Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa

 , , ,
, , , 
Abstract
1. Introduction
2. Results
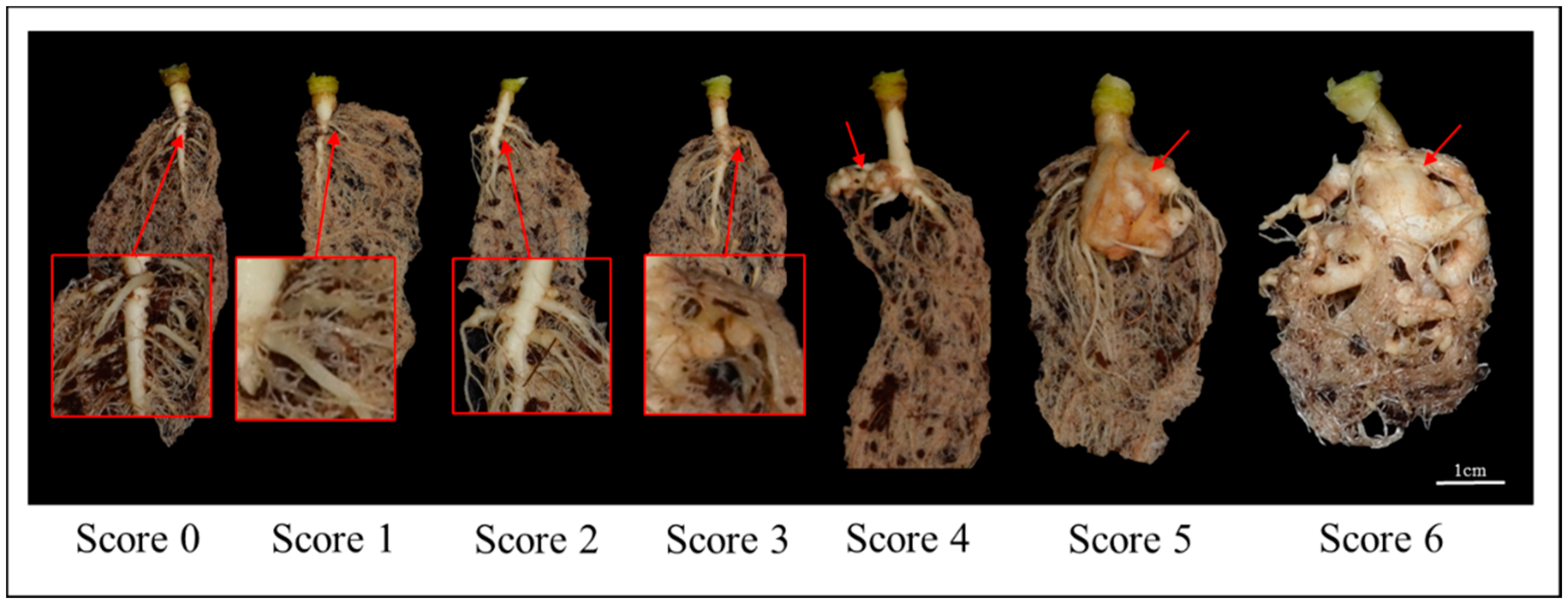
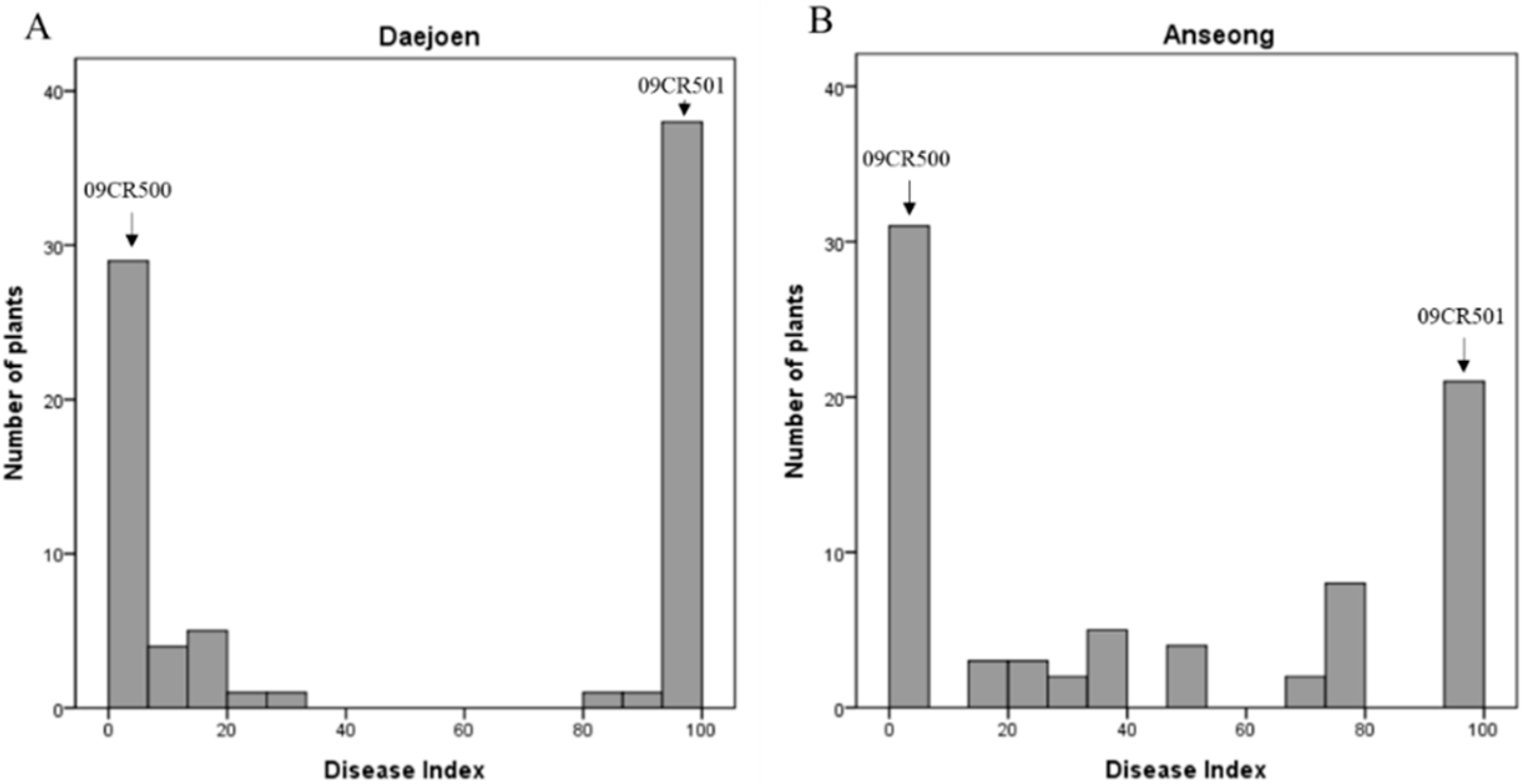
2.1. Inheritance of Clubroot Resistance in Line 09CR500
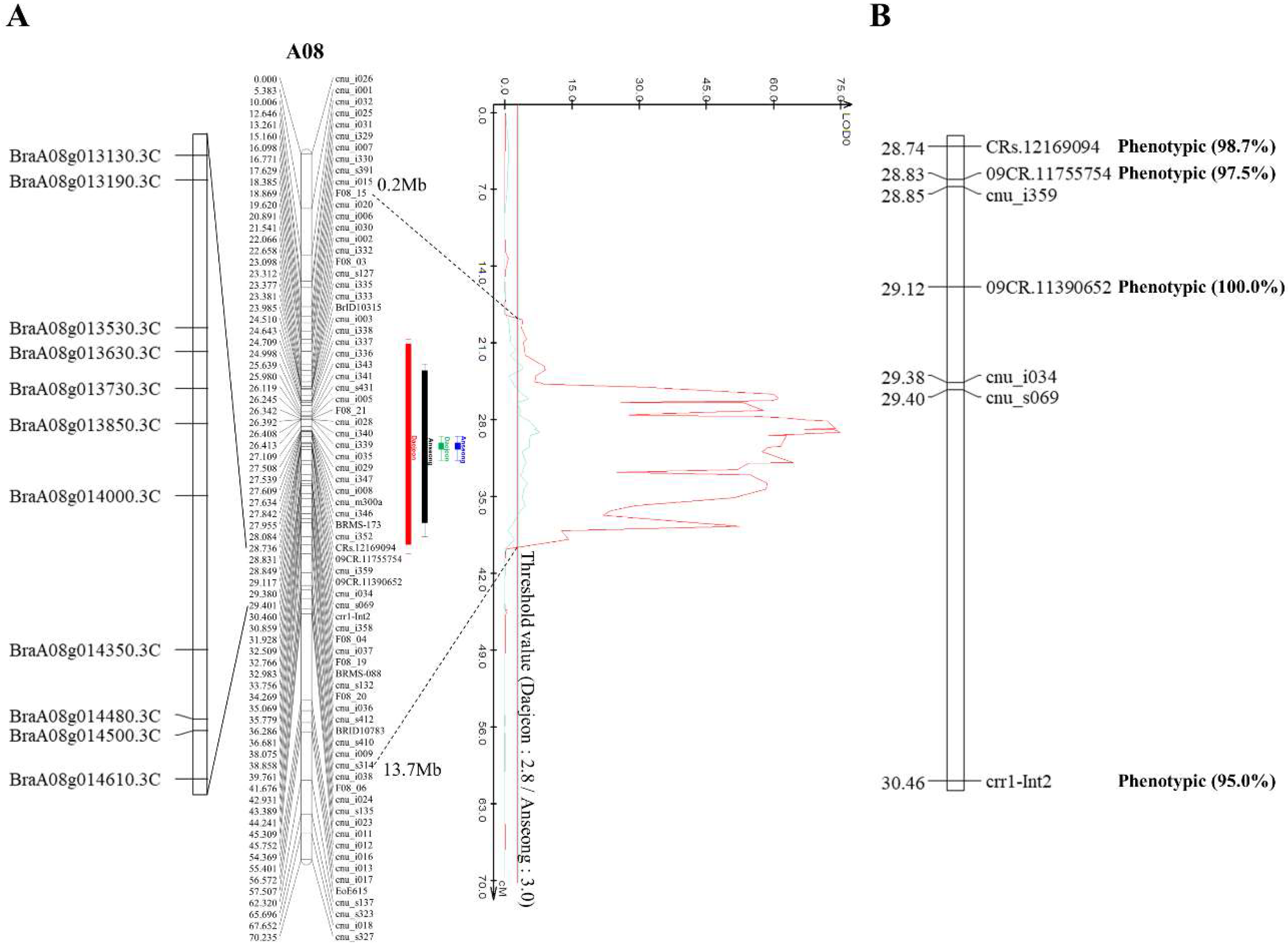
2.2. Whole-Genome Sequencing Analysis and Genetic Map Construction
2.3. QTL Identification and Marker Development
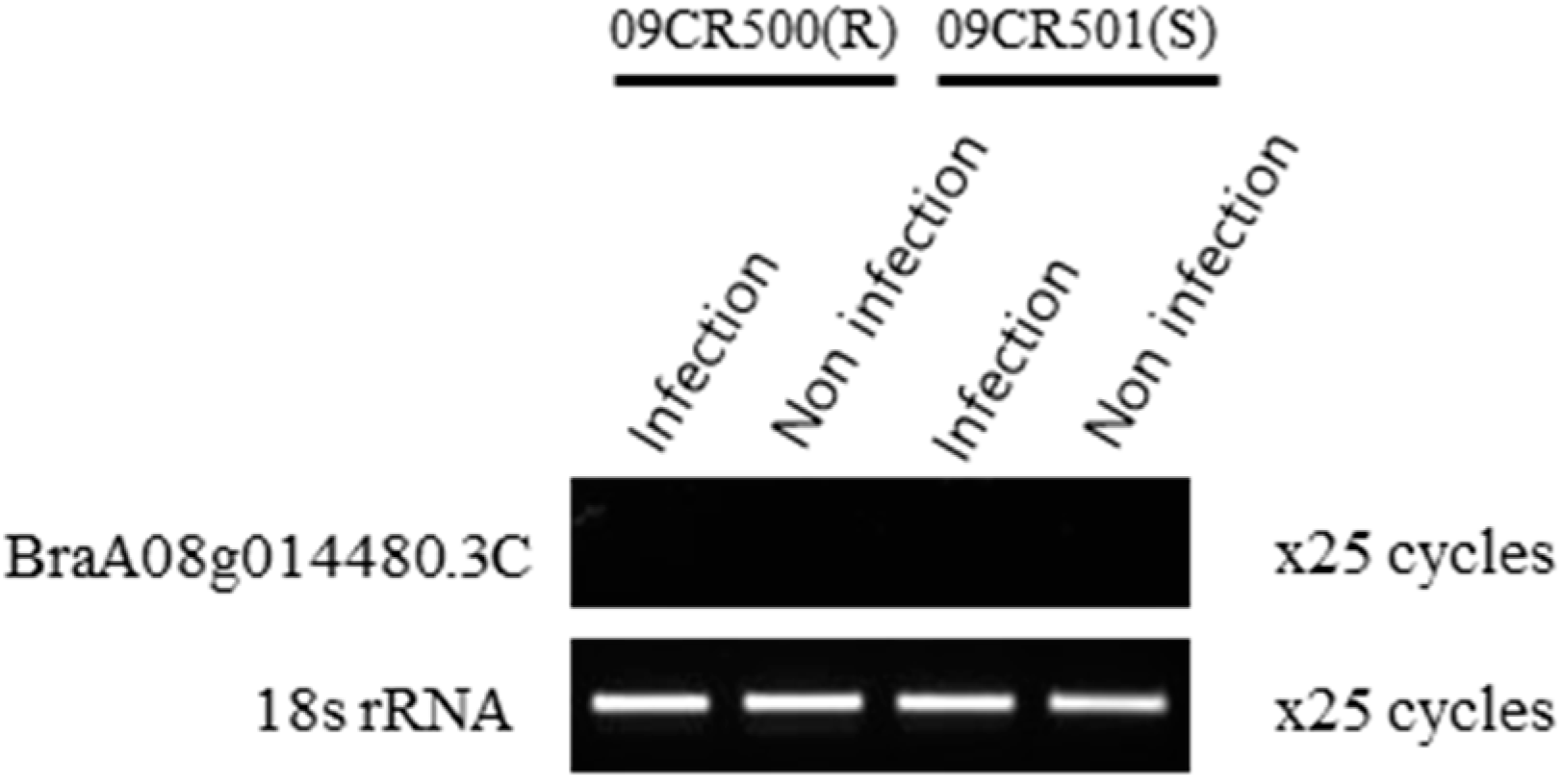
2.4. Validation of QTL Based on the Expression, High-Resolution Melting (HRM), and InDel Marker Analysis
3. Discussion
3.1. PbBrA08Banglim Is a Stable Large-Effect QTL
3.2. RT-PCR and HRM Analysis Established the Novelty of the PbBrA08Banglim Locus
4. Materials and Methods
4.1. Plant Materials
4.2. Pathogen Inoculation and Evaluation of Disease Symptoms
4.3. Marker Development and Genotyping
4.4. Genetic Map Construction and QTL Analysis
4.5. Annotation of Candidate Genes
4.6. Conversion of Physical Location to BRAD Version 3.0
4.7. Semiquantitative RT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| QTL | Quantitative trait loci |
| DH | Doubled haploid |
| LG | Linkage group |
| SNP | Single nucleotide polymorphism |
| InDel | Insertion and deletion |
| LOD | Logarithm of odds |
| CIM | Composite interval mapping |
| ICIM | Inclusive composite interval mapping |
| TIR | Toll interleukin-1 receptor |
| NBS | Nucleotide binding site |
| LRR | Leucine-rich repeat |
| RT-PCR | Real-time polymerase chain reaction |
References
- dos Santos Isaias, R.M.; de Oliveira, D.C.; da Silva Carneiro, R.G.; Kraus, J.E. Developmental anatomy of galls in the Neotropics: Arthropods stimuli versus host plant constraints. In Neotropical Insect Galls; Springer: Berlin, Germany, 2014; pp. 15–34. [Google Scholar]
- Walerowski, P.; Gündel, A.; Yahaya, N.; Truman, W.; Sobczak, M.; Olszak, M.; Rolfe, S.; Borisjuk, L.; Malinowski, R. Clubroot disease stimulates early steps of phloem differentiation and recruits SWEET sucrose transporters within developing galls. The Plant Cell 2018, 30, 3058–3073. [Google Scholar]
- Donald, C.; Porter, I. Integrated control of clubroot. J. Plant Growth Regul. 2009, 28, 289. [Google Scholar]
- Zhang, X.-Z.; Lee, S.-U.; Kim, J.-S.; Yoon, Y.-S.; Choi, G.-S.; Kim, H.-K.; Kim, B.-S. Control Efficacy of Flusulfamide GR on Chinese Cabbage Clubroot Caused by Plasmodiophora brassicae. Res. Plant Dis. 2005, 11, 43–47. [Google Scholar]
