The Potential of Metabolomics in the Diagnosis of Thyroid Cancer

 ,
,  ,
,  and
and 
Abstract

1. Introduction
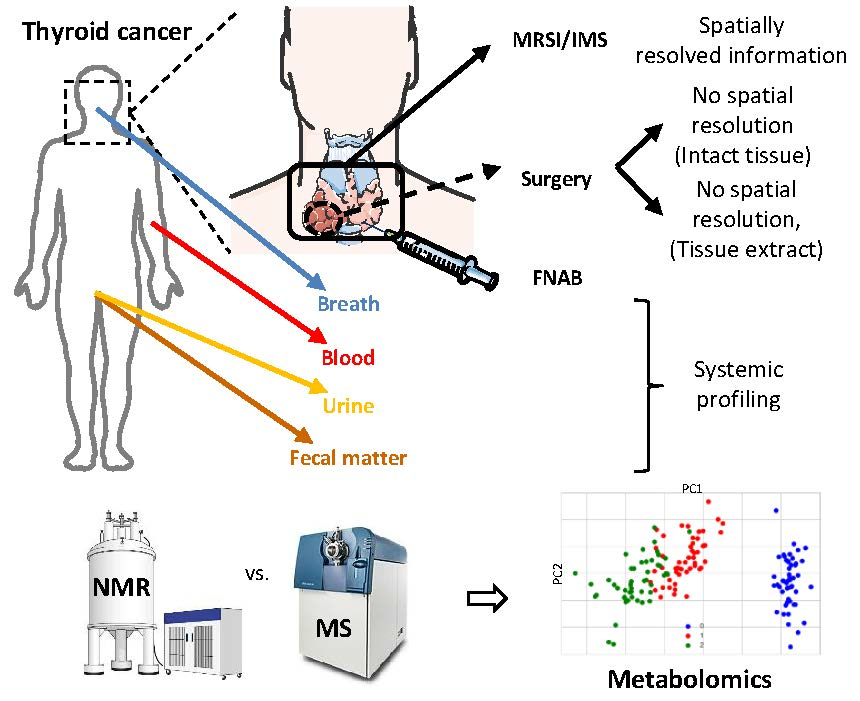
2. Metabolomics in Thyroid Cancer
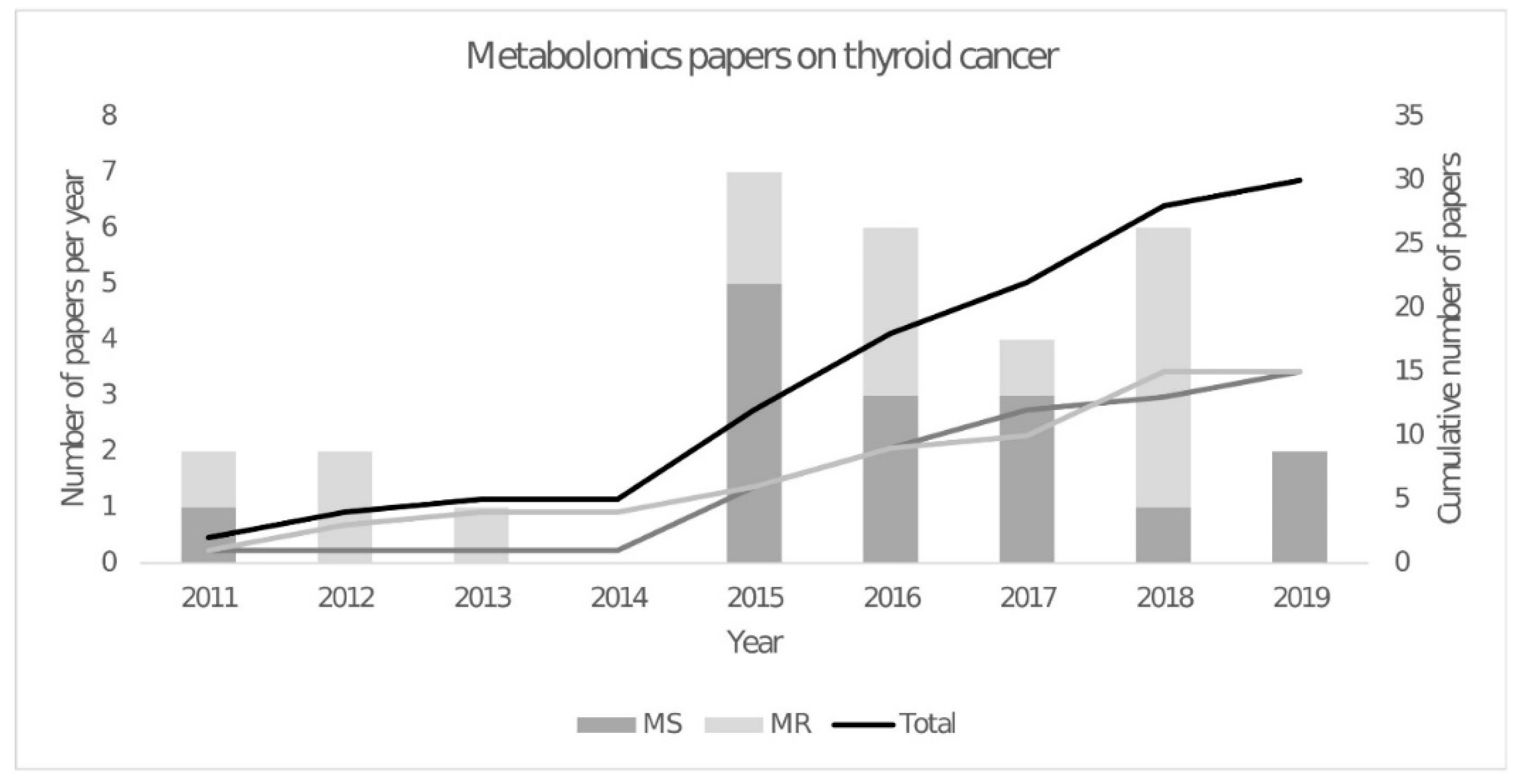
2.1. The Early Years—NMR Spectroscopy
2.2. The Rise of Mass Spectrometry
2.3. Peripheral Fluids
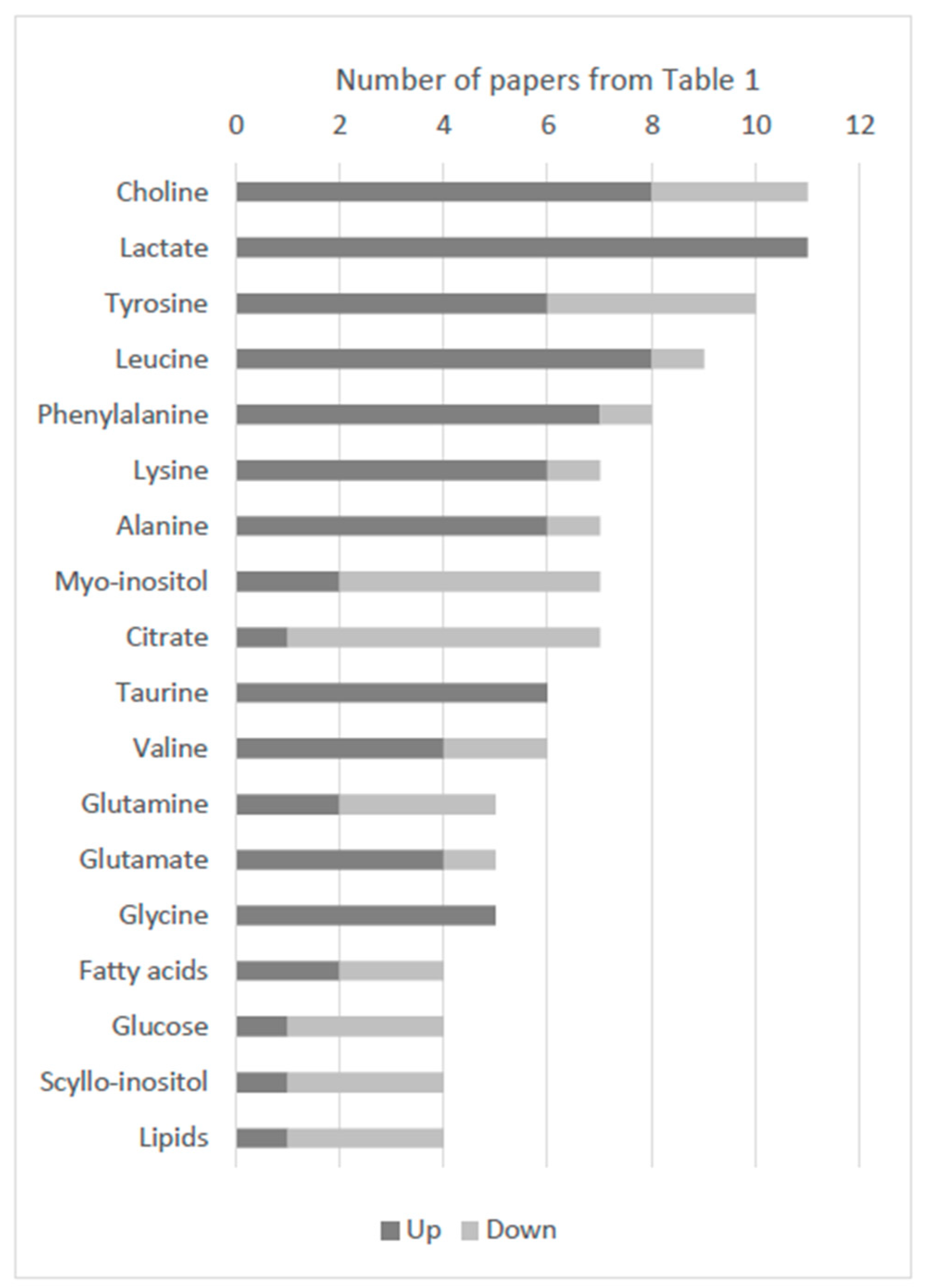
2.4. Most Referred Metabolites

{kind=link}
{kind=link}
{kind=link}
| Technique | Method | Study Site | Sample | Study Design | Altered Metabolites | Reference |
|---|---|---|---|---|---|---|
| NMR | MRS | Spatially resolved information | Non-invasive method | 8 TC vs. 5 CTR | Ch ↑ | [46] |
| MRS | Non-invasive method | 8 MN vs. 17 BN | Ch ↑ | [47] | ||
| MRS | Non-invasive method | 8 PTC vs. 18 BN | Ch ↑ | [48] | ||
| 1H-NMR | Tissue | Tissue | 19 MN vs. 24 BN | TGL, K ↑ | [44] | |
| 2D 1H-NMR | Tissue | 32 MN vs. 61 BN | Cross peaks from CHL/cholesteryl esters and di-/TGL ↑; two unassigned cross peaks ↓ | [45] | ||
| HR-MAS-NMR | Tissue | Tissue: 72 TC vs. 28 CTR; Tissue: 38 MN vs. 34 BN; Aspirate ex vivo: 12 TC vs. 12 CTR | Tissue TC vs. CTR: F, Y, S, K, TAU, Q, E, A, I, L and V ↑; Lip ↓. Tissue MN vs. BN: LAC and TAU ↑; Lip, Ch, PC, myo- and scyllo-IST ↓ | [53] | ||
| HR-MAS-NMR | Tissue | 38 MN vs. 34 BN | F, TAU and LAC ↑; Ch and Ch derivatives, myo- and scyllo-IST ↓ | [54] | ||
| HR-MAS-NMR | Tissue | 52 MN vs. 46 BN | Y, S, A, L, F↑; myo- and scyllo-IST and CIT ↓ | [57] | ||
| 1H-NMR | Tissue extracts | Tissue extracts | 15 TC vs. 19 BN and 27 CTR | CHL ↑; DLC ↓ | [49] | |
| 1H-NMR | Tissue extracts | 45 thyroid lesions vs. 19 CTR from the same participant | M, A, E, G, LAC, Y, F and HPX ↑; ACT ↓ | [58] | ||
| 1H-NMR | Tissue extracts | 32 LNM vs. 20 absence of LNM; 19 lateral LNM vs. 33 absence of lateral LNM | No statistically altered metabolites | [61] | ||
| 1H-NMR | Tissue extracts | 16 PTC vs. 16 CTR from the same participant | L, V, G, TAU, LAC, Ch, ETA, GPC and LDL↑; CIT, VLDL ↓ | [62] | ||
| 1H-NMR | Tissue extracts | 11 TC vs. 10 CTR from the same participant | LAC, F ↑ | [63] | ||
| 1H-NMR | FNAB | Aspirates | 34 PTC vs. 69 BN | LAC, Ch, O-PC, G↑; CIT, E, Q ↓ | [60] | |
| 31P-NMR | Systemic profiling | Plasma | 16 MN vs. 17 hypothyroid in remission and 14 euthyroidism in remission and 23 healthy euthyroid controls | MN vs. hypothyroid in remission: PE + SM and PC ↓ | [50] | |
| 1H-NMR | Serum | 20 PTC vs. 20 BN and 20 CTR | PTC vs. CTR: V, L, I, LACA, A, E, K, G ↑; Lip, Ch and Y ↓ | [84] | ||
| 1H-NMR | Serum | 17 PTC vs. 17 BN and 20 CTR | PTC vs. BN: KYN, HIP, NIC, XNT ↑; Q, CIT, O-ALC, GSH, W, Y, HoS, β-A ↓ PTC vs. CTR: myo- and scyllo-IST, W, PPN, LAC, HoC, 3-Me GTA, N, D, Ch ↑; ACM ↓ | [85] | ||
| 1H-NMR | Serum | 41 PTC vs. 55 BN and 40 CTR | L, LAC, A, G, K and Ch ↑; GLU ↓ | [86] | ||
| 1H-NMR | Serum and urine | 17 PTC vs. 33 BN and 17 CTR | PTC vs. CTR: Serum: CRE ↑; V, A, CRN and Y ↓; Urine: CIT and ACT ↓ | [88] | ||
| HR-MAS-NMR | Combination | Tissue and Aspirates | 4 PTC, 4 FA, 5 CTR | NA | [51] | |
| HR-MAS-NMR and 1H-NMR | Tissue and plasma | Tissue: 16 PTMC vs. 11 CTR tissues from the same participants; Plasma: 26 PTMC vs. 17 CTR volunteers | Tissue: F, Y, LAC, S, C, K, Q/E, TAU, L, A, I and V ↑; FA ↓. Plasma: same as tissue as well as GLU, MAN, PYR and 3-HBA ↑ and V, Y, P, K, L ↓ | [56] | ||
| MS | IMS and MS/MS | Spatially resolved information | Tissue | 7 PTC vs. 7 CTR from the same participants | PC (16:0/18:1) and (16:0/18:2) and SM (d18:0/16:1) ↑ | [71] |
| IMS and MALDI-FTIR MS | Tissue and serum | Tissue: 16 MN vs. 5 BN and 15 CTR Serum: 124 MN vs. 43 BN and 122 CTR | MN vs. BN: PA (36:2), (36:3), (38:3) ↑ PA (38:4), (38:5), (40:5) ↓ | [72] | ||
| DESI-MS | Tissue | Tissue | 8 PTC vs. 18 CTR lymph nodes from the same participant | Q in adjacent lymph node, GSH, CDL, PI, PS and CER↑ | [73] | |
| AFADESI-IMS | Tissue | 12 PTC vs. 12 CTR from the same participant | F, L, Y ↑; CRE ↓ | [74] | ||
| GC-MS | Tissue extracts | Tissue extracts | 16 PTC vs. 16 CTR from the same participant | MLO, IN, CHL and ARA altered; GLU, FRU, GAL, MAN, 2-keto-D-GLA and RHA ↓ | [76] | |
| GC-TOF-MS and UHPLC-qTOF-MS | Tissue extracts | 57 PTC vs. CTR from the same participant; 48 BN vs. CTR from the same participant | LACA, TCA cycle intermediates, Aa, one-carbon metabolism ↑, disrupted W metabolism in PTC and BN. TAU and HTAU and ECDA ↑ in only PTC | [109] | ||
