Exosomal Long Non-Coding RNAs in Lung Diseases

 , , ,
, , ,
Abstract
1. Introduction
1.1. Overview of Lung Diseases Covered in the Current Review
1.1.1. Idiopathic Pulmonary Fibrosis (IPF)
1.1.2. Chronic Obstructive Pulmonary Disease (COPD)
1.1.3. Asthma
1.1.4. Lung Cancer
1.2. Exosomes
1.2.1. Exosomes Are Essential Actors of Intercellular Communication
1.2.2. Exosomes Are Playing an Essential Role in Lung Diseases
1.3. Long Non-Coding Rnas
1.3.1. Transcript-Regulating LncRNAs
1.3.2. Epigenetics-Regulating LncRNAs
2. LncRNAs and Their Exosomes in Lung Diseases
2.1. H19
2.1.1. H19 and IPF
2.1.2. H19 and Asthma
2.1.3. H19 and COPD
2.1.4. H19 and Lung Cancer
- ⋄
- Regulators of H19: In NSCLC tumor tissues, FOXF2 can bind the promoter of H19 and can increase its expression, causing a PTEN downregulation [135]. Additionally, H19 may be a direct transcriptional target of and is induced by MYC in NSCLC tumor tissues. Indeed, MYC binds to H19 promoter’s E-boxes to facilitate histone acetylation and transcriptional initiation. Furthermore, MYC can downregulate the expression of IGF2 independently [136,137]. Besides, Shahdoust et al. found H19 among the seven most differentially expressed lncRNAs in the human airway epithelium of cigarette smokers when compared to non-smokers [138]. Similar results, obtained in cdk-4/hTERT-immortalized human bronchial epithelial cells (HBEC), described the H19 upregulation following prolonged CSC exposure. Interestingly, the same authors also found a general diminution of H4K16ac and H4K20me3 and an overall increase of H3K27me3 levels [74]. Nevertheless, H19 methylation status remains low, as RIOX2 can remove methyl groups from H3K9me3 on the H19 promoter, leading to de-repress H19 transcription [139]. Importantly, Liu et al. suggested that H19 demethylation may precede the methylations that silence tumor suppressor genes such as p16-CDKN2A, MGMT, DAPK, E-cadherin (CDH1), and CDH13 [74]. Moreover, the histone alterations coincided with a decreased DNMT1 and an increased DNMT3B expressions, as well as the activation of the WNT/β-catenin signaling pathway during prolonged CSC exposure [74]. Indeed, the authors found that WNT ligands, such as WNT2, WNT5A, WNT6, and WNT10A, and the Wnt signaling targets FOXN1 and TCF7, were up-regulated [74].
- ⋄
- H19 regulated genes: In NSCLC tumor tissues, H19 decreases the expression of CDH1 by inducing its promoter methylation and also increases CDH2 and VIM expressions [134,140]. Moreover, in human NSCLC cell lines (A549), H19 may regulate metastasis through the modulation of cell proliferation and cell adhesion proteins, including MACC1, EGFR, -catenin (CTNNB1), ERK1 (MAPK3) and ERK2 (MAPK1) [141].
- ⋄
- ⋄
- H19 as ceRNA: Among its known functions, H19 was reported as a heavy miRNA regulator. Indeed, H19 can sequester: • hsa-miR-107, to release the expression of NF1 in NSCLC tumor tissues [128] • hsa-miR-200a, to release the expression of ZEB1 and ZEB2 in NSCLC tumor tissues [130] • hsa-miR-29b-3p and hsa-miR-17, to release the expression of STAT3 in NSCLC tumor tissues [131,132] • hsa-miR-196b to release the expression of LIN28B and induce cell growth in NSCLC tumor tissues [142] • hsa-miR-138 (MIR138-1, MI0000476 or MIR138-2, MI0000455) to release the expression of PDK1 in NSCLC tumor tissues [129] • hsa-miR-484 to release the expression of ROCK2 and increase the levels of phosphorylated JUN as well as the mesenchymal markers N-cadherin (CDH2), vimentin (VIM), ZEB1 and SNAI1 while decreasing the level of the epithelial marker CDH1 in NSCLC tumor tissues and cell lines [133,134,143]. A similar observation can be made with the downregulation of hsa-miRNA-203 (hsa-miRNA-203a or hsa-miRNA-203b), which was associated with VIM and SNAI1 upregulation and CDH1 downregulation in NSCLC tumor tissues [143].
- ⋄
- 19 as a miRNA regulator: In NSCLC tumor tissues, H19 induces hsa-miR-675-5p expression, which, in turn, increases the expression of BCL2 and decreases the expression of TP53 as well as BAX [144,145,146]. Xu et al. hypothesized H19 as an epigenetic regulator of hsa-miR-6515-3p, which contributes to metastasis [147]. H19 expression was also positively correlated to hsa-miR-21 expression [97].
- ⋄
- H19 at the clinical level: H19 upregulation in NSCLC tumor tissues was associated with advanced tumor–node–metastasis (TNM) stages and negatively correlated with patient Overall Survival (OS) [96,97]. H19 expression was also higher in stage III and IV NSCLC, while hsa-miR-21 expression was higher in stage I and II NSCLC when compared to non-tumor lung tissues [97]. Besides, plasma levels of H19 were significantly increased in NSCLC patients when compared to patients with benign lung disease [89]. Additionally, a nucleotide polymorphism, the H19-rs217727 C>T, was found significantly associated with an increased risk of lung cancer [84]. Consequently, with further efforts to confirm these results in large independent cohorts, H19 would make a great biomarker to diagnose or to assess a genetic predisposition to lung cancers.
- ⋄
- H19 behavior against treatment: Wang et al. found a negative correlation between the H19 upregulation in NSCLC tumor tissues and the Cisplatin (DDP) response [96].
- ⋄
- Exosomal H19: Tumor-released lncRNA H19 (exo-H19) can promote gefitinib resistance via packaging into exosomes in NSCLCs. While the experiments were on gefitinib-resistant NSCLC cell lines, the authors assessed the expression of H19 in both gefitinib-resistant and parental sensitive cells. H19 expression was increased in gefitinib-resistant NSCLC cells and was described as secreted through the incorporation into exosomes, which was mediated by HNRNPA2B1. Moreover, exosome-mediated transfer of H19 conferred gefitinib resistance to the recipient NSCLC cells [148].
