The Role of Semaphorins in Metabolic Disorders

Abstract
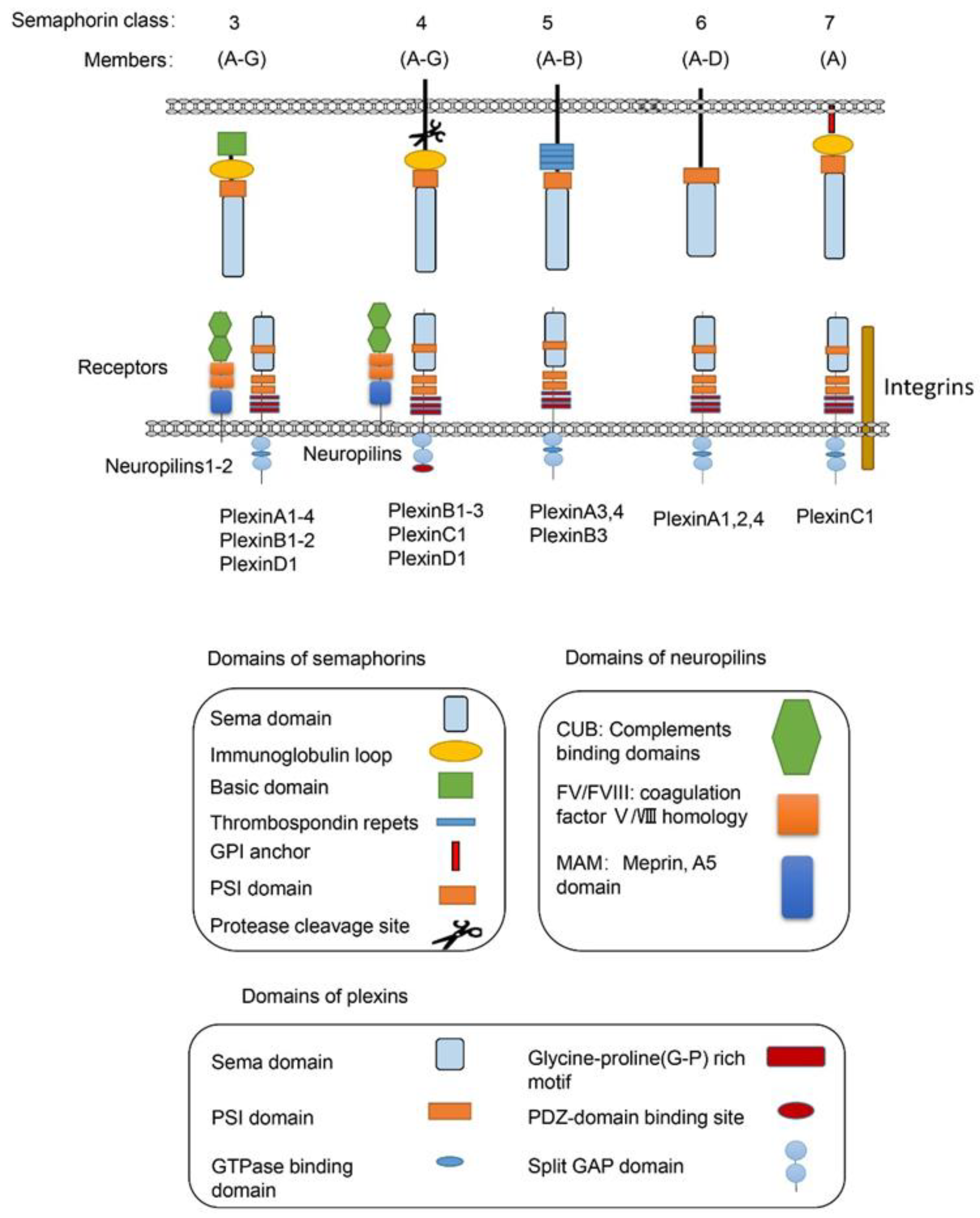
1. Introduction
2. Semaphorins in Metabolism
2.1. Semaphorins in Obesity
2.1.1. Semaphorins in Adipogenesis
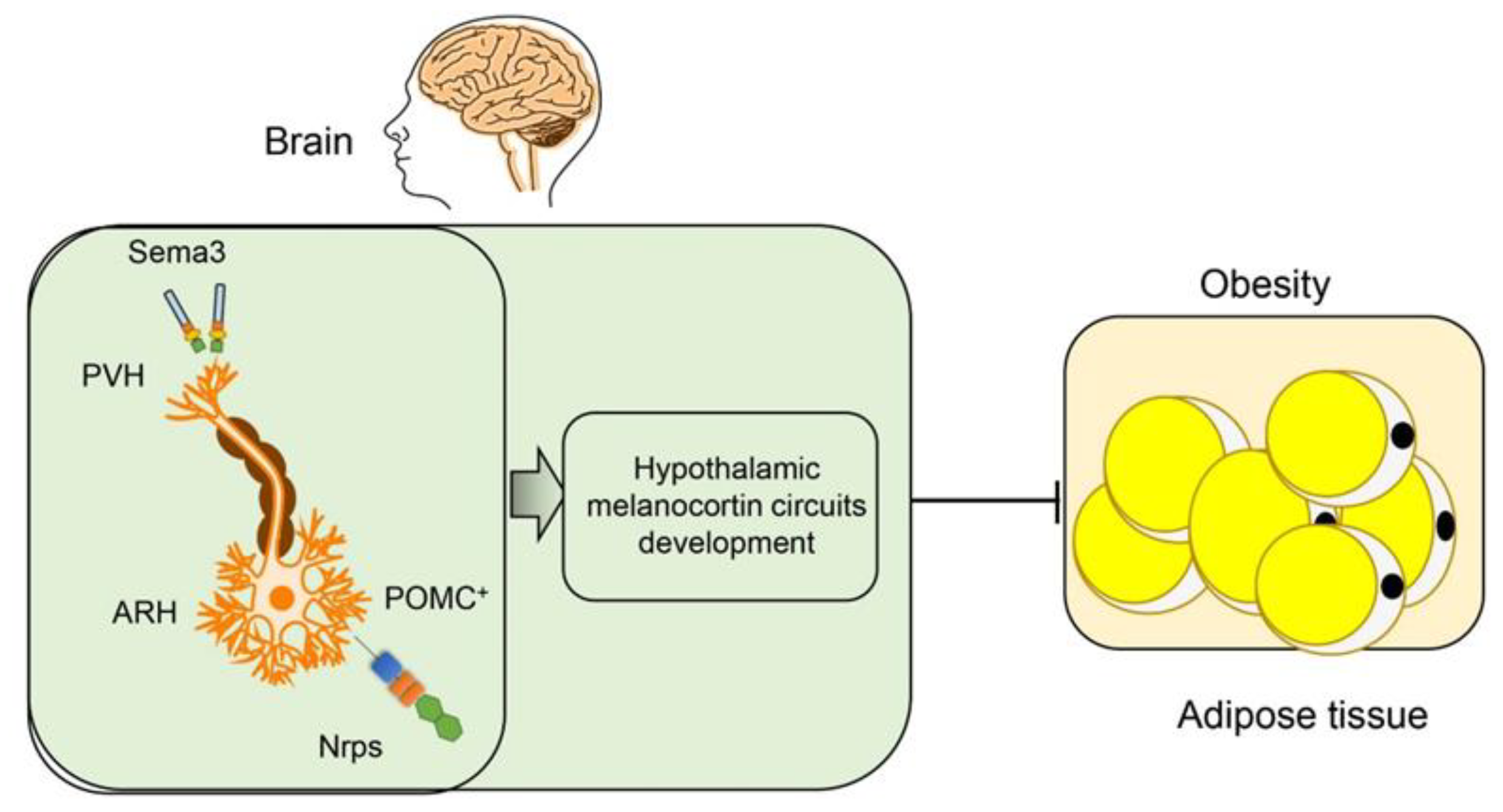
2.1.2. Semaphorins in Hypothalamic Regulation of Obesity
2.2. Semaphorins in Adipose Inflammation
2.3. Semaphorins in Brown Adipose Tissue
2.4. Semaphorins in Immune Cell Metabolism
3. Semaphorins in Diabetic Complications
3.1. Semaphorins in Diabetic Retinopathy
3.2. Semaphorins in Diabetic Nephropathy
3.3. Semaphorins in Diabetic Neuropathy
3.4. Semaphorins in Diabetic Wound Healing
3.5. Diabetic Osteoporosis
4. Miscellaneous
5. Perspectives
Funding
Conflicts of Interest
Abbreviations
| Sema3A | semaphorin 3A |
| ASCs | adipose mesenchymal stem cells |
| Nrp | neuropilin |
| POMC | proopiomelanocortin |
| ARH | arcuate nucleus of hypothalamus |
| NPY | neuropeptide Y |
| MC4R | melanocortin receptor 4 |
| GWAS | genome-wide association studies |
| PVH | paraventricular nucleus of the hypothalamus |
| WAT | white adipose tissue |
| BAT | brown adipose tissue |
| DR | diabetic retinopathy |
| OIR | oxygen-induced retinopathy |
| DN | diabetic nephropathy |
| ESRD | end-stage renal disease |
| DPN | diabetic peripheral neuropathy |
| NGF | neural growth factor |
| Arg | arginase |
References
- Yazdani, U.; Terman, J.R. The semaphorins. Genome Biol. 2006, 7, 211. [Google Scholar] [CrossRef] [PubMed]
- Bamberg, J.A.; Baumgartner, S.; Betz, H.; Bolz, J.; Chedotal, A.; Christensen, C.R.; Comoglio, P.M.; Culotti, J.G.; Doherty, P.; Drabkin, H.; et al. Unified nomenclature for the semaphorins/collapsins. Semaphorin Nomenclature Committee Cell 1999, 97, 551–552. [Google Scholar] [CrossRef]
- Cheng, S.; Ozkan, E. Finally on Track: Interactions of Off-Track with Plex-Sema Pathway and Glycosaminoglycans. Structure 2020, 28, 492–494. [Google Scholar] [CrossRef]
- Rozbesky, D.; Robinson, R.A.; Jain, V.; Renner, M.; Malinauskas, T.; Harlos, K.; Siebold, C.; Jones, E.Y. Diversity of oligomerization in Drosophila semaphorins suggests a mechanism of functional fine-tuning. Nat. Commun. 2019, 10, 3691. [Google Scholar] [CrossRef] [PubMed]
- Iragavarapu-Charyulu, V.; Wojcikiewicz, E.; Urdaneta, A. Semaphorins in Angiogenesis and Autoimmune Diseases: Therapeutic Targets? Front. Immunol. 2020, 11, 346. [Google Scholar] [CrossRef]
- Stedden, C.G.; Menegas, W.; Zajac, A.L.; Williams, A.M.; Cheng, S.; Ozkan, E.; Horne-Badovinac, S. Planar-Polarized Semaphorin-5c and Plexin A Promote the Collective Migration of Epithelial Cells in Drosophila. Curr. Biol. 2019, 29, 908–920. [Google Scholar] [CrossRef] [PubMed]
- Kolodkin, A.L.; Matthes, D.J.; Goodman, C.S. The semaphorin genes encode a family of transmembrane and secreted growth cone guidance molecules. Cell 1993, 75, 1389–1399. [Google Scholar] [CrossRef]
- Zhou, Y.; Gunput, R.A.; Pasterkamp, R.J. Semaphorin signaling: Progress made and promises ahead. Trends Biochem. Sci. 2008, 33, 161–170. [Google Scholar] [CrossRef]
- Tamagnone, L.; Artigiani, S.; Chen, H.; He, Z.; Ming, G.I.; Song, H.; Chedotal, A.; Winberg, M.L.; Goodman, C.S.; Poo, M.; et al. Plexins are a large family of receptors for transmembrane, secreted, and GPI-anchored semaphorins in vertebrates. Cell 1999, 99, 71–80. [Google Scholar] [CrossRef]
- Alto, L.T.; Terman, J.R. Semaphorins and their Signaling Mechanisms. Methods Mol. Biol. 2017, 1493, 1–25. [Google Scholar] [CrossRef]
- Wannemacher, K.M.; Wang, L.; Zhu, L.; Brass, L.F. The role of semaphorins and their receptors in platelets: Lessons learned from neuronal and immune synapses. Platelets 2011, 22, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.J.; Robinson, R.A.; Perez-Branguli, F.; Bell, C.H.; Mitchell, K.J.; Siebold, C.; Jones, E.Y. Structural basis of semaphorin-plexin signalling. Nature 2010, 467, 1118–1122. [Google Scholar] [CrossRef]
- Nogi, T.; Yasui, N.; Mihara, E.; Matsunaga, Y.; Noda, M.; Yamashita, N.; Toyofuku, T.; Uchiyama, S.; Goshima, Y.; Kumanogoh, A.; et al. Structural basis for semaphorin signalling through the plexin receptor. Nature 2010, 467, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.H.; Lohrum, M.; Klostermann, A.; Betz, H.; Puschel, A.W. The chemorepulsive activity of secreted semaphorins is regulated by furin-dependent proteolytic processing. EMBO J. 1997, 16, 6077–6086. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Yoshida, Y.; Livet, J.; Reimert, D.V.; Mann, F.; Merte, J.; Henderson, C.E.; Jessell, T.M.; Kolodkin, A.L.; Ginty, D.D. Semaphorin 3E and plexin-D1 control vascular pattern independently of neuropilins. Science 2005, 307, 265–268. [Google Scholar] [CrossRef]
