A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry




Abstract
1. Introduction
2. Results
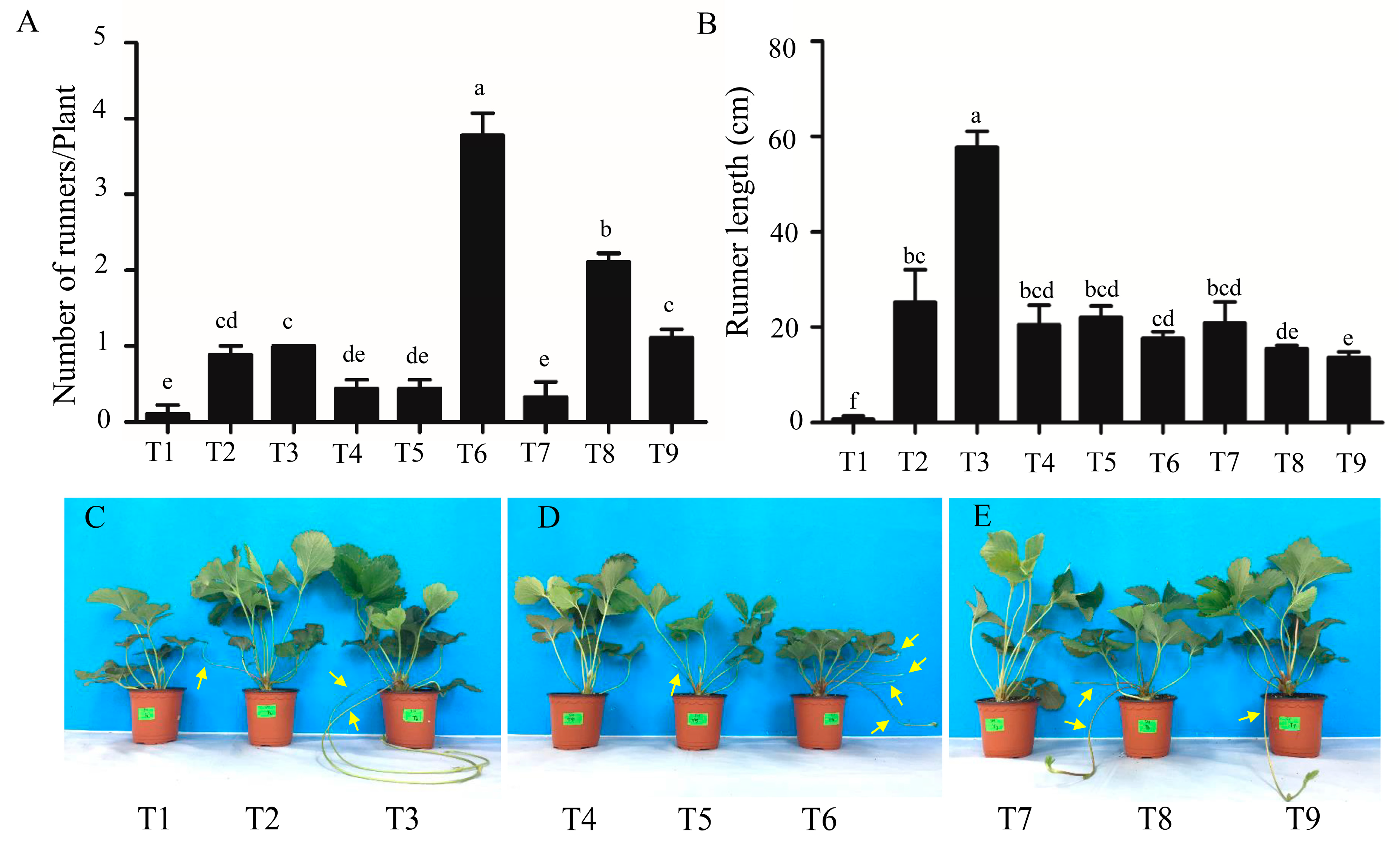
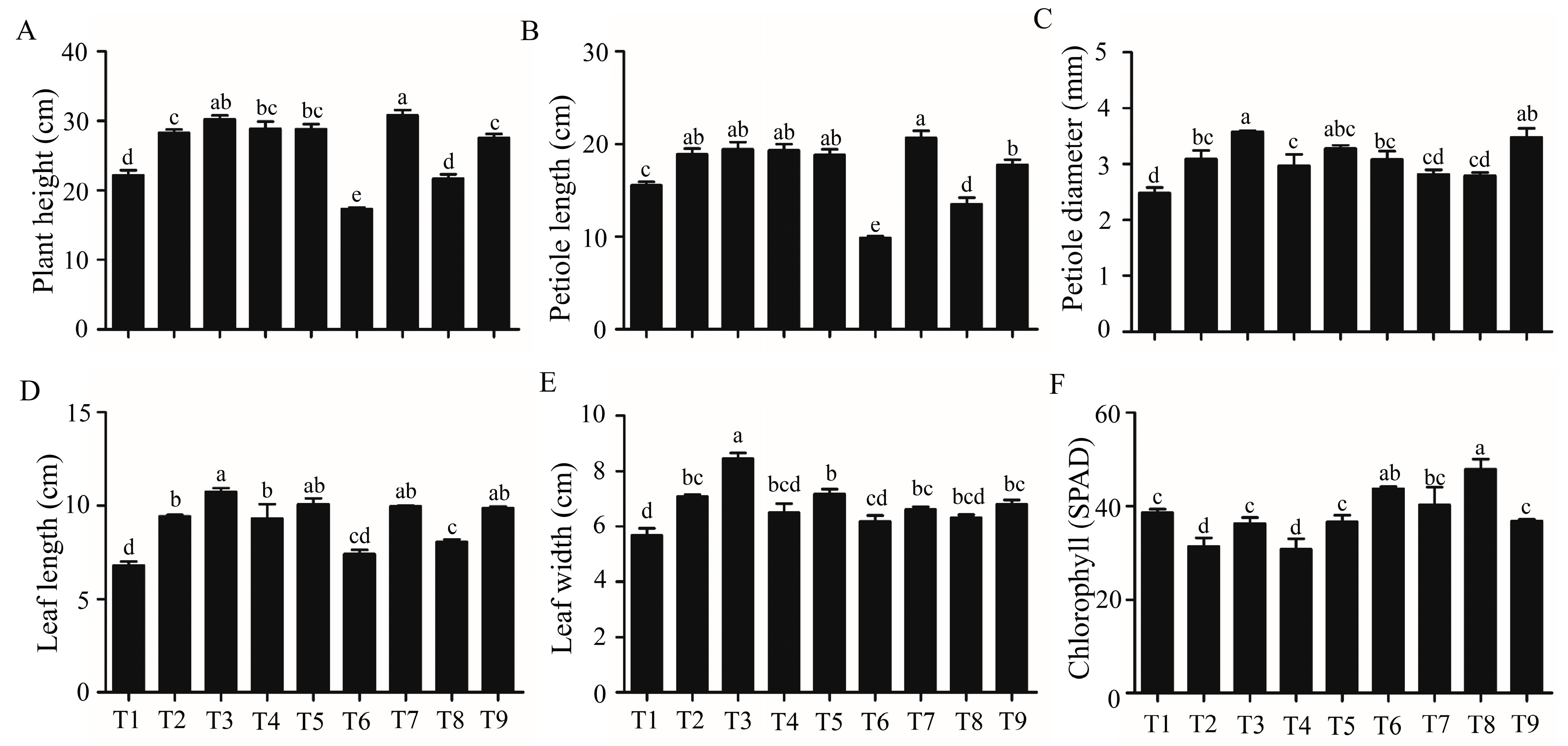
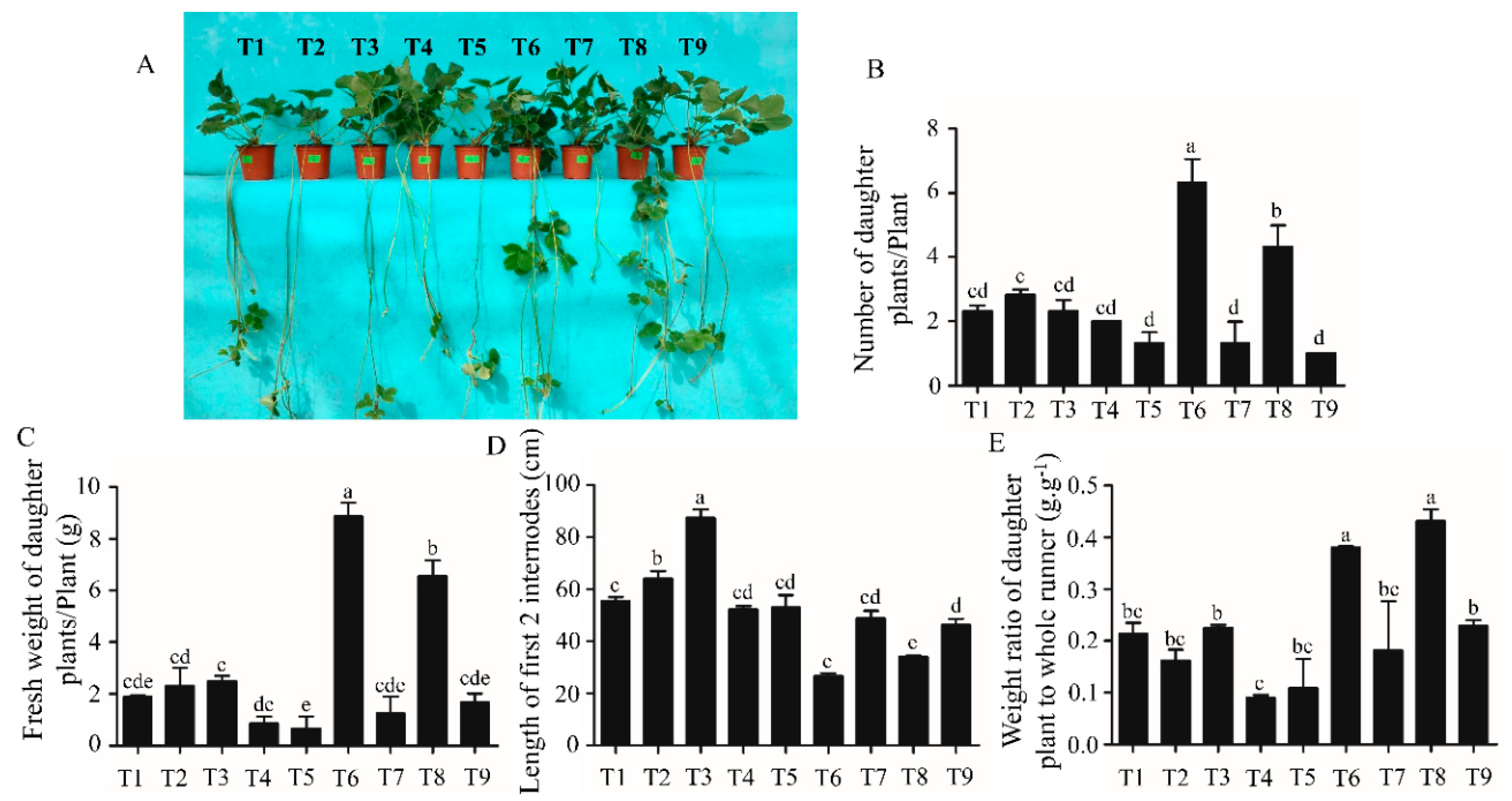
2.1. Analysis of the Morphological and Growth Parameters
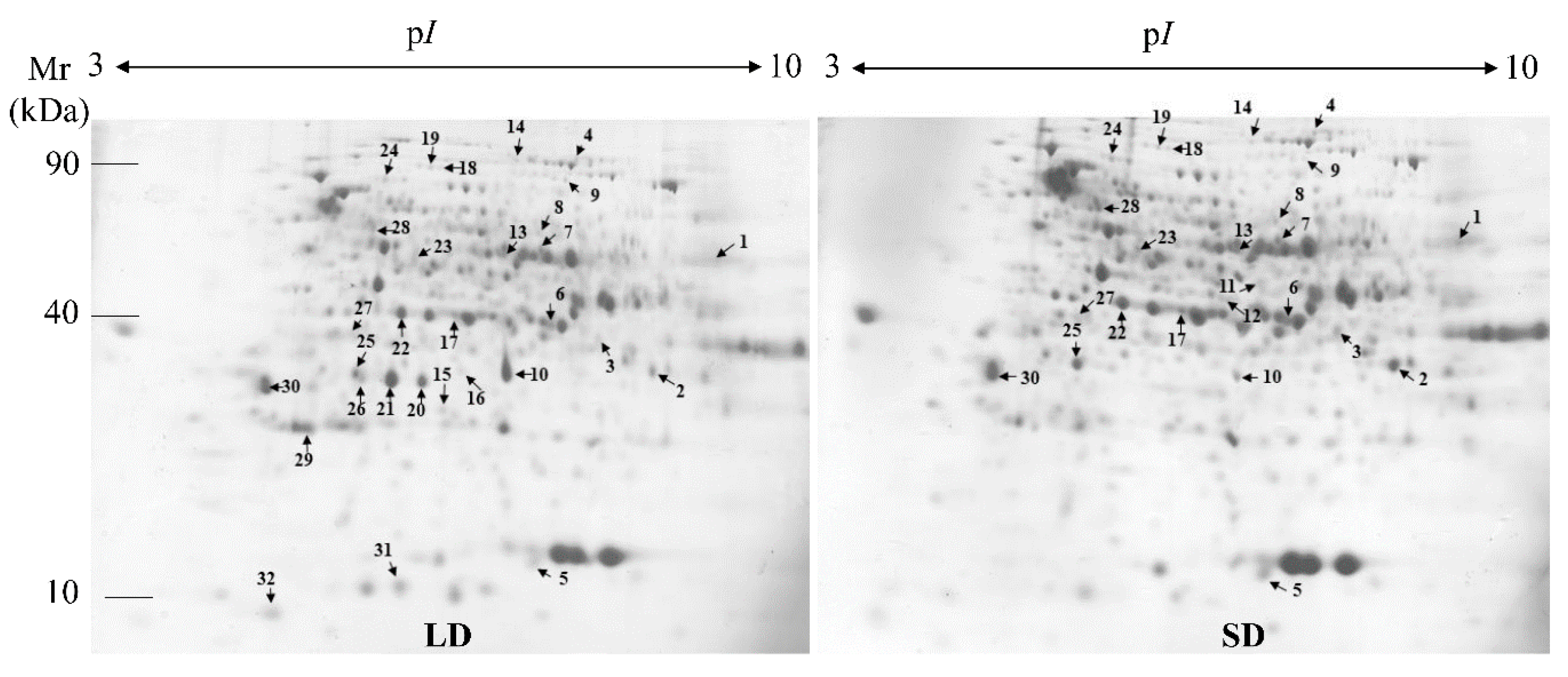
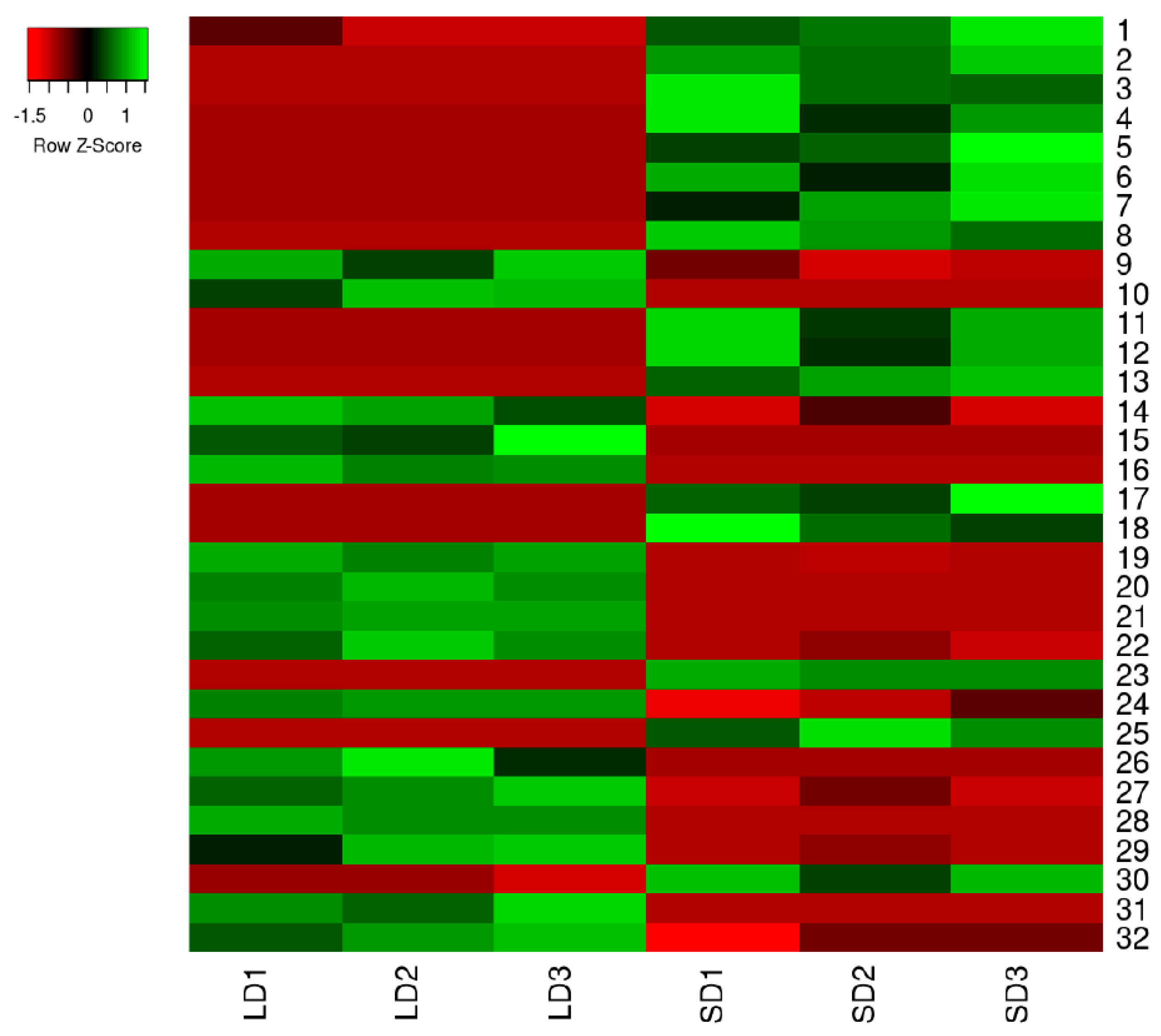
2.2. Protein Expression Profiles and Protein Identification
2.3. Functional Classification of the DEPs
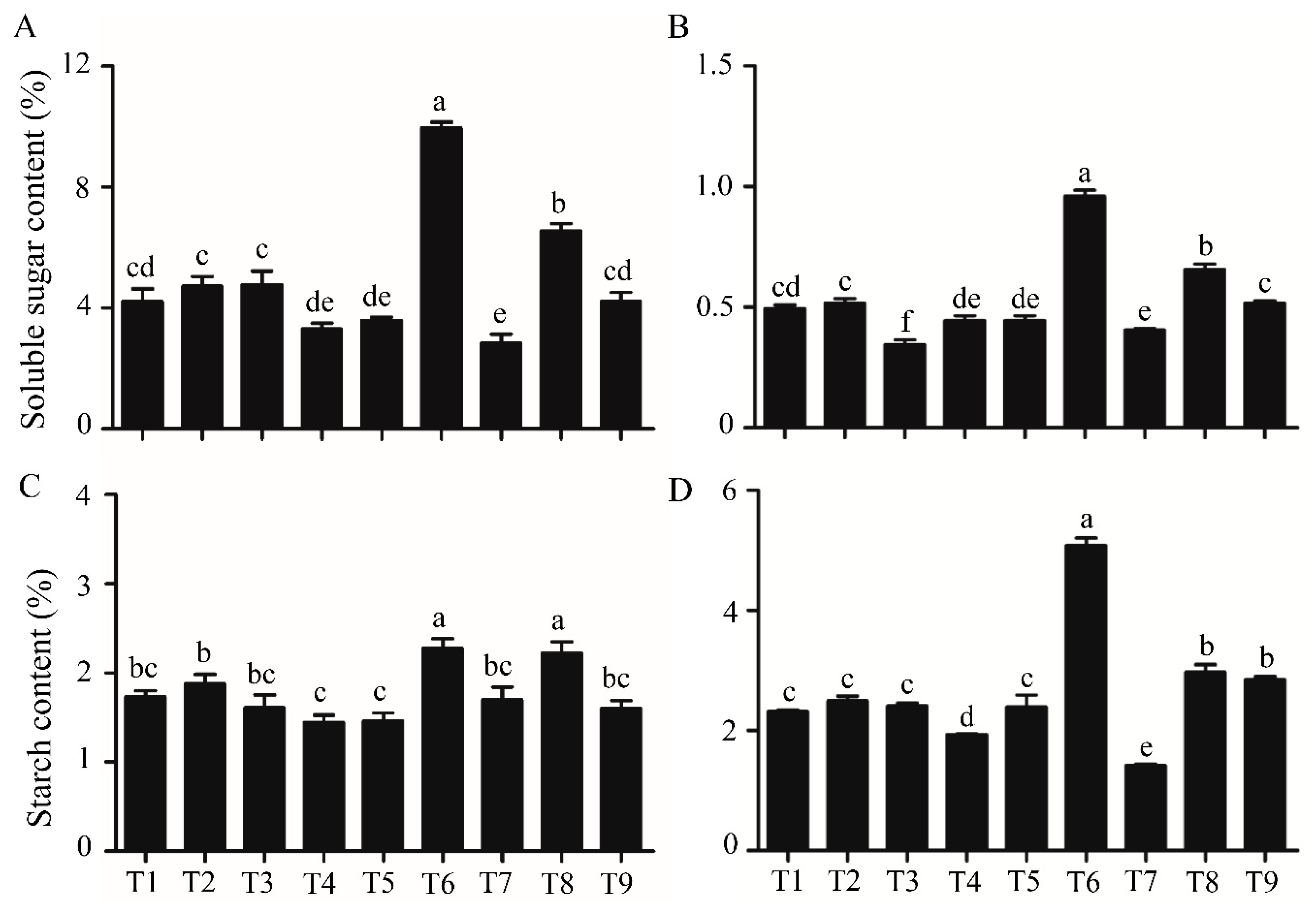
2.4. Soluble Sugar and Starch Contents
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Culture Condition
4.2. Measurement of the Chlorophyll Content
4.3. Proteomic Analysis
4.3.1. Materials for the Proteomic Analysis
4.3.2. Protein Extraction
4.3.3. Two-Dimensional Gel Electrophoresis and Image Analysis
4.3.4. In-Gel Digestion and MALDI-TOF/MS
4.3.5. Protein Functional Classification
4.4. Measurements of the Soluble Sugar and Starch Contents
4.5. Measurements of the Soluble Sugar and Starch Contents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hossain, M.R.; Natarajan, S.; Kim, H.-T.; Jesse, D.M.I.; Lee, C.-G.; Park, J.-I.; Nou, I.-S. High density linkage map construction and QTL mapping for runner production in allo-octoploid strawberry Fragaria× ananassa based on ddRAD-seq derived SNPs. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Caruana, J.C.; Sittmann, J.W.; Wang, W.; Liu, Z. Suppressor of runnerless encodes a DELLA protein that controls runner formation for asexual reproduction in strawberry. Mol. Plant. 2018, 11, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Hytönen, T.; Elomaa, P. Genetic and environmental regulation of flowering and runnering in strawberry. Genes Genom. Genom. 2011, 5, 56–64. [Google Scholar]
- Hytönen, T.; Elomaa, P.; Moritz, T.; Junttila, O. Gibberellin mediates daylength-controlled differentiation of vegetative meristems in strawberry (Fragaria× ananassa Duch). BMC Plant. Biol. 2009, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Heide, O.; Stavang, J.; Sønsteby, A. Physiology and genetics of flowering in cultivated and wild strawberries–a review. J. Hortic. Sci. Biotech. 2013, 88, 1–18. [Google Scholar] [CrossRef]
- Weebadde, C.; Wang, D.; Finn, C.; Lewers, K.; Luby, J.J.; Bushakra, J.; Sjulin, T.; Hancock, J. Using a linkage mapping approach to identify QTL for day-neutrality in the octoploid strawberry. Plant. Breeding 2008, 127, 94–101. [Google Scholar] [CrossRef]
- Castro, P.; Bushakra, J.; Stewart, P.; Weebadde, C.; Wang, D.; Hancock, J.; Finn, C.; Luby, J.J.; Lewers, K. Genetic mapping of day-neutrality in cultivated strawberry. Mol. Breeding 2015, 35, 79. [Google Scholar] [CrossRef]