2.2. MEG3
2.2.1. MEG3 and IPF
2.2.2. MEG3 and Asthma
2.2.3. MEG3 and COPD
2.2.4. MEG3 and Lung Cancer
- ⋄
- ⋄
- MEG3 regulated genes: Two studies realized in NSCLC tumor tissues described MEG3 as an activator of TP53 by decreasing the levels of MDM2 [149,153]. Additionally, MEG3 can decrease BCL2 expression by promoting BAX, and also decrease MAP1LC3A expression [156,157]. Besides, Su et al. described MEG3 as inversely correlated with PCNA [153].
- ⋄
- MEG3 recruits the PRC2: In NSCLC cell lines, MEG3 contributes to the recruitment of PRC2’s EZH2, through a possible interaction with JARID2. This PRC2 recruitment will eventually repress the expression of CDH1, ZEB family (ZEB1 and ZEB2), and hsa-miR-200 family ( MIR200A, MIR200B, and MIR200C ), which will lead to a decreased EMT [158].
- ⋄
- MEG3 as ceRNA: Among its known functions, MEG3 was reported as a heavy miRNA regulator. Indeed, MEG3 can sequester: • hsa-miR-650 to release the expression of SLC34A2, in NSCLC cell line (H1299) [159] • hsa-miR-7-5p to release the expression of BRCA1, in NSCLC tumor tissues and BEAS-2B, A549, and HCC823 cell lines [156] • hsa-miR-21-5p to release the expression of SOX7, in DDP-resistant NSCLC tumor tissues and A549 and H1299 cell lines [117] • hsa-miR-3163 to release the expression of SKP2 that will, in turn, promotes the ubiquitination-associated degradation of p27 (CDKN1B), in NSCLC tumor tissues and A549 cell lines [153] • hsa-miR-205-5p to release the expression of LRP1. hsa-miR-205-5p may also be involved in the inhibition of TP53, p21 (CDKN1A), and caspase-3 (CASP3) expressions, in NSCLC tumor tissues and MEG3-knockdown NSCLC cell lines [154,157,160]. Moreover, using lncRNA-miRNA-mRNA regulatory network modules, Li et al. showed the following interactions in LUAD tumors from The Cancer Genome Atlas (TCGA). MEG3 and MIAT may interact with hsa-miR-106 (hsa-miR-106a or hsa-miR-106b), which then would regulate the expression of MAPK9. For a full overview of the Lung Adenocarcinoma (LUAD) miRNAs-lncRNAs-mRNAs network, please report to Li et al. Figure 2 [161].
- ⋄
- MEG3 at the clinical level: MEG3 low-expression in NSCLC tumor tissues was associated with short-term survival in two independent public datasets [106]. Besides, MEG3 genotype rs4081134 SNP (AA) was associated with a lung cancer risk in Chinese patients [85]. While being promising, these findings should, however, be confirmed in additional large independent cohorts to classify MEG3 as a reliable biomarker.
- ⋄
- MEG3 behavior against treatment: MEG3 can promote NSCLC cell lines sensitivity (A549 and H292) to Vincristine, by inhibiting autophagy. Indeed, autophagy level was higher in resistant cells, and the overexpression of MEG3 significantly reduced the expression of autophagy-related proteins LC3-I (MAP1LC3A), and LC3-II (MAP1LC3B) were [120]. The overexpression of MEG3 can also increase the DDP-sensitivity of NSCLC cell lines (A549 and H1299) [117] and xenografts [118] by decreasing TP53, CTNNB1, survivin (BIRC5), therefore targeting the WNT/-catenin signaling pathway [118], and Bcl-xl (BCL2L1) [119]. Furthermore, in Xu et al., the authors showed that Paclitaxel (PTX) could upregulate MEG3 and TP53, thus inhibiting cell proliferation and promoting the death of A549 cells [162].
- ⋄
- Exosomal MEG3: From the PubMed screening, we found no studies on MEG3 associated with lung diseases and exosomes. However, three recent studies could describe exosomal MEG3 in high-grade serous carcinoma [163], cervical cancer [164], and Hunner-type interstitial cystitis [165]. These studies underline that MEG3 may be involved in intercellular communication, especially in cancers, and therefore further research on this topic is needed to assess its relevance in lung diseases.
2.3. MALAT1
2.3.1. MALAT1 and Asthma
2.3.2. MALAT1 and COPD
2.3.3. MALAT1 and Lung Cancer
- ⋄
- Regulators of MALAT1: TDP-43 (TARDBP) can upregulate MALAT1 expression through direct interaction in NSCLC cell lines (A549 and YTLMC-9) [169]. Moreover, in the tumor tissues of NSCLC female patients, ESR2 can upregulate MALAT1 expression by binding to estrogen-response-element I and II on the proximal 2-kb region of MALAT1 promoter [172]. Additionally, in the NSCLC cell line (A549), Oct3/4 (POU5F1) and SP1 can increase MALAT1 expression by physically binding its promoter [178,180]. Furthermore, in NSCLC cell lines (SPC-A1 and H1299) in vitro and in vivo, TFAP2C and ZEB1 can upregulate MALAT1 expression, leading to the sequestration of miR-200b, which, in turn, increases of E2F3 and ZEB1, creating, therefore, a positive feedback loop [181]. Some miRNAs may also regulate MALAT1. Indeed, hsa-miR-142-3p can inhibit MALAT1 and WNT/-catenin signaling pathway in NSCLC tumor tissues and H1299 cell lines [75]. Moreover, hsa-miR-101-3p can inhibit MALAT1, BCL2, MMP9, PI3K (PIK3CA) expressions in NSCLC cell lines (H1299 and H520) [77].
