In Experimental Dilated Cardiomyopathy Heart Failure and Survival Are Adversely Affected by a Lack of Sexual Interactions

 ,
,  and
and
Abstract
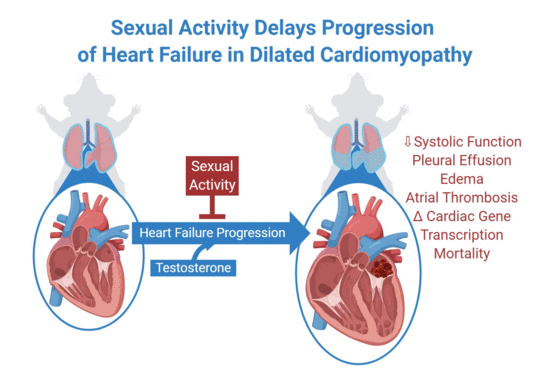
1. Introduction
2. Results and Discussion
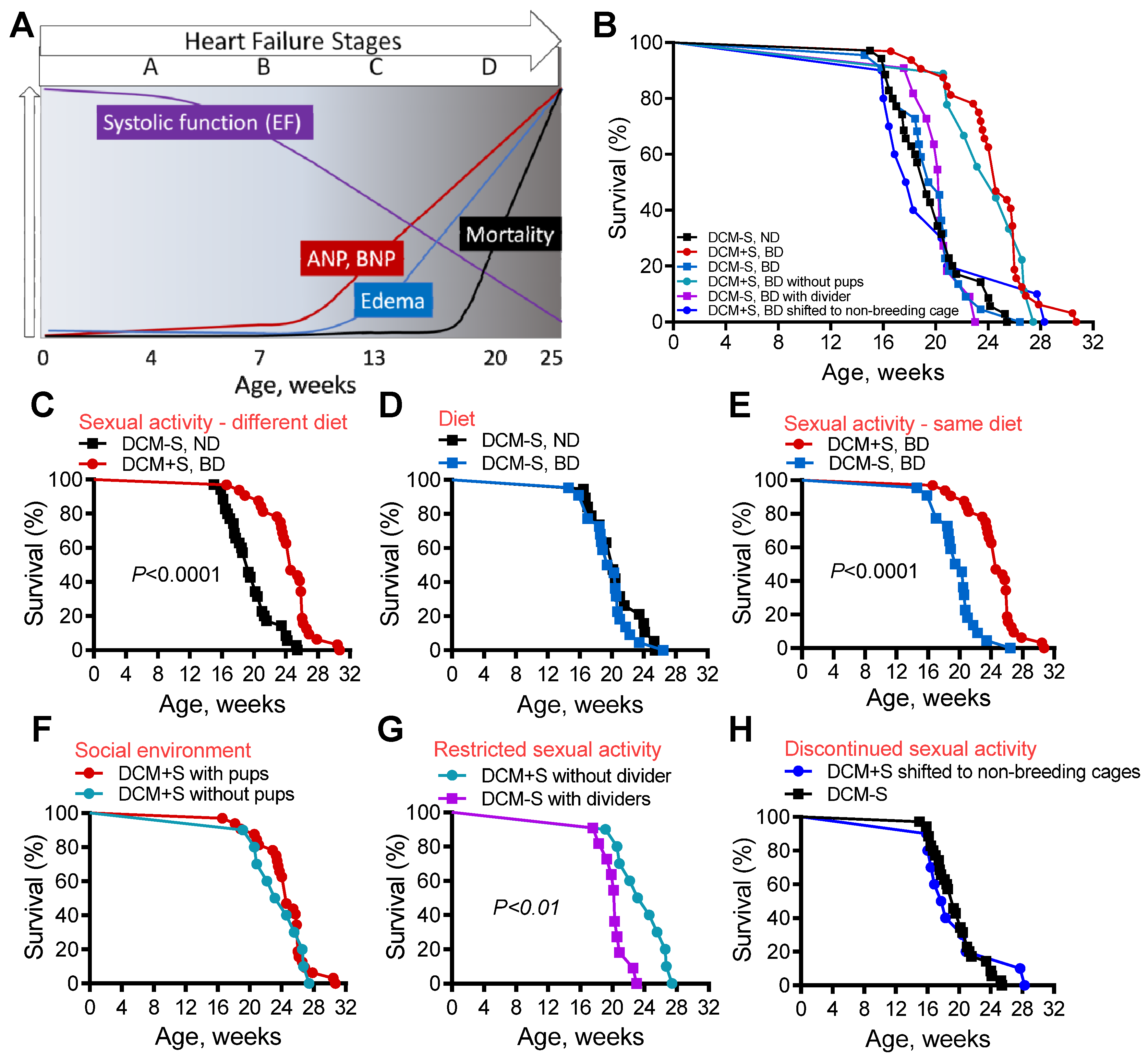
2.1. Access to Sexual Activity Improves Survival in Mice with Dilated Cardiomyopathy (DCM)
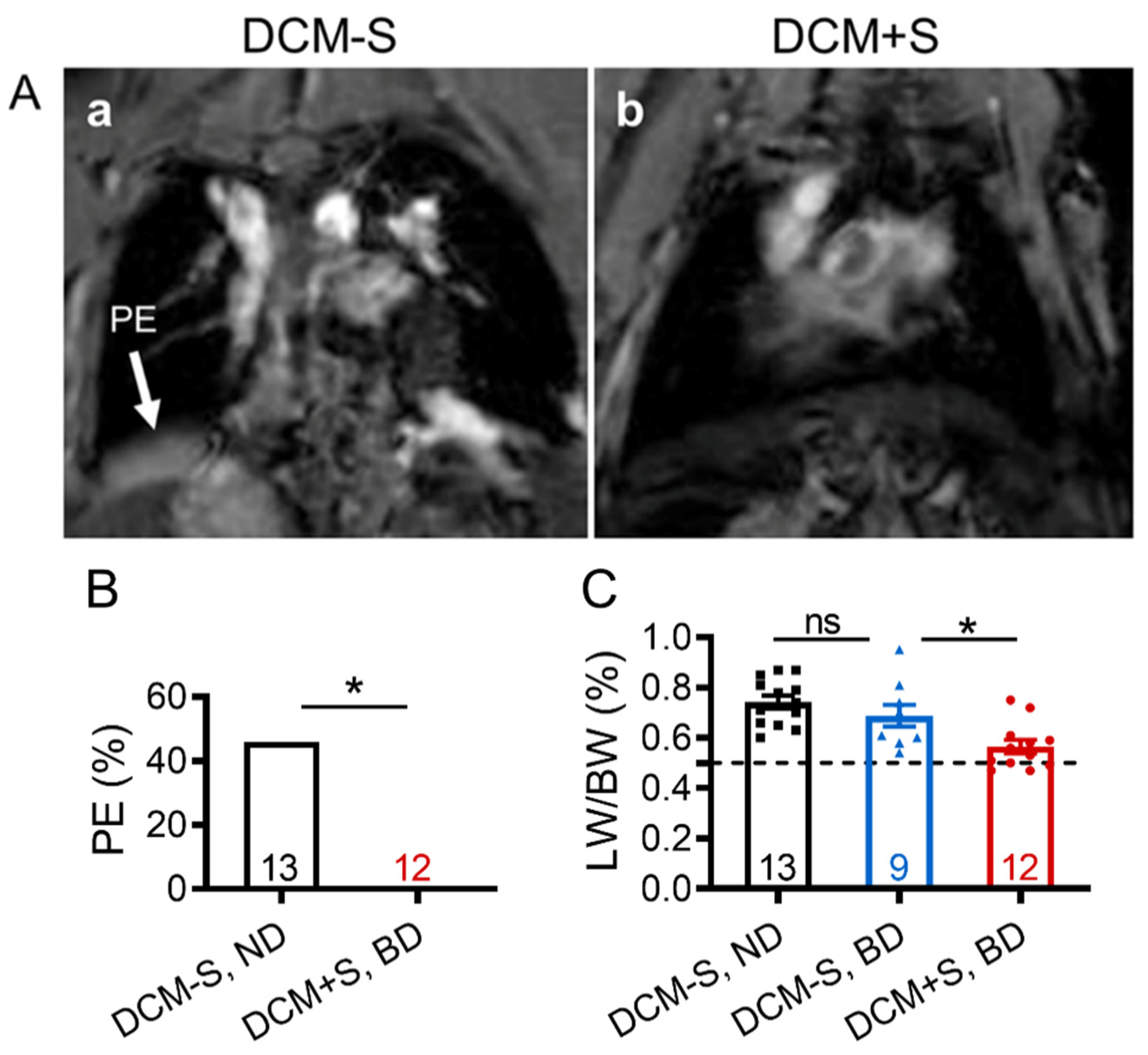
2.2. Sexual Interaction Reduces Pleural Effusion and Edema Development
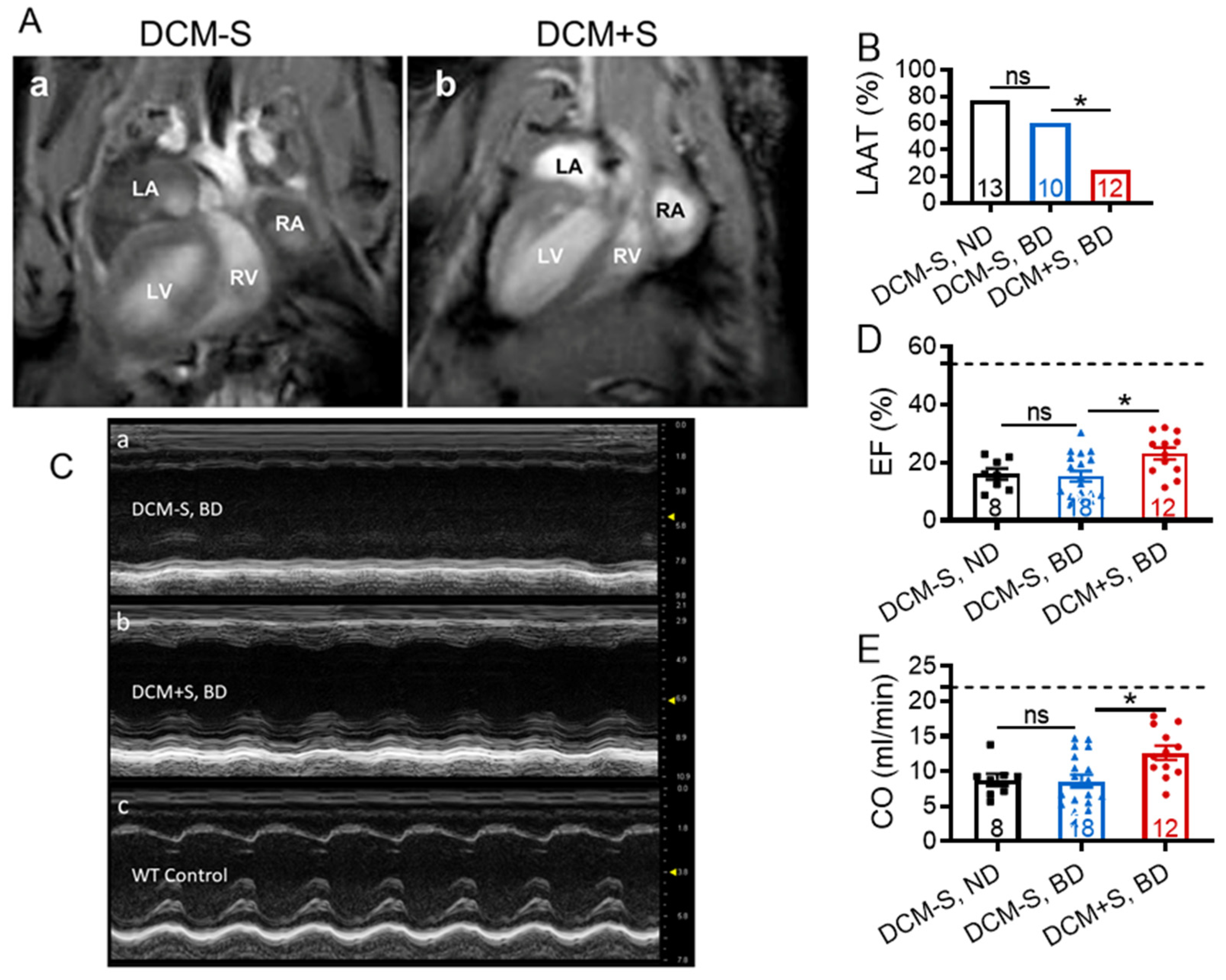
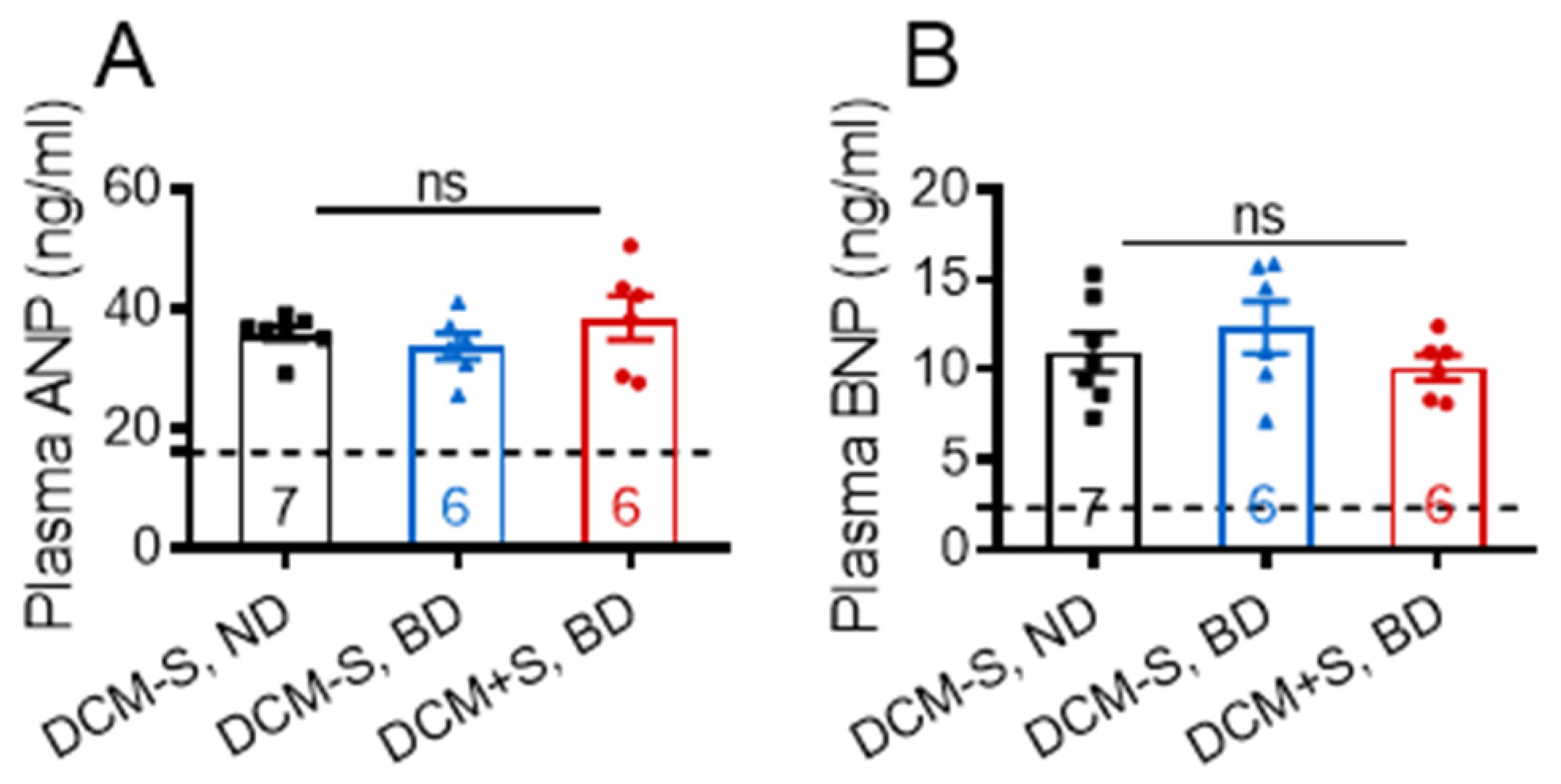
2.3. Sexual Interaction Improves Myocardial Contractile Function
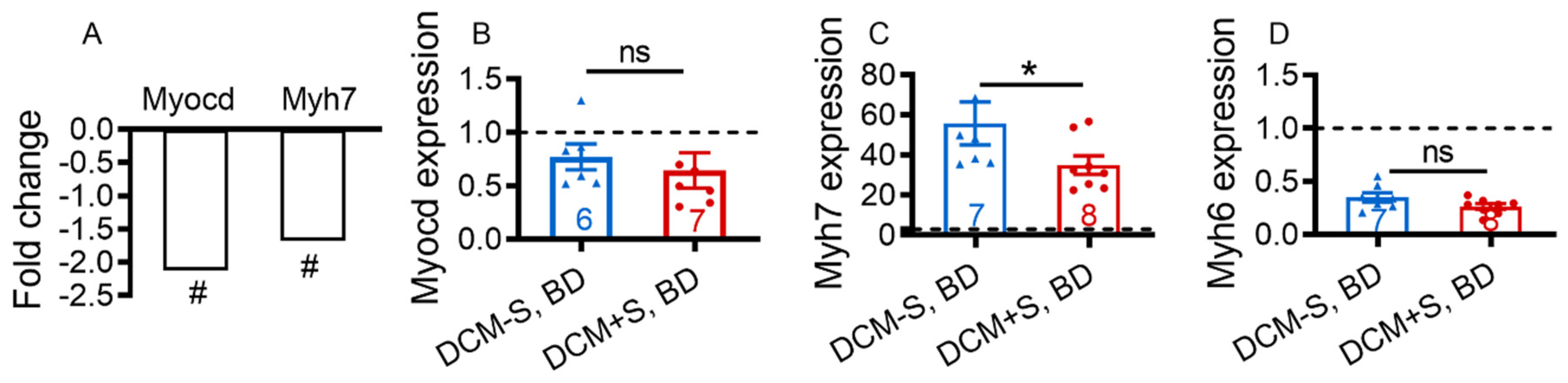
2.4. Sexual Activity Attenuates Heart Failure through Differentiall Activation of Cardiac Contractile Transcription Pathways
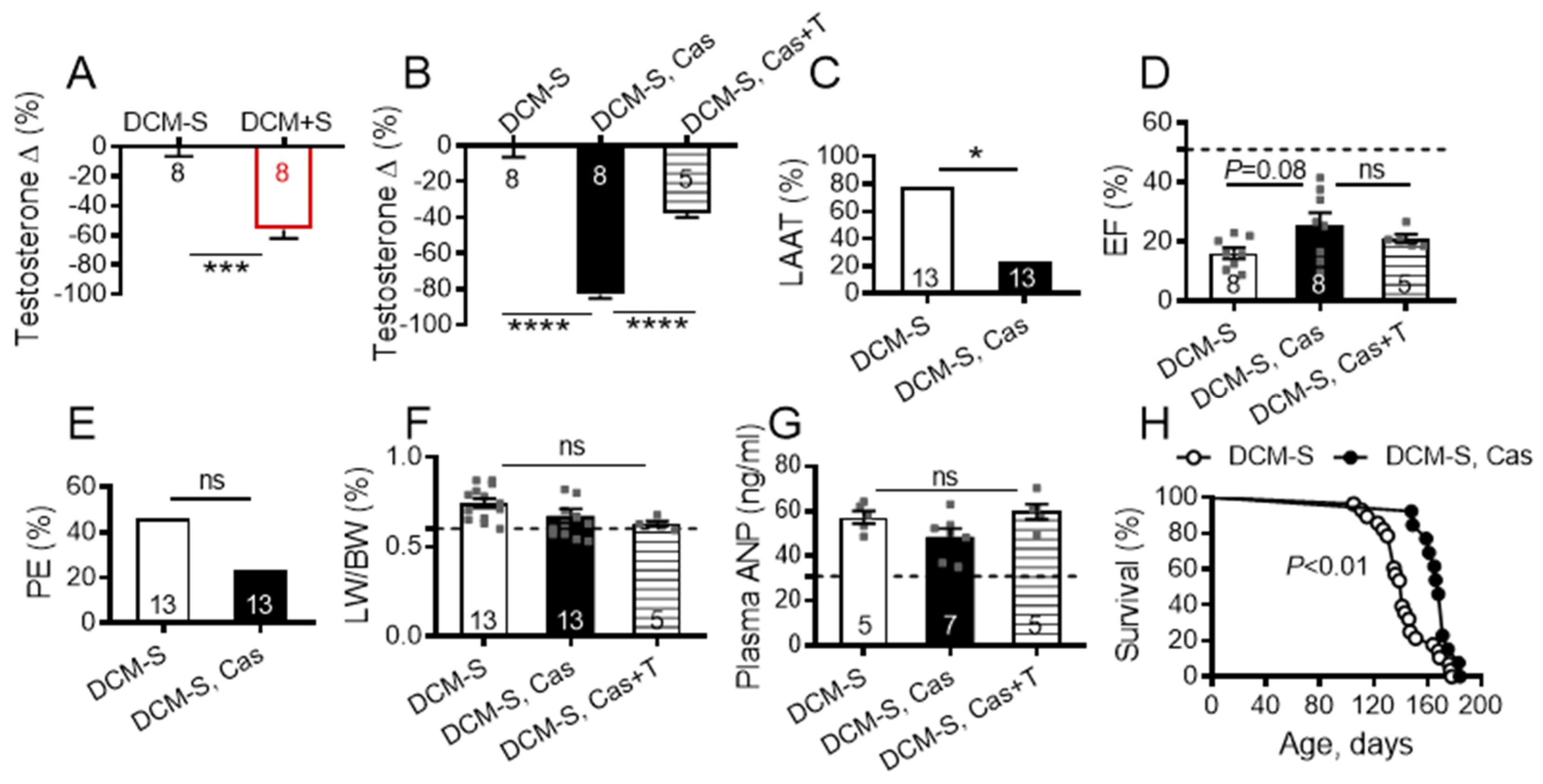
2.5. Sexual Activity and Lowered Testosterone, Attenuate HF and Prolong Survival
3. Materials and Methods
3.1. Animals
3.2. Echocardiography
3.3. Magnetic Resonance Imaging (MRI)
3.4. Left Atrial Appendage Thrombus, Lung Edema and Pleural Effusion Analysis
3.5. Surgical Castration
3.6. Testosterone Pellet Implantation
3.7. RNA Isolation and Microarray Analysis
3.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
3.9. Enzyme Immunoassay
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTC1 | α-actin-1 |
| ACTC2 | α-actinin-2 |
| ANP | atrial natriuretic peptide |
| BD | breeding diet |
| BNP | b-type natriuretic peptide |
| CO | cardiac output |
| Cas | castrated |
| Cas+T | castrated with supplemented testosterone |
| DES | desmin |
| DVR | digital video recorders |
| DCM | dilated cardiomyopathy |
| DCM+S | dilated cardiomyopathy with access to sexual activity |
| DCM-S | dilated cardiomyopathy without access to sexual activity |
| DMD | dystrophin |
| EF | ejection fraction |
| ELISA | enzyme-linked immunosorbent assay |
| FDR | false discovery rate |
| FC | fold-change |
| FS | fractional shortening |
| HF | heart failure |
