Assay Systems for Profiling Deubiquitinating Activity


Abstract
1. Introduction
2. In vitro Deubiquitination Assays
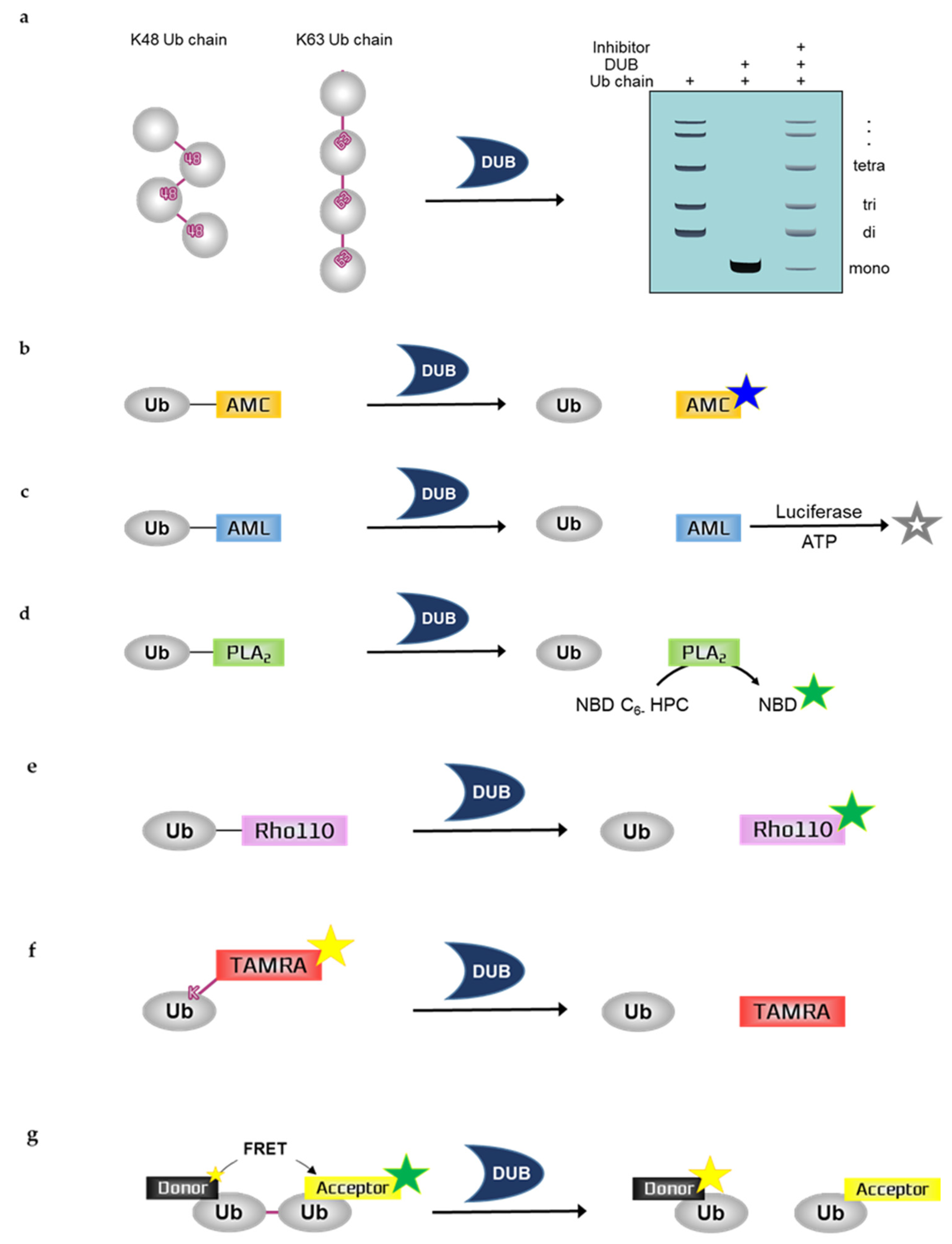
2.1. Ubiquitin Chain Cleavage Assay
2.2. Ubiquitin Probes
2.2.1. Ubiquitin-7-Amino-4-Methylcoumarin (Ub-AMC)
2.2.2. Ubiquitin-Aminoluciferin (Ub-AML)
2.2.3. Ubiquitin-Phospholipase A2 (Ub-PLA2)
2.2.4. Ubiquitin-Rhodamine110 (Ub-Rho110)
2.2.5. Fluorescence Polarization (FP)-Based Ubiquitin-Lys-5-Tetramethylrhodamine-Gly (Ub-Lys-TAMRA-Gly)
2.2.6. Fluorescence Resonance Energy Transfer (FRET)-Based Di-Ub Probes
2.2.7. Limitations
3. Deubiquitination Assays for Mammalian Cells
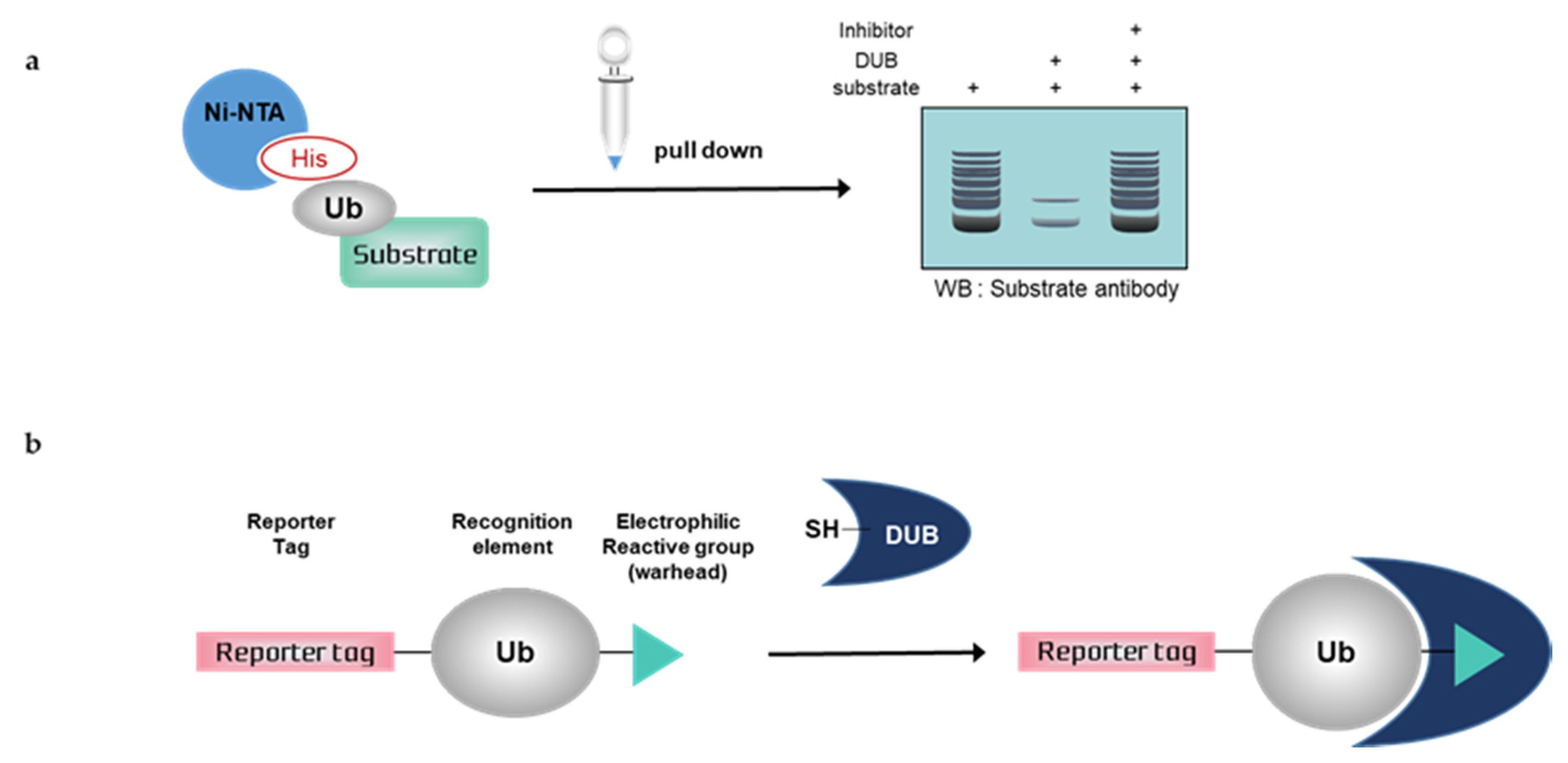
3.1. Nickel Pull-Down Assays
3.2. Activity-Based Probes (ABPs)
3.3. Cell-Permeable ABPs
3.3.1. Delivery of ABPs to Cells
3.3.2. Cell-Penetrating Peptide (CPP)-Based ABPs
3.3.3. Small Molecule-Based ABPs
3.4. Limitations
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DUB | Deubiquitinating Enzyme |
| ABP | Activity-Based Probe |
| HTS | High-Throughput Screening |
| Ub | Ubiquitin |
| AMC | 7-amino-4-methylcoumarin |
| AML | Aminoluciferin |
| PLA2 | Phospholipase A2 |
| NBD | 7-nitrobenz-2-oxa-1,3-diazole |
| NBD C6-HPC | 2-(6-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)hexanoyl-1-hexadecanoyl-sn-glycero-3-phosphocholine |
| Rho | Rhodamine 110 |
| FP | Fluorescence Polarization |
| TAMRA | 5-tetramethylrhodamine |
| FRET | Fluorescence Resonance Energy Transfer |
| PA | Propargylamide |
| VME | Vinylmethyl Ester |
| VS | Vinylsulfone |
| PFO | Perfringolysin O |
| CPP | Cell-Penetrating Peptide |
References
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Kulathu, Y.; Komander, D. Atypical ubiquitylation—the unexplored world of polyubiquitin beyond Lys48 and Lys63 linkages. Nat. Rev. Mol. Cell Biol. 2012, 13, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Komander, D. The emerging complexity of protein ubiquitination. Biochem. Soc. Trans. 2009, 37, 937–953. [Google Scholar] [CrossRef]
- Priolo, C.; Tang, D.; Brahamandan, M.; Benassi, B.; Sicinska, E.; Ogino, S.; Farsetti, A.; Porrello, A.; Finn, S.; Zimmermann, J.; et al. The isopeptidase USP2a protects human prostate cancer from apoptosis. Cancer Res. 2006, 66, 8625–8632. [Google Scholar] [CrossRef]
- Stevenson, L.F.; Sparks, A.; Allende-Vega, N.; Xirodimas, D.P.; Lane, D.P.; Saville, M.K. The deubiquitinating enzyme USP2a regulates the p53 pathway by targeting Mdm2. Embo. J. 2007, 26, 976–986. [Google Scholar] [CrossRef]
- Qu, Q.; Mao, Y.; Xiao, G.; Fei, X.; Wang, J.; Zhang, Y.; Liu, J.; Cheng, G.; Chen, X.; Wang, J.; et al. USP2 promotes cell migration and invasion in triple negative breast cancer cell lines. Tumour Biol. 2015, 36, 5415–5423. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Chakraborty, D.; Basu, M.; Ghosh, M.K. Emerging insights into HAUSP (USP7) in physiology, cancer and other diseases. Signal. Transduct Target Ther. 2018, 3, 17. [Google Scholar] [CrossRef]
- Wang, Z.; Kang, W.; You, Y.; Pang, J.; Ren, H.; Suo, Z.; Liu, H.; Zheng, Y. USP7: Novel Drug Target in Cancer Therapy. Front. Pharm. 2019, 10, 427. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Chen, C.; Yuan, H.; Wen, X.; Sun, H. USP7: Target Validation and Drug Discovery for Cancer Therapy. Med. Chem. 2018, 14, 3–18. [Google Scholar] [CrossRef]