- ⋄
- MALAT1 regulated genes: In NSCLC cell lines (A549 and H1299), MALAT1 can repress TP53 expression at the pre-mRNA level by binding a responsive region in the TP53 P1 promoter, leading to the downregulation of CDKN1A and FAS expressions [170]. In NSCLC cell lines (PC-9 and A549), MALAT1 also decreases cleaved-PARP1, cleaved-CASP3, and upregulates phospho-STAT3 [171], which in turn upregulates MDR1 (ABCB1) and MRP1 (ABCC1) [182]. Moreover, MALAT1 is involved in the upregulation of BCL2, MMP9, PIK3CA expressions, thus activating the PI3K/AKT signaling pathway in NSCLC tumor tissues and H1299 cell lines [77]. Furthermore, in NSCLC tumor tissues and cell lines (A549 and H1299), MALAT1 can upregulate VIM and downregulate CDH1 and is involved in the phosphorylation of AKT1, RPS6KB1, and MTOR [79,102]. Besides, in NSCLC tumor tissues and cell lines (A549, H661, and H460), MALAT1 can upregulate CXCL5, which in turn upregulates p-MAPK8 and down-regulates p-MAP2K1/2, p-MAPK3/1 proteins [176,183].
- ⋄
- MALAT1 as ceRNA: Among its known functions, MALAT1 was reported as a heavy miRNA regulator. Indeed, MALAT1 can sequester: • hsa-miR-145 to release the expressions of NEDD9 in the tumor tissues of NSCLC female patients [172], and KLF4 in NSCLC tumor tissues and cell lines (A549 and H1299) [184] • hsa-miR-204 to release the expression of SNAI2 in NSCLC tumor tissues and cell lines (A549, H1299, H460, and H446) [185] • hsa-miR-124-1 to release the expression of STAT3 and PI3K/AKT signaling pathway, in NSCLC cell lines (A549, H23, H522, H1299, H460) [78] • hsa-miR-206 and impact the AKT/MTOR signaling pathway in NSCLC tumor tissues and cell lines (A549 and H1299) [79] • hsa-miR-200b to release E2F3 and ZEB1 expressions, thus creating a positive feedback loop as ZEB1 can upregulate MALAT1 in NSCLC cell lines (SPC-A1 and H1299) in vitro and in vivo [181] • hsa-miR-200a-3p to release PD-L1 (CD274) expression in NSCLC tumor tissues and cell lines (A549 and CAL-12T) [186] • hsa-miR-429 to release Cyclin D1 (CCND1), MMP9, VIM, and CDH2 expression while repressing CDH1 expression in NSCLC tumor tissues [187] • hsa-miR-197-3p to release the Catenin-1 (CTNND1) expression in NSCLC tumor tissues and cell lines (A549, H1299, H460 and SPC-A-1) [188] • hsa-miR-101-3p to release the expressions of MCL1 in NSCLC tumor tissues and cell lines ((A549, H1299, H469, SPC-A1) [103], and SOX9 in NSCLC tumor tissues and cell lines (A549, H1299, HCC827, and H358) [76]. Moreover, in NSCLC tumor tissues and cell lines (A549, H1299, HCC827, and H358), SOX9 could activate MALAT1 expression by binding MALAT1 promoter on a specific site (5′-TCATTGTGT-3′), thus creating a positive feedback loop which dramatically increases MALAT1 downstream effects. Besides, SOX9 contributes to the upregulation of CTNNB1, a downstream target of MYC, thus activating the WNT/-catenin signaling pathway [76].
- ⋄
- MALAT1 at the clinical level: Several studies described a high MALAT1 level associated with a poor prognosis and short OS in NSCLC tumor tissues [76,79,101,102,103,105]. Besides, MALAT1 level was low in the serum of patients with NSCLC; however, it still lacks some specificity [90,91,92,93]. Furthermore, both MALAT1 and SOX9 expressions were associated with age, tumor size, and TNM stage, making these two genes potential candidates for prognosis tools [76,79,104].
- ⋄
- ⋄
- Exosomal MALAT1: Exosomal MALAT1 (exo-MALAT1) was described as upregulated in the serum of 77 NSCLC patients, and its expression was related to tumor stage and lymphatic metastasis [174]. While being a potential new biomarker for tumor stage diagnosis, further studies should be conducted on larger cohorts to confirm the predictive power of exo-MALAT1.
2.4. FENDRR
2.4.1. FENDRR and IPF
2.4.2. FENDRR and Lung Cancer
- ⋄
- ⋄
- FENDRR regulated genes: In NSCLC tumor tissues, FENDRR can specifically bind to the 3’UTR of ABCB1, thus blocking HuR (ELAVL1) binding to ABCB1 3’UTR, and therefore resulting in the decrease of ABCB1 expression [195]. Besides, Xu et al. found a negative correlation between FENDRR and ABCC10 expressions in NSCLC tumor tissues and cell lines (A549) [116].
- ⋄
- FENDRR as ceRNA: In NSCLC tumor tissues, cell lines (H1650, HCC827, A549, and H1975) and xenografts, FENDRR could also act as a miRNA regulator. Indeed, FENDRR can sequester hsa-miR-761 to release the expression of the metalloproteinase inhibitor TIMP2 [189,196]. Additionally, Liu et al. associated FENDRR to prognostic-significant ceRNA networks using TCGA-LUAD data. They also listed seven other molecules associated with these ceRNA networks; three mRNAs, EPAS1, FOXF1, and EDNRB, and four miRNAs, hsa-miR-148a, hsa-miR-195, hsa-miR-196b, and hsa-miR-301b). For an exhaustive overview of their FENDRR centered lncRNA–miRNA–mRNA ceRNA network, please refer to Figure 5 of their study [197].
- ⋄
- FENDRR at the clinical level: FENDRR low-expression in tumor tissues is strongly associated with TNM 1 stage in LUAD patients. Furthermore, when associated with LINC00312, FENDRR showed a diagnostic value in detecting these LUAD patients [88]. Nevertheless, to confirm the diagnostic power of FENDRR on the TNM 1 stage of LUAD patients, additional studies should be conducted on a broader spectrum of lung cancers, including different cancer subtypes and TNM stages.
- ⋄
- FENDRR behavior against treatment: The low expression of FENDRR observed in NSCLC tumor tissues was correlated with chemo-resistance to DDP [116]. However, additional studies are required to confirm this result.
- ⋄
- Exosomal FENDRR: From the PubMed search, we found no studies associating FENDRR to exosomes. However, FENDRR is an important lncRNA that controls the occurrence of metastasis. Indeed, low FENDRR expression was associated with distant metastasis and allowed the downregulation of the metalloproteinase inhibitor TIMP2 by the lack of competition with hsa-miR-761. Therefore, the metalloproteinase can degrade the extracellular matrix (ECM) and facilitates tumor metastasis [196]. It would, therefore, be interesting to seek for FENDRR in the extracellular vesicles of lung cancer associated with distant metastasis.