| LAAT | left atrial appendage thrombus |
| LA | left atrium |
| LV | left ventricle |
| LW/BW | lung weight-to-body weight ratio |
| MRI | magnetic resonance imaging |
| Myocd | myocardin |
| Myh6 | myosin heavy chain 6 |
| Myh7 | myosin heavy chain 7 |
| NP | natriuretic peptide |
| ND | normal diet |
| PE | pleural effusion |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| RA | right atrium |
| RV | right ventricle |
| T | testosterone |
| TPM1 | tropomyosin-1 |
References
- Jessup, M.; Brozena, S. Heart failure. N. Engl. J. Med. 2003, 348, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Metra, M.; Teerlink, J.R. Heart failure. Lancet 2017, 390, 1981–1995. [Google Scholar] [CrossRef]
- Weintraub, R.G.; Semsarian, C.; Macdonald, P. Dilated cardiomyopathy. Lancet 2017, 390, 400–414. [Google Scholar] [CrossRef]
- Lindau, S.T.; Schumm, L.P.; Laumann, E.O.; Levinson, W.; O’Muircheartaigh, C.A.; Waite, L.J. A Study of Sexuality and Health among Older Adults in the United States. N. Engl. J. Med. 2007, 357, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Levine, G.N.; Steinke, E.E.; Bakaeen, F.G.; Bozkurt, B.; Cheitlin, M.D.; Conti, J.B.; Foster, E.; Jaarsma, T.; Kloner, R.A.; Lange, R.A.; et al. Sexual Activity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 2012, 125, 1058–1072. [Google Scholar] [CrossRef]
- Baert, A.; Pardaens, S.; De Smedt, D.; Puddu, P.E.; Ciancarelli, M.C.; Dawodu, A.; De Sutter, J.; De Bacquer, D.; Clays, E. Sexual Activity in Heart Failure Patients: Information Needs and Association with Health-Related Quality of Life. Int. J. Environ. Res. Public Heal. 2019, 16, 1570. [Google Scholar] [CrossRef]
- Mandras, S.; Uber, P.A.; Mehra, M.R. Sexual Activity and Chronic Heart Failure. Mayo Clin. Proc. 2007, 82, 1203–1210. [Google Scholar] [CrossRef]
- Jaarsma, T. Reply to ‘Sexual function of patients with heart failure: Distinct phenotypes distinct sexual function?’ by Konstantinos Koutsampasopoulos. ESC Hear. Fail. 2017, 4, 320. [Google Scholar] [CrossRef][Green Version]