| GC-TOF-MS and UHPLC-QqQ-MS | Tissue extracts | Untargeted: 15 PTC vs. 15 CTR from the same participants; Targeted: 10 PTC vs. 10 CTR from the same participants | GOL, MLB and MEL ↓ | [77] | ||
| GC-MS | Formalin-fixed tissue | 7 FTC, 4 PTC, 4 PTC-FV, 6 MTC, 6 ATC, 3 FA, 5 CTR | Cancerous thyroid vs. normal tissue: LACA ↑; several FA and their esters ↓. MN vs. BN: myo-IST Ph, SCA and certain FA and their esters ↑; PTC vs. follicular thyroid lesions: CTA ↑; GLA ↓ | [78] | ||
| MALDI-Q-Ion Mobility-TOF-MS | Formalin-fixed tissue sections | 3 PTC vs. 3 BN from the same participant | PC (32:0), (32:1), (34:1) and (36:3), SM (34:1) and (36:1) and PA (36:2) and (36:3) ↑ | [75] | ||
| GC-MS | Systemic profiling | Exhaled breath | 39 PTC vs. 25 BN and 32 CTR | PTC vs. BN: 1, 1, 3-triMe-3-(2-Me-2-propenyl) CPT, trans-2-dodecen-1-ol ↑; (3-Me-oxiran-2-yl)-methanol ↓; PTC vs. CTR: PHN, ETG mono vinyl ester, CPR, 1-bromo-1-(3-Me-1-pentenylidene)-2,2,3,3-tetraMe CPR ↑; CHX, 4-HBA, 2,2-dimethyldecane, ETH ↓ | [93] | |
| GC-MS | Plasma | 19 PTC vs. 16 BN and 20 CTR | PTC vs. BN: SUC ↑; PTC vs. CTR: E, α-KTG, AD-5 monoPh, 3-HBA, CPA, URA ↑; CYS, C↓ | [83] | ||
| nUHPLC-ESI-MS/MS | Plasma | 10 TC vs. 74 other cancers and 20 CTR | TC vs. other cancers and CTR: Lyso PI (18:0) and (18:1) | [91] | ||
| LC-LTQ Orbitrap MS | Serum | 30 PTC vs. 80 BN and 30 CTR | FA, AC, SPG (SPG, SPG-1-Ph), OLM and 3-HBA ↑ | [52] | ||
| GC-TOF-MS | Serum | 37 PTC-DM vs. 40 PTC-AB | N, GABA, AOA, 4- DOP ↑; PGA ↓ | [79] | ||
| LC-DIA-MS | Serum | 30 PTC vs. 27 CTR | 392 significantly changed metabolites | [68] | ||
| UPLC-QTOF-MS | Fecal matter | 15 TC vs. 15 CTR | 3,7,11,15-tetraMe-6,10,14-hexadecatrien-1-ol, TGL (16:0/16:1(9Z)/18:2(9Z, 12Z)), 10-propyl-5,9-tridecadien-1-ol ↑; DHEAS, EPKSI ↓ | [92] | ||
| HUPLC/UHPLC-MS | Serum and urine | 124 PTC vs. 76 BN and 116 CTR | PTC vs. BN and CTR: Serum β-HBA, DHA, 1-MeAD ↑, pregnanediol-3-GLC, urinary NIC mononucleotide and XNTO ↓ | [89] | ||
| UPLC-Q/TOF-MS | Tissue and systemic profiling | Tissue, serum and plasma | 141 PTC vs. 93 BN and 100 CTR plus validation sets in 6 independent centers | PTC vs. CTR: Serum: 17 significantly changed metabolites; Plasma: 42 significantly changed metabolites, such as PB, L-E ↑; myo-IST, alpha-N-phenylacetyl-L-Q, lyso PC (18:0) and (18:1) ↓ PTC vs. BN: No significant differences in serum/plasma; Tissue: 16 significantly changed metabolites | [90] | |
| GC-MS | Culture cells | Thyrospheres with cancer stem-like cells | Cancer thyrospheres vs. cancer parental adherent cells and to non-cancer thyrospheres | SCA, MLI, D, E ↑; GLU, PYR, FRU ↓ | [80] | |
| NMR and MS | 1H-NMR and GC−FID/MS | Tissue extracts | Tissue extracts | 53 thyroid lesions vs. 46 CTR from the same participant | Ch, PC, GPC, PEA, LAC, GSH, TAU, myo- and scyllo-IST, IN, FUM, URD and Aa ↑; Lip ↓ | [59] |
| Other | FT-Raman | Tissue | Tissue | 6 MN vs. 10 BN | T3 and T4 hormones ↑ | [43] |
| Hyperspectral Raman microscopy | Tissue extracts | Single cells | 5 PTC vs. 5 BN | Lip; Nuc ↑; F, W, Prot, ↓ | [42] | |
| Capillary electrophoresis | Systemic profiling | Urine | 12 TC vs. 12 CTR | IN, N2-MG, N2,N2-DMG, 1-MG ↑ | [87] | |
| Amino acid analyser | Plasma | 33 TC vs. 137 CTR | M, L, Y and K ↑ | [70] |
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| AUS/FLUS | Atypical or follicular lesions of undetermined significance |
| DDA | Data-dependent acquisition |
| DIA | Data-independent acquisition |
| FNAB | Fine-needle aspiration biopsy |
| GABA | Gamma-amino butyric acid |
| GC-MS | Gas chromatography coupled to mass spectrometry |
| HMDB | Human metabolome database |
| HR-MAS | High-resolution magic angle spinning |
| J-Res | J-resolved spectroscopy |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LC-MS | Liquid chromatography coupled to mass spectrometry |
| MALDI-TOF/TOF | Matrix-assisted laser desorption/ionisation tandem time-of-flight |
| MAPK | Mitogen-activated protein kinase |
| miRNAs | Micro RNA molecules |
| MRSI | Magnetic resonance spectroscopy imaging |
| MS | Mass spectrometry |
| NMR | Nuclear magnetic resonance |
| OPLS-DA | Orthogonalized partial least squares discriminant analysis |
| ROC | Receiver operating characteristic |
| TOCSY | Total correlation spectroscopy |
| UPLC-MS | Ultra-performance liquid chromatography coupled to mass spectrometry |
References
