Differentiation of Human Cardiac Atrial Appendage Stem Cells into Adult Cardiomyocytes: A Role for the Wnt Pathway?


Abstract
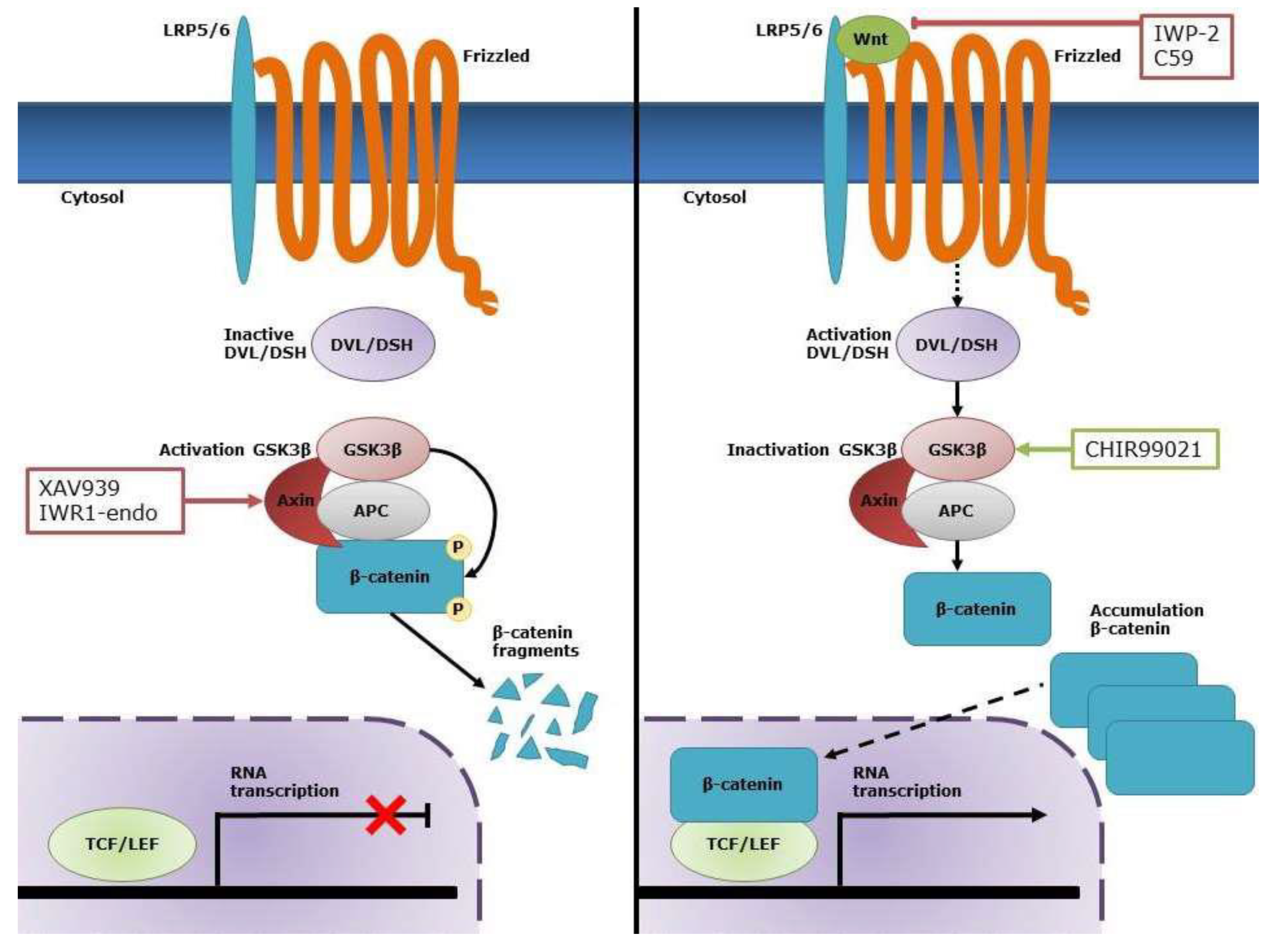
1. Introduction
2. Results
2.1. CASCs Are Predominantly Present in the RAA
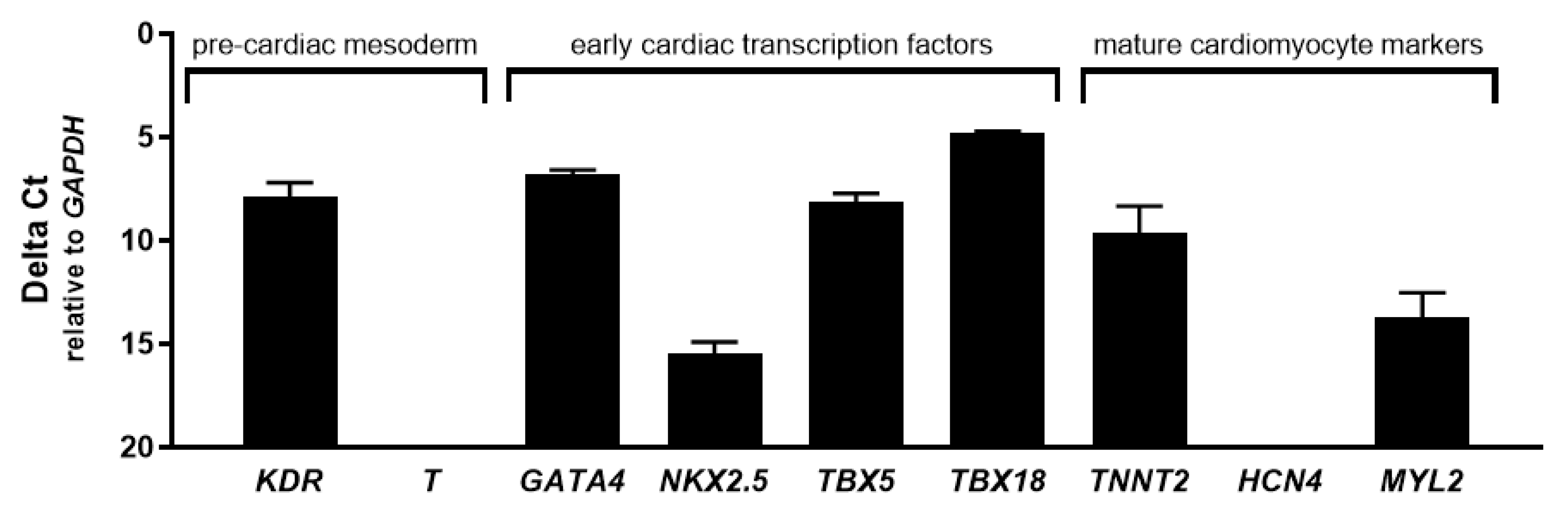
2.2. CASCs Express Early Cardiac Differentiation Markers during Expansion
2.3. Several FZD Receptor Subtypes are Expressed in CASCs
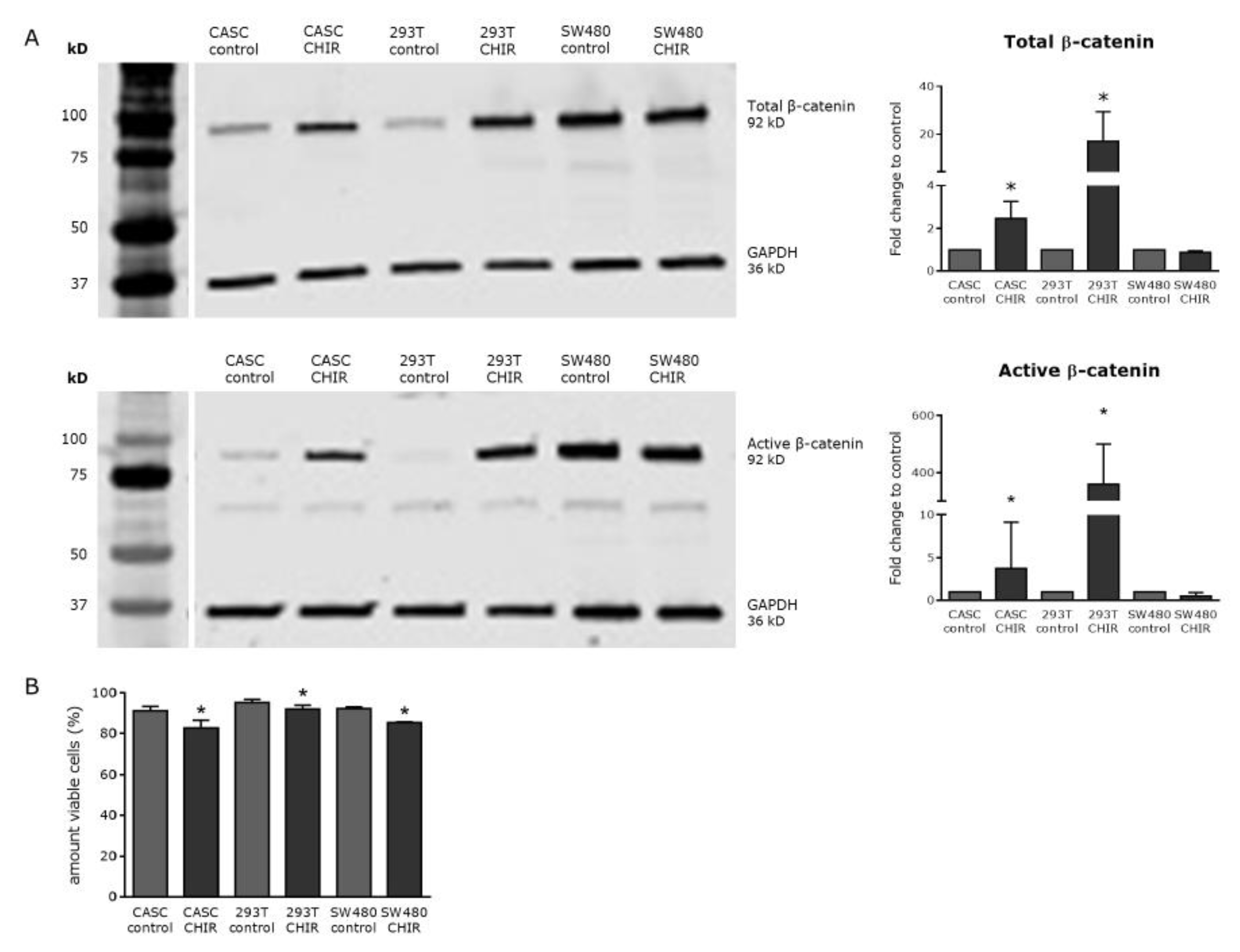
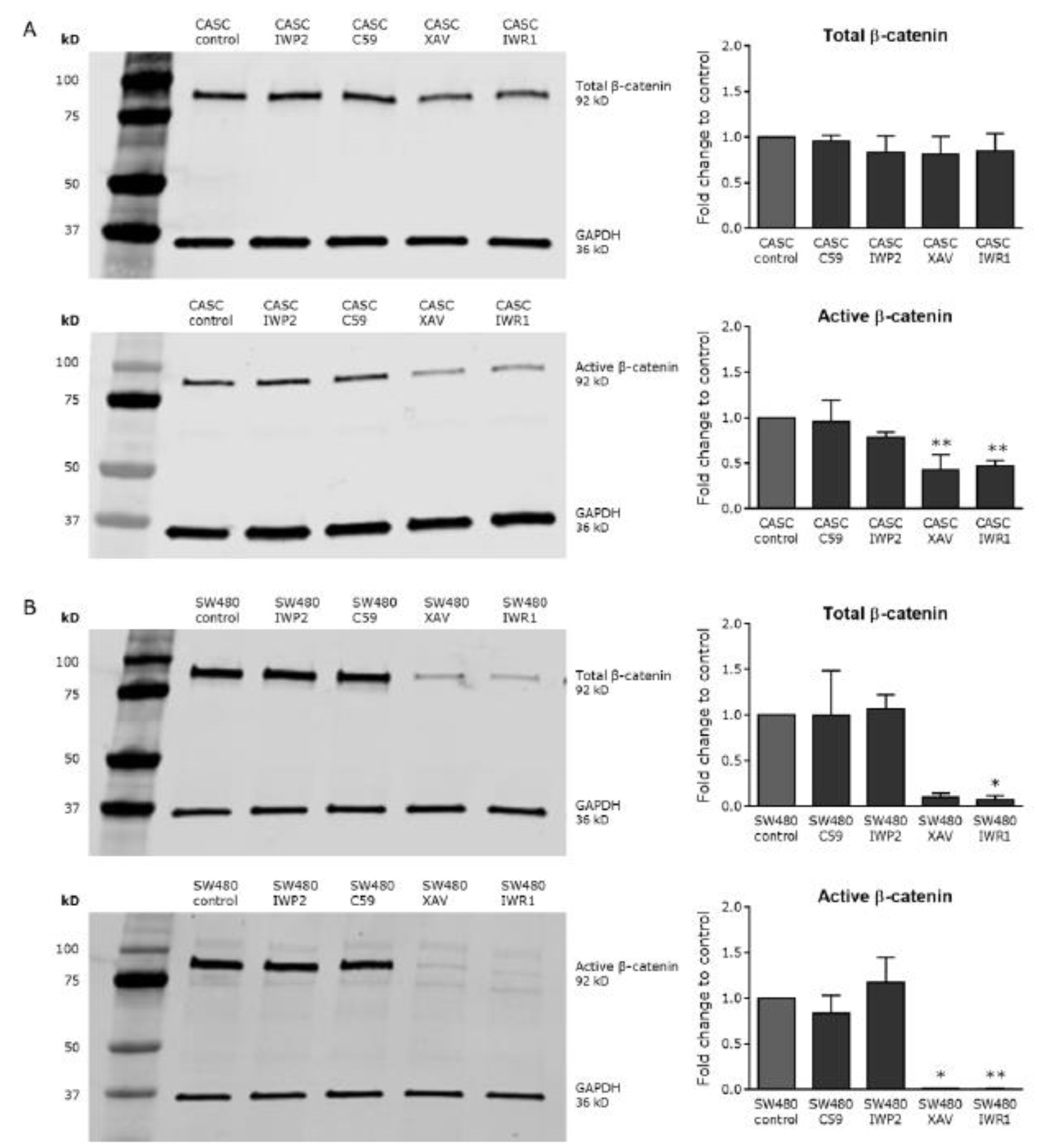
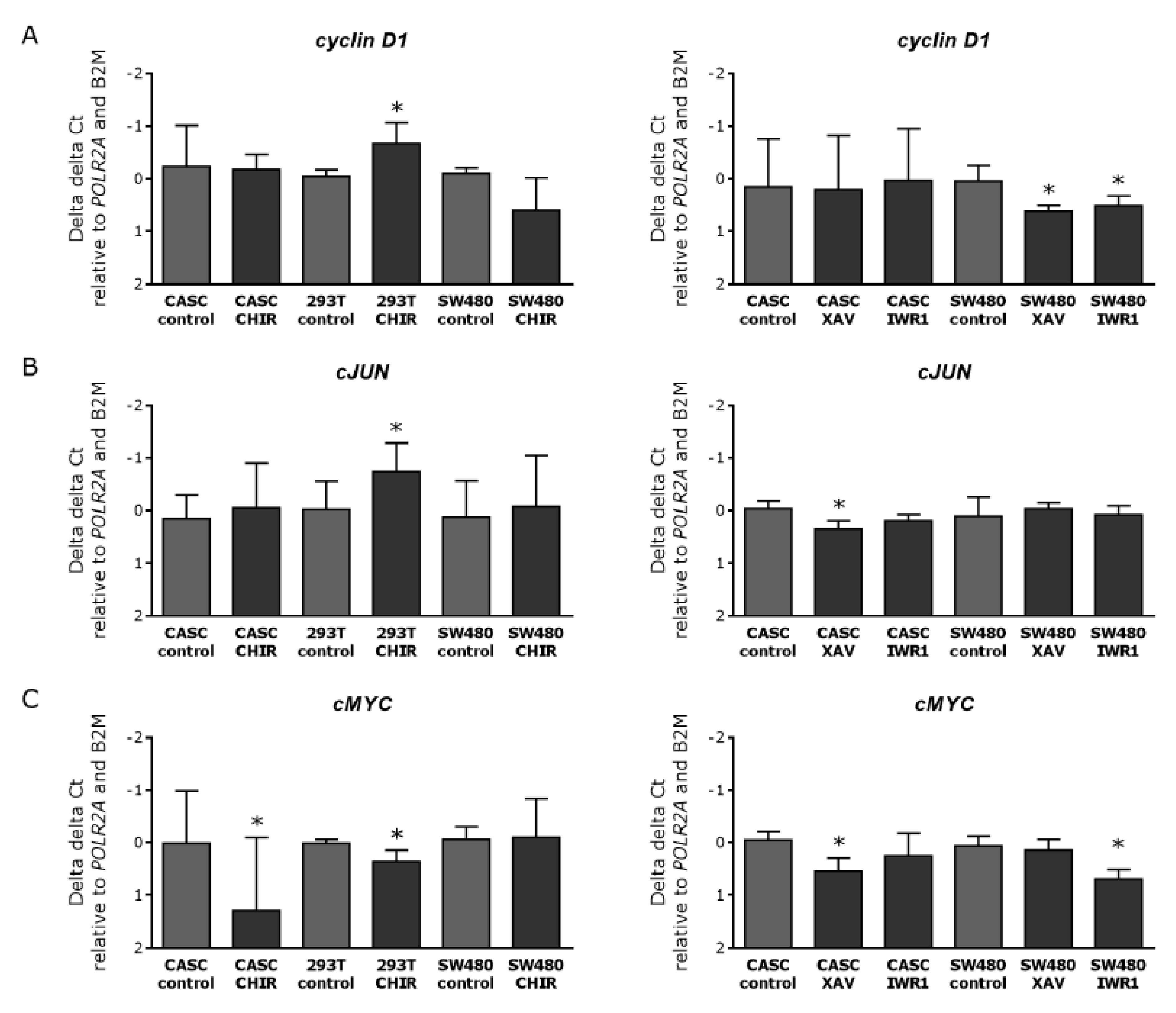
2.4. Wnt Signaling Can Be Modulated in CASCs by Specific Small-Molecule Activators and Inhibitors
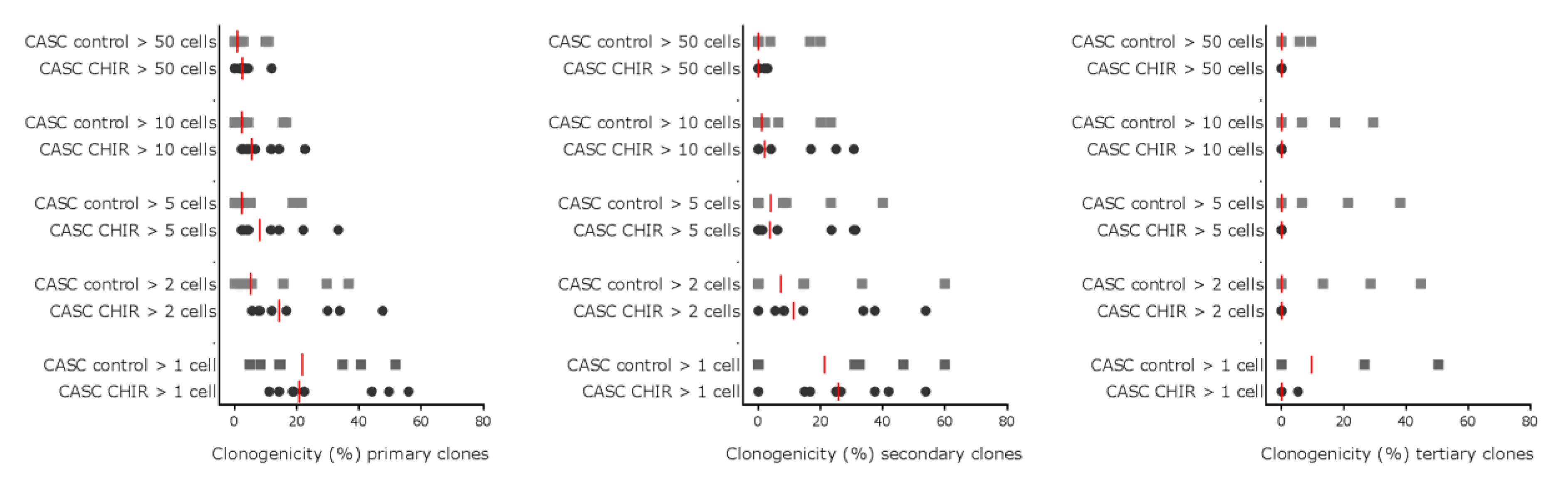
2.5. Modulating Wnt Signaling Does Not Influence CASC Clonogenicity or Proliferation
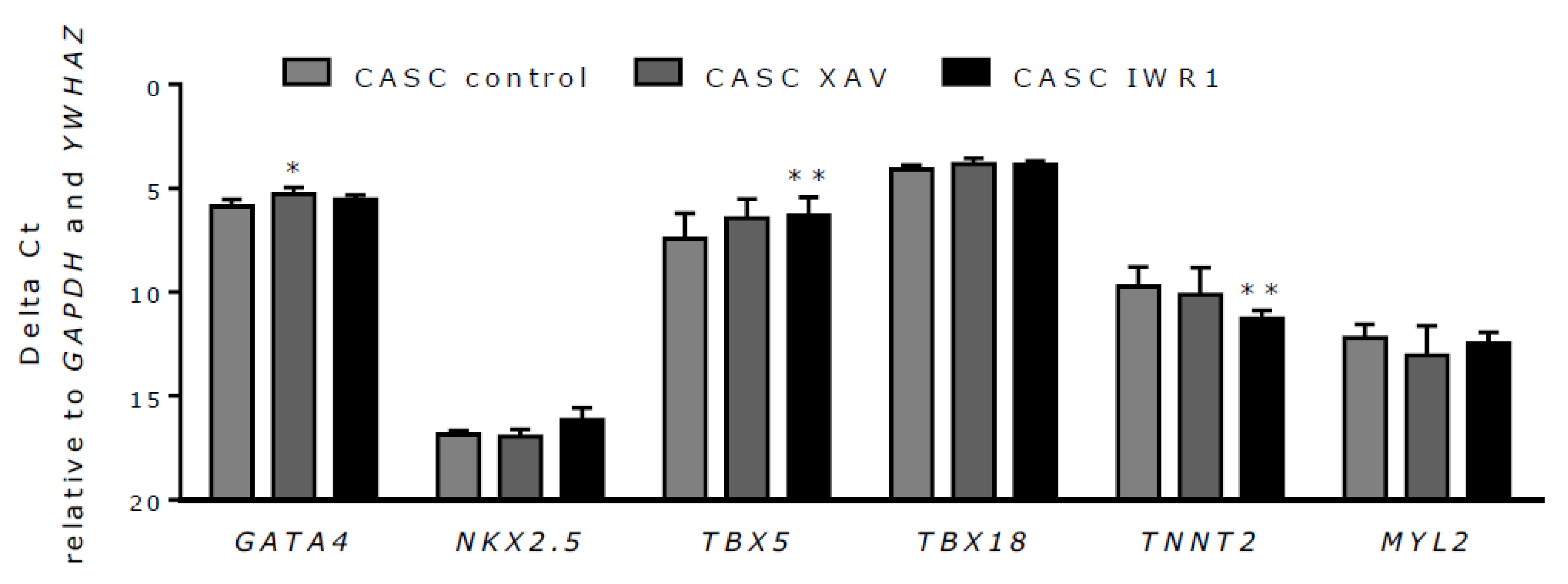
2.6. Wnt Inhibition in CASCs Does Not Induce Mature Cardiomyocyte Differentiation
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Cell Culture
4.3. Flow Cytometric Analyses of ALDHbr Cells in Different Compartments of the Heart
4.4. Drugs and Treatment
4.5. RNA Isolation and Complementary DNA (cDNA) Synthesis
4.6. Conventional PCR
4.7. Quantitative PCR
4.8. Western Blot Analysis
4.9. Viability Testing
4.10. Serial Clonogenic Assay
4.11. Statistics
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Korewicki, J. Cardiac transplantation is still the method of choice in the treatment of patients with severe heart failure. Cardiol. J. 2009, 16, 493–499. [Google Scholar] [PubMed]
- Gyongyosi, M.; Wojakowski, W.; Lemarchand, P.; Lunde, K.; Tendera, M.; Bartunek, J.; Marban, E.; Henry, T.D.; Traverse, J.H.; Moyé, L.A.; et al. Meta-Analysis of Cell-based CaRdiac stUdiEs (ACCRUE) in patients with acute myocardial infarction based on individual patient data. Circ. Res. 2015, 116, 1346–1360. [Google Scholar] [CrossRef] [PubMed]