- Jaarsma, T.; Fridlund, B.; Mårtensson, J. Sexual Dysfunction in Heart Failure Patients. Curr. Hear. Fail. Rep. 2014, 11, 330–336. [Google Scholar] [CrossRef]
- Kolbe, N.; Kugler, C.; Schnepp, W.; Jaarsma, T. Sexual Counseling in Patients with Heart Failure. J. Cardiovasc. Nurs. 2016, 31, 53–61. [Google Scholar] [CrossRef]
- Fentzke, R.C.; Korcarz, C.E.; Lang, R.M.; Lin, H.; Leiden, J.M. Dilated cardiomyopathy in transgenic mice expressing a dominant-negative CREB transcription factor in the heart. J. Clin. Investig. 1998, 101, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.T.; Collins, K.; Korcarz, C.E.; Fentzke, R.; Lang, R.M.; Leiden, J.M. Effects of exercise training on LV performance and mortality in a murine model of dilated cardiomyopathy. Am. J. Physiol. Circ. Physiol. 2000, 279, H210–H215. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.A.; Birdsey, N.; Huggins, G.S.; Svensson, E.; Heppe, D.; Knaub, L. Cardiac-specific overexpression of dominant-negative CREB leads to increased mortality and mitochondrial dysfunction in female mice. Am. J. Physiol. Circ. Physiol. 2010, 299, H2056–H2068. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, I.P.; Wang, N.; McNamee, R.; Houng, A.K.; A Mohamad, A.; Fan, T.M.; Reed, G. Corin overexpression improves cardiac function, heart failure, and survival in mice with dilated cardiomyopathy. Hypertension 2012, 61, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Gladysheva, I.P.; Fan, T.-H.M.; Sullivan, R.; Houng, A.K.; Reed, G. Atrial natriuretic peptide affects cardiac remodeling, function, heart failure, and survival in a mouse model of dilated cardiomyopathy. Hypertension 2013, 63, 514–519. [Google Scholar] [CrossRef]
- Tripathi, R.; Wang, D.; Sullivan, R.; Fan, T.-H.M.; Gladysheva, I.P.; Reed, G.L. Depressed Corin Levels Indicate Early Systolic Dysfunction Before Increases of Atrial Natriuretic Peptide/B-Type Natriuretic Peptide and Heart Failure Development. Hypertension 2016, 67, 362–367. [Google Scholar] [CrossRef]