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Chen, H.; Chedotal, A.; He, Z.; Goodman, C.S.; Tessier-Lavigne, M. Neuropilin-2, a novel member of the neuropilin family, is a high affinity receptor for the semaphorins Sema E and Sema IV but not Sema III. Neuron 1997, 19, 547–559. [Google Scholar] [CrossRef]
- Takahashi, T.; Fournier, A.; Nakamura, F.; Wang, L.H.; Murakami, Y.; Kalb, R.G.; Fujisawa, H.; Strittmatter, S.M. Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors. Cell 1999, 99, 59–69. [Google Scholar] [CrossRef]
- Neufeld, G.; Kessler, O. The semaphorins: Versatile regulators of tumour progression and tumour angiogenesis. Nat. Rev. Cancer 2008, 8, 632–645. [Google Scholar] [CrossRef]
- Gitler, A.D.; Lu, M.M.; Epstein, J.A. PlexinD1 and semaphorin signaling are required in endothelial cells for cardiovascular development. Dev. Cell 2004, 7, 107–116. [Google Scholar] [CrossRef]
- Toledano, S.; Nir-Zvi, I.; Engelman, R.; Kessler, O.; Neufeld, G. Class-3 Semaphorins and Their Receptors: Potent Multifunctional Modulators of Tumor Progression. Int. J. Mol. Sci. 2019, 20, 556. [Google Scholar] [CrossRef] [PubMed]
- Granziero, L.; Circosta, P.; Scielzo, C.; Frisaldi, E.; Stella, S.; Geuna, M.; Giordano, S.; Ghia, P.; Caligaris-Cappio, F. CD100/Plexin-B1 interactions sustain proliferation and survival of normal and leukemic CD5+ B lymphocytes. Blood 2003, 101, 1962–1969. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Li, Y.N.; Jin, H.J.; Wu, J.H.; He, Q.W.; Wang, X.X.; Lei, H.; Hu, B. Sema4D/PlexinB1 inhibition ameliorates blood-brain barrier damage and improves outcome after stroke in rats. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 2181–2196. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Jiang, S.; Dissanayaka, W.L.; Heng, B.C.; Xu, J.; Gong, T.; Huang, X.; Zhang, C. Sema4D/PlexinB1 promotes endothelial differentiation of dental pulp stem cells via activation of AKT and ERK1/2 signaling. J. Cell. Biochem. 2019, 120, 13614–13624. [Google Scholar] [CrossRef] [PubMed]
- Witherden, D.A.; Watanabe, M.; Garijo, O.; Rieder, S.E.; Sarkisyan, G.; Cronin, S.J.; Verdino, P.; Wilson, I.A.; Kumanogoh, A.; Kikutani, H.; et al. The CD100 receptor interacts with its plexin B2 ligand to regulate epidermal gammadelta T cell function. Immunity 2012, 37, 314–325. [Google Scholar] [CrossRef]
- He, Y.; Guo, Y.; Fan, C.; Lei, Y.; Zhou, Y.; Zhang, M.; Ye, C.; Ji, G.; Ma, L.; Lian, J.; et al. Interferon-alpha-Enhanced CD100/Plexin-B1/B2 Interactions Promote Natural Killer Cell Functions in Patients with Chronic Hepatitis C Virus Infection. Front. Immunol. 2017, 8, 1435. [Google Scholar] [CrossRef]
- Nishide, M.; Nojima, S.; Ito, D.; Takamatsu, H.; Koyama, S.; Kang, S.; Kimura, T.; Morimoto, K.; Hosokawa, T.; Hayama, Y.; et al. Semaphorin 4D inhibits neutrophil activation and is involved in the pathogenesis of neutrophil-mediated autoimmune vasculitis. Ann. Rheum. Dis. 2017, 76, 1440–1448. [Google Scholar] [CrossRef]
- Mizrahi, S.; Markel, G.; Porgador, A.; Bushkin, Y.; Mandelboim, O. CD100 on NK cells enhance IFNgamma secretion and killing of target cells expressing CD72. PLoS ONE 2007, 2, e818. [Google Scholar] [CrossRef]
- Kuklina, E.M. Receptor Functions of Semaphorin 4D. Biochemistry (Mosc) 2019, 84, 1021–1027. [Google Scholar] [CrossRef]
- Zhuang, J.; Li, X.; Zhang, Y.; Shi, R.; Shi, C.; Yu, D.; Bao, X.; Hu, M. Sema6A-plexin-A2 axis stimulates RANKL-induced osteoclastogenesis through PLCgamma-mediated NFATc1 activation. Life Sci. 2019, 222, 29–35. [Google Scholar] [CrossRef]
- Liu, H.; Juo, Z.S.; Shim, A.H.; Focia, P.J.; Chen, X.; Garcia, K.C.; He, X. Structural basis of semaphorin-plexin recognition and viral mimicry from Sema7A and A39R complexes with PlexinC1. Cell 2010, 142, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Liu, Y.; You, T.; Heath, J.; Xu, L.; Zheng, X.; Wang, A.; Wang, Y.; Li, F.; Yang, F.; et al. Vascular Semaphorin 7A Upregulation by Disturbed Flow Promotes Atherosclerosis Through Endothelial beta1 Integrin. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Jongbloets, B.C.; Lemstra, S.; Schellino, R.; Broekhoven, M.H.; Parkash, J.; Hellemons, A.J.; Mao, T.; Giacobini, P.; van Praag, H.; De Marchis, S.; et al. Stage-specific functions of Semaphorin7A during adult hippocampal neurogenesis rely on distinct receptors. Nat. Commun. 2017, 8, 14666. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, T.; Kikutani, H. Semaphorin signaling during cardiac development. Adv. Exp. Med. Biol. 2007, 600, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, T.; Yoshida, J.; Sugimoto, T.; Yamamoto, M.; Makino, N.; Takamatsu, H.; Takegahara, N.; Suto, F.; Hori, M.; Fujisawa, H.; et al. Repulsive and attractive semaphorins cooperate to direct the navigation of cardiac neural crest cells. Dev. Biol. 2008, 321, 251–262. [Google Scholar] [CrossRef]