- Heide, O.M. Photoperiod and temperature interactions in growth and flowering of strawberry. Physiol. Plant. 1977, 40, 21–26. [Google Scholar] [CrossRef]
- Hytönen, T.; Palonen, P.; Mouhu, K.; Junttila, O. Crown branching and cropping potential in strawberry (Fragaria ananassa Duch.) can be enhanced by daylength treatments. J. Hortic. Sci. Biotech. 2004, 79, 466–471. [Google Scholar] [CrossRef]
- Mouhu, K.; Kurokura, T.; Koskela, E.A.; Albert, V.A.; Elomaa, P.; Hytönen, T. The Fragaria vesca homolog of SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 represses flowering and promotes vegetative growth. Plant. Cell 2013, 25, 3296–3310. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O. Long-day control of flowering in everbearing strawberries. J. Hortic. Sci. Biotech. 2007, 82, 875–884. [Google Scholar] [CrossRef]
- Bradford, E.; Hancock, J.F.; Warner, R.M. Interactions of temperature and photoperiod determine expression of repeat flowering in strawberry. J. Am. Soc. Hortic. Sci. 2010, 135, 102–107. [Google Scholar] [CrossRef]
- Uddin, A.J.; Hossan, M.; Islam, M.; Ahsan, M.; Mehraj, H. Strawberry growth and yield responses to gibberellic acid concentrations. J. Expt. Biosci. 2012, 3, 51–56. [Google Scholar]
- Kumar, R.; Bakshi, M.; Singh, D. Influence of plant growth regulators on growth, yield and quality of strawberrry (Fragaria x ananassa Duch.) under UP sub tropics. Asian J. Hortic. 2012, 7, 434–436. [Google Scholar]
- Palei, S.; Das, K.; Sahoo, K.; Dash, D.K.; Swain, S. Influence of plant growth regulators on strawberry Cv. Chandler under Odisha condition. Int. J. Sci. Res. 2016, 7, 9945–9948. [Google Scholar]
- Greene, D.W.; Schloemann, S.G. Prohexadione-calcium inhibits runner formation and enhances yield of strawberry. J. Am. Pomol. Soc. 2010, 64, 125. [Google Scholar]
- Kim, H.M.; Lee, H.R.; Kang, J.H.; Hwang, S.J. Prohexadione-calcium application during vegetative growth affects growth of mother plants, runners, and runner plants of Maehyang strawberry. Agronomy 2019, 9, 155. [Google Scholar] [CrossRef]
- Qiu, Y.; Guan, S.C.; Wen, C.; Li, P.; Gao, Z.; Chen, X. Auxin and cytokinin coordinate the dormancy and outgrowth of axillary bud in strawberry runner. BMC Plant. Biol. 2019, 19, 528. [Google Scholar] [CrossRef]
- Kour, S.; Kumar, R.; Wali, V.; Sharma, A.; Bakshi, P. Impact of benzyladenine and gibberellic acid on quality and economics of runner production in Chandler strawberry (Fragaria× ananassa) under subtropical climate. Indian J. Agric. Sci. 2017, 87, 964–967. [Google Scholar]