- Pareja, F.; Ferraro, D.A.; Rubin, C.; Cohen-Dvashi, H.; Zhang, F.; Aulmann, S.; Ben-Chetrit, N.; Pines, G.; Navon, R.; Crosetto, N.; et al. Deubiquitination of EGFR by Cezanne-1 contributes to cancer progression. Oncogene 2012, 31, 4599–4608. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Gato, D.; Chuan, Y.C.; Jiang, N.; Svensson, C.; Bao, J.; Paul, I.; Egevad, L.; Kessler, B.M.; Wikström, P.; Niu, Y.; et al. OTUB1 de-ubiquitinating enzyme promotes prostate cancer cell invasion in vitro and tumorigenesis in vivo. Mol. Cancer 2015, 14, 8. [Google Scholar] [CrossRef]
- Liu, Y.; Fallon, L.; Lashuel, H.A.; Liu, Z.; Lansbury, P.T., Jr. The UCH-L1 gene encodes two opposing enzymatic activities that affect alpha-synuclein degradation and Parkinson’s disease susceptibility. Cell 2002, 111, 209–218. [Google Scholar] [CrossRef]
- Xilouri, M.; Kyratzi, E.; Pitychoutis, P.M.; Papadopoulou-Daifoti, Z.; Perier, C.; Vila, M.; Maniati, M.; Ulusoy, A.; Kirik, D.; Park, D.S.; et al. Selective neuroprotective effects of the S18Y polymorphic variant of UCH-L1 in the dopaminergic system. Hum. Mol. Genet. 2012, 21, 874–889. [Google Scholar] [CrossRef] [PubMed]
- Rott, R.; Szargel, R.; Haskin, J.; Bandopadhyay, R.; Lees, A.J.; Shani, V.; Engelender, S. alpha-Synuclein fate is determined by USP9X-regulated monoubiquitination. Proc. Natl. Acad. Sci. USA 2011, 108, 18666–18671. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, B.J.; Wilson, S.M.; Jung, H.; Miller, R.J. Altered neurotransmitter release machinery in mice deficient for the deubiquitinating enzyme Usp14. Am. J. Physiol. Cell Physiol. 2012, 302, C698–C708. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Serricchio, M.; Jauregui, M.; Shanbhag, R.; Stoltz, T.; Di Paolo, C.T.; Kim, P.K.; McQuibban, G.A. Deubiquitinating enzymes regulate PARK2-mediated mitophagy. Autophagy 2015, 11, 595–606. [Google Scholar] [CrossRef]
- Ahmed, N.; Zeng, M.; Sinha, I.; Polin, L.; Wei, W.Z.; Rathinam, C.; Flavell, R.; Massoumi, R.; Venuprasad, K. The E3 ligase Itch and deubiquitinase Cyld act together to regulate Tak1 and inflammation. Nat. Immunol. 2011, 12, 1176–1183. [Google Scholar] [CrossRef]
- Xu, D.; Lillico, S.G.; Barnett, M.W.; Whitelaw, C.B.; Archibald, A.L.; Ait-Ali, T. USP18 restricts PRRSV growth through alteration of nuclear translocation of NF-kappaB p65 and p50 in MARC-145 cells. Virus Res. 2012, 169, 264–267. [Google Scholar] [CrossRef]
- Nag, D.K.; Finley, D. A small-molecule inhibitor of deubiquitinating enzyme USP14 inhibits Dengue virus replication. Virus Res. 2012, 165, 103–106. [Google Scholar] [CrossRef]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Ghosh, M.K. Cell death and deubiquitinases: Perspectives in cancer. Biomed. Res. Int 2014, 2014, 435197. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Yan, F.; Ying, M.; Cao, J.; He, Q.; Zhu, H.; Yang, B. Inhibition of Ubiquitin-Specific Proteases as a Novel Anticancer Therapeutic Strategy. Front. Pharm. 2018, 9, 1080. [Google Scholar] [CrossRef]
- Smolskaya, S.; Logashina, Y.A.; Andreev, Y.A. Escherichia coli Extract-Based Cell-Free Expression System as an Alternative for Difficult-to-Obtain Protein Biosynthesis. Int. J. Mol. Sci. 2020, 21, 928. [Google Scholar] [CrossRef]
- Xu, D.; Shan, B.; Lee, B.H.; Zhu, K.; Zhang, T.; Sun, H.; Liu, M.; Shi, L.; Liang, W.; Qian, L.; et al. Phosphorylation and activation of ubiquitin-specific protease-14 by Akt regulates the ubiquitin-proteasome system. eLife 2015, 4, e10510. [Google Scholar] [CrossRef] [PubMed]