2.5. TUG1
2.5.1. TUG1 and Asthma
2.5.2. TUG1 and COPD
2.5.3. TUG1 and Lung Cancer
- ⋄
- Regulators of TUG1: Zhang et al. found that TP53 can regulate TUG1 expression in NSCLC tumor tissues and SPC-A1 cell line in vitro and in vivo [105,107]. This result requires, however, an independent validation. ⋄ TUG1 regulated genes: In NSCLC tumor tissues, TUG1 has been described to trans-downregulate the expression of Homeobox B7 (HOXB7), CELF1, and EZH2 (PRC2 subunit). Moreover, TUG1 decrease was also significantly associated with the differential expression of the following target genes. In both LUAD and SCC, TUG1 downregulation was associated with the downregulation of ELAVL1, PTBP1, IGF2BP1, IGF2BP2, IGF2BP3, PUM2, TNRC6A, DGCR8, FMR1, FXR1, FUS, MOV10, ZC3H7B, EWSR1, FUS-mutant, SRSF1, U2AF2, UPF1, and TARDBP [95]. It was otherwise associated with the upregulation of HNRNPC [95]. Conversely, in SCLCs, TUG1 may silence LIMK2 and BAX expression by interacting with EZH2 [86,105,107,108,198,199,200].
- ⋄
- ⋄
- TUG1 as ceRNA: In NSCLC tumor tissues, cell lines (SPC-A1, NCI-H520, NCI-H520, NCI-H1299) and xenograft, Guo et al. found that TUG1 can sequester hsa-miR-221, thus releasing the expression of PTEN [83].
- ⋄
- ⋄
- TUG1 behavior against treatment: The low expression of TUG1 observed in NSCLC cells was associated with chemo-resistance to DDP. When overexpressed, TUG1 promoted the sensitivity of NSCLC cells to DDP, leading to apoptosis, in vitro, and in vivo [83].
- ⋄
- Exosomal TUG1: From the PubMed search, we found no studies associating TUG1 with lung diseases and exosomes. However, two recent studies mentioned exosomal TUG1, in MCF-7 cells, the levels of TUG1 were moderately elevated in exosomes when compared to cells [201], and TUG1 was up-regulated in the serum exosomes of colorectal cancer (CRC) patients [202]. Besides, Guo et al. described TUG1 as ceRNA to hsa-miR-221, which enables PTEN expression [83]. Together, these results suggest that TUG1 may be involved in intercellular communication to synchronize cellular proliferation. Therefore, further research on this topic is needed to assess its relevance in lung cancers.
2.6. CDKN2B-AS1, ANRIL
2.6.1. CDKN2B-AS1 and IPF
2.6.2. CDKN2B-AS1 and Asthma
2.6.3. CDKN2B-AS1 and COPD
2.6.4. CDKN2B-AS1 and Lung Cancer
- ⋄
- Regulators of CDKN2B-AS1: In NSCLC tumor tissues and cell lines (A549, SPC-A1, and NCI-H1975), MYC can upregulate CDKN2B-AS1 by physically interacting with the c-Myc-responsive element (E-box) of CDKN2B-AS1 promoter [87,208]. Besides, Midkine (MDK) was described as often upregulated in the tumor microenvironment of SCC tumor tissues and cell lines (SCC4, OSCC3, HSC3, and CAL27). The authors also found that MDK can upregulate the CDKN2B-AS1 expression [121].
- ⋄
- DKN2B-AS1 regulated genes: In NSCLC tumor tissues, Alsibai et al. found a strong positive correlation between the expressions of CDKN2B-AS1 and the tumor suppressors p15-CDKN2B and p14-CDKN2A, but not p16-CDKN2A. Interestingly, expressed CDKN2B-AS1 can stabilize the PRC complexes to repress the expression of p15, p14, and p16, leading to activate the cell cycle [207]. Few additional studies described the ability of CDKN2B-AS1 to decrease the expression of PARP1, cleaved-PARP1, and cleaved-CASP3, and to increase BCL2 and CASP3 expressions [87,121].
- ⋄
- ⋄
- CDKN2B-AS1 at the clinical level: Lin et al. found a positive correlation between CDKN2B-AS1 high-expression and the differentiation grade and TNM stages in LUAD [87]. CDKN2B-AS1 high-expression in NSCLC tumor tissues was also associated with poor patient OS [94]. Intriguing results from Du et al. suggested that a low CDKN2B-AS1 expression in the peripheral blood of IPF patients may promote the occurrence of lung cancers by regulating the P53 signaling pathway [63]. However, further investigations should be conducted on IPF patients that developed lung cancer to confirm this hypothesis. Interestingly, CDKN2B-AS1 SNPs were strongly associated with the risk of developing a LUAD [210,211,212,213,214].
- ⋄
- CDKN2B-AS1 behavior against treatment: CDKN2B-AS1 high expression increases PTX resistance of A549 cells [87]. Interestingly, Zhang et al. showed that cancer-associated fibroblasts (CAFs) contribute to the high level of MDK in the tumor micro-environment of Oral Squamous Cell Carcinoma (OSCC) tissues, thus promoting a DDP resistance via a high expression of CDKN2B-AS1 [121].
- ⋄
- Exosomal CDKN2B-AS1: From the PubMed search, we did not find studies associating CDKN2B-AS1 with lung diseases and exosomes. Moreover, only one recent study mentioned exosomal CDKN2B-AS1 as significantly higher in the urine of BC patients when compared to healthy subjects [215]. However, CDKN2B-AS1 is an important lncRNA that can decrease the expression levels of PARP1, which plays a crucial role in DNA repair [87,121]. Since the alteration of the DNA repair mechanism is part of the hallmark of cancers, it would be interesting to seek for CDKN2B-AS1 in the extracellular vesicles of early-stage lung tumors.