- Tripathi, R.; Sullivan, R.; Fan, T.-H.M.; Wang, D.; Sun, Y.; Reed, G.L.; Gladysheva, I.P. Enhanced heart failure, mortality and renin activation in female mice with experimental dilated cardiomyopathy. PLoS ONE 2017, 12, e0189315. [Google Scholar] [CrossRef]
- Sullivan, R.; Mehta, R.M.; Tripathi, R.; Gladysheva, I.P.; Reed, G.L. Normalizing Plasma Renin Activity in Experimental Dilated Cardiomyopathy: Effects on Edema, Cachexia, and Survival. Int. J. Mol. Sci. 2019, 20, 3886. [Google Scholar] [CrossRef]
- Tripathi, R.; Sullivan, R.; Fan, T.-H.M.; Houng, A.K.; Mehta, R.M.; Reed, G.L.; Gladysheva, I.P. Cardiac-Specific Overexpression of Catalytically Inactive Corin Reduces Edema, Contractile Dysfunction, and Death in Mice with Dilated Cardiomyopathy. Int. J. Mol. Sci. 2019, 21, 203. [Google Scholar] [CrossRef]
- Merlo, M.; Cannata, A.; Gobbo, M.; Stolfo, D.; Elliott, P.M.; Sinagra, G. Evolving concepts in dilated cardiomyopathy. Eur. J. Hear. Fail. 2017, 20, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Hershberger, R.E.; Hedges, D.J.; Morales, A. Dilated cardiomyopathy: The complexity of a diverse genetic architecture. Nat. Rev. Cardiol. 2013, 10, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E.; Jr Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA guideline for the management of heart failure: Executive summary: A report of the American College of Cardiology Foundation/American Heart Association Task Force on practice guidelines. Circulation 2013, 128, 1810–1852. [Google Scholar] [CrossRef] [PubMed]
- Houser, S.R.; Margulies, K.B.; Murphy, A.M.; Spinale, F.G.; Francis, G.S.; Prabhu, S.D.; Rockman, H.A.; Kass, D.A.; Molkentin, J.D.; A Sussman, M.; et al. Animal Models of Heart Failure. Circ. Res. 2012, 111, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Tallent, B.R.; Lifshitz, J. Creating an Inexpensive Camera Array for Rodent Observation; Laboratory Animal Science Professional: Memphis, TN, USA, June 2018; pp. 47–50. [Google Scholar]