- Sonego, M.; Pellarin, I.; Costa, A.; Vinciguerra, G.L.R.; Coan, M.; Kraut, A.; D’Andrea, S.; Dall’Acqua, A.; Castillo-Tong, D.C.; Califano, D.; et al. USP1 links platinum resistance to cancer cell dissemination by regulating Snail stability. Sci Adv. 2019, 5, eaav3235. [Google Scholar] [CrossRef]
- Abdul Rehman, S.A.; Kristariyanto, Y.A.; Choi, S.Y.; Nkosi, P.J.; Weidlich, S.; Labib, K.; Hofmann, K.; Kulathu, Y. MINDY-1 Is a Member of an Evolutionarily Conserved and Structurally Distinct New Family of Deubiquitinating Enzymes. Mol. Cell 2016, 63, 146–155. [Google Scholar] [CrossRef]
- Park, J.; Kwon, M.S.; Kim, E.E.; Lee, H.; Song, E.J. USP35 regulates mitotic progression by modulating the stability of Aurora B. Nat. Commun. 2018, 9, 688. [Google Scholar] [CrossRef]
- Shi, T.; Bao, J.; Wang, N.X.; Zheng, J.; Wu, D. Identification of Small Molecule TRABID Deubiquitinase Inhibitors By Computation-Based Virtual Screen. BMC Chem. Biol. 2012, 12, 4. [Google Scholar] [CrossRef]
- Dang, L.C.; Melandri, F.D.; Stein, R.L. Kinetic and mechanistic studies on the hydrolysis of ubiquitin C-terminal 7-amido-4-methylcoumarin by deubiquitinating enzymes. Biochemistry 1998, 37, 1868–1879. [Google Scholar] [CrossRef]
- Morgan, M.T.; Wolberger, C. Competition Assay for Measuring Deubiquitinating Enzyme Substrate Affinity. Methods Mol. Biol. 2018, 1844, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.H.; Kim, T.; Nguyen, T.; Hahn, M.J.; Yun, S.I.; Kim, K.K. A Selective Inhibitor of Ubiquitin-Specific Protease 4 Suppresses Colorectal Cancer Progression by Regulating β-Catenin Signaling. Cell. Physiol. Biochem. 2019, 53, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Kapuria, V.; Peterson, L.F.; Fang, D.; Bornmann, W.G.; Talpaz, M.; Donato, N.J. Deubiquitinase inhibition by small-molecule WP1130 triggers aggresome formation and tumor cell apoptosis. Cancer Res. 2010, 70, 9265–9276. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, S.J.; Wu, J.; Eddins, M.J.; Leach, C.A.; Strickler, J.E. Bioluminescence assay platform for selective and sensitive detection of Ub/Ubl proteases. Biochim. Biophys. Acta 2012, 1823, 2079–2086. [Google Scholar] [CrossRef]
- Pinto, A.; Mace, Y.; Drouet, F.; Bony, E.; Boidot, R.; Draoui, N.; Lobysheva, I.; Corbet, C.; Polet, F.; Martherus, R.; et al. A new ER-specific photosensitizer unravels (1)O2-driven protein oxidation and inhibition of deubiquitinases as a generic mechanism for cancer PDT. Oncogene 2016, 35, 3976–3985. [Google Scholar] [CrossRef]
- Nicholson, B.; Leach, C.A.; Goldenberg, S.J.; Francis, D.M.; Kodrasov, M.P.; Tian, X.; Shanks, J.; Sterner, D.E.; Bernal, A.; Mattern, M.R.; et al. Characterization of ubiquitin and ubiquitin-like-protein isopeptidase activities. Protein Sci. 2008, 17, 1035–1043. [Google Scholar] [CrossRef]
- Altun, M.; Kramer, H.B.; Willems, L.I.; McDermott, J.L.; Leach, C.A.; Goldenberg, S.J.; Kumar, K.G.; Konietzny, R.; Fischer, R.; Kogan, E.; et al. Activity-based chemical proteomics accelerates inhibitor development for deubiquitylating enzymes. Chem. Bio.l 2011, 18, 1401–1412. [Google Scholar] [CrossRef]
- Hassiepen, U.; Eidhoff, U.; Meder, G.; Bulber, J.F.; Hein, A.; Bodendorf, U.; Lorthiois, E.; Martoglio, B. A sensitive fluorescence intensity assay for deubiquitinating proteases using ubiquitin-rhodamine110-glycine as substrate. Anal. Biochem. 2007, 371, 201–207. [Google Scholar] [CrossRef]