- Peng, G.; Lahlali, R.; Hwang, S.-F.; Pageau, D.; Hynes, R.K.; McDonald, M.R.; Gossen, B.D.; Strelkov, S.E. Crop rotation, cultivar resistance, and fungicides/biofungicides for managing clubroot (Plasmodiophora brassicae) on canola. Can. J. Plant Pathol. 2014, 36, 99–112. [Google Scholar]
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar]
- Diederichsen, E.; Frauen, M.; Linders, E.G.; Hatakeyama, K.; Hirai, M. Status and perspectives of clubroot resistance breeding in crucifer crops. J. Plant Growth Regul. 2009, 28, 265–281. [Google Scholar]
- Kulwal, P.; Thudi, M.; Varshney, R. Crop breeding for sustainable agriculture, genomics interventions in. In Sustainable Food Production; Springer: New York, NY, USA, 2013. [Google Scholar]
- Diederichsen, E.; Sacristan, M. Disease response of resynthesized Brassica napus L. lines carrying different combinations of resistance to Plasmodiophora brassicae Wor. Plant Breed. 1996, 115, 5–10. [Google Scholar]
- Neik, T.X.; Barbetti, M.J.; Batley, J. Current status and challenges in identifying disease resistance genes in Brassica napus. Front. Plant Sci. 2017, 8, 1788. [Google Scholar]
- Diederichsen, E.; Beckmann, J.; Schondelmeier, J.; Dreyer, F. Genetics of clubroot resistance in Brassica napus ’Mendel’. Acta Hortic. 2006, 307–311. [Google Scholar]
- Zhang, H.; Feng, J.; Hwang, S.F.; Strelkov, S.; Falak, I.; Huang, X.; Sun, R. Mapping of clubroot (Plasmodiophora brassicae) resistance in canola (Brassica napus). Plant Pathol. 2016, 65, 435–440. [Google Scholar]
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of novel QTLs for isolate-specific partial resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307. [Google Scholar]
- Crute, I.; Phelps, K.; Barnes, A.; Buczacki, S.; Crisp, P. The relationship between genotypes of three Brassica species and collections of Plasmodiophora brassicae. Plant Pathol. 1983, 32, 405–420. [Google Scholar]
- Crisp, P.; Crute, I.; Sutherland, R.; Angell, S.; Bloor, K.; Burgess, H.; Gordon, P. The exploitation of genetic resources of Brassica oleracea in breeding for resistance to clubroot (Plasmodiophora brassicae). Euphytica 1989, 42, 215–226. [Google Scholar]
- Matsumoto, E.; Yasui, C.; Ohi, M.; Tsukada, M. Linkage analysis of RFLP markers for clubroot resistance and pigmentation in Chinese cabbage (Brassica rapa ssp. pekinensis). Euphytica 1998, 104, 79. [Google Scholar]