- Grewal, J.; Siu, S.C.; Ross, H.J.; Mason, J.; Balint, O.H.; Sermer, M.; Colman, J.M.; Silversides, C.K. Pregnancy Outcomes in Women With Dilated Cardiomyopathy. J. Am. Coll. Cardiol. 2009, 55, 45–52. [Google Scholar] [CrossRef]
- Lompre, A.-M.; Schwartz, K.; D’Albis, A.; Lacombe, G.; Van Thiem, N.; Swynghedauw, B. Myosin isoenzyme redistribution in chronic heart overload. Nature 1979, 282, 105–107. [Google Scholar] [CrossRef]
- Edwards, B.S.; Ackermann, D.M.; Lee, M.E.; Reeder, G.S.; Wold, L.E.; Burnett, J.C., Jr. Identification of atrial natriuretic factor within ventricular tissue in hamsters and humans with congestive heart failure. J. Clin. Investig. 1988, 81, 82–86. [Google Scholar] [CrossRef]
- Razeghi, P.; Young, M.E.; Alcorn, J.L.; Moravec, C.S.; Frazier, O.; Taegtmeyer, H. Metabolic gene expression in fetal and failing human heart. Circulation 2001, 104, 2923–2931. [Google Scholar] [CrossRef]
- Rajabi, M.; Kassiotis, C.; Razeghi, P.; Taegtmeyer, H. Return to the fetal gene program protects the stressed heart: A strong hypothesis. Hear. Fail. Rev. 2007, 12, 331–343. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Chang, P.S.; Wang, Z.; Sutherland, L.; Richardson, J.A.; Small, E.; Krieg, P.A.; Olson, E.N. Activation of Cardiac Gene Expression by Myocardin, a Transcriptional Cofactor for Serum Response Factor. Cell 2001, 105, 851–862. [Google Scholar] [CrossRef]
- Mikhailov, A.T.; Torrado, M. In Search of Novel Targets for Heart Disease: Myocardin and Myocardin-Related Transcriptional Cofactors. Biochem. Res. Int. 2012, 2012, 973723. [Google Scholar] [CrossRef]