- Lamberto, I.; Liu, X.; Seo, H.S.; Schauer, N.J.; Iacob, R.E.; Hu, W.; Das, D.; Mikhailova, T.; Weisberg, E.L.; Engen, J.R.; et al. Structure-Guided Development of a Potent and Selective Non-covalent Active-Site Inhibitor of USP7. Cell Chem. Biol. 2017, 24, 1490–1500.e1411. [Google Scholar] [CrossRef]
- Lee, J.H.; Shin, S.K.; Jiang, Y.; Choi, W.H.; Hong, C.; Kim, D.E.; Lee, M.J. Facilitated Tau Degradation by USP14 Aptamers via Enhanced Proteasome Activity. Sci. Rep. 2015, 5, 10757. [Google Scholar] [CrossRef]
- Moerke, N.J. Fluorescence Polarization (FP) Assays for Monitoring Peptide-Protein or Nucleic Acid-Protein Binding. Curr. Protoc. Chem. Biol. 2009, 1, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Aulabaugh, A. Application of Fluorescence Polarization in HTS Assays. Methods Mol. Biol. 2016, 1439, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Du, Y. Fluorescence polarization assay to quantify protein-protein interactions in an HTS format. Methods Mol. Biol. 2015, 1278, 529–544. [Google Scholar] [CrossRef]
- Geurink, P.P.; El Oualid, F.; Jonker, A.; Hameed, D.S.; Ovaa, H. A general chemical ligation approach towards isopeptide-linked ubiquitin and ubiquitin-like assay reagents. Chembiochem 2012, 13, 293–297. [Google Scholar] [CrossRef]
- De Cesare, V.; Lopez, D.C.; Mabbitt, P.D.; Antico, O.; Wood, N.T.; Virdee, S. Deubiquitinating Enzyme Amino Acid Profiling Reveals a Class of Ubiquitin Esterases. bioRxiv 2020. [Google Scholar] [CrossRef]
- Juenemann, K.; Jansen, A.H.P.; van Riel, L.; Merkx, R.; Mulder, M.P.C.; An, H.; Statsyuk, A.; Kirstein, J.; Ovaa, H.; Reits, E.A. Dynamic recruitment of ubiquitin to mutant huntingtin inclusion bodies. Sci. Rep. 2018, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, P.; Ohayon, S.; Nawatha, M.; Brik, A. Chemical and semisynthetic approaches to study and target deubiquitinases. Chem. Soc. Rev. 2016, 45, 4171–4198. [Google Scholar] [CrossRef]
- Flierman, D.; van der Heden van Noort, G.J.; Ekkebus, R.; Geurink, P.P.; Mevissen, T.E.; Hospenthal, M.K.; Komander, D.; Ovaa, H. Non-hydrolyzable Diubiquitin Probes Reveal Linkage-Specific Reactivity of Deubiquitylating Enzymes Mediated by S2 Pockets. Cell Chem. Biol. 2016, 23, 472–482. [Google Scholar] [CrossRef]
- Piston, D.W.; Rizzo, M.A. FRET by fluorescence polarization microscopy. Methods Cell Biol. 2008, 85, 415–430. [Google Scholar] [CrossRef]
- Geurink, P.P.; van Tol, B.D.; van Dalen, D.; Brundel, P.J.; Mevissen, T.E.; Pruneda, J.N.; Elliott, P.R.; van Tilburg, G.B.; Komander, D.; Ovaa, H. Development of Diubiquitin-Based FRET Probes to Quantify Ubiquitin Linkage Specificity of Deubiquitinating Enzymes. Chembiochem 2016, 17, 816–820. [Google Scholar] [CrossRef]
- Hameed, D.S.; Sapmaz, A.; Ovaa, H. How Chemical Synthesis of Ubiquitin Conjugates Helps To Understand Ubiquitin Signal Transduction. Bioconjug. Chem. 2017, 28, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Y.; Zhang, J.; Cui, H.; Shao, Z.S.; Cheng, C.; Wang, Y.B.; Wang, H.S. Fluorescence resonance energy transfer (FRET)-based nanoarchitecture for monitoring deubiquitinating enzyme activity. Chem. Commun. (Camb) 2020, 56, 3183–3186. [Google Scholar] [CrossRef] [PubMed]