- Piao, Z.; Deng, Y.; Choi, S.; Park, Y.; Lim, Y. SCAR and CAPS mapping of CRb, a gene conferring resistance to Plasmodiophora brassicae in Chinese cabbage (Brassica rapa ssp. pekinensis). Theor. Appl. Genet. 2004, 108, 1458–1465. [Google Scholar]
- Kato, T.; Hatakeyama, K.; Fukino, N.; Matsumoto, S. Fine mapping of the clubroot resistance gene CRb and development of a useful selectable marker in Brassica rapa. Breed. Sci. 2013, 63, 116–124. [Google Scholar]
- Sakamoto, K.; Saito, A.; Hayashida, N.; Taguchi, G.; Matsumoto, E. Mapping of isolate-specific QTLs for clubroot resistance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Theor. Appl. Genet. 2008, 117, 759–767. [Google Scholar]
- Pang, W.; Fu, P.; Li, X.; Zhan, Z.; Yu, S.; Piao, Z. Identification and mapping of the Clubroot resistance gene CRd in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Front. Plant Sci. 2018, 9, 653. [Google Scholar]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Kondo, M.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Hirai, M.; Matsumoto, S. Simple sequence repeat-based comparative genomics between Brassica rapa and Arabidopsis thaliana: The genetic origin of clubroot resistance. Genetics 2006, 173, 309–319. [Google Scholar]
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar]
- Hirai, M.; Harada, T.; Kubo, N.; Tsukada, M.; Suwabe, K.; Matsumoto, S. A novel locus for clubroot resistance in Brassica rapa and its linkage markers. Theor. Appl. Genet. 2004, 108, 639–643. [Google Scholar]
- Nguyen, M.; Monakhos, G.; Komakhin, R.; Monakhos, S. The new Clubroot resistance locus is located on chromosome A05 in Chinese cabbage (Brassica rapa L.). Russ. J. Genet. 2018, 54, 296–304. [Google Scholar]
- Chu, M.; Song, T.; Falk, K.C.; Zhang, X.; Liu, X.; Chang, A.; Lahlali, R.; McGregor, L.; Gossen, B.D.; Yu, F. Fine mapping of Rcr1 and analyses of its effect on transcriptome patterns during infection by Plasmodiophora brassicae. BMC Genom. 2014, 15, 1166. [Google Scholar]
- Yu, F.; Zhang, X.; Peng, G.; Falk, K.C.; Strelkov, S.E.; Gossen, B.D. Genotyping-by-sequencing reveals three QTL for clubroot resistance to six pathotypes of Plasmodiophora brassicae in Brassica rapa. Sci. Rep. 2017, 7, 4516. [Google Scholar]
- Laila, R.; Park, J.I.; Robin, A.H.K.; Natarajan, S.; Vijayakumar, H.; Shirasawa, K.; Isobe, S.; Kim, H.T.; Nou, I.S. Mapping of a novel clubroot resistance QTL using ddRAD-seq in Chinese cabbage (Brassica rapa L.). BMC Plant Biol. 2019, 19, 13. [Google Scholar]
