Beneficial Effects of Inflammatory Cytokine-Targeting Aptamers in an Animal Model of Chronic Prostatitis

 ,
,
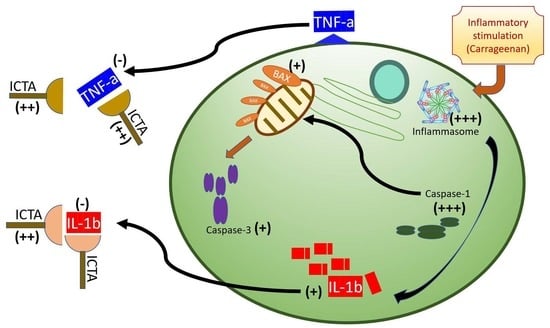
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. AptIL-1b Binds to IL-1b With High Affinity and Targets IL-1b In Vitro
2.2. ICTA Attenuated Local Carrageenan-Induced Mechanical Allodynia and Tactile Hyperalgesia
2.3. ICTA Modified Local Carrageenan-Induced Glandular Hyperplasia and Inflammatory Responses in the Prostate
2.4. ICTA Promoted Local Carrageenan-Induced Apoptosis in the Prostate
3. Discussion
3.1. Severity of Clinical Symptoms is Related to the Inflammasome Pathway
3.2. Inflammasome is Important in CP/CPPS
3.3. Cytokines Responsible for Inflammation
3.4. Besides Cytokines, Inflammasome is Also a Good Target for Treating Inflammation
3.5. Aptamers Were Used to Detect and Treat Inflammation-Related Diseases
3.6. Apoptosis in Inflammation
3.7. Concomitant Neutralization of Cytokines
3.8. Potential Problems and Research Topics With ICTA Administration
4. Materials and Methods
4.1. Oligonucleotides
4.2. Construction of IL-1b-Targeting Aptamer
4.3. Antibodies
4.4. Animals
4.5. Animal Treatment
4.6. Von Frey Filament Test
4.7. Immunohistochemistry
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IL-1b | Interleukin-1 beta |
| TNF-a | Tumor necrosis factor-alpha |
| ICTA | Inflammatory cytokine-targeting aptamers |
| CP | Chronic prostatitis |
| CPPS | Chronic pelvic pain syndrome |
| NALP | Nucleotide-binding oligomerization domain, Leucine rich Repeat and Pyrin domain containing |
| Casp3 | Caspase-3 |
| AptIL-1b | Interleukin-1 beta-targeting aptamer |
| AptTNF-a | Tumor necrosis factor-alpha-targeting aptamer |
| BAX | B-cell lymphoma 2-associated X protein |
| IFN-r | Interferon-gamma |
| ASC | Apoptosis-associated Speck-Like Protein Containing a Caspase Recruitment Domain |
| SD rat | Sprague Dawley rat |
References
- Batstone, G.R.; Doble, A. Chronic prostatitis. Curr. Opin. Urol. 2003, 13, 23–29. [Google Scholar] [PubMed]
- Dimitrakov, J.; Joffe, H.V.; Soldin, S.J.; Bolus, R.; Buffington, C.A.; Nickel, J.C. Adrenocortical hormone abnormalities in men with chronic prostatitis/chronic pelvic pain syndrome. Urology 2008, 71, 261–266. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Theoharides, T.C.; Cochrane, D.E. Critical role of mast cells in inflammatory diseases and the effect of acute stress. J. Neuroimmunol. 2004, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.U.; Wise, D.; Sawyer, T.; Glowe, P.; Orenberg, E.K. 6-day intensive treatment protocol for refractory chronic prostatitis/chronic pelvic pain syndrome using myofascial release and paradoxical relaxation training. J. Urol. 2011, 185, 1294–1299. [Google Scholar] [CrossRef]
- Pontari, M.A. Chronic prostatitis/chronic pelvic pain syndrome in elderly men: Toward better understanding and treatment. Drugs Aging 2003, 20, 1111–1125. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shang, X.; Huang, Y. The effects of interleukin-10 and -8 in chronic prostatitis. Zhonghua Nan Ke Xue 2004, 10, 486–487. [Google Scholar] [PubMed]
- Nadler, R.B.; Koch, A.E.; Calhoun, E.A.; Campbell, P.L.; Pruden, D.L.; Bennett, C.L.; Yarnold, P.R.; Schaeffer, A.J. IL-1beta and TNF-alpha in prostatic secretions are indicators in the evaluation of men with chronic prostatitis. J. Urol. 2000, 164, 214–218. [Google Scholar] [CrossRef]
- Chen, C.S.; Chang, P.J.; Lin, W.Y.; Huang, Y.C.; Ho, D.R. Evidences of the inflammasome pathway in chronic prostatitis and chronic pelvic pain syndrome in an animal model. Prostate 2013, 73, 391–397. [Google Scholar] [CrossRef]