- Holmboe, S.A.; Priskorn, L.; Jørgensen, N.; Skakkebaek, N.E.; Linneberg, A.; Juul, A.; Andersson, A.-M. Influence of marital status on testosterone levels—A ten year follow-up of 1113 men. Psychoneuroendocrinology 2017, 80, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Alvergne, A.; Faurie, C.; Raymond, M. Variation in testosterone levels and male reproductive effort: Insight from a polygynous human population. Horm. Behav. 2009, 56, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Basaria, S.; Coviello, A.D.; Travison, T.G.; Storer, T.W.; Farwell, W.R.; Jette, A.M.; Eder, R.; Tennstedt, S.; Ulloor, J.; Zhang, A.; et al. Adverse Events Associated with Testosterone Administration. N. Engl. J. Med. 2010, 363, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Freeman, G.; Cowling, B.J.; Schooling, M. Testosterone therapy and cardiovascular events among men: A systematic review and meta-analysis of placebo-controlled randomized trials. BMC Med. 2013, 11, 108. [Google Scholar] [CrossRef]
- Kloner, R.A.; Carson, C.; Dobs, A.; Kopecky, S.; Mohler, E.R. Testosterone and Cardiovascular Disease. J. Am. Coll. Cardiol. 2016, 67, 545–557. [Google Scholar] [CrossRef]
- Cavasin, M.A.; Sankey, S.S.; Yu, A.-L.; Menon, S.; Yang, X.-P. Estrogen and testosterone have opposing effects on chronic cardiac remodeling and function in mice with myocardial infarction. Am. J. Physiol. Circ. Physiol. 2003, 284, H1560–H1569. [Google Scholar] [CrossRef]
- Gao, X.M.; Agrotis, A.; Autelitano, D.J.; Percy, E.; Woodcock, E.A.; Jennings, G.L.; Dart, A.M.; Du, X.J. Sex hormones and cardiomyopathic phenotype induced by cardiac beta 2-adrenergic receptor overexpression. Endocrinology 2003, 144, 4097–4105. [Google Scholar] [CrossRef]
- Bocchi, E.A.; Carvalho, V.O.; Guimaraes, G.V. Inverse correlation between testosterone and ventricle ejection fraction, hemodynamics and exercise capacity in heart failure patients with erectile dysfunction. Int. Braz. J. Urol. 2008, 34, 302–312. [Google Scholar] [CrossRef][Green Version]
- Baggish, A.L.; Weiner, R.B.; Kanayama, G.; Hudson, J.I.; Picard, M.H.; Hutter, A.M.; Pope, H.G. Long-Term Anabolic-Androgenic Steroid Use Is Associated With Left Ventricular Dysfunction. Circ. Hear. Fail. 2010, 3, 472–476. [Google Scholar] [CrossRef]