- Nagaoka, T.; Doullah, M.; Matsumoto, S.; Kawasaki, S.; Ishikawa, T.; Hori, H.; Okazaki, K. Identification of QTLs that control clubroot resistance in Brassica oleracea and comparative analysis of clubroot resistance genes between B. rapa and B. oleracea. Theor. Appl. Genet. 2010, 120, 1335–1346. [Google Scholar]
- Lee, J.; Izzah, N.K.; Choi, B.-S.; Joh, H.J.; Lee, S.-C.; Perumal, S.; Seo, J.; Ahn, K.; Jo, E.J.; Choi, G.J. Genotyping-by-sequencing map permits identification of clubroot resistance QTLs and revision of the reference genome assembly in cabbage (Brassica oleracea L.). DNA Res. 2016, 23, 29–41. [Google Scholar]
- Dakouri, A.; Zhang, X.; Peng, G.; Falk, K.C.; Gossen, B.D.; Strelkov, S.E.; Yu, F. Analysis of genome-wide variants through bulked segregant RNA sequencing reveals a major gene for resistance to Plasmodiophora brassicae in Brassica oleracea. Sci. Rep. 2018, 8, 1–10. [Google Scholar]
- Nomura, K.; Minegishi, Y.; Kimizuka-Takagi, C.; Fujioka, T.; Moriguchi, K.; Shishido, R.; Ikehashi, H. Evaluation of F2 and F3 plants introgressed with QTLs for clubroot resistance in cabbage developed by using SCAR markers. Plant Breed. 2005, 124, 371–375. [Google Scholar]
- Peng, L.; Zhou, L.; Li, Q.; Wei, D.; Ren, X.; Song, H.; Mei, J.; Si, J.; Qian, W. Identification of quantitative trait loci for clubroot resistance in Brassica oleracea with the use of Brassica SNP microarray. Front. Plant Sci. 2018, 9, 822. [Google Scholar]
- Chang, A.; Lamara, M.; Wei, Y.; Hu, H.; Parkin, I.A.; Gossen, B.D.; Peng, G.; Yu, F. Clubroot resistance gene Rcr6 in Brassica nigra resides in a genomic region homologous to chromosome A08 in B. rapa. BMC Plant Biol. 2019, 19, 224. [Google Scholar]
- Manzanares-Dauleux, M.; Delourme, R.; Baron, F.; Thomas, G. Mapping of one major gene and of QTLs involved in resistance to clubroot in Brassica napus. Theor. Appl. Genet. 2000, 101, 885–891. [Google Scholar]
- Hejna, O.; Havlickova, L.; He, Z.; Bancroft, I.; Curn, V. Analysing the genetic architecture of clubroot resistance variation in Brassica napus by associative transcriptomics. Mol. Breed. 2019, 39, 112. [Google Scholar]
- Werner, S.; Diederichsen, E.; Frauen, M.; Schondelmaier, J.; Jung, C. Genetic mapping of clubroot resistance genes in oilseed rape. Theor. Appl. Genet. 2008, 116, 363–372. [Google Scholar]