2.7. HOTAIR
2.7.1. HOTAIR and COPD
2.7.2. HOTAIR and Lung Cancer
- ⋄
- Regulators of HOTAIR: In the NSCLC cell line A549, HOTAIR was described as upregulated by hypoxia and CSE [216,217], and is a direct target of HIF-1 (HIF1A), which acts through interaction with putative hypoxia-responsive elements (HREs) in the upstream region of HOTAIR [216]. Besides, pro-inflammatory IL6 can activate STAT3 in an autocrine path, and STAT3 will then increase HOTAIR expression by interacting with its promoter [217]. In NSCLC tumor tissues and A549 cell line, Caveolin 1 (CAV1) was described to upregulate HOTAIR [219]. Interestingly, a specific negative regulation loop involves HOTAIR and TP53 in NSCLC tumor tissues. Indeed, two TP53 binding sites were found on HOTAIR’s promoter and can suppress HOTAIR transcription after TP53 binding. HOTAIR can, in turn, modify the promoter of TP53 by increasing H3K27me3 leading to TP53 repression [220].
- ⋄
- HOTAIR regulated genes: In SCLC cell lines (H69 and H446), HOTAIR can activate the NF-κB signaling pathway through the methylation of HOXA1 [73]. Indeed, HOTAIR regulates the HOXA1 methylation level by decreasing DNMT1 and DNMT3B expression [115]. In an NSCLC cell line (PC9), HOTAIR is involved in cellular growth with p65 (RELA), DNMT1, and EZH2. Moreover, HOTAIR can inhibit JUN and CDKN1A [221]. Furthermore, in NSCLC tumors tissues and PC9 cell line, HOTAIR can activate WNT3A, CTNNB1, APC, ABCC1, and ABCB1, and can also promote the expression of 14-3-3 (SFN) [222,223]. In NSCLC tumor tissues and A459 cell line, HOTAIR would also upregulate CSC-related biomarkers such as NANOG, POU5F1, SOX2, MYC, CTNNB1, and KLF4 [113]. In NSCLC tumor tissues and cell lines (A549, PC9, H1299, and H520), Besides, HOTAIR was associated with LSH (HELLS) to regulate the FOXA1 to FOXA2 ratio and promote cell migration and invasion. Importantly, HELLS regulates this ratio by binding to the promoter of FOXA1, not FOXA2 [224]. Additionally, in Lung cancer cell lines (A549, H460, H1299, NCI-H460, HCC-827), HOTAIR regulates the expression of BECN1, phospho-ULK1, and the LC3II/I (MAP1LC3A / MAP1LC3B) ratio [81].
- ⋄
- HOTAIR recruits the PRC2: Fang et al. recently described a negative feedback regulator loop involving HOTAIR in SCLC cell lines (NCI-H69 and NCI-H446). Indeed, HOTAIR may upregulate EZH2 and H3K27me3 levels, which in turn can repress HOTAIR, leading to change HOXA1 methylation [225]. Interestingly, both HOTAIR 5’ and 3’ ends may be involved in the cell cycle dysregulation. Indeed, in NSCLC cell lines (95C, 95D, and YTMLC-90), Liu et al. demonstrated that RB1 and E2F1 are both regulated by HOTAIR5’ via the PRC2 (EZH2, SUZ12, and EED) complex and by HOTAIR3’ via the LSD1/ CoREST/ REST complex. Both complexes may, therefore, act on the WNT/-catenin signaling pathway and promote EMT when coupled with histone H3 lysine 27 methylation and lysine 4 demethylation [80].
- ⋄
- HOTAIR as ceRNA: Among its known functions, HOTAIR was reported as a miRNA regulator. Indeed, HOTAIR can sequester: • hsa-miR-214-3p to release the expression of PDPK1, in NSCLC cell lines (A549 and PC9 cells) [226] • hsa-miR-217 to release the expression of DACH1, in NSCLC cell lines (H23, H292, H1299, and A549) [227] • hsa-miR-326 to release the expression of SP1, in NSCLC tumor tissues and A549 cell line [228], and PHOX2A in NSCLC cell lines (A549, 95D, NCI-H460, HLamp, and H838) [229]. HOTAIR may also sequester hsa-miR-613 in NSCLC tumor tissues and cell lines (H1299, H23, H292, and A549) [230], and hsa-miR-221 in NSCLC tumor tissues and cell lines (A549, H322, and H1299) [100].
- ⋄
- HOTAIR at the clinical level: HOTAIR high expression in NSCLC tumor tissues coincides with greater tumor size, advanced TNM stage, lymph node metastasis or lymph-vascular invasion, and short disease-free interval [98]. Its expression was also related to a reduced OS in NSCLC tumor tissues [99]. Furthermore, the expressions of HOTAIR in patients with stage I and II were lower than those with stage III and IV NSCLC tumors [100]. Besides, both H19 and HOTAIR were identified as non-invasive diagnostic biomarkers in the sputum of lung cancer patients [109]. The diagnosis of head-and-neck squamous cell carcinoma (HNSCC) can also be improved by combining the high expression of HOTAIR to the high expression of CASC9 [231]. Altogether, these encouraging results underline the potential of HOTAIR as a diagnostic biomarker.
- ⋄
- HOTAIR behavior against treatment: HOTAIR high-expression contributes to DDP resistance via CDKN1A downregulation in LUAD tumor tissues, and experimental downregulated HOTAIR in A549 cells promoted DDP sensitivity [112,113]. It also contributes to Atractylenolide 1 and Erlotinib resistances by activating PDK1 and EZH2, in LUAD cells, in vitro and in vivo [114]. Besides, the downregulation of HOTAIR can increase the SCLC cell lines’ sensitivity to DDP, Adriamycin, and Etoposide, through decreasing DNMT1 and DNMT3B expressions, leading to the reduction of HOXA1 methylation [115]. Furthermore, the downregulation of HOTAIR can increase NSCLC cell lines sensitivity (A549, H460, H1299, NCI-H460, HCC-827) to Crizotinib through the inhibition of ULK1-phosphorylation. This sensitivity leads to the suppression of tumor growth and triggers the cell cycle arrest and the apoptosis signaling pathway [81].