- Huang, C.-K.; Lee, S.O.; Chang, E.; Pang, H.; Chang, C. Androgen receptor (AR) in cardiovascular diseases. J. Endocrinol. 2016, 229, R1–R16. [Google Scholar] [CrossRef]
- Junior, R.R.; Ronconi, K.; Jesus, I.; Almeida, P.; Forechi, L.; Vassallo, D.V.; Guatimosim, S.; Stefanon, I.; Fernandes, A. Testosterone deficiency prevents left ventricular contractility dysfunction after myocardial infarction. Mol. Cell. Endocrinol. 2018, 460, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Kattih, B.; Elling, L.S.; Weiss, C.; Bea, M.; Zwadlo, C.; Bavendiek, U.; Bauersachs, J.; Heineke, J. Anti-androgenic therapy with finasteride in patients with chronic heart failure—A retrospective propensity score based analysis. Sci. Rep. 2019, 9, 10139. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Lin, M.-T.; Cheng, R.-T.; Chen, S.-H. Testosterone Depletion by Castration May Protect Mice from Heat-Induced Multiple Organ Damage and Lethality. J. Biomed. Biotechnol. 2010, 2010, 485306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Moore, D.J.; Kim, J.I.; Lee, K.M.; O’Connor, M.R.; Duff, P.E.; Yang, M.; Lei, J.; Markmann, J.F.; Deng, S. Inhibition of Transplantation Tolerance by Immune Senescence Is Reversed by Endocrine Modulation. Sci. Transl. Med. 2011, 3, 87ra52. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.B.; Mestler, G.E. Mortality and Survival: Comparison of Eunuchs with Intact Men and Women in a Mentally Retarded Population. J. Gerontol. 1969, 24, 395–411. [Google Scholar] [CrossRef]
- Min, K.-J.; Lee, C.-K.; Park, H.-N. The lifespan of Korean eunuchs. Curr. Boil. 2012, 22, R792–R793. [Google Scholar] [CrossRef]
- Gems, D. Evolution of sexually dimorphic longevity in humans. Aging 2014, 6, 84–91. [Google Scholar] [CrossRef]