- Li, L.; Luo, Y.; Chen, B.; Xu, K.; Zhang, F.; Li, H.; Huang, Q.; Xiao, X.; Zhang, T.; Hu, J. A genome-wide association study reveals new loci for resistance to clubroot disease in Brassica napus. Front. Plant Sci. 2016, 7, 1483. [Google Scholar]
- Kamei, A.; Tsuro, M.; Kubo, N.; Hayashi, T.; Wang, N.; Fujimura, T.; Hirai, M. QTL mapping of clubroot resistance in radish (Raphanus sativus L.). Theor. Appl. Genet. 2010, 120, 1021–1027. [Google Scholar]
- Gan, C.; Deng, X.; Cui, L.; Yu, X.; Yuan, W.; Dai, Z.; Yao, M.; Pang, W.; Ma, Y.; Yu, X. Construction of a high-density genetic linkage map and identification of quantitative trait loci associated with clubroot resistance in radish (Raphanus sativus L.). Mol. Breed. 2019, 39, 116. [Google Scholar]
- Varshney, R.K. Exciting journey of 10 years from genomes to fields and markets: Some success stories of genomics-assisted breeding in chickpea, pigeonpea and groundnut. Plant Sci. 2016, 242, 98–107. [Google Scholar]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar]
- Bolger, M.E.; Weisshaar, B.; Scholz, U.; Stein, N.; Usadel, B.; Mayer, K.F. Plant genome sequencing—applications for crop improvement. Curr. Opin. Biotech. 2014, 26, 31–37. [Google Scholar]
- Yang, Z.; Huang, D.; Tang, W.; Zheng, Y.; Liang, K.; Cutler, A.J.; Wu, W. Mapping of quantitative trait loci underlying cold tolerance in rice seedlings via high-throughput sequencing of pooled extremes. PLoS ONE 2013, 8, e68433. [Google Scholar]
- Zhu, J.; Chen, J.; Gao, F.; Xu, C.; Wu, H.; Chen, K.; Si, Z.; Yan, H.; Zhang, T. Rapid mapping and cloning of the virescent-1 gene in cotton by bulked segregant analysis–next generation sequencing and virus-induced gene silencing strategies. J. Exp. Bot. 2017, 68, 4125–4135. [Google Scholar]
- Williams, P.H. A system for the determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga. Phytopathology 1966, 56, 624–626. [Google Scholar]
- Dobson, R. Pathotypes of Plasmodiophora brassicae in Washington, Oregon, and California. Plant Dis. 1983, 67, 269. [Google Scholar] [CrossRef]
- Kim, H.; Jo, E.J.; Choi, Y.H.; Jang, K.S.; Choi, G.J. Pathotype classification of Plasmodiophora brassicae isolates using Clubroot-resistant cultivars of Chinese cabbage. Plant Pathol. J. 2016, 32, 423–430. [Google Scholar]