- Sipos, J.A.; Mazzaferri, E.L. Thyroid cancer epidemiology and prognostic variables. Clin. Oncol. 2010, 22, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Sherma, S.I. Thyroid carcinoma. Lancet 2003, 361, 501–511. [Google Scholar] [CrossRef]
- Kondo, T.; Ezzat, S.; Asa, S.L. Pathogenetic mechanisms in thyroid follicular-cell neoplasia. Nat. Rev. Cancer 2006, 6, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Figge, J.J. Epidemiology of thyroid cancer. In Thyroid Cancer: A Comprehensive Guide to Clinical Management; Wartofsky, L., Van Nostrand, D., Eds.; Springer: New York, NY, USA, 2016; Volume 2, pp. 9–15. [Google Scholar]
- Knudsen, N.; Laurberg, P.; Perrild, H.; Bülow, I.; Ovesen, L.; Jørgensen, T. Risk factors for goiter and thyroid nodules. Thyroid 2002, 12, 879–888. [Google Scholar] [CrossRef]
- Albi, E.; Cataldi, S.; Lazzarini, A.; Codini, M.; Beccari, T.; Ambesi-Impiombato, F.S.; Curcio, F. Radiation and thyroid cancer. Int. J. Mol. Sci. 2017, 18, 911. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Zaballos, M.A.; Santisteban, P. Key signaling pathways in thyroid cancer. J. Endocrinol. 2017, 235, R43–R61. [Google Scholar] [CrossRef]
- Alshaikh, S.; Harb, Z.; Aljufairi, E.; Almahari, S.A. Classification of thyroid fine-needle aspiration cytology into Bethesda categories: An institutional experience and review of the literature. Cytojournal 2018, 15, 4. [Google Scholar] [CrossRef]
- Bongiovanni, M.; Spitale, A.; Faquin, W.C.; Mazzucchelli, L.; Baloch, Z.W. The Bethesda system for reporting thyroid cytopathology: A meta-analysis. Acta Cytol. 2012, 56, 333–339. [Google Scholar] [CrossRef]
- Gill, K.S.; Tassone, P.; Hamilton, J.; Hjelm, N.; Luginbuhl, A.; Cognetti, D.; Tuluc, M.; Martinez-Outschoorn, U.; Johnson, J.M.; Curry, J.M. Thyroid cancer metabolism: A review. J. Thyroid Disord. 2016, 5. [Google Scholar] [CrossRef]
- Mazzaferri, E.L. Thyroid cancer in thyroid nodules: Finding a needle in the haystack. Am. J. Med. 1992, 93, 359–362. [Google Scholar] [CrossRef]
- Nikiforov, Y.E. Thyroid carcinoma: Molecular pathways and therapeutic targets. Mod. Pathol. 2008, 21, S37–S43. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Hwang, T.S.; Choi, Y.L.; Kim, W.Y.; Han, H.S.; Lim, S.D.; Kim, W.S.; Yoo, Y.B.; Kim, S.K. Molecular profiling of papillary thyroid carcinoma in Korea with a high prevalence of BRAF(V600E) mutation. Thyroid 2017, 27, 802–810. [Google Scholar] [CrossRef]
- Pak, K.; Suh, S.; Kim, S.J.; Kim, I.J. Prognostic value of genetic mutations in thyroid cancer: A meta-analysis. Thyroid 2015, 25, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Kitabayashi, N.; Zhou, X.K.; Fahey, T.J., 3rd; Scognamiglio, T. MicroRNA analysis as a potential diagnostic tool for papillary thyroid carcinoma. Mod. Pathol. 2008, 21, 1139–1146. [Google Scholar] [CrossRef]
- Yip, L.; Kelly, L.; Shuai, Y.; Armstrong, M.J.; Nikiforov, Y.E.; Carty, S.E.; Nikiforova, M.N. MicroRNA signature distinguishes the degree of aggressiveness of papillary thyroid carcinoma. Ann. Surg. Oncol. 2011, 18, 2035–2041. [Google Scholar] [CrossRef]
- Pallante, P.; Visone, R.; Ferracin, M.; Ferraro, A.; Berlingieri, M.T.; Troncone, G.; Chiappetta, G.; Liu, C.G.; Santoro, M.; Negrini, M.; et al. MicroRNA deregulation in human thyroid papillary carcinomas. Endocr. Relat. Cancer 2006, 13, 497–508. [Google Scholar] [CrossRef]
- Pagni, F.; L’Imperio, V.; Bono, F.; Garancini, M.; Roversi, G.; De Sio, G.; Galli, M.; Smith, A.J.; Chinello, C.; Magni, F. Proteome analysis in thyroid pathology. Expert Rev. Proteom. 2015, 12, 375–390. [Google Scholar] [CrossRef]
- Damante, G.; Scaloni, A.; Tell, G. Thyroid tumors: Novel insights from proteomic studies. Expert Rev. Proteom. 2009, 6. [Google Scholar] [CrossRef]
- Nipp, M.; Elsner, M.; Balluff, B.; Meding, S.; Sarioglu, H.; Ueffing, M.; Rauser, S.; Unger, K.; Hofler, H.; Walch, A.; et al. S100-A10, thioredoxin, and S100-A6 as biomarkers of papillary thyroid carcinoma with lymph node metastasis identified by MALDI imaging. J. Mol. Med. 2012, 90, 163–174. [Google Scholar] [CrossRef]
- Sofiadis, A.; Dinets, A.; Orre, L.M.; Branca, R.M.; Juhlin, C.C.; Foukakis, T.; Wallin, G.; Hoog, A.; Hulchiy, M.; Zedenius, J.; et al. Proteomic study of thyroid tumors reveals frequent up-regulation of the Ca2+ -binding protein S100A6 in papillary thyroid carcinoma. Thyroid 2010, 20, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.; Brown, S.M.H.; Stephen, W.; Hunsucker Romana, T.; Netea, M.; Simon, A.; Chiang, D.E.H.; Kenneth, R.; Shroyer Mark, W.; Duncan, B.; et al. Quantitative and qualitative differences in protein expression between papillary thyroid carcinoma and normal thyroid tissue. Mol. Carcinog. 2006, 45, 613–626. [Google Scholar]