- Tirat, A.; Schilb, A.; Riou, V.; Leder, L.; Gerhartz, B.; Zimmermann, J.; Worpenberg, S.; Eidhoff, U.; Freuler, F.; Stettler, T.; et al. Synthesis and characterization of fluorescent ubiquitin derivatives as highly sensitive substrates for the deubiquitinating enzymes UCH-L3 and USP-2. Anal. Biochem. 2005, 343, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, S.J.; McDermott, J.L.; Butt, T.R.; Mattern, M.R.; Nicholson, B. Strategies for the identification of novel inhibitors of deubiquitinating enzymes. Biochem. Soc. Trans. 2008, 36, 828–832. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, J.; Wu, J.; Cao, P.; Kingsbury, W.D.; McDermott, J.L.; Kodrasov, M.P.; McKelvey, D.M.; Suresh Kumar, K.G.; Goldenberg, S.J.; Mattern, M.R.; et al. Selective Dual Inhibitors of the Cancer-Related Deubiquitylating Proteases USP7 and USP47. ACS Med. Chem. Lett. 2012, 3, 789–792. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.A. 2.32-Heterologous Protein Expression. In Comprehensive Biotechnology, 2nd ed.; Moo-Young, M., Ed.; Academic Press: Burlington, MA, USA, 2011; pp. 441–449. [Google Scholar]
- Spriestersbach, A.; Kubicek, J.; Schäfer, F.; Block, H.; Maertens, B. Purification of His-Tagged Proteins. Methods Enzym. 2015, 559, 1–15. [Google Scholar] [CrossRef]
- Cho, J.; Park, J.; Shin, S.C.; Kim, J.H.; Kim, E.E.; Song, E.J. Ribosomal protein S2 interplays with MDM2 to induce p53. Biochem. Biophys. Res. Commun. 2020, 523, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Park, J.; Shin, S.C.; Jang, M.; Kim, J.H.; Kim, E.E.; Song, E.J. USP47 Promotes Tumorigenesis by Negative Regulation of p53 through Deubiquitinating Ribosomal Protein S2. Cancers (Basel) 2020, 12, 1137. [Google Scholar] [CrossRef] [PubMed]
- Reverdy, C.; Conrath, S.; Lopez, R.; Planquette, C.; Atmanene, C.; Collura, V.; Harpon, J.; Battaglia, V.; Vivat, V.; Sippl, W.; et al. Discovery of specific inhibitors of human USP7/HAUSP deubiquitinating enzyme. Chem. Biol. 2012, 19, 467–477. [Google Scholar] [CrossRef]
- Hewings, D.S.; Flygare, J.A.; Bogyo, M.; Wertz, I.E. Activity-based probes for the ubiquitin conjugation-deconjugation machinery: New chemistries, new tools, and new insights. FEBS J. 2017, 284, 1555–1576. [Google Scholar] [CrossRef]
- Ward, J.A.; McLellan, L.; Stockley, M.; Gibson, K.R.; Whitlock, G.A.; Knights, C.; Harrigan, J.A.; Jacq, X.; Tate, E.W. Quantitative Chemical Proteomic Profiling of Ubiquitin Specific Proteases in Intact Cancer Cells. ACS Chem. Biol. 2016, 11, 3268–3272. [Google Scholar] [CrossRef] [PubMed]
- Kramer, H.B.; Nicholson, B.; Kessler, B.M.; Altun, M. Detection of ubiquitin-proteasome enzymatic activities in cells: Application of activity-based probes to inhibitor development. Biochim. Biophys. Acta 2012, 1823, 2029–2037. [Google Scholar] [CrossRef] [PubMed]
- Sanman, L.E.; Bogyo, M. Activity-based profiling of proteases. Annu. Rev. Biochem. 2014, 83, 249–273. [Google Scholar] [CrossRef]
- Claessen, J.H.L.; Witte, M.D.; Yoder, N.C.; Zhu, A.Y.; Spooner, E.; Ploegh, H.L. Catch-and-release probes applied to semi-intact cells reveal ubiquitin-specific protease expression in Chlamydia trachomatis infection. Chembiochem 2013, 14, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.; Claessen, J.H.; Ploegh, H.L. A viral deubiquitylating enzyme restores dislocation of substrates from the endoplasmic reticulum (ER) in semi-intact cells. J. Biol. Chem. 2012, 287, 23594–23603. [Google Scholar] [CrossRef]