- Liu, C.; Guo, Z.; Park, Y.G.; Wei, H.; Jeong, B.R. PGR and its application method affect number and length of runners produced in ‘Maehyang’and ‘Sulhyang’strawberries. Agronomy 2019, 9, 59. [Google Scholar] [CrossRef]
- Slovin, J.P.; Schmitt, K.; Folta, K.M. An inbred line of the diploid strawberry Fragaria vesca f. semperflorens for genomic and molecular genetic studies in the Rosaceae. Plant. Methods 2009, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Tenreira, T.; Lange, M.J.P.; Lange, T.; Bres, C.; Labadie, M.; Monfort, A.; Hernould, M.; Rothan, C.; Denoyes, B. A specific gibberellin 20-oxidase dictates the flowering-runnering decision in diploid strawberry. Plant. Cell 2017, 29, 2168–2182. [Google Scholar] [CrossRef]
- Li, W.; Zhang, J.; Sun, H.; Wang, S.; Chen, K.; Liu, Y.; Li, H.; Ma, Y.; Zhang, Z. FveRGA1, encoding a DELLA protein, negatively regulates runner production in Fragaria vesca. Planta 2018, 247, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.O.; Nunes-Nesi, A.; Araújo, W.L.; Fernie, A.R. To bring flowers or do a runner: Gibberellins make the decision. Mol. Plant. 2018, 11, 4–6. [Google Scholar] [CrossRef]
- Shaw, D.V.; Famula, T.R. Complex segregation analysis of day-neutrality in domestic strawberry (Fragaria× ananassa Duch.). Euphytica 2005, 145, 331–338. [Google Scholar] [CrossRef]
- Sugimoto, T.; Tamaki, K.; Matsumoto, J.; Yamamoto, Y.; Shiwaku, K.; Watanabe, K. Detection of RAPD markers linked to the everbearing gene in Japanese cultivated strawberry. Plant. Breeding 2005, 124, 498–501. [Google Scholar] [CrossRef]
- Gaston, A.; Perrotte, J.; Lerceteau-Köhler, E.; Rousseau-Gueutin, M.; Petit, A.; Hernould, M.; Rothan, C.; Denoyes, B. PFRU, a single dominant locus regulates the balance between sexual and asexual plant reproduction in cultivated strawberry. J. Exp. Bot. 2013, 64, 1837–1848. [Google Scholar] [CrossRef]
- Sooriyapathirana, S.S.; Mookerjee, S.; Weebadde, C.K.; Finn, C.E.; Lewers, K.; Bushakra, J.M.; Luby, J.J.; Stewart, P.; Neils, S.; Hancock, J.F. Identification of QTL associated with flower and runner production in octoploid strawberry (Fragaria× ananassa). J. Berry Res. 2015, 5, 107–116. [Google Scholar] [CrossRef]
- Durner, E.F.; Barden, J.; Himelrick, D.; Poling, E. Photoperiod and temperature effects on flower and runner development in day-neutral, Junebearing, and everbearing strawberries. J. Am. Soc. Hortic. Sci. 1984, 109, 396–400. [Google Scholar]
- Heide, O.M.; Sønsteby, A. Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca). Physiol. Plant. 2007, 130, 280–289. [Google Scholar] [CrossRef]
- Kender, W.; Carpenter, S.; Braun, J. Runner formation in everbearing strawberry as influenced by growth-promoting and inhibiting substances. Ann. Bot. 1971, 35, 1045–1052. [Google Scholar] [CrossRef]