- Giordano, S.; Corso, S.; Conrotto, P.; Artigiani, S.; Gilestro, G.; Barberis, D.; Tamagnone, L.; Comoglio, P.M. The semaphorin 4D receptor controls invasive growth by coupling with Met. Nat. Cell Biol. 2002, 4, 720–724. [Google Scholar] [CrossRef]
- Capparuccia, L.; Tamagnone, L. Semaphorin signaling in cancer cells and in cells of the tumor microenvironment--two sides of a coin. J. Cell Sci. 2009, 122, 1723–1736. [Google Scholar] [CrossRef]
- Neufeld, G.; Mumblat, Y.; Smolkin, T.; Toledano, S.; Nir-Zvi, I.; Ziv, K.; Kessler, O. The semaphorins and their receptors as modulators of tumor progression. Drug Resist. Updates 2016, 29, 1–12. [Google Scholar] [CrossRef]
- Li, Z.; Hao, J.; Duan, X.; Wu, N.; Zhou, Z.; Yang, F.; Li, J.; Zhao, Z.; Huang, S. The Role of Semaphorin 3A in Bone Remodeling. Front. Cell. Neurosci. 2017, 11, 40. [Google Scholar] [CrossRef]
- Takegahara, N.; Takamatsu, H.; Toyofuku, T.; Tsujimura, T.; Okuno, T.; Yukawa, K.; Mizui, M.; Yamamoto, M.; Prasad, D.V.; Suzuki, K.; et al. Plexin-A1 and its interaction with DAP12 in immune responses and bone homeostasis. Nat. Cell Biol. 2006, 8, 615–622. [Google Scholar] [CrossRef]
- Verlinden, L.; Vanderschueren, D.; Verstuyf, A. Semaphorin signaling in bone. Mol. Cell. Endocrinol. 2016, 432, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Lontos, K.; Adamik, J.; Tsagianni, A.; Galson, D.L.; Chirgwin, J.M.; Suvannasankha, A. The Role of Semaphorin 4D in Bone Remodeling and Cancer Metastasis. Front. Endocrinol. 2018, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Serini, G.; Valdembri, D.; Zanivan, S.; Morterra, G.; Burkhardt, C.; Caccavari, F.; Zammataro, L.; Primo, L.; Tamagnone, L.; Logan, M.; et al. Class 3 semaphorins control vascular morphogenesis by inhibiting integrin function. Nature 2003, 424, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, T.; Yabuki, M.; Kamei, J.; Kamei, M.; Makino, N.; Kumanogoh, A.; Hori, M. Semaphorin-4A, an activator for T-cell-mediated immunity, suppresses angiogenesis via Plexin-D1. EMBO J. 2007, 26, 1373–1384. [Google Scholar] [CrossRef]
- Iyer, A.S.; Chapoval, S.P. Neuroimmune Semaphorin 4A in Cancer Angiogenesis and Inflammation: A Promoter or a Suppressor? Int. J. Mol. Sci. 2018, 20, 124. [Google Scholar] [CrossRef]
- Ito, D.; Nojima, S.; Kumanogoh, A. The role of semaphorin family in immune systems. Nihon Rinsho Meneki Gakkai Kaishi 2014, 37, 1–10. [Google Scholar] [CrossRef][Green Version]
- Nishide, M.; Kumanogoh, A. The role of semaphorins in immune responses and autoimmune rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 19–31. [Google Scholar] [CrossRef]
- Suzuki, K.; Kumanogoh, A.; Kikutani, H. Semaphorins and their receptors in immune cell interactions. Nat. Immunol. 2008, 9, 17–23. [Google Scholar] [CrossRef]
- Lee, C.C.W.; Munuganti, R.S.N.; Peacock, J.W.; Dalal, K.; Jiao, I.Z.F.; Shepherd, A.; Liu, L.; Tam, K.J.; Sedgwick, C.G.; Bhasin, S.; et al. Targeting Semaphorin 3C in Prostate Cancer With Small Molecules. J. Endocr. Soc. 2018, 2, 1381–1394. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, C.; Sun, Q.; Wu, J.; Qiu, L.; Gao, C.; Liu, W.; Yang, J.; Jun, N.; Dong, J. The role of semaphorin 4D in tumor development and angiogenesis in human breast cancer. Oncotargets Ther. 2016, 9, 5737–5750. [Google Scholar] [CrossRef]
- Tarullo, S.E.; Hill, R.C.; Hansen, K.C.; Behbod, F.; Borges, V.F.; Nelson, A.C.; Lyons, T.R. Postpartum breast cancer progression is driven by semaphorin 7a-mediated invasion and survival. Oncogene 2020, 39, 2772–2785. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, D.; Son, E.; Yoo, S.J.; Sa, J.K.; Shin, Y.J.; Yoon, Y.; Nam, D.H. Pharmacokinetics, Biodistribution, and Toxicity Evaluation of Anti-SEMA3A (F11) in In Vivo Models. Anticancer Res. 2018, 38, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zeng, Z.; Qiao, L.; Jiang, X.; Ma, J.; Wang, J.; Ye, S.; Ma, Q.; Wei, J.; Wu, M.; et al. Semaphorin 4C Promotes Macrophage Recruitment and Angiogenesis in Breast Cancer. Mol. Cancer Res. Mcr 2019, 17, 2015–2028. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, L.; Liu, W.X.; Wang, K. VEGF and SEMA4D have synergistic effects on the promotion of angiogenesis in epithelial ovarian cancer. Cell. Mol. Biol. Lett. 2018, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Franco, A.; Eixarch, H.; Costa, C.; Gil, V.; Castillo, M.; Calvo-Barreiro, L.; Montalban, X.; Del Rio, J.A.; Espejo, C. Semaphorin 7A as a Potential Therapeutic Target for Multiple Sclerosis. Mol. Neurobiol. 2017, 54, 4820–4831. [Google Scholar] [CrossRef]
- Xie, J.; Wang, H. Semaphorin 7A as a potential immune regulator and promising therapeutic target in rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 10. [Google Scholar] [CrossRef]