- Ponomareva, L.; Liu, H.; Duan, X.; Dickerson, E.; Shen, H.; Panchanathan, R.; Choubey, D. AIM2, an IFN-inducible cytosolic DNA sensor, in the development of benign prostate hyperplasia and prostate cancer. Mol. Cancer Res. 2013, 11, 1193–1202. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Chen, W.; Yao, X.; Xing, Y.; Wang, X.; Zhong, J.; Meng, G. Mycoplasma hyorhinis activates the NLRP3 inflammasome and promotes migration and invasion of gastric cancer cells. PLoS ONE 2013, 8, e77955. [Google Scholar] [CrossRef]
- Gu, N.Y.; Kim, J.H.; Han, I.H.; Im, S.J.; Seo, M.Y.; Chung, Y.H.; Ryu, J.S. Trichomonas vaginalis induces IL-1beta production in a human prostate epithelial cell line by activating the NLRP3 inflammasome via reactive oxygen species and potassium ion efflux. Prostate 2016, 76, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, F.; Li, W.W.; Stary, C.; Clark, J.D.; Xu, S.; Xiong, X. The inflammasome as a target for pain therapy. Br. J. Anaesth. 2016, 117, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.-Z.; Cao, Q.; Liu, C. Targeting NLRP3 Inflammasome in the Treatment of CNS Diseases. Front. Mol. Neurosci. 2018, 11, 320. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, X.; Liu, Y.; Xia, Y.; Chang, R.; Zhang, C. Inflammasome inhibitors: Promising therapeutic approaches against cancer. J. Hematol. Oncol. 2019, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Wolf, G.; Yirmiya, R.; Goshen, I.; Iverfeldt, K.; Holmlund, L.; Takeda, K.; Shavit, Y. Impairment of interleukin-1 (IL-1) signaling reduces basal pain sensitivity in mice: Genetic, pharmacological and developmental aspects. Pain 2003, 104, 471–480. [Google Scholar] [CrossRef]
- Ashok, A.; Keener, R.; Rubenstein, M.; Stookey, S.; Bajpai, S.; Hicks, J.; Alme, A.K.; Drake, C.G.; Zheng, Q.; Trabzonlu, L.; et al. Consequences of interleukin 1beta-triggered chronic inflammation in the mouse prostate gland: Altered architecture associated with prolonged CD4(+) infiltration mimics human proliferative inflammatory atrophy. Prostate 2019, 79, 732–745. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Simon, A.; van der Meer, J.W.M. Treating inflammation by blocking interleukin-1 in a broad spectrum of diseases. Nat. Rev. Drug Discov. 2012, 11, 633–652. [Google Scholar] [CrossRef]
- Schlesinger, N.; de Meulemeester, M.; Pikhlak, A.; Yücel, A.E.; Richard, D.; Murphy, V.; Arulmani, U.; Sallstig, P.; So, A. Canakinumab relieves symptoms of acute flares and improves health-related quality of life in patients with difficult-to-treat Gouty Arthritis by suppressing inflammation: Results of a randomized, dose-ranging study. Arthritis Res. Ther. 2011, 13, R53. [Google Scholar] [CrossRef]
- Everett, B.M.; Cornel, J.H.; Lainscak, M.; Anker, S.D.; Abbate, A.; Thuren, T.; Libby, P.; Glynn, R.J.; Ridker, P.M. Anti-Inflammatory Therapy With Canakinumab for the Prevention of Hospitalization for Heart Failure. Circulation 2019, 139, 1289–1299. [Google Scholar] [CrossRef]
- Lai, W.Y.; Wang, J.W.; Huang, B.T.; Lin, E.P.; Yang, P.C. A Novel TNF-alpha-Targeting Aptamer for TNF-alpha-Mediated Acute Lung Injury and Acute Liver Failure. Theranostics 2019, 9, 1741–1751. [Google Scholar] [CrossRef]