- Momenpour, A.; Taghavi, T.S.; Manochehr, S. Effects of banzyladenine and gibberellin on runner production and some vegetative traits of three strawberry cultivars. Afr. J. Agric. Res. 2011, 6, 4357–4361. [Google Scholar]
- Surapornpiboon, P.; Surapornpiboon, P. Effect of benzyladenine and gibberellic acid on runner production in strawberry. In Proceedings of the 19. Rajamangala University of Technology Conference, Pathum Thani, Thailand, 22–27 January 2002. [Google Scholar]
- Dai, F.; Wang, Z.; Luo, G.; Tang, C. Phenotypic and transcriptomic analyses of autotetraploid and diploid mulberry (Morus alba L.). Int. J. Mol. Sci. 2015, 16, 22938–22956. [Google Scholar] [CrossRef] [PubMed]
- Jaskani, M.J.; Kwon, S.W.; Kim, D.H. Comparative study on vegetative, reproductive and qualitative traits of seven diploid and tetraploid watermelon lines. Euphytica 2005, 145, 259–268. [Google Scholar] [CrossRef]
- Bagheri, M.; Mansouri, H. Effect of induced polyploidy on some biochemical parameters in Cannabis sativa L. Appl. Biochem. Biotech. 2015, 175, 2366–2375. [Google Scholar] [CrossRef] [PubMed]
- Barratt, D.P.; Derbyshire, P.; Findlay, K.; Pike, M.; Wellner, N.; Lunn, J.; Feil, R.; Simpson, C.; Maule, A.J.; Smith, A.M. Normal growth of Arabidopsis requires cytosolic invertase but not sucrose synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 13124–13129. [Google Scholar] [CrossRef]
- Rennie, E.A.; Turgeon, R. A comprehensive picture of phloem loading strategies. Proc. Natl. Acad. Sci. USA 2009, 106, 14162–14167. [Google Scholar] [CrossRef]
- Xu, S.-M.; Brill, E.; Llewellyn, D.J.; Furbank, R.T.; Ruan, Y.-L. Overexpression of a potato sucrose synthase gene in cotton accelerates leaf expansion, reduces seed abortion, and enhances fiber production. Mol. Plant. 2012, 5, 430–441. [Google Scholar] [CrossRef]
- Gupta, N.; Zargar, S.M.; Salgotra, R.; Dar, T.A. Identification of drought stress-responsive proteins in common bean. J. Proteins Proteom. 2019, 10, 45–53. [Google Scholar] [CrossRef]
- Bojarová, P.; Křen, V. Glycosidases: A key to tailored carbohydrates. Trends Biotechnol. 2009, 27, 199–209. [Google Scholar] [CrossRef]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial malate dehydrogenase lowers leaf respiration and alters photorespiration and plant growth in Arabidopsis. Plant. Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef] [PubMed]