- Sofiadis, A.; Becker, S.; Hellman, U.; Hultin-Rosenberg, L.; Dinets, A.; Hulchiy, M.; Zedenius, J.; Wallin, G.; Foukakis, T.; Hoog, A.; et al. Proteomic profiling of follicular and papillary thyroid tumors. Eur. J. Endocrinol. 2012, 166, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Gawin, M.; Wojakowska, A.; Pietrowska, M.; Marczak, L.; Chekan, M.; Jelonek, K.; Lange, D.; Jaksik, R.; Gruca, A.; Widlak, P. Proteome profiles of different types of thyroid cancers. Mol. Cell Endocrinol. 2018, 472, 68–79. [Google Scholar] [CrossRef]
- Ito, Y.; Miyoshi, E.; Uda, E.; Yoshida, H.; Uruno, T.; Takamura, Y.; Miya, A.; Kobayashi, K.; Matsuzuka, F.; Matsuura, N.; et al. 14-3-3 σ possibly plays a constitutive role in papillary carcinoma, but not in follicular tumor of the thyroid. Cancer Lett. 2003, 200, 161–166. [Google Scholar] [CrossRef]
- Soudry, E.; Stern Shavit, S.; Hardy, B.; Morgenstern, S.; Hadar, T.; Feinmesser, R. Heat shock proteins HSP90, HSP70 and GRP78 expression in medullary thyroid carcinoma. Ann. Diagn. Pathol. 2017, 26, 52–56. [Google Scholar] [CrossRef]
- Mo, J.H.; Choi, I.J.; Jeong, W.J.; Jeon, E.H.; Ahn, S.H. HIF-1alpha and HSP90: Target molecules selected from a tumorigenic papillary thyroid carcinoma cell line. Cancer Sci. 2012, 103, 464–471. [Google Scholar] [CrossRef]
- Marsee, D.K.; Venkateswaran, A.; Tao, H.; Vadysirisack, D.; Zhang, Z.; Vandre, D.D.; Jhiang, S.M. Inhibition of heat shock protein 90, a novel RET/PTC1-associated protein, increases radioiodide accumulation in thyroid cells. J. Biol. Chem. 2004, 279, 43990–43997. [Google Scholar] [CrossRef]
- White, P.T.; Subramanian, C.; Zhu, Q.; Zhang, H.; Zhao, H.; Gallagher, R.; Timmermann, B.N.; Blagg, B.S.; Cohen, M.S. Novel HSP90 inhibitors effectively target functions of thyroid cancer stem cell preventing migration and invasion. Surgery 2016, 159, 142–151. [Google Scholar] [CrossRef]
- Thomas Ratajczak, T.; Ward, B.; Walsh, J.; Cluning, C. Hsp90 as a therapeutic target in endocrinology: Current evidence. Res. Rep. Endocr. Disord. 2015, 5, 141–155. [Google Scholar] [CrossRef]
- Lin, S.F.; Lin, L.J.; Hsueh, C.; Chou, T.C.; Yeh, C.N.; Chen, M.H.; Wong, R.J. Efficacy of an HSP90 inhibitor, ganetespib, in preclinical thyroid cancer models. Oncotarget 2017, 8, 41294–41304. [Google Scholar] [CrossRef] [PubMed]
- Deidda, M.; Piras, C.; Binaghi, G.; Congia, D.; Pani, A.; Boi, A.; Sanna, F.; Rossi, A.; Loi, B.; Cadeddu Dessalvi, C.; et al. Metabolomic fingerprint of coronary blood in STEMI patients depends on the ischemic time and inflammatory state. Sci. Rep. 2019, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Piras, C.; Arisci, N.; Poddighe, S.; Liggi, S.; Mariotti, S.; Atzori, L. Metabolomic profile in hyperthyroid patients before and after antithyroid drug treatment: Correlation with thyroid hormone and TSH concentration. Int. J. Biochem. Cell Biol. 2017, 93, 119–128. [Google Scholar] [CrossRef]
- Griffin, J.L.; Atherton, H.; Shockcor, J.; Atzori, L. Metabolomics as a tool for cardiac research. Nat. Rev. Cardiol. 2011, 8, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Deidda, M.; Piras, C.; Cadeddu Dessalvi, C.; Locci, E.; Barberini, L.; Orofino, S.; Musu, M.; Mura, M.N.; Manconi, P.E.; Finco, G.; et al. Distinctive metabolomic fingerprint in scleroderma patients with pulmonary arterial hypertension. Int. J. Cardiol. 2017, 241, 401–406. [Google Scholar] [CrossRef]
- Fanos, V.; Pintus, M.C.; Lussu, M.; Atzori, L.; Noto, A.; Stronati, M.; Guimaraes, H.; Marcialis, M.A.; Rocha, G.; Moretti, C.; et al. Urinary metabolomics of bronchopulmonary dysplasia (BPD): Preliminary data at birth suggest it is a congenital disease. J. Matern. Fetal Neonatal Med. 2014, 27 (Suppl. 2), 39–45. [Google Scholar] [CrossRef]
- Iuculano, A.; Murgia, F.; Peddes, C.; Santoru, M.L.; Tronci, L.; Deiana, M.; Balsamo, A.; Euser, A.; Atzori, L.; Monni, G. Metabolic scharacterisation of amniotic fluids of fetuses with enlarged nuchal translucency. J. Perinat. Med. 2019, 47, 311–318. [Google Scholar] [CrossRef]