- Bruyneel, A.A.; Sehgal, A.; Malandraki-Miller, S.; Carr, C. Stem Cell Therapy for the Heart: Blind Alley or Magic Bullet? J. Cardiovasc. Trans. Res. 2016, 9, 405–4018. [Google Scholar] [CrossRef] [PubMed]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef]
- Bearzi, C.; Rota, M.; Hosoda, T.; Tillmans, J.; Nascimbene, A.; De Angelis, A.; Yasuzawa-Amano, S.; Trofimona, I.; Siggins, R.W.; Lecapitaine, N.; et al. Human cardiac stem cells. Proc. Nat. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar] [CrossRef]
- Bolli, R.; Chugh, A.R.; D’Amario, D.; Loughran, J.H.; Stoddard, M.F.; Ikram, S.; Beache, G.M.; Wagner, S.G.; Leri, A.; Hosoda, T.; et al. Cardiac stem cells in patients with ischemic cardiomyopathy (SCIPIO): Initial results of a randomised phase 1 trial. Lancet 2011, 378, 1847–1857. [Google Scholar] [CrossRef]
- The Lancet Editors. RETRACTION-Cardiac stem cells in patients with ischemic cardiomyopathy (SCIPIO): Initial results of a randomised phase 1 trial. Lancet 2019, 393, 1084. [Google Scholar] [CrossRef]
- Van Berlo, J.H.; Kanisicak, O.; Maillet, M.; Vagnozzi, R.J.; Karch, J.; Lin, S.C.; Middleton, R.C.; Marban, E.; Molkentin, J.D. C-kit+ cells minimally contribute cardiomyocytes to the heart. Nature 2014, 509, 337–341. [Google Scholar] [CrossRef]
- Sultana, N.; Zhang, L.; Yan, J.; Chen, J.; Cai, W.; Razzaque, S.; Jeong, D.; Sheng, W.; Bu, L.; Xu, M.; et al. Resident c-kit(+) cells in the heart are not cardiac stem cells. Nat. Commun. 2015, 6, 8701. [Google Scholar] [CrossRef]
- Koninckx, R.; Daniels, A.; Windmolders, S.; Mees, U.; Macianskiene, R.; Mubagwa, K.; Steels, P.; Jamaer, L.; Dubois, J.; Robic, B.; et al. The cardiac atrial appendage stem cell: A new and promising candidate for myocardial repair. Cardiovasc. Res. 2013, 97, 413–423. [Google Scholar] [CrossRef]