- Hanse, E.A.; Ruan, C.; Kachman, M.; Wang, D.; Lowman, X.H.; Kelekar, A. Cytosolic malate dehydrogenase activity helps support glycolysis in actively proliferating cells and cancer. Oncogene 2017, 36, 3915–3924. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Carrari, F.; Lytovchenko, A.; Smith, A.M.; Loureiro, M.E.; Ratcliffe, R.G.; Sweetlove, L.J.; Fernie, A.R. Enhanced photosynthetic performance and growth as a consequence of decreasing mitochondrial malate dehydrogenase activity in transgenic tomato plants. Plant. Physiol. 2005, 137, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Siddappaji, M.H.; Scholes, D.R.; Bohn, M.; Paige, K.N. Overcompensation in response to herbivory in Arabidopsis thaliana: The role of glucose-6-phosphate dehydrogenase and the oxidative pentose-phosphate pathway. Genetics 2013, 195, 589–598. [Google Scholar] [CrossRef]
- Agapito-Tenfen, S.Z.; Guerra, M.P.; Wikmark, O.-G.; Nodari, R.O. Comparative proteomic analysis of genetically modified maize grown under different agroecosystems conditions in Brazil. Proteome Sci. 2013, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino acid catabolism in plants. Mol. Plant. 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Czub, B.; Shah, A.Z.; Alfano, G.; Kruczek, P.M.; Chakarova, C.F.; Bhattacharya, S.S. TOPORS, a dual E3 ubiquitin and sumo1 ligase, interacts with 26 S protease regulatory subunit 4, encoded by the PSMC1 gene. PLoS ONE 2016, 11, e0148678. [Google Scholar] [CrossRef]
- Natesh, R.; Clare, D.K.; Farr, G.W.; Horwich, A.L.; Saibil, H.R. A two-domain folding intermediate of RuBisCO in complex with the GroEL chaperonin. Int. J. Biol. Macromol. 2018, 118, 671–675. [Google Scholar] [CrossRef]
- Aigner, H.; Wilson, R.; Bracher, A.; Calisse, L.; Bhat, J.; Hartl, F.; Hayer-Hartl, M. Plant RuBisCo assembly in E. coli with five chloroplast chaperones including BSD2. Science 2017, 358, 1272–1278. [Google Scholar] [CrossRef]
- Sjögren, L.L.; MacDonald, T.M.; Sutinen, S.; Clarke, A.K. Inactivation of the clpC1 gene encoding a chloroplast Hsp100 molecular chaperone causes growth retardation, leaf chlorosis, lower photosynthetic activity, and a specific reduction in photosystem content. Plant. Physiol. 2004, 136, 4114–4126. [Google Scholar] [CrossRef]
- Hayer-Hartl, M. From chaperonins to Rubisco assembly and metabolic repair. Protein Sci. 2017, 26, 2324–2333. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Poovaiah, B. Arabidopsis chloroplast chaperonin 10 is a calmodulin-binding protein. Biochem. Biophys. Res. Commun. 2000, 275, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Li, Y.; Chen, L.S. Magnesium deficiency–induced impairment of photosynthesis in leaves of fruiting Citrus reticulata trees accompanied by up-regulation of antioxidant metabolism to avoid photo-oxidative damage. J. Plant. Nutr. Soil Sci. 2012, 175, 784–793. [Google Scholar] [CrossRef]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhi, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant. 2016, 38, 145. [Google Scholar] [CrossRef]