- Saperstein, S.; Chen, L.; Oakes, D.; Pryhuber, G.; Finkelstein, J. IL-1beta augments TNF-alpha-mediated inflammatory responses from lung epithelial cells. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2009, 29, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Pontari, M.A.; Ruggieri, M.R. Mechanisms in prostatitis/chronic pelvic pain syndrome. J. Urol. 2004, 172, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Luzzi, G.A. Chronic prostatitis and chronic pelvic pain in men: Aetiology, diagnosis and management. J. Eur. Acad. Dermatol. Venereol. 2002, 16, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Hua, V.N.; Williams, D.H.; Schaeffer, A.J. Role of bacteria in chronic prostatitis/chronic pelvic pain syndrome. Curr. Urol. Rep. 2005, 6, 300–306. [Google Scholar] [CrossRef]
- Pontari, M.A. Etiology of chronic prostatitis/chronic pelvic pain syndrome: Psychoimmunoneurendocrine dysfunction (PINE syndrome) or just a really bad infection? World J. Urol. 2013, 31, 725–732. [Google Scholar] [CrossRef]
- Vykhovanets, E.V.; Resnick, M.I.; MacLennan, G.T.; Gupta, S. Experimental rodent models of prostatitis: Limitations and potential. Prostate Cancer Prostatic Dis. 2007, 10, 15–29. [Google Scholar] [CrossRef]
- Kang, S.W.; Park, J.H.; Seok, H.; Park, H.J.; Chung, J.H.; Kim, C.J.; Kim, Y.O.; Han, Y.R.; Hong, D.; Kim, Y.S.; et al. The Effects of Korea Red Ginseng on Inflammatory Cytokines and Apoptosis in Rat Model with Chronic Nonbacterial Prostatitis. Biomed. Res. Int. 2019, 2019, 2462561. [Google Scholar] [CrossRef]
- Ponniah, S.; Arah, I.; Alexander, R.B. PSA is a candidate self-antigen in autoimmune chronic prostatitis/chronic pelvic pain syndrome. Prostate 2000, 44, 49–54. [Google Scholar] [CrossRef]
- Bai, J.; Wang, S.; Liu, J.; Ye, Z.; Yu, X.; Xi, Q.; Hu, D.; Su, S. Characterization of circulating CD4+CD25high regulatory T cells in men with chronic prostatitis/chronic pelvic pain syndrome. Urology 2010, 75, 938–942. [Google Scholar] [CrossRef]
- Panchanathan, R.; Liu, H.; Choubey, D. Hypoxia primes human normal prostate epithelial cells and cancer cell lines for the NLRP3 and AIM2 inflammasome activation. Oncotarget 2016, 7, 28183–28199. [Google Scholar] [CrossRef]
- Lu, J.; Su, Y.; Chen, X.; Chen, Y.; Luo, P.; Lin, F.; Zhang, J. Rapamycininduced autophagy attenuates hormoneimbalanceinduced chronic nonbacterial prostatitis in rats via the inhibition of NLRP3 inflammasomemediated inflammation. Mol. Med. Rep. 2019, 19, 221–230. [Google Scholar] [PubMed]
- Kashyap, M.; Pore, S.; Wang, Z.; Gingrich, J.; Yoshimura, N.; Tyagi, P. Inflammasomes are important mediators of prostatic inflammation associated with BPH. J. Inflamm. (Lond.) 2015, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.G.; Chen, J.; Meng, J.L.; Zhang, Y.; Liu, Y.; Zhan, C.S.; Chen, X.G.; Zhang, L.; Liang, C.Z. Effect of alcohol on chronic pelvic pain and prostatic inflammation in a mouse model of experimental autoimmune prostatitis. Prostate 2019, 79, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Furuoka, M.; Ozaki, K.; Sadatomi, D.; Mamiya, S.; Yonezawa, T.; Tanimura, S.; Takeda, K. TNF-alpha Induces Caspase-1 Activation Independently of Simultaneously Induced NLRP3 in 3T3-L1 Cells. J. Cell Physiol. 2016, 231, 2761–2767. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Wojtczak, A.; Cieslak, M. Role of pro-inflammatory cytokines of pancreatic islets and prospects of elaboration of new methods for the diabetes treatment. Acta. Biochim. Pol. 2015, 62, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.W.; Fedorak, R.N. Tumor necrosis factor-alpha monoclonal antibodies in the treatment of inflammatory bowel disease: Clinical practice pharmacology. Gastroenterol. Clin. N. Am. 2010, 39, 543–557. [Google Scholar] [CrossRef]