- ⋄
- Exosomal HOTAIR: Exosomal HOTAIR (exo-HOTAIR) appeared in the exosomes from bronchoalveolar lavage (BAL) of smokers, NSCLC, and healthy patients, but without significant differences between the three conditions [232]. However, in a recent letter to editors, Zhang et al. found exo-HOTAIR more expressed in the blood samples from LCC patients when compared to LUAD or SCC patients. Moreover, these authors described that A549 and H1299 cells treated with exo-HOTAIR increased the level of cellular HOTAIR. The authors concluded that exo-HOTAIR promotes proliferation, migration, and invasion of the cells through the sequestration of hsa-miR-203 (hsa-mir-203a or hsa-mir-203b) [233]. Interestingly, this interaction between hsa-miR-203 and HOTAIR was also reported with similar effects in renal cell carcinoma cells. In this study, Dasgupta et al. described that the sequestration of hsa-miR-203 decreases CDH1, PTEN, CDKN1A, and CDKN1B levels, while it increases the expression of VIM [234].
2.8. GAS5
2.8.1. GAS5 and Asthma
2.8.2. GAS5 and Lung Cancer
- ⋄
- GAS5 regulated genes: In NSCLC tumor tissues, GAS5 decrease correlates strongly and significantly to the decrease of FAS-AS1 and THRIL in male donors and combined tumors, as well as the increase of NEAT1 in male donors and combined tumors. GAS5 decrease also correlates with TUG1 increase in female donors and combined tumors, and with PVT1 increase in female donors when compared to ANCTs. GAS5 expression is also associated with the upregulation of IGF2BP2 and the downregulation of FXR1 [95]. Moreover, GAS5 downregulates the expression of TNRC6A, ZC3H7B, and UPF1, while it can upregulate the expression of EIF4A3, TIA1, TIAL1, and HNRNPC [95]. Besides, in NSCLC tumor tissues and cell lines (A549, H1299, H1975, HCC827), GAS5 can deregulate the expression of phospho-EGFR, phospho-MAPK1, phospho-AKT1, and IGF1R. Interestingly, GAS5 overexpression inversely correlates with the activation of the EGFR pathway [110].
- ⋄
- GAS5 as ceRNA: Among its known functions, GAS5 can act as a miRNA regulator. Indeed, GAS5 may sequester hsa-miR-21-5p in NSCLC tumor tissues and the cell lines (NCI-H460, A549, NCI-H1299, H460, SK-MES-1, H157, and H358) [82,111]. GAS5 may also sequester hsa-miR-205-5p in the NSCLC cell lines (A549, H460, 95D, H1299, SPC-A-1, and H522) [235]. Both hsa-miR-21-5p and hsa-miR-205-5p sequestration would release the expression of PTEN [82,111,235]. Additionally, GAS5 suspected of sequestering hsa-miR-135b-5p in NSCLC tumor tissues and cell lines (A549 and H1975) [236]. It is also suspected to sequester hsa-miR-23a in NSCLC tumor tissues and cell lines (A549, H838, H157, and HCC827) [237].
- ⋄
- GAS5 at the clinical level: Esfandi et al. emphasized the GAS5 low expression in tumor tissues as a promising biomarker for the diagnosis of the NSCLCs [95].
- ⋄
- GAS5 behavior against treatment: GAS5 may regulate chemo-resistance to DDP of NSCLC tumor tissues and cell lines (H460 and H157), through the PTEN signaling pathway [82]. Besides, GAS5 low expression contributes to resistance to gefitinib to LUAD cell lines and tumor tissues [110]. Furthermore, its low expression can promote the resistance to ionizing radiation in NSCLC cell lines [111] and tumor tissues [236].
- ⋄
- Exosomal GAS5: In Cheng et al. study on urethane-induced lung cancer mouse model, lung cancer-derived exosomal GAS5 (exo-GAS5) affects the proliferation, apoptosis, and tube formation of human umbilical vein endothelial cells (HUVECs). The overexpression of GAS5 leads to an increase of exo-GAS5, upregulates PTEN expression, and inhibits the phosphorylation of PI3K/AKT, through hsa-miR-29b-3p sequestration [238]. Furthermore, exo-GAS5 expression was lower in the serum of 64 NSCLC patients when compared to healthy controls. This low expression of exo-GAS5 was associated with larger tumor size and advanced TNM [239]. While being a potential new biomarker for the diagnosis of Stage I NSCLCs, further studies should be conducted on larger cohorts to confirm the predictive power of exo-GAS5.
3. Additional LncRNAs Not Yet Described in Lung Cancer Studies
3.1. LINC00861
3.1.1. LINC00861 in Asthma
3.1.2. LINC00861 in COPD
3.2. CCDC18-AS1, RP4-717I23.3
3.2.1. CCDC18-AS1 in Asthma
3.2.2. CCDC18-AS1 in COPD
4. Additional Interesting Exosomal-LncRNAs Described with Lung Cancer
4.1. Exo-UCA1
4.2. Exo-lncMMP2-2
4.3. Exo-GAPLINC
4.4. Exo-TBILA and Exo-AGAP2-AS1
4.5. Exo-SOX2-OT
5. Concluding Remarks and Future Perspectives
- ⋄
- Genome instability and mutation: MALAT1 and CDKN2B-AS1 may be key players of the “Genome instability and mutation” hallmark since they can decrease the expression levels of PARP1 [87,121,171]. H19 may also contribute to the “Genome instability and mutation” as well as the “Evading growth suppressors” hallmarks. CSC exposure induces an overall increase of H3K27me3 levels, which would repress many genes [74]. However, RIOX2 may demethylate H19 before the DNA-repair gene MGMT and the cyclin-dependent kinase inhibitor p16-CDKN2A are methylated [74,139]. Subsequently, a decreased level of MGMT would lead to chromosomal alterations, while a decrease in p16-CDKN2A would lead to inhibit the cell cycle arrest in G1 and G2 phases.
- ⋄
- Activating invasion and metastasis: FENDRR may be a key player of the “Activating invasion and metastasis” hallmark. The low FENDRR expression observed in NSCLC tumor tissues allows the ECM degradation by the metalloproteinases and thus facilitates the metastasis. Indeed, under normal conditions, FENDRR is supposed to sequester hsa-miR-761, which will permit the increase of the metalloproteinase inhibitor TIMP2, leading to the degradation of the extracellular matrix [196].
- ⋄
- Resisting cell death: MALAT1, MEG3, and CDKN2B-AS1 may be key actors of the “Resisting cell death” hallmark. CDKN2B-AS1 decreases cleaved-CASP3 while increasing BCL2 and CASP3 expression [87,121]. High levels of MEG3 reduces CASP3 through hsa-miR-205-5 sequestration [154,157,160]. These high levels can also decrease the expression of BIRC5 [118]. MALAT1 can also decrease cleaved-CASP3 levels [171]. Subsequently, in NSCLCs, cleaved-CASP3 is decreased, while BCL2, BIRC5, and CASP3 is increased, which leads to escape apoptosis.