- Cui, T.Z.; Kawamukai, M. Coq10, a mitochondrial coenzyme Q binding protein, is required for proper respiration in Schizosaccharomyces pombe. FEBS J. 2009, 276, 748–759. [Google Scholar] [CrossRef]
- Amthor, J.S.; Baldocchi, D.D. Terrestrial higher plant respiration and net primary production. Terr. Global Prod. 2001, 33–59. [Google Scholar]
- Amthor, J. Higher plant respiration and its relationships to photosynthesis. In Ecophysiology of Photosynthesis; Springer: Berlin, Germany, 1995; pp. 71–101. [Google Scholar]
- Morris, S.E.; Cox, M.C.; Ross, J.J.; Krisantini, S.; Beveridge, C.A. Auxin dynamics after decapitation are not correlated with the initial growth of axillary buds. Plant. Physiol. 2005, 138, 1665–1672. [Google Scholar] [CrossRef]
- Mason, M.G.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar demand, not auxin, is the initial regulator of apical dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef]
- Bertheloot, J.; Barbier, F.; Boudon, F.; Perez-Garcia, M.D.; Péron, T.; Citerne, S.; Dun, E.; Beveridge, C.; Godin, C.; Sakr, S. Sugar availability suppresses the auxin-induced strigolactone pathway to promote bud outgrowth. New Phytol. 2020, 225, 866–879. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, V.P.; Gupta, N. Efficacy of putrescine and benzyladenine on photosynthesis and productivity in relation to drought tolerance in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Pla. 2012, 18, 331–336. [Google Scholar] [CrossRef]
- Youssef, A.; Mady, M. Influence of light intensity and benzyladenine on growth performance of Aspidistra elatior Blume plant. Res. J. Agric. Biol. Sci. 2013, 9, 248–257. [Google Scholar]
- Bianco, L.; Lopez, L.; Scalone, A.G.; Di Carli, M.; Desiderio, A.; Benvenuto, E.; Perrotta, G. Strawberry proteome characterization and its regulation during fruit ripening and in different genotypes. J. Proteom. 2009, 72, 586–607. [Google Scholar] [CrossRef] [PubMed]
- Görg, A.; Obermaier, C.; Boguth, G.; Harder, A.; Scheibe, B.; Wildgruber, R.; Weiss, W. The current state of two-dimensional electrophoresis with immobilized pH gradients. Electrophoresis 2000, 21, 1037–1053. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Lopez, S.; Maroto, J.; San Bautista, A.; Pascual, B.; Alagarda, J. Differences in carbohydrate content of waiting-bed strawberry plants during development in the nursery. Sci. Hortic. 2002, 94, 53–62. [Google Scholar] [CrossRef]