- Rider, P.; Carmi, Y.; Cohen, I. Biologics for Targeting Inflammatory Cytokines, Clinical Uses, and Limitations. Int. J. Cell Biol. 2016, 2016, 9259646. [Google Scholar] [CrossRef]
- Joosten, L.A.; Helsen, M.M.; Saxne, T.; van de Loo, F.A.; Heinegard, D.; van den Berg, W.B. IL-1 alpha beta blockade prevents cartilage and bone destruction in murine type II collagen-induced arthritis, whereas TNF-alpha blockade only ameliorates joint inflammation. J. Immunol. 1999, 163, 5049–5055. [Google Scholar]
- Surguladze, D.; Deevi, D.; Claros, N.; Corcoran, E.; Wang, S.; Plym, M.J.; Wu, Y.; Doody, J.; Mauro, D.J.; Witte, L.; et al. Tumor Necrosis Factor-α and Interleukin-1 Antagonists Alleviate Inflammatory Skin Changes Associated with Epidermal Growth Factor Receptor Antibody Therapy in Mice. Cancer Res. 2009, 69, 5643–5647. [Google Scholar] [CrossRef]
- de Rivero Vaccari, J.P.; Lotocki, G.; Alonso, O.F.; Bramlett, H.M.; Dietrich, W.D.; Keane, R.W. Therapeutic neutralization of the NLRP1 inflammasome reduces the innate immune response and improves histopathology after traumatic brain injury. J. Cereb. Blood Flow Metab. 2009, 29, 1251–1261. [Google Scholar] [CrossRef]
- Cheng, C.; Xu, J.M.; Yu, T. Neutralizing IL-6 reduces heart injury by decreasing nerve growth factor precursor in the heart and hypothalamus during rat cardiopulmonary bypass. Life Sci. 2017, 178, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Koike, K.; Beatman, E.L.; Schweitzer, K.S.; Justice, M.J.; Mikosz, A.M.; Ni, K.; Clauss, M.A.; Petrache, I. Subcutaneous administration of neutralizing antibodies to endothelial monocyte-activating protein II attenuates cigarette smoke-induced lung injury in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L558–L566. [Google Scholar] [CrossRef] [PubMed]
- Hoehlig, K.; Maasch, C.; Shushakova, N.; Buchner, K.; Huber-Lang, M.; Purschke, W.G.; Vater, A.; Klussmann, S. A novel C5a-neutralizing mirror-image (l-)aptamer prevents organ failure and improves survival in experimental sepsis. Mol. Ther. 2013, 21, 2236–2246. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.L.; Gan, S.T.; Ho, B. Single-stranded DNA oligoaptamers: Molecular recognition and LPS antagonism are length- and secondary structure-dependent. J. Innate. Immun. 2009, 1, 46–58. [Google Scholar] [CrossRef]
- Chen, L.; Li, D.Q.; Zhong, J.; Wu, X.L.; Chen, Q.; Peng, H.; Liu, S.Q. IL-17RA aptamer-mediated repression of IL-6 inhibits synovium inflammation in a murine model of osteoarthritis. Osteoarthr. Cartil. 2011, 19, 711–718. [Google Scholar] [CrossRef]
- Hu, J.; Al-Waili, D.; Hassan, A.; Fan, G.C.; Xin, M.; Hao, J. Inhibition of cerebral vascular inflammation by brain endothelium-targeted oligodeoxynucleotide complex. Neuroscience 2016, 329, 30–42. [Google Scholar] [CrossRef]
- Boshtam, M.; Asgary, S.; Kouhpayeh, S.; Shariati, L.; Khanahmad, H. Aptamers Against Pro- and Anti-Inflammatory Cytokines: A Review. Inflammation 2017, 40, 340–349. [Google Scholar] [CrossRef]
- Shen, C.; Assche, G.V.; Colpaert, S.; Maerten, P.; Geboes, K.; Rutgeerts, P.; Ceuppens, J.L. Adalimumab induces apoptosis of human monocytes: A comparative study with infliximab and etanercept. Aliment. Pharmacol. Ther. 2005, 21, 251–258. [Google Scholar] [CrossRef]
- Atreya, R.; Zimmer, M.; Bartsch, B.; Waldner, M.J.; Atreya, I.; Neumann, H.; Hildner, K.; Hoffman, A.; Kiesslich, R.; Rink, A.D.; et al. Antibodies against tumor necrosis factor (TNF) induce T-cell apoptosis in patients with inflammatory bowel diseases via TNF receptor 2 and intestinal CD14(+) macrophages. Gastroenterology 2011, 141, 2026–2038. [Google Scholar] [CrossRef]