- Bingol, K. Recent advances in targeted and untargeted metabolomics by NMR and MS/NMR methods. High Throughput 2018, 7, 9. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Cui, L.; Lu, H.; Lee, Y.H. Challenges and emergent solutions for LC-MS/MS based untargeted metabolomics in diseases. Mass Spectrom. Rev. 2018, 37, 772–792. [Google Scholar] [CrossRef]
- De Oliveira, M.A.S.; Campbell, M.; Afify, A.M.; Huang, E.C.; Chan, J.W. Hyperspectral Raman microscopy can accurately differentiate single cells of different human thyroid nodules. Biomed. Opt. Express 2019, 10, 4411–4421. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.S.; Bitar, R.A.; Martinho, H.S.; Santos, A.B.; Kulcsar, M.A.; Friguglietti, C.U.; da Costa, R.B.; Arisawa, E.A.; Martin, A.A. Thyroid tissue analysis through Raman spectroscopy. Analytic 2009, 134, 2361–2370. [Google Scholar] [CrossRef]
- Russell, P.; Lean, C.; Delbridge, L.; May, G.; Dowd, S.; Mountford, C. Proton magnetic resonance and human thyroid neoplasia I: Discrimination between benign and malignant neoplasms. Am. J. Med. 1994, 96, 383–388. [Google Scholar] [CrossRef]
- Mackinnon, W.B.; Delbridge, L.; Russell, P.; Lean, C.L.; May, G.L.; Doran, S.; Dowd, S.; Mountford, C.E. Two-dimensional proton magnetic resonance spectroscopy for tissue scharacterisation of thyroid neoplasms. World J. Surg. 1996, 20, 841–847. [Google Scholar] [CrossRef] [PubMed]
- King, A.D.; Yeung, D.K.; Ahuja, A.T.; Tse, G.M.; Chan, A.B.; Lam, S.S.; van Hasselt, A.C. In vivo 1H MR spectroscopy of thyroid carcinoma. Eur. J. Radiol. 2005, 54, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Goswami, B.; Chowdhury, V.; RaviShankar, L.; Kakar, A. Evaluation of the role of magnetic resonance spectroscopy in the diagnosis of follicular malignancies of thyroid. Arch. Surg. 2011, 146, 179–182. [Google Scholar] [CrossRef]
- Gupta, N.; Kakar, A.K.; Chowdhury, V.; Gulati, P.; Shankar, L.R.; Vindal, A. Magnetic resonance spectroscopy as a diagnostic modality for carcinoma thyroid. Eur. J. Radiol. 2007, 64, 414–418. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Sasaki, J.; Yamamoto, M.; Saitoh, K.; Nakaya, S.; Kubokawa, M. Quantitation by (1)H-NMR of dolichol, cholesterol and choline-containing lipids in extracts of normal and phathological thyroid tissue. Nmr Biomed. 2000, 13, 377–383. [Google Scholar] [CrossRef]
- Raffelt, K.; Moka, D.; Süllentrop, F.; Dietlein, M.; Hahn, J.; Schicha, H. Systemic alterations in phospholipid concentrations of blood plasma in patients with thyroid carcinoma: An in-vitro 31P high-resolution NMR study. Nmr Biomed. 2000, 13, 8–13. [Google Scholar] [CrossRef]
- Jordan, K.W.; Adkins, C.B.; Cheng, L.L.; Faquin, W.C. Application of magnetic-resonance-spectroscopy- based metabolomics to the fine-needle aspiration diagnosis of papillary thyroid carcinoma. Acta Cytol. 2011, 55, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Yin, P.; Su, D.; Peng, Z.; Zhou, L.; Ma, L.; Guo, W.; Ma, L.; Xu, G.; Shi, J.; et al. Serum metabolic profiling and features of papillary thyroid carcinoma and nodular goiter. Mol. Biosyst. 2011, 7, 2608–2614. [Google Scholar] [CrossRef] [PubMed]
- Miccoli, P.; Torregrossa, L.; Shintu, L.; Magalhaes, A.; Chandran, J.; Tintaru, A.; Ugolini, C.; Minuto, M.N.; Miccoli, M.; Basolo, F.; et al. Metabolomics approach to thyroid nodules: A high-resolution magic-angle spinning nuclear magnetic resonance-based study. Surgery 2012, 152, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Torregrossa, L.; Shintu, L.; Nambiath Chandran, J.; Tintaru, A.; Ugolini, C.; Magalhaes, A.; Basolo, F.; Miccoli, P.; Caldarelli, S. Toward the reliable diagnosis of indeterminate thyroid lesions: A HRMAS NMR-based metabolomics case of study. J. Proteome Res. 2012, 11, 3317–3325. [Google Scholar] [CrossRef]
- Carter, J.V.; Pan, J.; Rai, S.N.; Galandiuk, S. ROC-ing along: Evaluation and interpretation of receiver operating characteristic curves. Surgery 2016, 159, 1638–1645. [Google Scholar] [CrossRef]
- Lu, J.; Hu, S.; Miccoli, P.; Zeng, Q.; Liu, S.; Ran, L.; Hu, C. Non-invasive diagnosis of papillary thyroid microcarcinoma: A NMR-based metabolomics approach. Oncotarget 2016, 7, 81768–81777. [Google Scholar] [CrossRef]
- Rezig, L.; Servadio, A.; Torregrossa, L.; Miccoli, P.; Basolo, F.; Shintu, L.; Caldarelli, S. Diagnosis of post-surgical fine-needle aspiration biopsies of thyroid lesions with indeterminate cytology using HRMAS NMR-based metabolomics. Metabolomics 2018, 14, 141. [Google Scholar] [CrossRef]