- Pang, W.; Li, X.; Choi, S.R.; Dhandapani, V.; Im, S.; Park, M.Y.; Jang, C.S.; Yang, M.S.; Ham, I.K.; Lee, E.M.; et al. Development of a leafy Brassica rapa fixed line collection for genetic diversity and population structure analysis. Mol. Breed. 2015, 35, 54. [Google Scholar]
- Choi, S.R.; Oh, S.H.; Dhandapani, V.; Jang, C.S.; Ahn, C.-H.; Rameneni, J.J.; Kim, H.; Jeon, I.; Lim, Y.P. Development of SNP markers for marker-assisted breeding in Chinese cabbage using Fluidigm genotyping assays. Hortic. Environ. Biote. 2020, 61, 327–338. [Google Scholar] [CrossRef]
- Kim, H.; Choi, S.R.; Bae, J.; Hong, C.P.; Lee, S.Y.; Hossain, M.J.; Van Nguyen, D.; Jin, M.; Park, B.-S.; Bang, J.-W. Sequenced BAC anchored reference genetic map that reconciles the ten individual chromosomes of Brassica rapa. BMC genomics 2009, 10, 432. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods. 2012, 9, 357. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed]
- Marcotty, M.; Ledgard, H.F. The World of Programming Languages; Springer: Berlin, German, 1986; p. 41. [Google Scholar]
- De Vicente, M.; Metz, T.; Alercia, A. Descriptors for Genetic Markers Technologies-Version 1.0, February 2004; Bioversity International: Rome, Italy, 2004. [Google Scholar]
- Li, X.; Ramchiary, N.; Dhandapani, V.; Choi, S.R.; Hur, Y.; Nou, I.S.; Yoon, M.K.; Lim, Y.P. Quantitative trait loci mapping in Brassica rapa revealed the structural and functional conservation of genetic loci governing morphological and yield component traits in the A, B, and C subgenomes of Brassica species. DNA Res. 2013, 20, 1–16. [Google Scholar] [CrossRef]
- Matsumoto, E.; Ueno, H.; Aruga, D.; Sakamoto, K.; Hayashida, N. Accumulation of three Clubroot resistance genes through marker-assisted selection in Chinese cabbage (Brassica rapa ssp. pekinensis). J. Jpn. Soc. Hortic. Sci. 2012, 81, 184–190. [Google Scholar] [CrossRef]
- Suwabe, K.; Suzuki, G.; Nunome, T.; Hatakeyama, K.; Mukai, Y.; Fukuoka, H.; Matsumoto, S. Microstructure of a Brassica rapa genome segment homoeologous to the resistance gene cluster on Arabidopsis chromosome 4. Breed. Sci. 2012, 62, 170–177. [Google Scholar] [CrossRef]
- Ueno, H.; Matsumoto, E.; Aruga, D.; Kitagawa, S.; Matsumura, H.; Hayashida, N. Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol. Biol. 2012, 80, 621–629. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Z.; Zhang, C.; Pang, W.; Choi, S.R.; Lim, Y.P.; Piao, Z. Fine genetic and physical mapping of the CRb gene conferring resistance to clubroot disease in Brassica rapa. Mol. Breed. 2014, 34, 1173–1183. [Google Scholar] [CrossRef]