- Torene, R.; Nirmala, N.; Obici, L.; Cattalini, M.; Tormey, V.; Caorsi, R.; Starck-Schwertz, S.; Letzkus, M.; Hartmann, N.; Abrams, K.; et al. Canakinumab reverses overexpression of inflammatory response genes in tumour necrosis factor receptor-associated periodic syndrome. Ann. Rheum. Dis. 2017, 76, 303. [Google Scholar] [CrossRef]
- Sharma, R.; Anker, S.D. Cytokines, apoptosis and cachexia: The potential for TNF antagonism. Int. J. Cardiol. 2002, 85, 161–171. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Nakajima, S.; Hosojima, S.; Thi Nguyen, D.; Hattori, T.; Manh Le, T.; Hori, O.; Mahib, M.R.; Yamaguchi, Y.; Miura, M.; et al. Caspase-1 initiates apoptosis in the absence of gasdermin D. Nat. Commun. 2019, 10, 2091. [Google Scholar] [CrossRef] [PubMed]
- William, R.; Watson, G.; Rotstein, O.D.; Parodo, J.; Bitar, R.; Marshall, J.C. The IL-1β-Converting Enzyme (Caspase-1) Inhibits Apoptosis of Inflammatory Neutrophils through Activation of IL-1β. J. Immunol. 1998, 161, 957–962. [Google Scholar]
- Szondy, Z.; Sarang, Z.; Kiss, B.; Garabuczi, É.; Köröskényi, K. Anti-inflammatory Mechanisms Triggered by Apoptotic Cells during Their Clearance. Front. Immunol. 2017, 8, 909. [Google Scholar] [CrossRef] [PubMed]
- Voll, R.E.; Herrmann, M.; Roth, E.A.; Stach, C.; Kalden, J.R.; Girkontaite, I. Immunosuppressive effects of apoptotic cells. Nature 1997, 390, 350–351. [Google Scholar] [CrossRef] [PubMed]
- Sharma-Chawla, N.; Stegemann-Koniszewski, S.; Christen, H.; Boehme, J.D.; Kershaw, O.; Schreiber, J.; Guzmán, C.A.; Bruder, D.; Hernandez-Vargas, E.A. In vivo Neutralization of Pro-inflammatory Cytokines During Secondary Streptococcus pneumoniae Infection Post Influenza A Virus Infection. Front. Immunol. 2019, 10, 1864. [Google Scholar] [CrossRef]
- Ross, M.E.; Caligiuri, M.A. Cytokine-induced apoptosis of human natural killer cells identifies a novel mechanism to regulate the innate immune response. Blood 1997, 89, 910–918. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Kowsar, R.; Keshtegar, B.; Miyamoto, A. Understanding the hidden relations between pro- and anti-inflammatory cytokine genes in bovine oviduct epithelium using a multilayer response surface method. Sci. Rep. 2019, 9, 3189. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, D.-R.; Chang, P.-J.; Lin, W.-Y.; Huang, Y.-C.; Lin, J.-H.; Huang, K.-T.; Chan, W.-N.; Chen, C.-S. Beneficial Effects of Inflammatory Cytokine-Targeting Aptamers in an Animal Model of Chronic Prostatitis. Int. J. Mol. Sci. 2020, 21, 3953. https://doi.org/10.3390/ijms21113953
Ho D-R, Chang P-J, Lin W-Y, Huang Y-C, Lin J-H, Huang K-T, Chan W-N, Chen C-S. Beneficial Effects of Inflammatory Cytokine-Targeting Aptamers in an Animal Model of Chronic Prostatitis. International Journal of Molecular Sciences. 2020; 21(11):3953. https://doi.org/10.3390/ijms21113953
Chicago/Turabian StyleHo, Dong-Ru, Pey-Jium Chang, Wei-Yu Lin, Yun-Ching Huang, Jian-Hui Lin, Kuo-Tsai Huang, Wai-Nga Chan, and Chih-Shou Chen. 2020. "Beneficial Effects of Inflammatory Cytokine-Targeting Aptamers in an Animal Model of Chronic Prostatitis" International Journal of Molecular Sciences 21, no. 11: 3953. https://doi.org/10.3390/ijms21113953
APA StyleHo, D.-R., Chang, P.-J., Lin, W.-Y., Huang, Y.-C., Lin, J.-H., Huang, K.-T., Chan, W.-N., & Chen, C.-S. (2020). Beneficial Effects of Inflammatory Cytokine-Targeting Aptamers in an Animal Model of Chronic Prostatitis. International Journal of Molecular Sciences, 21(11), 3953. https://doi.org/10.3390/ijms21113953