- Yoshida, Y.; Ogata, A.; Kang, S.; Ebina, K.; Shi, K.; Nojima, S.; Kimura, T.; Ito, D.; Morimoto, K.; Nishide, M.; et al. Semaphorin 4D Contributes to Rheumatoid Arthritis by Inducing Inflammatory Cytokine Production: Pathogenic and Therapeutic Implications. Arthritis Rheumatol. 2015, 67, 1481–1490. [Google Scholar] [CrossRef]
- Eissa, N.; Hussein, H.; Diarra, A.; Elgazzar, O.; Gounni, A.S.; Bernstein, C.N.; Ghia, J.E. Semaphorin 3E regulates apoptosis in the intestinal epithelium during the development of colitis. Biochem. Pharmacol. 2019, 166, 264–273. [Google Scholar] [CrossRef]
- Kang, S.; Nakanishi, Y.; Kioi, Y.; Okuzaki, D.; Kimura, T.; Takamatsu, H.; Koyama, S.; Nojima, S.; Nishide, M.; Hayama, Y.; et al. Semaphorin 6D reverse signaling controls macrophage lipid metabolism and anti-inflammatory polarization. Nat. Immunol. 2018, 19, 561–570. [Google Scholar] [CrossRef]
- Delgoffe, G.M.; Woo, S.R.; Turnis, M.E.; Gravano, D.M.; Guy, C.; Overacre, A.E.; Bettini, M.L.; Vogel, P.; Finkelstein, D.; Bonnevier, J.; et al. Stability and function of regulatory T cells is maintained by a neuropilin-1-semaphorin-4a axis. Nature 2013, 501, 252–256. [Google Scholar] [CrossRef]
- Kenan, S.; Onur, O.D.; Solakoglu, S.; Kotil, T.; Ramazanoglu, M.; Celik, H.H.; Ocak, M.; Uzuner, B.; Firatli, E. Investigation of the effects of semaphorin 3A on new bone formation in a rat calvarial defect model. J. Cranio-Maxillo-Facial Surg. Off. Publ. Eur. Assoc. Cranio-Maxillo-Facial Surg. 2019, 47, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Parkash, J.; Messina, A.; Langlet, F.; Cimino, I.; Loyens, A.; Mazur, D.; Gallet, S.; Balland, E.; Malone, S.A.; Pralong, F.; et al. Semaphorin7A regulates neuroglial plasticity in the adult hypothalamic median eminence. Nat. Commun. 2015, 6, 6385. [Google Scholar] [CrossRef] [PubMed]
- Oleari, R.; Caramello, A.; Campinoti, S.; Lettieri, A.; Ioannou, E.; Paganoni, A.; Fantin, A.; Cariboni, A.; Ruhrberg, C. PLXNA1 and PLXNA3 cooperate to pattern the nasal axons that guide gonadotropin-releasing hormone neurons. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.H.; Li, Y.; Zhou, Y.F.; Haslam, J.; Elvis, O.N.; Mao, L.; Xia, Y.P.; Hu, B. Semaphorin-3E attenuates neointimal formation via suppressing VSMCs migration and proliferation. Cardiovasc. Res. 2017, 113, 1763–1775. [Google Scholar] [CrossRef]
- Sun, Q.; Peng, Y.; Zhao, Q.; Yan, S.; Liu, S.; Yang, Q.; Liu, K.; Rokosh, D.G.; Jiao, K. SEMA6D regulates perinatal cardiomyocyte proliferation and maturation in mice. Dev. Biol. 2019, 452, 1–7. [Google Scholar] [CrossRef]
- Sandireddy, R.; Cibi, D.M.; Gupta, P.; Singh, A.; Tee, N.; Uemura, A.; Epstein, J.A.; Singh, M.K. Semaphorin 3E/PlexinD1 signaling is required for cardiac ventricular compaction. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Lee, H.; Macpherson, L.J.; Parada, C.A.; Zuker, C.S.; Ryba, N.J.P. Rewiring the taste system. Nature 2017, 548, 330–333. [Google Scholar] [CrossRef]
- Frias, C.P.; Liang, J.; Bresser, T.; Scheefhals, L.; van Kesteren, M.; van Dorland, R.; Hu, H.Y.; Bodzeta, A.; van Bergen En Henegouwen, P.M.P.; Hoogenraad, C.C.; et al. Semaphorin4D Induces Inhibitory Synapse Formation by Rapid Stabilization of Presynaptic Boutons via MET Coactivation. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 4221–4237. [Google Scholar] [CrossRef]
- Liu, X.; Tan, N.; Zhou, Y.; Zhou, X.; Chen, H.; Wei, H.; Chen, J.; Xu, X.; Zhang, S.; Yang, G.; et al. Semaphorin 3A Shifts Adipose Mesenchymal Stem Cells towards Osteogenic Phenotype and Promotes Bone Regeneration In Vivo. Stem Cells Int. 2016, 2016, 2545214. [Google Scholar] [CrossRef]
- Liu, M.; Xie, S.; Liu, W.; Li, J.; Li, C.; Huang, W.; Li, H.; Song, J.; Zhang, H. Mechanism of SEMA3G knockdown-mediated attenuation of high-fat diet-induced obesity. J. Endocrinol. 2020, 244, 223–236. [Google Scholar] [CrossRef]
- van der Klaauw, A.A.; Croizier, S.; Mendes de Oliveira, E.; Stadler, L.K.J.; Park, S.; Kong, Y.; Banton, M.C.; Tandon, P.; Hendricks, A.E.; Keogh, J.M.; et al. Human Semaphorin 3 Variants Link Melanocortin Circuit Development and Energy Balance. Cell 2019, 176, 729–742 e718. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Yoshida, Y.; Moriya, J.; Nojima, A.; Uemura, A.; Kobayashi, Y.; Minamino, T. Semaphorin3E-induced inflammation contributes to insulin resistance in dietary obesity. Cell Metab. 2013, 18, 491–504. [Google Scholar] [CrossRef]
- Yoshida, Y.; Shimizu, I.; Hayashi, Y.; Ikegami, R.; Suda, M.; Katsuumi, G.; Wakasugi, T.; Nakao, M.; Nakagami, H.; Morishita, R.; et al. Peptide vaccine for semaphorin3E ameliorates systemic glucose intolerance in mice with dietary obesity. Sci. Rep. 2019, 9, 3858. [Google Scholar] [CrossRef] [PubMed]