- Deja, S.; Dawiskiba, T.; Balcerzak, W.; Orczyk-Pawilowicz, M.; Glod, M.; Pawelka, D.; Mlynarz, P. Follicular adenomas exhibit a unique metabolic profile. (1)H NMR studies of thyroid lesions. PLoS ONE 2013, 8, e84637. [Google Scholar] [CrossRef]
- Tian, Y.; Nie, X.; Xu, S.; Li, Y.; Huang, T.; Tang, H.; Wang, Y. Integrative metabonomics as potential method for diagnosis of thyroid malignancy. Sci. Rep. 2015, 5, 14869. [Google Scholar] [CrossRef]
- Ryoo, I.; Kwon, H.; Kim, S.C.; Jung, S.C.; Yeom, J.A.; Shin, H.S.; Cho, H.R.; Yun, T.J.; Choi, S.H.; Sohn, C.H.; et al. Metabolomic analysis of percutaneous fine-needle aspiration specimens of thyroid nodules: Potential application for the preoperative diagnosis of thyroid cancer. Sci. Rep. 2016, 6, 30075. [Google Scholar] [CrossRef]
- Seo, J.W.; Han, K.; Lee, J.; Kim, E.K.; Moon, H.J.; Yoon, J.H.; Park, V.Y.; Baek, H.M.; Kwak, J.Y. Application of metabolomics in prediction of lymph node metastasis in papillary thyroid carcinoma. PLoS ONE 2018, 13, e0193883. [Google Scholar] [CrossRef]
- Li, Y.; Chen, M.; Liu, C.; Xia, Y.; Xu, B.; Hu, Y.; Chen, T.; Shen, M.; Tang, W. Metabolic changes associated with papillary thyroid carcinoma: A nuclear magnetic resonance-based metabolomics study. Int. J. Mol. Med. 2018, 41, 3006–3014. [Google Scholar] [CrossRef] [PubMed]
- Metere, A.; Graves, C.E.; Chirico, M.; Caramujo, M.J.; Pisanu, M.E.; Iorio, E. Metabolomic reprogramming detected by (1)H-NMR spectroscopy in human thyroid cancer tissues. Biology 2020, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, S.; Liu, P.; Bjorndahl, T.C.; Mandal, R.; Grant, J.R.; Wilson, M.; Eisner, R.; Sinelnikov, I.; Hu, X.; Luchinat, C.; et al. Correction: Accurate, fully-automated NMR spectral profiling for metabolomics. PLoS ONE 2015, 10, e0132873. [Google Scholar] [CrossRef]
- Jimenez, B.; Mirnezami, R.; Kinross, J.; Cloarec, O.; Keun, H.C.; Holmes, E.; Goldin, R.D.; Ziprin, P.; Darzi, A.; Nicholson, J.K. 1H HR-MAS NMR spectroscopy of tumor-induced local metabolic “field-effects” enables colorectal cancer staging and prognostication. J. Proteome Res. 2013, 12, 959–968. [Google Scholar] [CrossRef]
- Lutz, N.W.; Sweedler, J.V.; Wevers, R.A. Methodologies for Metabolomics: Experimental Strategies and Techniques; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Zhou, J.; Li, Y.; Chen, X.; Zhong, L.; Yin, Y. Development of data-independent acquisition workflows for metabolomic analysis on a quadrupole-orbitrap platform. Talanta 2017, 164, 128–136. [Google Scholar] [CrossRef]
- Mendes, V.M.; Coelho, M.; Manadas, B. Untargeted metabolomics relative quantification by SWATH mass spectrometry applied to cerebrospinal fluid. Methods Mol. Biol. 2019, 2044, 321–336. [Google Scholar] [CrossRef]
- Gu, Y.; Chen, T.; Fu, S.; Sun, X.; Wang, L.; Wang, J.; Lu, Y.; Ding, S.; Ruan, G.; Teng, L.; et al. Perioperative dynamics and significance of amino acid profiles in patients with cancer. J. Transl. Med. 2015, 13, 35. [Google Scholar] [CrossRef]
- Ishikawa, S.; Tateya, I.; Hayasaka, T.; Masaki, N.; Takizawa, Y.; Ohno, S.; Kojima, T.; Kitani, Y.; Kitamura, M.; Hirano, S.; et al. Increased expression of phosphatidylcholine (16:0/18:1) and (16:0/18:2) in thyroid papillary cancer. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Guo, S.; Qiu, L.; Wang, Y.; Qin, X.; Liu, H.; He, M.; Zhang, Y.; Li, Z.; Chen, X. Tissue imaging and serum lipidomic profiling for screening potential biomarkers of thyroid tumors by matrix-assisted laser desorption/ionization-Fourier transform ion cyclotron resonance mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 4357–4370. [Google Scholar] [CrossRef]
- Zhang, J.; Feider, C.L.; Nagi, C.; Yu, W.; Carter, S.A.; Suliburk, J.; Cao, H.S.T.; Eberlin, L.S. Detection of metastatic breast and thyroid cancer in lymph nodes by desorption electrospray sionisation mass spectrometry imaging. J. Am. Soc. Mass Spectrom. 2017, 28, 1166–1174. [Google Scholar] [CrossRef]