- ⋄
- Sustaining and proliferative signaling: GAS5, TUG1, MALAT1, H19, and HOTAIR may be key actors of the “Sustaining and proliferative signaling” hallmark in NSCLCs. High GAS5 and low HOTAIR levels combined with EGFR inhibitors, increase the sensitivity to treatment [110,114]. GAS5, TUG1, H19, and MALAT1 are upstream regulators of the PTEN/PI3K/AKT signaling pathway. H19 recruits EZH2 to repress PTEN expression [135]. MALAT1 is involved in the upregulation of PIK3CA [77] and phospho-STAT3 [171], and in the phosphorylation of AKT1 and MTOR [79,102]. Finally, TUG1 and GAS5 can release PTEN expression, respectively, through hsa-miR-221 and hsa-miR-21-5p, hsa-miR-205-5p sequestration [82,83,111,235]. Subsequently, in NSCLCs, PTEN is downregulated, while PIK3CA, phospho-STAT3, phospho-AKT1, and phospho-MTOR are increased, thus enhancing the cellular proliferation.
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Abbreviation | Full name |
| AECOPD | acute exacerbations of COPD |
| ANCT | Adjacent Non-Cancerous Tissue |
| ASM | Airway Smooth Muscle |
| BA-E | Bronchial asthma at exacerbation |
| BA-R | Bronchial asthma at remission |
| BEAS-2B | Bronchial epithelial cells |
| ceRNA | competing endogenous RNA |
| CDK | Cyclin Dependent Kinase |
| COPD | Chronic Obstructive Pulmonary Disease |
| CS | Cigarette Smoke |
| CSC | Cancer Stem Cell |
| CSE | Cigarette Smoke Extract |
| DDP | Cisplatine |
| EMT | Epithelial Mesenchymal Transition |
| FEV_1 | Forced expiratory volume in 1 second |
| FFMI | Fat-free mass index |
| FVC | Forced vital capacity |
| GINA | Global Initiative for Asthma |
| H3K9me3 | H3 lysine 9 tri-methylation |
| H3K27me3 | H3 lysine 27 tri-methylation |
| HBE | Human Bronchial Epithelial cells |
| HRE | Hypoxia-Responsive Element |
| HNSCC | head-and-neck squamous cell carcinoma |
| IPF | Idiopathic Pulmonary Fibrosis |
| LD | linear dichroism |
| LDL | low-density lipoprotein |
| LSCC | Lung Squamous Cell Carcinoma |
| lncRNA | long non-coding RNA |
| LTB4 | Leukotriene B4 |
| LUAD | Lung Adenocarcinoma |
| MET | Mesenchymal Epithelial Transition |
| NAT | Natural Antisense transcripts |
| NSCLC | Non Small Cell Lung Cancer |
| OS | Overall Survival |
| OSCC | Oral Squamous Cell Carcinoma |
| PRC | Polycomb Repressive Complex |
| PTX | Paclitaxel |
| SCC | Squamous Cell Carcinoma |
| SCLC | Small Cell Lung Cancer |
| SNP | Single Nucleotide polymorphism |
| TCGA | The Cancer Genome Atlas |
| TNM | Tumor-Node–Metastasis: |
| TNM | T: Extent of the primary tumor, |
| TNM | N: lymph node involvement, |
| TNM | M: metastatic disease |
| WHO | World Health Organization |
| Signaling Pathway | KEGG ID |
| AKT/MTOR signaling pathway | hsa04150 |
| Apoptosis signaling pathway | hsa04210 |
| EGFR pathway | hsa01521 |
| NF-B signaling pathway | hsa04064 |
| PTEN/PI3K/AKT signaling pathway | hsa04151 |
| TGFB/SMAD3 signaling pathway | hsa04350 |
| T-cell receptor signaling pathway | hsa04660 |
| Th17 cell differentiation | hsa04659 |
| P53 signaling pathway | hsa04115 |
| WNT/-catenin signaling pathway | hsa04310 |
| Mirbase ID | HGNC symbol | Mirbase accession |
| hsa-miR-101-3p | MIR101-1 | MIMAT0000099 |
| hsa-miR-106a | MIR106A | MI0000113 |
| hsa-miR-106b | MIR106B | MI0000734 |
| hsa-miR-107 | MIR107 | MI0000114 |
| hsa-miR-124-1 | MIR124-1 | MI0000443 |
| hsa-miR-125a | MIR125A | MI0000469 |
| hsa-miR-135b-5p | MIR135B | MIMAT0000758 |
| hsa-miR-140 | MIR140 | MI0000456 |
| hsa-miR-142-3p | MIR142 | MIMAT0000434 |
| hsa-miR-145 | MIR145 | MI0000461 |
| hsa-miR-145-5p | MIR145 | MIMAT0000437 |
| hsa-miR-148a | MIR148A | MI0000253 |
| hsa-miR-15a | MIR15A | MI0000069 |
| hsa-miR-195 | MIR195 | MI0000489 |
| hsa-miR-196b | MIR196B | MI0001150 |
| hsa-miR-197-3p | MIR197 | MIMAT0000227 |
| hsa-miR-17 | MIR17 | MI0000071 |
| hsa-miR-196a-1 | MIR196A1 | MI0000238 |
| hsa-miR-196a-2 | MIR196A2 | MI0000279 |
| hsa-miR-200 | MIR200 family | hsa-miR-200 family |
| hsa-miR-200a | MIR200A | MI0000737 |
| hsa-miR-200a-3p | MIR200A | MIMAT0000682 |
| hsa-miR-200b | MIR200B | MI0000342 |
| hsa-miR-200c | MIR200C | MI0000650 |
| hsa-miR-203a | MIR203A | MI0000283 |
| hsa-miR-203b | MIR203B | MI0017343 |