- Mejhert, N.; Wilfling, F.; Esteve, D.; Galitzky, J.; Pellegrinelli, V.; Kolditz, C.I.; Viguerie, N.; Tordjman, J.; Naslund, E.; Trayhurn, P.; et al. Semaphorin 3C is a novel adipokine linked to extracellular matrix composition. Diabetologia 2013, 56, 1792–1801. [Google Scholar] [CrossRef]
- Giordano, A.; Coppari, R.; Castellucci, M.; Cinti, S. Sema3a is produced by brown adipocytes and its secretion is reduced following cold acclimation. J. Neurocytol. 2001, 30, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.; Boura-Halfon, S.; Cortese, N.; Haimon, Z.; Sar Shalom, H.; Kuperman, Y.; Kalchenko, V.; Brandis, A.; David, E.; Segal-Hayoun, Y.; et al. Brown-adipose-tissue macrophages control tissue innervation and homeostatic energy expenditure. Nat. Immunol. 2017, 18, 665–674. [Google Scholar] [CrossRef]
- Rabiee, A.; Kruger, M.; Ardenkjaer-Larsen, J.; Kahn, C.R.; Emanuelli, B. Distinct signalling properties of insulin receptor substrate (IRS)-1 and IRS-2 in mediating insulin/IGF-1 action. Cell. Signal. 2018, 47, 1–15. [Google Scholar] [CrossRef]
- Vecchie, A.; Dallegri, F.; Carbone, F.; Bonaventura, A.; Liberale, L.; Portincasa, P.; Fruhbeck, G.; Montecucco, F. Obesity phenotypes and their paradoxical association with cardiovascular diseases. Eur. J. Intern. Med. 2018, 48, 6–17. [Google Scholar] [CrossRef]
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef]
- Broughton, D.E.; Moley, K.H. Obesity and female infertility: Potential mediators of obesity’s impact. Fertil. Steril. 2017, 107, 840–847. [Google Scholar] [CrossRef]
- Liu, W.; Li, J.; Liu, M.; Zhang, H.; Wang, N. PPAR-gamma promotes endothelial cell migration by inducing the expression of Sema3g. J. Cell. Biochem. 2015, 116, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Masuda, T.; Fukaya, M.; Kataoka, H.; Mishina, M.; Yaginuma, H.; Watanabe, M.; Shimizu, T. Identification and characterization of a novel member of murine semaphorin family. Genes Cells Devoted Mol. Cell. Mech. 2005, 10, 785–792. [Google Scholar] [CrossRef]
- O’Brien, P.D.; Hinder, L.M.; Callaghan, B.C.; Feldman, E.L. Neurological consequences of obesity. Lancet Neurol. 2017, 16, 465–477. [Google Scholar] [CrossRef]
- Sohn, J.W. Network of hypothalamic neurons that control appetite. BMB Rep. 2015, 48, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.P.; Vong, L.; Olson, D.P.; Koda, S.; Krashes, M.J.; Ye, C.; Yang, Z.; Fuller, P.M.; Elmquist, J.K.; Lowell, B.B. MC4R-expressing glutamatergic neurons in the paraventricular hypothalamus regulate feeding and are synaptically connected to the parabrachial nucleus. Proc. Natl. Acad. Sci. USA 2014, 111, 13193–13198. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.D. Anatomy and regulation of the central melanocortin system. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Zeltser, L.M. Feeding circuit development and early-life influences on future feeding behaviour. Nat. Rev. Neurosci. 2018, 19, 302–316. [Google Scholar] [CrossRef]
- Zeltser, L.M. Axon Guidance Molecules Implicated in Early-Onset Obesity. Trends Neurosci. 2019, 42, 439–440. [Google Scholar] [CrossRef]
- Engin, A.B. Adipocyte-Macrophage Cross-Talk in Obesity. Adv. Exp. Med. Biol. 2017, 960, 327–343. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Moore, K.J. The Semaphorin 3E/PlexinD1 axis regulates macrophage inflammation in obesity. Cell Metab. 2013, 18, 461–462. [Google Scholar] [CrossRef]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochim. Biophys. Acta 2014, 1842, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.L.; Pearce, E.J. Metabolic pathways in immune cell activation and quiescence. Immunity 2013, 38, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Vijan, S. Type 2 Diabetes. Ann. Intern. Med. 2019, 171, ITC65–ITC80. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Wu, J.H.; Li, Y.N.; Chen, A.Q.; Hong, C.D.; Zhang, C.L.; Wang, H.L.; Zhou, Y.F.; Li, P.C.; Wang, Y.; Mao, L.; et al. Inhibition of Sema4D/PlexinB1 signaling alleviates vascular dysfunction in diabetic retinopathy. Embo Mol. Med. 2020, 12, e10154. [Google Scholar] [CrossRef]
- Kwon, S.H.; Shin, J.P.; Kim, I.T.; Park, D.H. Association of Plasma Semaphorin 3A With Phenotypes of Diabetic Retinopathy and Nephropathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2983–2989. [Google Scholar] [CrossRef]
- Cerani, A.; Tetreault, N.; Menard, C.; Lapalme, E.; Patel, C.; Sitaras, N.; Beaudoin, F.; Leboeuf, D.; De Guire, V.; Binet, F.; et al. Neuron-derived semaphorin 3A is an early inducer of vascular permeability in diabetic retinopathy via neuropilin-1. Cell Metab. 2013, 18, 505–518. [Google Scholar] [CrossRef]
- Dejda, A.; Mawambo, G.; Cerani, A.; Miloudi, K.; Shao, Z.; Daudelin, J.F.; Boulet, S.; Oubaha, M.; Beaudoin, F.; Akla, N.; et al. Neuropilin-1 mediates myeloid cell chemoattraction and influences retinal neuroimmune crosstalk. J. Clin. Investig. 2014, 124, 4807–4822. [Google Scholar] [CrossRef]