- Roehrich, M.E.; Spicher, A.; Milano, G.; Vassalli, G. Characterization of cardiac-resident progenitor cells expressing high aldehyde dehydrogenase activity. BioMed Res. Int. 2013, 2013, 503047. [Google Scholar] [CrossRef] [PubMed]
- Fanton, Y.; Robic, B.; Rummens, J.L.; Daniels, A.; Windmolders, S.; Willems, L.; Jamaer, L.; Dubois, J.; Bijnens, E.; Heuts, N.; et al. Cardiac atrial appendage stem cells engraft and differentiate into cardiomyocytes in vivo: A new tool for cardiac repair after MI. Int. J. Cardiol. 2015, 201, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Fanton, Y.; Houbrechts, C.; Willems, L.; Daniels, A.; Linsen, L.; Ratajczak, J.; Bronckaers, A.; Lambrichts, I.; Declercq, J.; Rummens, J.L.; et al. Cardiac atrial appendage stem cells promote angiogenesis in vitro and in vivo. J. Mol. Cell. Cardiol. 2016, 97, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Ozhan, G.; Weidinger, G. Wnt/beta-catenin signaling in heart regeneration. Cell Regener. 2015, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R. Wnt signaling in disease and in development. Cell Res. 2005, 15, 28–32. [Google Scholar] [CrossRef]
- Gessert, S.; Kühl, M. The multiple phases and faces of wnt signaling during cardiac differentiation and development. Circ. Res. 2010, 107, 186–199. [Google Scholar] [CrossRef]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/beta-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef]
- Windmolders, S.; Willems, L.; Daniels, A.; Linsen, L.; Fanton, Y.; Hendrikx, M.; Koninckx, R.; Rummens, J.L.; Hensen, K. Clinical-scale in vitro expansion preserves biological characteristics of cardiac atrial appendage stem cells. Cell Prolif. 2015, 48, 175–186. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, W.; Evans, P.M.; Chen, X.; He, X.; Liu, C. Adenomatous polyposis coli (APC) differentially regulates beta-catenin phosphorylation and ubiquitination in colon cancer cells. J. Biol. Chem. 2006, 281, 17751–17757. [Google Scholar] [CrossRef]