- Saito, M.; Kubo, N.; Matsumoto, S.; Suwabe, K.; Tsukada, M.; Hirai, M. Fine mapping of the clubroot resistance gene, Crr3, in Brassica rapa. Theor. Appl. Genet. 2006, 114, 81. [Google Scholar] [CrossRef]
- Stam, P. Construction of integrated genetic linkage maps by means of a new computer package: Join Map. Plant J. 1993, 3, 739–744. [Google Scholar] [CrossRef]
- Van Ooijen, J.W. Joinmap® 4.1 Software for the Calculation of Genetic Linkage Maps in Experimental Populations of Diploid Species; Kyazma B.V.: Wageningen, The Netherlands, 2006. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann Eugen 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2012. Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 10 April 2020).
- Doerge, R.W.; Churchill, G.A. Permutation tests for multiple loci affecting a quantitative character. Genetics 1996, 142, 285–294. [Google Scholar]
- Li, H.; Ye, G.; Wang, J. A modified algorithm for the improvement of composite interval mapping. Genetics 2007, 175, 361–374. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef]
- Qi, J.N.; Yu, S.C.; Zhang, F.L.; Shen, X.Q.; Zhao, X.Y.; Yu, Y.J.; Zhang, D.S. Reference gene selection for real-time quantitative polymerase chain reaction of mRNA transcript levels in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Plant Mol. Biol. Rep. 2010, 28, 597–604. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
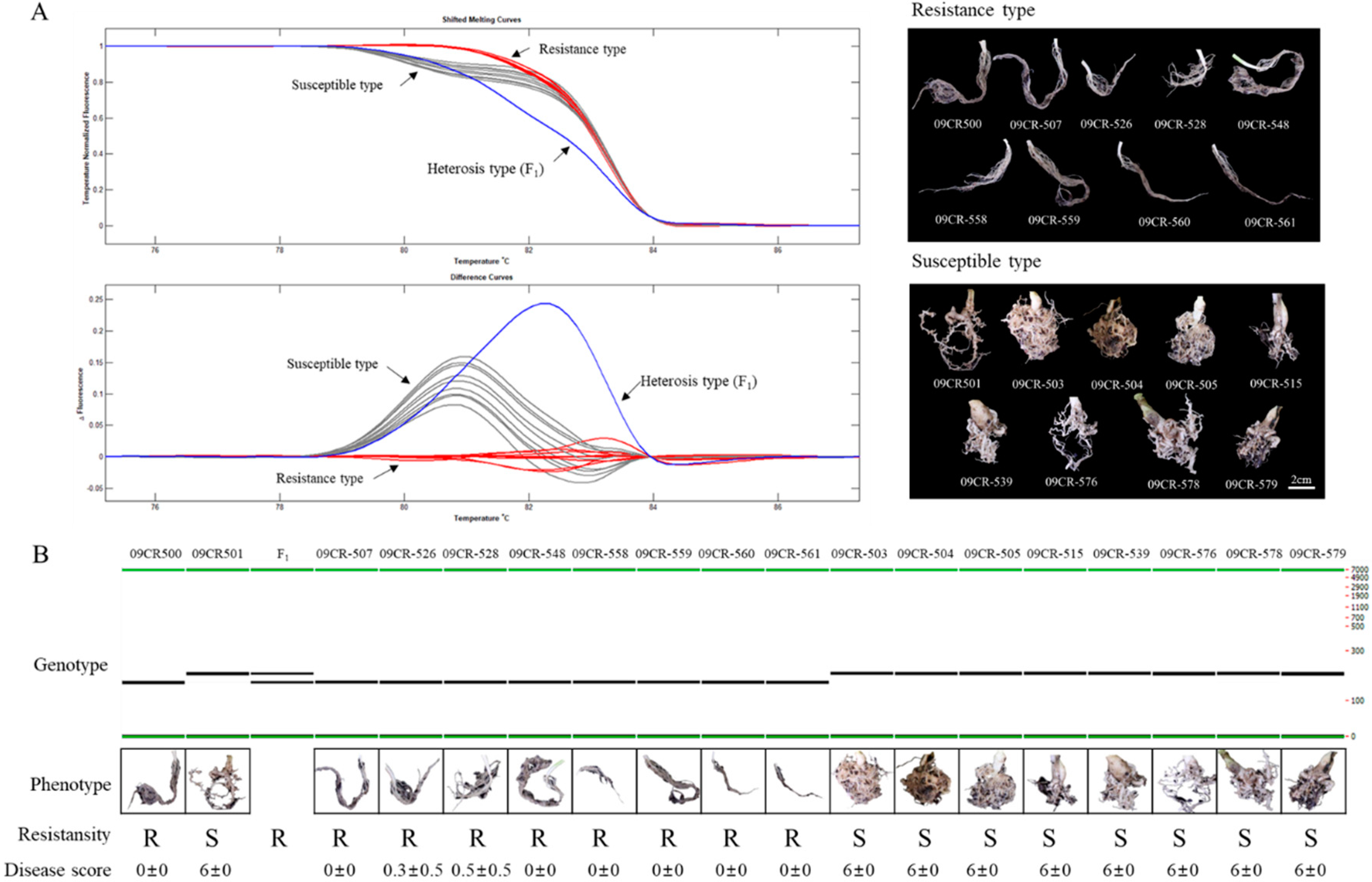{kind=link}
| Location | Plant Materials | Total No. | Phenotype of | Expected Rtio (R:S) | χ2 | ||
|---|---|---|---|---|---|---|---|
| Resistance (R) | Susceptible (S) | Missing (-) | |||||
| Daejoen | 09CR500 | 10 | 10 | 0 | 0 | - | - |
| 09CR501 | 10 | 0 | 10 | 0 | - | - | |
| DH population | 81 | 40 | 40 | 1 | 1:1 | 0.01 | |
| Anseong | 09CR500 | 10 | 10 | 0 | 0 | - | - |
| 09CR501 | 10 | 0 | 10 | 0 | - | - | |
| DH population | 81 | 39 | 40 | 2 | 1:1 | 0.06 | |
| Loci Name | Location | Chr No. | Closest Marker | LOD | R2 (%) | Additive |
|---|---|---|---|---|---|---|
| PbBrA08Banglim-1 | Daejeon | 8 | 09CR.11390652 | 74.8 | 97.1 | −47.0 |
| PbBrA08Banglim-2 | Anseong | 8 | 09CR.11390652 | 7.9 | 26.0 | −26.1 |
| Gene ID | Gene Position (bp) a | |||
|---|---|---|---|---|
| v1.2 | v3.0 | Start | End | Protein Description b |