- Joyal, J.S.; Sitaras, N.; Binet, F.; Rivera, J.C.; Stahl, A.; Zaniolo, K.; Shao, Z.; Polosa, A.; Zhu, T.; Hamel, D.; et al. Ischemic neurons prevent vascular regeneration of neural tissue by secreting semaphorin 3A. Blood 2011, 117, 6024–6035. [Google Scholar] [CrossRef]
- Fukushima, Y.; Okada, M.; Kataoka, H.; Hirashima, M.; Yoshida, Y.; Mann, F.; Gomi, F.; Nishida, K.; Nishikawa, S.; Uemura, A. Sema3E-PlexinD1 signaling selectively suppresses disoriented angiogenesis in ischemic retinopathy in mice. J. Clin. Investig. 2011, 121, 1974–1985. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.; Ranganathan, P.; Jayakumar, C.; Nauta, F.L.; Gansevoort, R.T.; Weintraub, N.L.; Brands, M.; Ramesh, G. Urinary semaphorin 3A correlates with diabetic proteinuria and mediates diabetic nephropathy and associated inflammation in mice. J. Mol. Med. 2014, 92, 1245–1256. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Veron, D.; Bertuccio, C.A.; Marlier, A.; Reidy, K.; Garcia, A.M.; Jimenez, J.; Velazquez, H.; Kashgarian, M.; Moeckel, G.W.; Tufro, A. Podocyte vascular endothelial growth factor (Vegf(1)(6)(4)) overexpression causes severe nodular glomerulosclerosis in a mouse model of type 1 diabetes. Diabetologia 2011, 54, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.K.; Veron, D.; Thomas, D.B.; Siegel, D.; Moeckel, G.; Kashgarian, M.; Tufro, A. Semaphorin3a promotes advanced diabetic nephropathy. Diabetes 2015, 64, 1743–1759. [Google Scholar] [CrossRef]
- Chittka, D.; Banas, B.; Lennartz, L.; Putz, F.J.; Eidenschink, K.; Beck, S.; Stempfl, T.; Moehle, C.; Reichelt-Wurm, S.; Banas, M.C. Long-term expression of glomerular genes in diabetic nephropathy. Nephrol. Dial Transpl. 2018, 33, 1533–1544. [Google Scholar] [CrossRef]
- Ishibashi, R.; Takemoto, M.; Akimoto, Y.; Ishikawa, T.; He, P.; Maezawa, Y.; Sakamoto, K.; Tsurutani, Y.; Ide, S.; Ide, K.; et al. A novel podocyte gene, semaphorin 3G, protects glomerular podocyte from lipopolysaccharide-induced inflammation. Sci. Rep. 2016, 6, 25955. [Google Scholar] [CrossRef]
- Sambo, F.; Malovini, A.; Sandholm, N.; Stavarachi, M.; Forsblom, C.; Makinen, V.P.; Harjutsalo, V.; Lithovius, R.; Gordin, D.; Parkkonen, M.; et al. Novel genetic susceptibility loci for diabetic end-stage renal disease identified through robust naive Bayes classification. Diabetologia 2014, 57, 1611–1622. [Google Scholar] [CrossRef]
- Koeck, F.X.; Bobrik, V.; Fassold, A.; Grifka, J.; Kessler, S.; Straub, R.H. Marked loss of sympathetic nerve fibers in chronic Charcot foot of diabetic origin compared to ankle joint osteoarthritis. J. Orthop. Res. 2009, 27, 736–741. [Google Scholar] [CrossRef]
- Lee, P.S.; Gao, N.; Dike, M.; Shkilnyy, O.; Me, R.; Zhang, Y.; Yu, F.X. Opposing Effects of Neuropilin-1 and -2 on Sensory Nerve Regeneration in Wounded Corneas: Role of Sema3C in Ameliorating Diabetic Neurotrophic Keratopathy. Diabetes 2019, 68, 807–818. [Google Scholar] [CrossRef]
- Wu, L.Y.; Li, M.; Qu, M.L.; Li, X.; Pi, L.H.; Chen, Z.; Zhou, S.L.; Yi, X.Q.; Shi, X.J.; Wu, J.; et al. High glucose up-regulates Semaphorin 3A expression via the mTOR signaling pathway in keratinocytes: A potential mechanism and therapeutic target for diabetic small fiber neuropathy. Mol. Cell. Endocrinol. 2018, 472, 107–116. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, Q.; Luo, Y.; Nguyen, T.; Rosenblatt, M.I.; Guaiquil, V.H. Semaphorin3A induces nerve regeneration in the adult cornea-a switch from its repulsive role in development. PLoS ONE 2018, 13, e0191962. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chopp, M.; Szalad, A.; Lu, X.; Zhang, Y.; Wang, X.; Cepparulo, P.; Lu, M.; Li, C.; Zhang, Z.G. Exosomes Derived From Schwann Cells Ameliorate Peripheral Neuropathy in Type 2 Diabetic Mice. Diabetes 2020, 69, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, B.; Yu, Z.; Wang, T.; Song, Y.; Zhuang, R.; Wu, Y.; Su, Y.; Guo, S. Effects of CD100 promote wound healing in diabetic mice. J. Mol. Histol. 2018, 49, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Tao, J.; Chen, D.D.; Cai, J.J.; Irani, K.; Wang, Q.; Yuan, H.; Chen, A.F. MicroRNA miR-27b rescues bone marrow-derived angiogenic cell function and accelerates wound healing in type 2 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Wang, L.; Zhao, B.; Liu, C.; Liu, H.; Zhu, R.; Chen, B.; Li, L.; Zhao, D.; Mo, F.; et al. Diabetes Perturbs Bone Microarchitecture and Bone Strength through Regulation of Sema3A/IGF-1/beta-Catenin in Rats. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 55–66. [Google Scholar] [CrossRef]
- Qiao, Q.; Xu, X.; Song, Y.; Song, S.; Zhu, W.; Li, F. Semaphorin 3A promotes osteogenic differentiation of BMSC from type 2 diabetes mellitus rats. J. Mol. Histol. 2018, 49, 369–376. [Google Scholar] [CrossRef]