- Mulder, M.P.; Witting, K.; Berlin, I.; Pruneda, J.N.; Wu, K.P.; Chang, J.G.; Merkx, R.; Bialas, J.; Groettrup, M.; Vertegaal, A.C.; et al. A cascading activity-based probe sequentially targets E1-E2-E3 ubiquitin enzymes. Nat. Chem. Biol. 2016, 12, 523–530. [Google Scholar] [CrossRef]
- Van Wijk, S.J.; Fulda, S.; Dikic, I.; Heilemann, M. Visualizing ubiquitination in mammalian cells. EMBO Rep. 2019, 20, e46520. [Google Scholar] [CrossRef]
- Raucher, D.; Ryu, J.S. Cell-penetrating peptides: Strategies for anticancer treatment. Trends Mol. Med. 2015, 21, 560–570. [Google Scholar] [CrossRef]
- Gui, W.; Ott, C.A.; Yang, K.; Chung, J.S.; Shen, S.; Zhuang, Z. Cell-Permeable Activity-Based Ubiquitin Probes Enable Intracellular Profiling of Human Deubiquitinases. J. Am. Chem. Soc. 2018, 140, 12424–12433. [Google Scholar] [CrossRef]
- Safa, N.; Anderson, J.C.; Vaithiyanathan, M.; Pettigrew, J.H.; Pappas, G.A.; Liu, D.; Gauthier, T.J.; Melvin, A.T. CPProtectides: Rapid uptake of well-folded beta-hairpin peptides with enhanced resistance to intracellular degradation. Pept. Sci. (Hoboken) 2019, 111, e24092. [Google Scholar] [CrossRef]
- Safa, N.; Pettigrew, J.H.; Gauthier, T.J.; Melvin, A.T. Direct measurement of deubiquitinating enzyme activity in intact cells using a protease-resistant, cell-permeable, peptide-based reporter. Biochem. Eng. J. 2019, 151, 107320. [Google Scholar] [CrossRef]
- Panyain, N.; Godinat, A.; Lanyon-Hogg, T.; Lachiondo-Ortega, S.; Will, E.J.; Soudy, C.; Mason, K.; Elkhalifa, S.; Smith, L.M.; Harrigan, J.A. Discovery of a potent and selective covalent inhibitor and activity-based probe for the deubiquitylating enzyme UCHL1, with anti-fibrotic activity. ChemRxiv 2019. [Google Scholar] [CrossRef]
- Geurink, P.P.; Kooij, R.; Sapmaz, A.; Liu, S.; Xin, B.-T.; Janssen, G.M.; van Veelen, P.A.; ten Dijke, P.; Ovaa, H. A small-molecule activity-based probe for monitoring ubiquitin C-terminal hydrolase L1 (UCHL1) activity in live cells and zebrafish embryos. BioRxiv 2019, 827642. [Google Scholar] [CrossRef]
- Borodovsky, A.; Ovaa, H.; Kolli, N.; Gan-Erdene, T.; Wilkinson, K.D.; Ploegh, H.L.; Kessler, B.M. Chemistry-based functional proteomics reveals novel members of the deubiquitinating enzyme family. Chem. Biol. 2002, 9, 1149–1159. [Google Scholar] [CrossRef]
- Love, K.R.; Pandya, R.K.; Spooner, E.; Ploegh, H.L. Ubiquitin C-terminal electrophiles are activity-based probes for identification and mechanistic study of ubiquitin conjugating machinery. ACS Chem. Biol. 2009, 4, 275–287. [Google Scholar] [CrossRef]
- McGouran, J.F.; Gaertner, S.R.; Altun, M.; Kramer, H.B.; Kessler, B.M. Deubiquitinating enzyme specificity for ubiquitin chain topology profiled by di-ubiquitin activity probes. Chem. Biol. 2013, 20, 1447–1455. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yin, L.; Cooper, E.M.; Lai, M.Y.; Dickey, S.; Pickart, C.M.; Fushman, D.; Wilkinson, K.D.; Cohen, R.E.; Wolberger, C. Evidence for bidentate substrate binding as the basis for the K48 linkage specificity of otubain 1. J. Mol. Biol. 2009, 386, 1011–1023. [Google Scholar] [CrossRef]
- Iphofer, A.; Kummer, A.; Nimtz, M.; Ritter, A.; Arnold, T.; Frank, R.; van den Heuvel, J.; Kessler, B.M.; Jansch, L.; Franke, R. Profiling ubiquitin linkage specificities of deubiquitinating enzymes with branched ubiquitin isopeptide probes. Chembiochem 2012, 13, 1416–1420. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Davidson, G.A.; Gui, W.; Yang, K.; Bozza, W.P.; Zhuang, Z. Activity-based ubiquitin-protein probes reveal target protein specificity of deubiquitinating enzymes. Chem. Sci. 2018, 9, 7859–7865. [Google Scholar] [CrossRef]