- Cai, C.L.; Martin, J.C.; Sun, Y.; Cui, L.; Wang, L.; Ouyang, K.; Yang, L.; Bu, L.; Liang, X.; Zhang, X.; et al. A myocardial lineage derives from Tbx18 epicardial cells. Nature 2008, 454, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Wang, G.; Lin, L.; Lowe, J.; Zhang, Q.; Bu, L.; Chen, Y.; Chen, J.; Sun, Y.; Evans, S.M. HCN4 dynamically marks the first heart field and conduction system precursors. Circ. Res. 2013, 113, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, J.K.; Ohgi, M.; Koshiba-Takeuchi, K.; Shiratori, H.; Sakaki, I.; Ogura, K.; Saijoh, Y.; Ogura, T. Tbx5 specifies the left/right ventricles and ventricular septum position during cardiogenesis. Development 2003, 130, 5953–5964. [Google Scholar] [CrossRef] [PubMed]
- Santini, M.P.; Forte, E.; Harvey, R.P.; Kovacic, J.C. Developmental origin and lineage plasticity of endogenous cardiac stem cells. Development 2016, 143, 1242–1258. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.M.; Fujiwara, Y.; Cibulsky, S.M.; Clapham, D.E.; Lien, C.L.; Schultheiss, T.M.; Orkin, S.H. Developmental origin of a bipotential myocardial and smooth muscle cell precursor in the mammalian heart. Cell 2006, 127, 1137–1150. [Google Scholar] [CrossRef]
- Freire, A.G.; Resende, T.P.; Pinto-do-O, P. Building and repairing the heart: What can we learn from embryonic development? BioMed Res. Int. 2014, 2014, 679168. [Google Scholar] [CrossRef]
- Naujok, O.; Lentes, J.; Diekmann, U.; Davenport, C.; Lenzen, S. Cytotoxicity and activation of the Wnt/beta-catenin pathway in mouse embryonic stem cells treated with four GSK3 inhibitors. BMC Res. Notes 2014, 7, 273. [Google Scholar] [CrossRef]
- Xiao, G.; Mao, S.; Baumgarten, G.; Serrano, J.; Jordan, M.C.; Roos, K.P.; Fishbein, M.C.; MacLellan, W.R. Inducible activation of c-Myc in adult myocardium in vivo provokes cardiac myocyte hypertrophy and reactivation of DNA synthesis. Circ. Res. 2001, 89, 1122–1129. [Google Scholar] [CrossRef]