- Li, S.; Huang, P.; Ding, G.; Zhou, L.; Tang, P.; Sun, M.; Zheng, Y.; Lin, S. Optimization of hormone combinations for root growth and bud germination in Chinese fir (Cunninghamia lanceolata) clone leaf cuttings. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Temperature | Photoperiod | GA3 | BA |
|---|---|---|---|---|
| K1 | 6.00 | 2.67 | 18.00 | 5.00 |
| K2 | 14.00 | 10.33 | 7.33 | 15.00 |
| K3 | 10.67 | 17.67 | 5.33 | 10.67 |
| R | 8.00 | 15.00 | 12.67 | 10.00 |
| Optimal level | 25/15 °C | 16/8 h | 0 mg·L−1 | 50 mg·L−1 |
| Spot No. | Accession No. | Species | Protein Name | Score a | Theor./Exp. Mr (kDa) b | Theor./Exp. pI c | SC (%) d |
|---|---|---|---|---|---|---|---|
| Carbohydrate Metabolism | |||||||
| 3 | gi|470120564 | Fragaria vesca | Malate dehydrogenase, cytoplasmic-like | 59 | 36.0/40.7 | 6.01/6.35 | 23 |
| 8 | gi|470136472 | Fragaria vesca | Glucose-6-phosphate 1-Dehydrogenase, cytoplasmic isoform 2-like | 184 | 58.8/81.7 | 5.93/6.07 | 36 |
| 9 | gi|470109046 | Fragaria vesca | Sucrose synthase 2-like | 98 | 92.8/90.8 | 5.94/6.01 | 23 |
| 20 | gi|470148237 | Fragaria vesca | Glucan endo-1,3-beta-glucosidase | 157 | 16/31.9 | 5.33/5.58 | 32 |
| 25 | gi|470114187 | Fragaria vesca | Putative lactoylglutathione lyase-like isoform 2 | 114 | 32.6/31.0 | 5.28/5.33 | 33 |
| Proteometabolism | |||||||
| 6 | gi|470132774 | Fragaria vesca | Probable protein disulfide-isomerase A6-like | 81 | 40.1/47.2 | 6.12/6.02 | 30 |
| 7 | gi|470136937 | Fragaria vesca | Probable mitochondrial-processing peptidase subunit beta-like | 219 | 58.5/60.7 | 6.42/6.07 | 29 |
| 12 | gi|470114303 | Fragaria vesca | Fumarylacetoacetase-like | 69 | 45.8/45.3 | 5.57/5.89 | 32 |
| 23 | gi|470142843 | Fragaria vesca | 26S protease regulatory subunit 6B homolog | 99 | 46.2/55.6 | 5.48/5.54 | 26 |
| Photosynthesis | |||||||
| 13 | gi|7008071 | Baccharis halimifolia | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit | 68 | 54.5/57.5 | 6.29/5.92 | 20 |
| 14 | NES2_FRAAN | Fragaria ananassa | (3S,6E)-nerolidol synthase 2, chloroplastic/mitochondrial OS | 40 | 66.6/88.6 | 6.27/5.99 | 20 |
| 19 | gi|470122943 | Fragaria vesca | Chaperone protein ClpC, Chloroplastic-like | 93 | 102.4/94.1 | 6.59/5.63 | 27 |
| 24 | gi|237637006 | Linum tenue | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit | 94 | 51.2/89.2 | 6.23/5.46 | 17 |
| 28 | gi|470107388 | Fragaria vesca | Chaperonin CPN60-2, mitochondrial-like | 89 | 61.8/69.9 | 5.89/5.41 | 28 |
| Respiration | |||||||
| 10 | gi|470109651 | Fragaria vesca | Coenzyme Q-binding protein COQ10 homolog, mitochondrial-like | 45 | 28.6/31.5 | 8.85/5.92 | 24 |
| Signaling Proteins | |||||||