- Gjonaj, L.; Sapmaz, A.; Gonzalez-Prieto, R.; Vertegaal, A.C.O.; Flierman, D.; Ovaa, H. USP7: Combining tools towards selectivity. Chem. Commun. (Camb) 2019, 55, 5075–5078. [Google Scholar] [CrossRef] [PubMed]
- Gjonaj, L.; Sapmaz, A.; Flierman, D.; Janssen, G.M.C.; van Veelen, P.A.; Ovaa, H. Development of a DUB-selective fluorogenic substrate. Chem. Sci. 2019, 10, 10290–10296. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; De Maziere, A.; Iaea, D.B.; Arthur, C.P.; Klumperman, J.; Ciferri, C.; Hannoush, R.N. Visualizing the cellular route of entry of a cystine-knot peptide with Xfect transfection reagent by electron microscopy. Sci. Rep. 2019, 9, 6907. [Google Scholar] [CrossRef] [PubMed]
- Etxaniz, A.; Gonzalez-Bullon, D.; Martin, C.; Ostolaza, H. Membrane Repair Mechanisms against Permeabilization by Pore-Forming Toxins. Toxins (Basel) 2018, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, P.; Brnjic, S.; Olofsson, M.H.; Fryknas, M.; Lindsten, K.; De Cesare, M.; Perego, P.; Sadeghi, B.; Hassan, M.; Larsson, R.; et al. Inhibition of proteasome deubiquitinating activity as a new cancer therapy. Nat. Med. 2011, 17, 1636–1640. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.A.; Pinto-Fernandez, A.; Cornelissen, L.; Bonham, S.; Diaz-Saez, L.; Riant, O.; Huber, K.V.M.; Kessler, B.M.; Feron, O.; Tate, E.W. Re-Evaluating the Mechanism of Action of alpha, beta-Unsaturated Carbonyl DUB Inhibitors b-AP15 and VLX1570: A Paradigmatic Example of Unspecific Protein Cross-linking with Michael Acceptor Motif-Containing Drugs. J. Med. Chem 2020, 63, 3756–3762. [Google Scholar] [CrossRef]
- Shibayama, H.; Takai, E.; Matsumura, I.; Kouno, M.; Morii, E.; Kitamura, Y.; Takeda, J.; Kanakura, Y. Identification of a cytokine-induced antiapoptotic molecule anamorsin essential for definitive hematopoiesis. J. Exp. Med. 2004, 199, 581–592. [Google Scholar] [CrossRef]
- Das, T.; Kim, E.E.; Song, E.J. Phosphorylation of USP15 and USP4 Regulates Localization and Spliceosomal Deubiquitination. J. Mol. Biol. 2019, 431, 3900–3912. [Google Scholar] [CrossRef]
- Das, D.S.; Das, A.; Ray, A.; Song, Y.; Samur, M.K.; Munshi, N.C.; Chauhan, D.; Anderson, K.C. Blockade of Deubiquitylating Enzyme USP1 Inhibits DNA Repair and Triggers Apoptosis in Multiple Myeloma Cells. Clin. Cancer Res. 2017, 23, 4280–4289. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, M.J. Isolation and Characterization of RNA Aptamers against a Proteasome-Associated Deubiquitylating Enzyme UCH37. Chembiochem 2017, 18, 171–175. [Google Scholar] [CrossRef]
- Todi, S.V.; Winborn, B.J.; Scaglione, K.M.; Blount, J.R.; Travis, S.M.; Paulson, H.L. Ubiquitination directly enhances activity of the deubiquitinating enzyme ataxin-3. EMBO J. 2009, 28, 372–382. [Google Scholar] [CrossRef]