- Qyang, Y.; Martin-Puig, S.; Chiravuri, M.; Chen, S.; Xu, H.; Bu, L.; Jiang, X.; Lin, L.; Granger, A.; Moretti, A.; et al. The renewal and differentiation of Isl1+ cardiovascular progenitors are controlled by a Wnt/beta-catenin pathway. Cell Stem Cell 2007, 1, 165–179. [Google Scholar] [CrossRef]
- Oikonomopoulos, A.; Sereti, K.I.; Conyers, F.; Bauer, M.; Liao, A.; Guan, J.; Crapps, D.; Han, J.K.; Dong, H.; Bayomy, A.F.; et al. Wnt signaling exerts an antiproliferative effect on adult cardiac progenitor cells through IGFBP3. Circ. Res. 2011, 109, 1363–1374. [Google Scholar] [CrossRef]
- Afouda, B.A.; Martin, J.; Liu, F.; Ciau-Uitz, A.; Patient, R.; Hoppler, S. GATA transcription factors integrate Wnt signalling during heart development. Development 2008, 135, 3185–3190. [Google Scholar] [CrossRef] [PubMed]
- Zelarayan, L.C.; Noack, C.; Sekkali, B.; Kmecova, J.; Gehrke, C.; Renger, A.; Zafiriou, M.P.; van der Nagel, R.; Dietz, R.; de Windt, L.J.; et al. Beta-Catenin downregulation attenuates ischemic cardiac remodeling through enhanced resident precursor cell differentiation. Proc. Natl. Acad. Sci. USA 2008, 105, 19762–19767. [Google Scholar] [CrossRef] [PubMed]
- Foley, A.C.; Mercola, M. Heart induction by Wnt antagonists depends on the homeodomain transcription factor Hex. Genes Dev. 2005, 19, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Koninckx, R.; Daniels, A.; Windmolders, S.; Carlotti, F.; Mees, U.; Steels, P.; Rummens, J.L.; Hendrikx, M.; Hensen, K. Mesenchymal stem cells or cardiac progenitors for cardiac repair? A comparative study. Cell. Mol. Life Sci. 2011, 68, 2141–2156. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Müller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALDHbr Cells (%) | |||
|---|---|---|---|
| Pig heart 1 | Pig heart 2 | Pig heart 3 | |