| 2 | gi|470103566 | Fragaria vesca | Annexin D1-like | 263 | 36.4/31.5 | 6.44/6.54 | 44 |
| 5 | gi|470114874 | Fragaria vesca | Transcription factor bHLH135-like | 53 | 10.4/12.2 | 5.07/6.02 | 44 |
| 16 | gi|571467514 | Glycine max | Probable calcium-binding protein CML45-like | 70 | 19.5/32.3 | 4.64/5.74 | 40 |
| Stress-Related Proteins | |||||||
| 1 | gi|470129560 | Fragaria vesca | Catalase-like | 87 | 57.1/59.7 | 6.58/6.74 | 33 |
| 4 | gi|470126676 | Fragaria vesca | Chaperone protein ClpB1-like | 85 | 101.8/100.5 | 5.91/6.21 | 21 |
| 31 | gi|470121677 | Fragaria vesca | Major allergen Pru ar 1-like | 36 | 17.7/22.9 | 5.39/5.51 | 44 |
| 32 | gi|470106694 | Fragaria vesca | Pathogenesis-related protein 1C-like | 70 | 18.1/34.2 | 5.1/5.05 | 24 |
| General Metabolism-Related Proteins | |||||||
| 11 | gi|470134340 | Fragaria vesca | Monodehydroascorbate reductase-like | 72 | 47.2/48.6 | 6.22/5.98 | 23 |
| 17 | gi|51493451 | Fragaria ananassa | Flavanone 3-hydroxylase | 307 | 41.4/43.1 | 5.61/5.72 | 53 |
| 29 | gi|470144168 | Fragaria vesca | Chalcone-flavonone isomerase-like | 95 | 23.6/25.4 | 4.86/5.16 | 46 |
| Treatment | Temperature (A, D/N, °C) | Photoperiod (B, D/N, h) | GA3 (C, mg·L−1) | 6-BA (D, mg·L−1) | ||||
|---|---|---|---|---|---|---|---|---|
| T1 | A1 | 23/17 | B1 | 8/16 | C1 | 0 | D1 | 0 |
| T2 | A1 | 23/17 | B2 | 12/12 | C2 | 50 | D2 | 50 |
| T3 | A1 | 23/17 | B3 | 16/8 | C3 | 100 | D3 | 100 |
| T4 | A2 | 25/15 | B1 | 8/16 | C2 | 50 | D3 | 100 |
| T5 | A2 | 25/15 | B2 | 12/12 | C3 | 100 | D1 | 0 |
| T6 | A2 | 25/15 | B3 | 16/8 | C1 | 0 | D2 | 50 |
| T7 | A3 | 27/13 | B1 | 8/16 | C3 | 100 | D2 | 50 |
| T8 | A3 | 27/13 | B2 | 12/12 | C1 | 0 | D3 | 100 |
| T9 | A3 | 27/13 | B3 | 16/8 | C2 | 50 | D1 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Hu, J.; Wei, H.; Jeong, B.R. A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry. Int. J. Mol. Sci. 2020, 21, 4917. https://doi.org/10.3390/ijms21144917
Li Y, Hu J, Wei H, Jeong BR. A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry. International Journal of Molecular Sciences. 2020; 21(14):4917. https://doi.org/10.3390/ijms21144917
Chicago/Turabian StyleLi, Yali, Jiangtao Hu, Hao Wei, and Byoung Ryong Jeong. 2020. "A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry" International Journal of Molecular Sciences 21, no. 14: 4917. https://doi.org/10.3390/ijms21144917
APA StyleLi, Y., Hu, J., Wei, H., & Jeong, B. R. (2020). A Long-Day Photoperiod and 6-Benzyladenine Promote Runner Formation through Upregulation of Soluble Sugar Content in Strawberry. International Journal of Molecular Sciences, 21(14), 4917. https://doi.org/10.3390/ijms21144917