- Qiao, Q.; Song, Y.L.; Li, F.L. Semaphorin 3A-stimulated bone marrow mesenchymal stem cells sheets promotes osteogenesis of type 2 diabetic rat. Zhonghua Kou Qiang Yi Xue Za Zhi 2018, 53, 333–338. [Google Scholar] [CrossRef]
- Fang, K.; Song, W.; Wang, L.; Xu, X.; Tan, N.; Zhang, S.; Wei, H.; Song, Y. Semaphorin 3A-modified adipose-derived stem cell sheet may improve osseointegration in a type 2 diabetes mellitus rat model. Mol. Med. Rep. 2016, 14, 2449–2456. [Google Scholar] [CrossRef][Green Version]
- Honasoge, A.; Nudleman, E.; Smith, M.; Rajagopal, R. Emerging Insights and Interventions for Diabetic Retinopathy. Curr. Diabetes Rep. 2019, 19, 100. [Google Scholar] [CrossRef]
- Moran, E.P.; Wang, Z.; Chen, J.; Sapieha, P.; Smith, L.E.; Ma, J.X. Neurovascular cross talk in diabetic retinopathy: Pathophysiological roles and therapeutic implications. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H738–H749. [Google Scholar] [CrossRef]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Guttmann-Raviv, N.; Shraga-Heled, N.; Varshavsky, A.; Guimaraes-Sternberg, C.; Kessler, O.; Neufeld, G. Semaphorin-3A and semaphorin-3F work together to repel endothelial cells and to inhibit their survival by induction of apoptosis. J. Biol. Chem. 2007, 282, 26294–26305. [Google Scholar] [CrossRef] [PubMed]
- Hua, N.; Liu, H.; Qian, X.; Dong, L.; Wu, J.; Li, X. The effect of semaphorin 3A in the process of apoptosis in oxygen induced retinopathy in rats. Zhonghua yan ke za zhi 2014, 50, 440–447. [Google Scholar] [PubMed]
- Sapieha, P. Eyeing central neurons in vascular growth and reparative angiogenesis. Blood 2012, 120, 2182–2194. [Google Scholar] [CrossRef] [PubMed]
- Motani, K.; Kosako, H. Activation of stimulator of interferon genes (STING) induces ADAM17-mediated shedding of the immune semaphorin SEMA4D. J. Biol. Chem. 2018, 293, 7717–7726. [Google Scholar] [CrossRef] [PubMed]
- Meza Letelier, C.E.; San Martin Ojeda, C.A.; Ruiz Provoste, J.J.; Frugone Zaror, C.J. Pathophysiology of diabetic nephropathy: A literature review. Medwave 2017, 17, e6839. [Google Scholar] [CrossRef]
- Podgorski, P.; Konieczny, A.; Lis, L.; Witkiewicz, W.; Hruby, Z. Glomerular podocytes in diabetic renal disease. Adv. Clin. Exp. Med. 2019, 28, 1711–1715. [Google Scholar] [CrossRef]
- Woroniecka, K.I.; Park, A.S.; Mohtat, D.; Thomas, D.B.; Pullman, J.M.; Susztak, K. Transcriptome analysis of human diabetic kidney disease. Diabetes 2011, 60, 2354–2369. [Google Scholar] [CrossRef]
- Vincent, A.M.; Callaghan, B.C.; Smith, A.L.; Feldman, E.L. Diabetic neuropathy: Cellular mechanisms as therapeutic targets. Nat. Rev. Neurol. 2011, 7, 573–583. [Google Scholar] [CrossRef]
- Apelqvist, J. Diagnostics and treatment of the diabetic foot. Endocrine 2012, 41, 384–397. [Google Scholar] [CrossRef]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenges. Lancet. Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef]
- Sawano, T.; Tsuchihashi, R.; Watanabe, F.; Niimi, K.; Yamaguchi, W.; Yamaguchi, N.; Furuyama, T.; Tanaka, H.; Matsuyama, T.; Inagaki, S. Changes in L-arginine metabolism by Sema4D deficiency induce promotion of microglial proliferation in ischemic cortex. Neuroscience 2019, 406, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Pauerstein, P.T.; Tellez, K.; Willmarth, K.B.; Park, K.M.; Hsueh, B.; Efsun Arda, H.; Gu, X.; Aghajanian, H.; Deisseroth, K.; Epstein, J.A.; et al. A radial axis defined by semaphorin-to-neuropilin signaling controls pancreatic islet morphogenesis. Development 2017, 144, 3744–3754. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Jensen, P.; Huang, H.; Lund Christensen, G.; Billestrup, N.; Larsen, M.R. Characterization of the Molecular Mechanisms Underlying Glucose Stimulated Insulin Secretion from Isolated Pancreatic beta-cells Using Post-translational Modification Specific Proteomics (PTMomics). Mol. Cell. Proteom. 2018, 17, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, M.; Xia, J.; Friedel, R.H.; Offermanns, S.; Worzfeld, T. A systematic expression analysis implicates Plexin-B2 and its ligand Sema4C in the regulation of the vascular and endocrine system. Exp. Cell Res. 2010, 316, 2477–2486. [Google Scholar] [CrossRef]
- Maione, F.; Molla, F.; Meda, C.; Latini, R.; Zentilin, L.; Giacca, M.; Seano, G.; Serini, G.; Bussolino, F.; Giraudo, E. Semaphorin 3A is an endogenous angiogenesis inhibitor that blocks tumor growth and normalizes tumor vasculature in transgenic mouse models. J. Clin. Investig. 2009, 119, 3356–3372. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diseases | References | Major Findings | Semaphorins Involved |
|---|---|---|---|
| Cancer | Lee, Munuganti et al., 2018 [49] | Promising small molecule inhibitors bind to SEMA3C, and attenuate prostate cancer growth | Sema3C |