| LAA | 4.4 | 4.9 | 3.5 |
| RAA | 10.5 | 5.6 | 8.7 |
| Left atrium | 0.4 | 1.5 | 1.5 |
| Right atrium | 2.8 | 1.5 | 3.9 |
| Left-ventricular free wall | 0.5 | 0.4 | 0.2 |
| Right-ventricular free wall | 1.1 | 1.9 | 1.9 |
| Apex | 3.1 | 1.6 | 0.2 |
| Septum | 0.3 | 0.4 | 0.2 |
| Patient Characteristics | |
|---|---|
| Age (years) | 70 ± 14 a |
| Gender: male/female (%) | 70/30 a |
| Risk factors | |
| Weight (kg) | 80 ± 15 a |
| Body mass index (kg/m²) | 28 ± 5 a |
| Last creatinine level pre-operation (mg/dL) | 1.0 ± 0.3 a |
| Smoking: current smoker/ex-smoker ≥1 month/never smoked (%) | 15/36/48 a |
| Diabetes (%) | 33 a |
| Hypercholesterolemia (%) | 87 c |
| Renal dysfunction (%) | 9 b |
| Hypertension (%) | 61 c |
| Chronic lung disease (%) | 16 b |
| Peripheral vascular disease (%) | 13 b |
| Cerebrovascular disease (%) | 19 b |
| Pre-operative cardiac status | |
| Myocardial infarction (%) | 34 b |
| Congestive heart failure (%) | 16 c |
| Angina: CCS 0/I/II/III/IV (%) | 18/18/57/4/4 d |
| Classification: NYHA I/II/III/IV (%) | 91/18/14/0 d |
| Surgical procedure | |
| Number of vessels: 0/1/2/3 (%) | 9/3/27/61 a |
| CABG/valve/other (%) | 91/15/3 a, e |
| Primer | Primer Sequence | Annealing Temperature | Fragment Size |
|---|---|---|---|
| β-actin | Forward: 5′–AGCGGGAAATCGTGCGTGACA–3′ Reverse: 5′–CCTGTAACAATGCATCTCATATTTGG–3′ | 56 °C | 791 bp |
| FZD1 | Forward: 5′–CCTTTCTTTCCTGGCTTGA–3′ Reverse: 5′–CTCACCCTGTAACCAACTAAG–3′ | 56 °C | 285 bp |
| FZD2 | Forward: 5′–CCCTACTCATTTGTCCTGTC–3′ Reverse: 5′–TGAATAGACTGCAGGGAAAG–3′ | 56 °C | 405 bp |
| FZD3 | Forward: 5′–CTCCTGAGGGATCCAAATAC–3′ Reverse: 5′–GAGCCGATGAGAACTACTATG–3′ | 56 °C | 282 bp |
| FZD4 | Forward: 5′–GAGAGAGAAGAGAGGAAATGG–3′ Reverse: 5′–GGTCACTTAATTGTTGCTAGTT–3′ | 56 °C | 185 bp |
| FZD5 | Forward: 5′–CCCAGAGCTAGGAAATGTAG–3′ Reverse: 5′–GATGTGCTCTGTCCTGTT–3′ | 56 °C | 165 bp |