| Bra020979 | BraA08g013130.3C | 11,388,577 | 11,391,460 | Receptor-like protein/Leucine rich repeat |
| Bra020974 | BraA08g013190.3C | 11,427,768 | 11,428,159 | LRR receptor-like serine/ threonine-protein kinase |
| Bra020945/ Bra020942 | BraA08g013530.3C | 11,658,233 | 11,659,423 | F-box/kelch-repeat protein |
| Bra020936 | BraA08g013630.3C | 11,696,216 | 11,696,785 | TIR domain containing disease resistance protein |
| Bra020928 | BraA08g013730.3C | 11,752,980 | 11,756,408 | Leucine-rich repeat receptor-like serine/threonine-protein kinase |
| Bra020918 | BraA08g013850.3C | 11,809,250 | 11,815,278 | LRR receptor-like serine/ threonine-protein kinase |
| Bra020901/ Bra020902 | BraA08g014000.3C | 11,921,301 | 11,924,248 | F-box protein |
| Bra020876 | BraA08g014350.3C | 12,162,113 | 12,169,372 | LRR receptor-like serine/ threonine-protein kinase |
| Bra020861 | BraA08g014480.3C | 12,271,553 | 12,276,276 | Disease resistance protein |
| Bra020860 | BraA08g014500.3C | 12,289,217 | 12,290,401 | F-box protein |
| Bra020847 | BraA08g014610.3C | 12,365,097 | 12,366,311 | F-box protein |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.R.; Oh, S.H.; Chhapekar, S.S.; Dhandapani, V.; Lee, C.Y.; Rameneni, J.J.; Ma, Y.; Choi, G.J.; Lee, S.-S.; Lim, Y.P. Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa. Int. J. Mol. Sci. 2020, 21, 4157. https://doi.org/10.3390/ijms21114157
Choi SR, Oh SH, Chhapekar SS, Dhandapani V, Lee CY, Rameneni JJ, Ma Y, Choi GJ, Lee S-S, Lim YP. Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa. International Journal of Molecular Sciences. 2020; 21(11):4157. https://doi.org/10.3390/ijms21114157
Chicago/Turabian StyleChoi, Su Ryun, Sang Heon Oh, Sushil Satish Chhapekar, Vignesh Dhandapani, Chang Yeol Lee, Jana Jeevan Rameneni, Yinbo Ma, Gyung Ja Choi, Soo-Seong Lee, and Yong Pyo Lim. 2020. "Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa" International Journal of Molecular Sciences 21, no. 11: 4157. https://doi.org/10.3390/ijms21114157
APA StyleChoi, S. R., Oh, S. H., Chhapekar, S. S., Dhandapani, V., Lee, C. Y., Rameneni, J. J., Ma, Y., Choi, G. J., Lee, S.-S., & Lim, Y. P. (2020). Quantitative Trait Locus Mapping of Clubroot Resistance and Plasmodiophora brassicae Pathotype Banglim-Specific Marker Development in Brassica rapa. International Journal of Molecular Sciences, 21(11), 4157. https://doi.org/10.3390/ijms21114157