| Jiang, Chen et al., 2016 [50] | Sema4D influences cell proliferation, invasion, migration, and apoptosis of breast cancer cells | Sema4D | |
| Tarullo, Hill et al., 2020 [51] | Sema7A promotes breast cancer progression | Sema7A | |
| Angiogenesis | Lee, Kim et al., 2018 [52] | Therapeutic Sema3A antibody F11 attenuated angiogenesis in glioblastoma | Sema3A |
| Yang, Zeng et al., 2019 [53] | Sema4C promotes angiogenesis in breast Cancer | Sema4C | |
| Chen, Zhang et al., 2018 [54] | Sema4D has synergistic effects with vegf on the promotion of angiogenesis | Sema4D | |
| Multiple sclerosis | Gutierrez-Franco, Eixarch et al., 2017 [55] | Sema7A is involved in peripheral immunity and CNS inflammation in MS pathogenesis | Sema7A |
| Rheumatoid arthritis | Xie and Wang 2017 [56] | Sema7A promotes rheumatoid arthritis | Sema7A |
| Yoshida, Ogata et al., 2015 [57] | Sema4D Contributes to rheumatoid arthritis by inducing inflammatory cytokine production | Sema4D | |
| Colitis | Eissa, Hussein et al., 2019 [58] | Sema3E regulates apoptosis in the intestinal epithelium and inhibits colitis | Sema3E |
| Kang, Nakanishi et al., 2018 [59] | Sema6D is important for generation of intestinal resident CX3CR1hi macrophages and prevents development of colitis | Sema6D | |
| Delgoffe, Woo et al., 2013 [60] | Sema4A regulates established inflammatory colitis through Nrp1 | Sema4A | |
| Bone remodeling | Kenan, Onur et al., 2019 [61] | Sema3A prevents bone resorption by inhibiting osteoclasts and increases bone formation by inducing osteoblasts | Sema3A |
| Endocrine diseases | Parkash, Messina et al., 2015 [62] | Sema7A promotes projection of gonadotropin-releasing hormone (GnRH) neurons and maintains normal oestrous cyclicity and fertility | Sema7A |
| Oleari, Caramello et al., 2019 [63] | Sema3A regulates Gonadotropin-releasing hormone neurons through its receptor neuropilins, PlexinA1 and PlexinA3 | Sema3A | |
| Atherosclerosis | Hu, Liu et al., 2018 [32] | Disturbed flow regulated Sema7A promotes atherosclerosis | Sema7A |
| Wu, Li et al., 2017 [64] | Sema3E attenuates neoinitimal formation via suppressing VSMCs migration and proliferation | Sema3E | |
| Cardiovascular diseases | Sun, Peng et al., 2019 [65] | Sema6D regulates perinatal cardiomyocyte proliferation and maturation in mice | Sema6D |
| Sandireddy, Cibi et al., 2019 [66] | Sema3E-PlexinD1 signaling is required for cardiac ventricular compaction | Sema3E | |
| Neuronal diseases | Lee, Macpherson et al., 2017 [67] | Sema3A and Sema7A regulates bitter and sweet neurons, respectively | Sema7A Sema3A |
| Frias, Liang et al., 2019 [68] | Sema4D induces inhibitory synapse formation | Sema4D |
| Metabolic Functions | Semaphorins Involved | Receptors Involved | References |
|---|---|---|---|
| Adipogenesis | Sema3A | Not mentioned | [69] |
| Sema3G | Nrp 2 | [70] | |
| Hypothalamus regulation of obesity | Sema3A, Sema3B, Sema3C, Sema3D Sema3E, Sema3F, Sema3G | PlexinA1, PlexinA2, PlexinA3, PlexinA4, Nrp1, Nrp2 | [71] |
| Adipose inflammation and fibrosis | Sema3E | PlexinD1 | [72,73] |
| Sema3C | Not mentioned | [74] | |
| Brown adipose tissue function | Sema3A | Not mentioned | [75] |
| Sema6A | PlexinA4 | [76] | |
| Sema4B | Not mentioned | [77] | |
| Immune cell metabolism | Sema6D | PlexinA4 | [59] |
| Diabetic Complications | Semaphorins Involved | Receptors Involved | References |
|---|---|---|---|
| Diabetic retinopathy | Sema4D | PlexinB1 | [96] |
| Sema3A | Nrp1 | [97,98,99,100] | |
| Sema3E | PlexinD1 | [101] | |
| Diabetic nephropathy | Sema3A | Nrp1, PlexinA1 | [97,102,103,104] |
| Sema3E | Not mentioned | [105] | |
| Sema3G | Not mentioned | [105,106] | |
| Sema5A, Sema5E | Not mentioned | [105] | |
| Sema6D | Not mentioned | [107] | |
| Diabetic neuropathy | Sema3C | Nrp1, Nrp2 | [108,109] |
| Sema3A | Not mentioned | [110,111] | |
| Sema6A | Not mentioned | [112] | |
| Diabetic wound healing | Sema4D | PlexinB2 | [113] |
| Sema6A | Not mentioned | [114] | |
| Diabetic osteoporosis | Sema3A | Not mentioned | [115,116,117,118] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Zhu, L. The Role of Semaphorins in Metabolic Disorders. Int. J. Mol. Sci. 2020, 21, 5641. https://doi.org/10.3390/ijms21165641
Lu Q, Zhu L. The Role of Semaphorins in Metabolic Disorders. International Journal of Molecular Sciences. 2020; 21(16):5641. https://doi.org/10.3390/ijms21165641
Chicago/Turabian StyleLu, Qiongyu, and Li Zhu. 2020. "The Role of Semaphorins in Metabolic Disorders" International Journal of Molecular Sciences 21, no. 16: 5641. https://doi.org/10.3390/ijms21165641
APA StyleLu, Q., & Zhu, L. (2020). The Role of Semaphorins in Metabolic Disorders. International Journal of Molecular Sciences, 21(16), 5641. https://doi.org/10.3390/ijms21165641