| FZD6 | Forward: 5′–CATCAATGAGAGAGGTGAAAG–3′ Reverse: 5′–GGGTGAACAAGCAGAGAT–3′ | 56 °C | 286 bp |
| FZD7 | Forward: 5′–CATTTGGATCCTTTGAGGTAAA–3′ Reverse: 5′–CTCTTCGTTCACTATGGTATCT–3′ | 56 °C | 203 bp |
| FZD8 | Forward: 5′–CTCTTCCTACGTAAACTCCC–3′ Reverse: 5′–GAGAGGGCAATGGTTAAATC–3′ | 56 °C | 356 bp |
| FZD9 | Forward: 5′–CTCCAAGACTTTCCAGACC–3′ Reverse: 5′–GTCCGTCTTAGTCATGTGC–3′ | 56 °C | 157 bp |
| FZD10 | Forward: 5′–GATTCAGCCCTCAGAAGAA–3′ Reverse: 5′–GCAGAGAGACTATTGGTGAA–3′ | 56 °C | 292 bp |
| Primer | Primer Sequence | Annealing Temperature | Concentration |
|---|---|---|---|
| B2M | Forward: 5′–AAGATGAGTATGCCTGCCGT–3′ Reverse: 5′–TTCATCCAATCCAAATGCGGC–3′ | 60 °C | 300 nM |
| cJUN | Forward: 5′–AGGTGGAGTTGAAAGAGTTAAGA–3′ Reverse: 5′–ACCATAGCATCAGGTACATCAG–3′ | 60 °C | 300 nM |
| cMYC | Forward: 5′–TTCTCTGAAAGGCTCTCCT–3′ Reverse: 5′–GTGAAGCTAACGTTGAGGG–3′ | 60 °C | 300 nM |
| Cyclin D1 | Forward: 5′–TCAAATGTGTGCAGAAGGAG–3′ Reverse: 5′–TCTCCTTCATCTTAGAGGCC–3′ | 60 °C | 100 nM |
| GAPDH | Forward: 5′–AGTCAACGGATTTGGTCGTATTG–3′ Reverse: 5′–ATCTCGCTCCTGGAAGATGGT–3′ | 60 °C | 300 nM |
| GATA4 | Forward: 5′–ACCTGAATAAATCTAAGACACCAG–3′ Reverse: 5′–CATCGCACTGACTGAGAAC–3′ | 60 °C | 100 nM |
| HCN4 | Forward: 5′–AGTCGGCCGGATTTTGGATT–3′ Reverse: 5′–AGGTGATGCCCACAGGAATG–3′ | 58 °C | 100 nM |
| KDR | Forward: 5′–CTAGGTAAGCCTCTTGGCCG–3′ Reverse: 5′–CGATGCTCACTGTGTGTTGC–3′ | 66 °C | 300 nM |
| MYL2 | Forward: 5′–TTGGGCGAGTGAACGTGAAA–3′ Reverse: 5′–GGTCCGCTCCCTTAAGTTTCT–3′ | 60 °C | 300 nM |
| MYL7 | Forward: 5′–GGAGTTCAAAGAAGCCTTCAGC–3′ Reverse: 5′–GTCAGGGCGAACATCTGCT–3′ | 60 °C | 300 nM |
| NKX2.5 | Forward: 5′–CAAGGACCCTAGAGCCGAAA–3′ Reverse: 5′–CACCGACACGTCTCACTCAG–3′ | 58 °C | 450 nM |
| POLR2A | Forward: 5′–TCACAGCAGTGCGCAAATTC–3′ Reverse: 5′–CCACGTCGACAGGAACATCA–3′ | 60 °C | 300 nM |
| T | Forward: 5′–ACTCCCAATCCTATTCTGACAACT–3′ Reverse: 5′–CGTTGCTCACAGACCACAGG–3′ | 60 °C | 450 nM |