- Huang, L.; Mao, X.; Sun, C.; Luo, Z.; Song, X.; Li, X.; Zhang, R.; Lv, Y.; Chen, J.; He, J.; et al. A graphical data processing pipeline for mass spectrometry imaging-based spatially resolved metabolomics on tumor heterogeneity. Anal. Chim. Acta 2019, 1077, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Wojakowska, A.; Cole, L.M.; Chekan, M.; Bednarczyk, K.; Maksymiak, M.; Oczko-Wojciechowska, M.; Jarzab, B.; Clench, M.R.; Polanska, J.; Pietrowska, M.; et al. Discrimination of papillary thyroid cancer from non-cancerous thyroid tissue based on lipid profiling by mass spectrometry imaging. Endokrynol. Pol. 2018, 69, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Shen, M.; Li, Y.; Liu, C.; Zhou, K.; Hu, W.; Xu, B.; Xia, Y.; Tang, W. GC-MS-based metabolomic analysis of human papillary thyroid carcinoma tissue. Int. J. Mol. Med. 2015, 36, 1607–1614. [Google Scholar] [CrossRef]
- Shang, X.; Zhong, X.; Tian, X. Metabolomics of papillary thyroid carcinoma tissues: Potential biomarkers for diagnosis and promising targets for therapy. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 11163–11175. [Google Scholar] [CrossRef]
- Wojakowska, A.; Chekan, M.; Marczak, L.; Polanski, K.; Lange, D.; Pietrowska, M.; Widlak, P. Detection of metabolites discriminating subtypes of thyroid cancer: Molecular profiling of FFPE samples using the GC/MS approach. Mol. Cell Endocrinol. 2015, 417, 149–157. [Google Scholar] [CrossRef]
- Shen, C.T.; Zhang, Y.; Liu, Y.M.; Yin, S.; Zhang, X.Y.; Wei, W.J.; Sun, Z.K.; Song, H.J.; Qiu, Z.L.; Wang, C.R.; et al. A distinct serum metabolic signature of distant metastatic papillary thyroid carcinoma. Clin. Endocrinol. 2017, 87, 844–852. [Google Scholar] [CrossRef]
- Caria, P.; Tronci, L.; Dettori, T.; Murgia, F.; Santoru, M.L.; Griffin, J.L.; Vanni, R.; Atzori, L. Metabolomic alterations in thyrospheres and adherent parental cells in papillary thyroid carcinoma cell lines: A pilot study. Int. J. Mol. Sci. 2018, 19, 2948. [Google Scholar] [CrossRef]
- Tronci, L.; Caria, P.; Frau, D.V.; Liggi, S.; Piras, C.; Murgia, F.; Santoru, M.L.; Pibiri, M.; Deiana, M.; Griffin, J.L.; et al. Crosstalk between metabolic alterations and altered redox balance in PTC-derived cell lines. Metabolites 2019, 9, 23. [Google Scholar] [CrossRef]
- Agrawal, S.; Kumar, S.; Sehgal, R.; George, S.; Gupta, R.; Poddar, S.; Jha, A.; Pathak, S. El-MAVEN: A fast, robust, and user-friendly mass spectrometry data processing engine for metabolomics. Methods Mol. Biol. 2019, 1978, 301–321. [Google Scholar] [CrossRef] [PubMed]
- Abooshahab, R.; Hooshmand, K.; Razavi, S.A.; Gholami, M.; Sanoie, M.; Hedayati, M. Plasma Metabolic Profiling of Human Thyroid Nodules by Gas Chromatography-Mass Spectrometry (GC-MS)-Based Untargeted Metabolomics. Front. Cell Dev. Biol. 2020, 8, 385. [Google Scholar] [CrossRef]
- Zhao, W.-X.; Wang, B.; Zhang, L.-Y.; Yan, S.-Y.; Yang, Y.-H. Analysis on the metabolite composition of serum samples from patients with papillary thyroid carcinoma using nuclear magnetic resonance. Int. J. Clin. Exp. Med. 2015, 8, 18013. [Google Scholar] [PubMed]
- Farrokhi Yekta, R.; Rezaei Tavirani, M.; Arefi Oskouie, A.; Mohajeri-Tehrani, M.R.; Soroush, A.R.; Akbarzadeh Baghban, A. Serum-based metabolic alterations in patients with papillary thyroid carcinoma unveiled by non-targeted 1H-NMR metabolomics approach. Iran J. Basic Med. Sci. 2018, 21, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, L.Y.; Xie, C.; Zhang, M.L.; Wang, Y.J.; Liu, G.H. Metabolomics as a potential method for predicting thyroid malignancy in children and adolescents. Pediatr. Surg. Int. 2019. [Google Scholar] [CrossRef]
- La, S.; Cho, J.; Kim, J.-H.; Kim, K.-R. Capillary electrophoretic profiling and pattern recognition analysis of urinary nucleosides from thyroid cancer patients. Anal. Chim. Acta 2003, 486, 171–182. [Google Scholar] [CrossRef]
- Wojtowicz, W.; Zabek, A.; Deja, S.; Dawiskiba, T.; Pawelka, D.; Glod, M.; Balcerzak, W.; Mlynarz, P. Serum and urine (1)H NMR-based metabolomics in the diagnosis of selected thyroid diseases. Sci. Rep. 2017, 7, 9108. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, Q.; Hou, H.; Wang, S.; Zhang, Y.; Luo, Y.; Chen, H.; Deng, H.; Zhu, H.; Zhang, L.; et al. Metabolite analysis-aided diagnosis of papillary thyroid cancer. Endocr. Relat. Cancer 2019, 26, 829–841. [Google Scholar] [CrossRef]
- Huang, F.Q.; Li, J.; Jiang, L.; Wang, F.X.; Alolga, R.N.; Wang, M.J.; Min, W.J.; Ma, G.; Zhao, Y.J.; Wang, S.L.; et al. Serum-plasma matched metabolomics for comprehensive scharacterisation of benign thyroid nodule and papillary thyroid carcinoma. Int. J. Cancer 2019, 144, 868–876. [Google Scholar] [CrossRef]
- Lee, G.B.; Lee, J.C.; Moon, M.H. Plasma lipid profile comparison of five different cancers by nanoflow ultrahigh performance liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2019, 1063, 117–126. [Google Scholar] [CrossRef]