- Yang, W.L.; Jin, G.; Li, C.F.; Jeong, Y.S.; Moten, A.; Xu, D.; Feng, Z.; Chen, W.; Cai, Z.; Darnay, B.; et al. Cycles of ubiquitination and deubiquitination critically regulate growth factor-mediated activation of Akt signaling. Sci. Signal. 2013, 6, ra3. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kong, L.; Yang, Q.; Duan, A.; Ju, X.; Cai, B.; Chen, L.; An, T.; Li, Y. Parthenolide inhibits ubiquitin-specific peptidase 7 (USP7), Wnt signaling, and colorectal cancer cell growth. J. Biol. Chem. 2020, 295, 3576–3589. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Fernández, A.; Davis, S.; Schofield, A.B.; Scott, H.C.; Zhang, P.; Salah, E.; Mathea, S.; Charles, P.D.; Damianou, A.; Bond, G.; et al. Comprehensive Landscape of Active Deubiquitinating Enzymes Profiled by Advanced Chemoproteomics. Front. Chem. 2019, 7, 592. [Google Scholar] [CrossRef] [PubMed]
- Conole, D.; Mondal, M.; Majmudar, J.D.; Tate, E.W. Recent Developments in Cell Permeable Deubiquitinating Enzyme Activity-Based Probes. Front. Chem. 2019, 7, 876. [Google Scholar] [CrossRef] [PubMed]
- Heal, W.P.; Dang, T.H.; Tate, E.W. Activity-based probes: Discovering new biology and new drug targets. Chem Soc. Rev. 2011, 40, 246–257. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Reporter Tag | Cys-Reactive Electrophilic Warhead | |||
|---|---|---|---|---|
| Affinity | Fluorescent | Direct Addition | Conjugate Addition | Nucleophilic Substitution |
| HA | Cy5 | Nitrile (Ub-CN) | Vinyl methyl ester(Ub-VME) | Chloroethylamine (Ub-Cl) |
| Biotin | TAMRA | Propargyl amide (Ub-PA/Prg) | Vinyl methyl sulfone (Ub-VS) | Bromoethylamine (Ub-Br2) |
| Flag | TER | Aldehyde (Ub-al) | Vinyl cyanide (Ub-CN) | Bromopropylamine (Ub-Br3) |
| - | - | - | Dehydroalanine (Ub-Dha) | Acyloxymethyl ketone (Ub-TF3BOK) |
| - | - | - | - | α-Amino-β-lactone (Ub-Lac) |
| Detection Method | Ubiquitin Substrates | Ref. | |
|---|---|---|---|
| in vitro deubiquitinating assay | Fluorescence | Ub-AMC | [30,31,32,33,89] |
| Ub-PLA2 | [36,37] | ||
| Ub-Rho110 | [38,39,40,90] | ||
| FP-based Ub-Lys-TAMRA-Gly | [44,45,46,53] | ||
| FRET di-Ub | [50,51,52] | ||
| Luminescence | Ub-AML | [34,35] | |
| Gel staining | Ubiquitin chain | [27,32,77] | |
| Deubiquitinating assay with mammalian cells | Western blotting | Ubiquitin chain (with immunoprecipitated DUB) | [28,29,91] |
| Nickel pull-down assay | [59,60,92] | ||
| Activity-based probes | [37,62,93] | ||
| Mass spectrometry | Activity-based probes | [37,74,94] | |
| Microscopy (fluorescence) | Activity-based probes (cell permeable only) | [70,95,96] | |
| Purpose | Deubiquitination Assays |
|---|---|
| Substrate specificity | Nickel pulldown assay |
| Ubiquitin chain linkage specificity | Ubiquitin chain cleavage assay, Nickel pulldown assay, FRET di-Ub |
| Kinetics | Ub-AMC, Ub-AML, Ub-PLA2, Ub-Rho110, FP-based Ub-Lys-TAMRA-Gly, FRET di-Ub |
| DUB activity | Ubiquitin chain cleavage assay, Nickel pulldown assay, ABP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.; Park, J.; Kim, E.E.; Song, E.J. Assay Systems for Profiling Deubiquitinating Activity. Int. J. Mol. Sci. 2020, 21, 5638. https://doi.org/10.3390/ijms21165638
Cho J, Park J, Kim EE, Song EJ. Assay Systems for Profiling Deubiquitinating Activity. International Journal of Molecular Sciences. 2020; 21(16):5638. https://doi.org/10.3390/ijms21165638
Chicago/Turabian StyleCho, Jinhong, Jinyoung Park, Eunice EunKyeong Kim, and Eun Joo Song. 2020. "Assay Systems for Profiling Deubiquitinating Activity" International Journal of Molecular Sciences 21, no. 16: 5638. https://doi.org/10.3390/ijms21165638
APA StyleCho, J., Park, J., Kim, E. E., & Song, E. J. (2020). Assay Systems for Profiling Deubiquitinating Activity. International Journal of Molecular Sciences, 21(16), 5638. https://doi.org/10.3390/ijms21165638