| TBX5 | Forward: 5′–CAGGAGCATAGCCAAATTTACCA–3′ Reverse: 5′–GGATAGCTAGAGCGGTAGAAGGA–3′ | 62 °C | 300 nM |
| TBX18 | Forward: 5′–ATGCATTCTGGCGACCATCA–3′ Reverse: 5′–ACGCCATTCCCAGTACCTTG–3′ | 62 °C | 300 nM |
| TNNT2 | Forward: 5′–ACTTGGAGGCAGAGAAGTTCG–3′ Reverse: 5′–CGGTGACTTTAGCCTTCCCG–3′ | 66 °C | 100 nM |
| YHWAZ | Forward: 5′–AGACGGAAGGTGCTGAGAAAA–3′ Reverse: 5′–TGTGAAGCATTGGGGATCAAGA–3′ | 60 °C | 300 nM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willems, L.; Daniëls, A.; Fanton, Y.; Linsen, L.; Evens, L.; Bito, V.; Declercq, J.; Rummens, J.-L.; Hensen, K.; Hendrikx, M. Differentiation of Human Cardiac Atrial Appendage Stem Cells into Adult Cardiomyocytes: A Role for the Wnt Pathway? Int. J. Mol. Sci. 2020, 21, 3931. https://doi.org/10.3390/ijms21113931
Willems L, Daniëls A, Fanton Y, Linsen L, Evens L, Bito V, Declercq J, Rummens J-L, Hensen K, Hendrikx M. Differentiation of Human Cardiac Atrial Appendage Stem Cells into Adult Cardiomyocytes: A Role for the Wnt Pathway? International Journal of Molecular Sciences. 2020; 21(11):3931. https://doi.org/10.3390/ijms21113931
Chicago/Turabian StyleWillems, Leen, Annick Daniëls, Yanick Fanton, Loes Linsen, Lize Evens, Virginie Bito, Jeroen Declercq, Jean-Luc Rummens, Karen Hensen, and Marc Hendrikx. 2020. "Differentiation of Human Cardiac Atrial Appendage Stem Cells into Adult Cardiomyocytes: A Role for the Wnt Pathway?" International Journal of Molecular Sciences 21, no. 11: 3931. https://doi.org/10.3390/ijms21113931
APA StyleWillems, L., Daniëls, A., Fanton, Y., Linsen, L., Evens, L., Bito, V., Declercq, J., Rummens, J.-L., Hensen, K., & Hendrikx, M. (2020). Differentiation of Human Cardiac Atrial Appendage Stem Cells into Adult Cardiomyocytes: A Role for the Wnt Pathway? International Journal of Molecular Sciences, 21(11), 3931. https://doi.org/10.3390/ijms21113931