- Feng, J.; Zhao, F.; Sun, J.; Lin, B.; Zhao, L.; Liu, Y.; Jin, Y.; Li, S.; Li, A.; Wei, Y. Alterations in the gut microbiota and metabolite profiles of thyroid carcinoma patients. Int. J. Cancer 2019, 144, 2728–2745. [Google Scholar] [CrossRef]
- Guo, L.; Wang, C.; Chi, C.; Wang, X.; Liu, S.; Zhao, W.; Ke, C.; Xu, G.; Li, E. Exhaled breath volatile biomarker analysis for thyroid cancer. Transl. Res. J. Lab. Clin. Med. 2015, 166, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the origin of cancer cells. Science 1956, 123. [Google Scholar] [CrossRef]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef]
- San-Millan, I.; Brooks, G.A. Reexamining cancer metabolism: Lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis 2017, 38, 119–133. [Google Scholar] [CrossRef]
- An, Y.J.; Cho, H.R.; Kim, T.M.; Keam, B.; Kim, J.W.; Wen, H.; Park, C.K.; Lee, S.H.; Im, S.A.; Kim, J.E.; et al. An NMR metabolomics approach for the diagnosis of leptomeningeal carcinomatosis in lung adenocarcinoma cancer patients. Int. J. Cancer 2015, 136, 162–171. [Google Scholar] [CrossRef]
- Suman, S.; Sharma, R.K.; Kumar, V.; Sinha, N.; Shukla, Y. Metabolic fingerprinting in breast cancer stages through (1)H NMR spectroscopy-based metabolomic analysis of plasma. J. Pharm. Biomed. Anal. 2018, 160, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Michalkova, L.; Hornik, S.; Sykora, J.; Habartova, L.; Setnicka, V. Diagnosis of pancreatic cancer via(1)H NMR metabolomics of human plasma. Analyst 2018, 143, 5974–5978. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.S.; Etten, R.A.V. Tyrosine kinases as targets for cancer therapy. N. Engl. J. Med. 2005, 353, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Groot, J.W.B.d.; Kema, I.P.; Breukelman, H.; Veer, E.v.d.; Wiggers, T.; Plukker, J.T.M.; Wolffenbuttel, B.H.R.; Links, T.P. Biochemical markers in the follow-up of medullary thyroid cancer. Thyroid 2006, 16, 1163–1170. [Google Scholar] [CrossRef]
- Jia, M.; Andreassen, T.; Jensen, L.; Bathen, T.F.; Sinha, I.; Gao, H.; Zhao, C.; Haldosen, L.A.; Cao, Y.; Girnita, L.; et al. Estrogen receptor alpha promotes breast cancer by reprogramming choline metabolism. Cancer Res. 2016, 76, 5634–5646. [Google Scholar] [CrossRef]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, E.O.; Bhujwalla, Z.M. Malignant transformation alters membrane choline phospholipid metabolism of human mammary epithelial cells. Cancer Res. 1999, 59, 80–84. [Google Scholar] [PubMed]
- Khatami, F.; Payab, M.; Sarvari, M.; Gilany, K.; Larijani, B.; Arjmand, B.; Tavangar, S.M. Oncometabolites as biomarkers in thyroid cancer: A systematic review. Cancer Manag. Res. 2019, 11, 1829–1841. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. Publ. Protein Soc. 2019, 28, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an soptimised workflow for global metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zheng, X.; Qiu, Y.; Jia, W.; Wang, J.; Yin, S. Distinct metabolomic profiles of papillary thyroid carcinoma and benign thyroid adenoma. J. Proteome Res. 2015, 14, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Wiltshire, J.J.; Drake, T.M.; Uttley, L.; Balasubramanian, S.P. Systematic review of trends in the incidence rates of thyroid cancer. Thyroid 2016, 26, 1541–1552. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, M.; Raposo, L.; Goodfellow, B.J.; Atzori, L.; Jones, J.; Manadas, B. The Potential of Metabolomics in the Diagnosis of Thyroid Cancer. Int. J. Mol. Sci. 2020, 21, 5272. https://doi.org/10.3390/ijms21155272
Coelho M, Raposo L, Goodfellow BJ, Atzori L, Jones J, Manadas B. The Potential of Metabolomics in the Diagnosis of Thyroid Cancer. International Journal of Molecular Sciences. 2020; 21(15):5272. https://doi.org/10.3390/ijms21155272
Chicago/Turabian StyleCoelho, Margarida, Luis Raposo, Brian J. Goodfellow, Luigi Atzori, John Jones, and Bruno Manadas. 2020. "The Potential of Metabolomics in the Diagnosis of Thyroid Cancer" International Journal of Molecular Sciences 21, no. 15: 5272. https://doi.org/10.3390/ijms21155272
APA StyleCoelho, M., Raposo, L., Goodfellow, B. J., Atzori, L., Jones, J., & Manadas, B. (2020). The Potential of Metabolomics in the Diagnosis of Thyroid Cancer. International Journal of Molecular Sciences, 21(15), 5272. https://doi.org/10.3390/ijms21155272