- Brooks, R.C.; Garratt, M.G. Life history evolution, reproduction, and the origins of sex-dependent aging and longevity. Ann. N. Y. Acad. Sci. 2016, 1389, 92–107. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group: | WT, ND | DCM-S, ND | DCM-S, BD | DCM+S, BD | 1p-Value |
|---|---|---|---|---|---|
| Group Size (n): | 13 | 8 | 18 | 12 | |
| Age (days): | 137 ± 1 | 139 ± 1 | 139 ± 0.5 | 137 ± 1 | 0.31 |
| Body Weight (g): | 29 ± 0.7 | 29 ± 0.6 | 29 ± 0.5 | 30 ± 0.6 | 0.20 |
| Heart Rate (bpm): | 479 ± 12 | 420 ± 21 | 440 ± 22 | 480 ± 20 | 0.11 |
| Ejection Fraction (%): | 54 ± 2 | 16 ± 2 | 15 ± 2 | 23 ± 2 | 0.0001 |
| Fractional Shortening (%): | 28 ± 1 | 7 ± 1 | 7 ± 1 | 11 ± 1 | 0.0001 |
| Cardiac Output (mL/min): | 22 ± 1 | 9 ± 1 | 9 ± 1 | 13 ± 1 | 0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, R.; Sullivan, R.D.; Fan, T.-H.M.; Mehta, R.M.; Gladysheva, I.P.; Reed, G.L. In Experimental Dilated Cardiomyopathy Heart Failure and Survival Are Adversely Affected by a Lack of Sexual Interactions. Int. J. Mol. Sci. 2020, 21, 5450. https://doi.org/10.3390/ijms21155450
Tripathi R, Sullivan RD, Fan T-HM, Mehta RM, Gladysheva IP, Reed GL. In Experimental Dilated Cardiomyopathy Heart Failure and Survival Are Adversely Affected by a Lack of Sexual Interactions. International Journal of Molecular Sciences. 2020; 21(15):5450. https://doi.org/10.3390/ijms21155450
Chicago/Turabian StyleTripathi, Ranjana, Ryan D. Sullivan, Tai-Hwang M. Fan, Radhika M. Mehta, Inna P. Gladysheva, and Guy L. Reed. 2020. "In Experimental Dilated Cardiomyopathy Heart Failure and Survival Are Adversely Affected by a Lack of Sexual Interactions" International Journal of Molecular Sciences 21, no. 15: 5450. https://doi.org/10.3390/ijms21155450
APA StyleTripathi, R., Sullivan, R. D., Fan, T.-H. M., Mehta, R. M., Gladysheva, I. P., & Reed, G. L. (2020). In Experimental Dilated Cardiomyopathy Heart Failure and Survival Are Adversely Affected by a Lack of Sexual Interactions. International Journal of Molecular Sciences, 21(15), 5450. https://doi.org/10.3390/ijms21155450