277–280. [Google Scholar] [CrossRef]
- Cau, L.; Pendaries, V.; Lhuillier, E.; Thompson, P.R.; Serre, G.; Takahara, H.; Méchin, M.-C.; Simon, M. Lowering relative humidity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 2017, 86, 106–113. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Lu, M.C.; Lai, N.S.; Yin, W.Y.; Yu, H.C.; Huang, H.B.; Tung, C.H. Anti-Citrullinated protein antibodies activated ERK1/2 and JNK mitogen-activated protein kinases via binding to surface-expressed citrullinated GRP78 on mononuclear cells. J. Clin. Immunol. 2013, 33, 558–566. [Google Scholar] [CrossRef]
- Lu, M.; Lai, N.; Chen, H.; Yu, H.; Huang, K.; Tung, C.; Huang, H.-B.; Yu, C.-L. Decreased microRNA(miR)-145 and increased miR-224 expression in T cells from patients with systemic lupus erythematosus involved in lupus immunopathogenesis. Clin. Exp. Immunol. 2012, 171, 91–99. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-C.; Tung, C.-H.; Huang, K.-Y.; Huang, H.-B.; Lu, M.-C. The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage. Int. J. Mol. Sci. 2020, 21, 5720. https://doi.org/10.3390/ijms21165720
Yu H-C, Tung C-H, Huang K-Y, Huang H-B, Lu M-C. The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage. International Journal of Molecular Sciences. 2020; 21(16):5720. https://doi.org/10.3390/ijms21165720
Chicago/Turabian StyleYu, Hui-Chun, Chien-Hsueh Tung, Kuang-Yung Huang, Hsien-Bin Huang, and Ming-Chi Lu. 2020. "The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage" International Journal of Molecular Sciences 21, no. 16: 5720. https://doi.org/10.3390/ijms21165720
APA StyleYu, H.-C., Tung, C.-H., Huang, K.-Y., Huang, H.-B., & Lu, M.-C. (2020). The Essential Role of Peptidylarginine Deiminases 2 for Cytokines Secretion, Apoptosis, and Cell Adhesion in Macrophage. International Journal of Molecular Sciences, 21(16), 5720. https://doi.org/10.3390/ijms21165720