| hsa-miR-204 | MIR204 | MI0000284 |
| hsa-miR-205-5p | MIR205 | MIMAT0000266 |
| hsa-miR-206 | MIR206 | MI0000490 |
| hsa-miR-21 | MIR21 | MI0000077 |
| hsa-miR-21-5p | MIR21 | MIMAT0000076 |
| hsa-miR-210 | MIR210 | MI0000286 |
| hsa-miR-214 | MIR214 | MI0000290 |
| hsa-miR-214-3p | MIR214 | MIMAT0000271 |
| hsa-miR-217 | MIR217 | MI0000293 |
| hsa-miR-218-1 | MIR218-1 | MI0000294 |
| hsa-miR-218-5p | MIR218-1 | MIMAT0000275 |
| hsa-miR-218-2 | MIR218-2 | MI0000295 |
| hsa-miR-221 | MIR221 | MI0000298 |
| hsa-miR-23a | MIR23A | MI0000079 |
| hsa-miR-29b-3p | MIR29B1 | MIMAT0000100 |
| hsa-miR-301b | MIR301B | MI0005568 |
| hsa-miR-3163 | MIR3163 | MI0014193 |
| hsa-miR-326 | MIR326 | MI0000808 |
| hsa-miR-33a-5p | MIR33A | MIMAT0000091 |
| hsa-miR-409-3p | MIR409 | MIMAT0001639 |
| hsa-miR-429 | MIR429 | MI0001641 |
| hsa-miR-484 | MIR484 | MI0002468 |
| hsa-miR-519a-1 | MIR519A1 | MI0003178 |
| hsa-miR-519a-2 | MIR519A2 | MI0003182 |
| hsa-miR-590-5p | MIR590 | MIMAT0003258 |
| hsa-miR-613 | MIR613 | MI0003626 |
| hsa-miR-650 | MIR650 | MI0003665 |
| hsa-miR-675 | MIR675 | MI0005416 |
| hsa-miR-675-5p | MIR675 | MIMAT0004284 |
| hsa-miR-6515-3p | MIR6515 | MIMAT0025487 |
| hsa-miR-7-5p | MIR7-1 | MIMAT0000252 |
| hsa-miR-761 | MIR761 | MI0003941 |
| hsa-let-7 | MIRLET7 family | hsa-let-7 family |
| hsa-let-7d-5p | MIRLET7D | MIMAT0000065 |
| HGNC symbol | Alias | Full name | Type | Accession ID |
| AGAP2-AS1 | AGAP2 Antisense RNA 1 | NAT | ENSG00000255737 | |
| CCDC18-AS1 | CCDC18 Antisense RNA 1 | NAT | ENSG00000223745 | |
| CDKN2B-AS1 | ANRIL | CDKN2B Antisense RNA 1 | NAT | ENSG00000240498 |
| FAS-AS1 | FAS Antisense RNA 1 | NAT | HGNC:37128 | |
| FENDRR | FOXF1 adjacent non-coding | |||
| developmental regulatory RNA | NAT | ENSG00000268388 | ||
| GAPLINC | RP11-838N2.4 | Gastric adenocarcinoma associated, | ||
| positive CD44 regulator, lincRNA | lincRNA | ENSG00000266835 | ||
| GAS5 | Growth Arrest Specific 5 | NAT | ENSG00000234741 | |
| H19 | H19 Imprinted Maternally Expressed Transcript | NAT, | ENSG00000130600 | |
| lincRNA | ||||
| HAGLR | HOXD Antisense Growth-Associated lncRNA | NAT | ENSG00000224189 | |
| HOTAIR | HOX Transcript Antisense RNA | NAT | ENSG00000228630 | |
| HOTAIRM1 | HOXA Transcript Antisense RNA, | |||
| Myeloid-Specific 1 | NAT | ENSG00000233429 | ||
| HOTTIP | HOXA Distal Transcript Antisense RNA | NAT | ENSG00000243766 | |
| HOXA-AS3 | HOXA Cluster Antisense RNA 3 | NAT | ENSG00000254369 | |
| HOXA10-AS | HOXA10 Antisense RNA | NAT | ENSG00000253187 | |
| HOXA11-AS | HOXA11 Antisense RNA | NAT | ENSG00000240990 | |
| LINC00312 | Long Intergenic Non-Protein Coding RNA 312 | lincRNA | HGNC:6662 | |
| LINC00861 | Long Intergenic Non-Protein Coding RNA 861 | lincRNA | ENSG00000245164 | |
| - | lnc-MMP2-2 | lnc-MMP2-2-1_dup1 | lincRNA | NONHSAT142627 |
| MEG3 | Maternally Expressed 3 | NAT | ENSG00000214548 | |
| MALAT1 | Metastasis Associated LUAD Transcript 1 | NAT | ENSG00000251562 | |
| NEAT1 | Nuclear Paraspeckle Assembly Transcript 1 | lincRNA | ENSG00000245532 | |
| PVT1 | Pvt1 Oncogene | NAT | ENSG00000249859 | |
| THRIL | TNF And HNRNPL Related | |||
| Immunoregulatory lncRNA | lincRNA | ENSG00000280634 | ||
| TUG1 | Taurine Up-Regulated 1 | lincRNA | ENSG00000253352 | |
| TBILA | TGF-Beta Induced LncRNA | lincRNA | ENSG00000261488 | |
| SOX2-OT | SOX2 Overlapping Transcript | NAT | ENSG00000242808 | |
| UCA1 | Urothelial Cancer Associated 1 | lincRNA | ENSG00000214049 | |
| ZEB1-AS1 | ZEB1 Antisense RNA 1 | NAT | ENSG00000237036 |
| HGNC symbol | Alias | Full name | Accession ID |
| ABCB1 | MDR1 | ATP Binding Cassette Subfamily B Member 1 | ENSG00000085563 |
| ABCC1 | MRP1 | ATP Binding Cassette Subfamily C Member 1 | ENSG00000103222 |
| ABCC10 | ATP binding cassette subfamily C member 10 | ENSG00000124574 | |
| ACO1 | IRP1 | aconitase 1 | ENSG00000122729 |
| ACTA2 | SMA | Actin 2, Smooth Muscle | ENSG00000107796 |
| AGO1 | Argonaute RISC Catalytic Components 1 | ENSG00000092847 | |
| AGO2 | Argonaute RISC Catalytic Components 2 | ENSG00000123908 | |
| AGO3 | Argonaute RISC Catalytic Components 3 | ENSG00000126070 | |
| AGO4 | Argonaute RISC Catalytic Components 4 | ENSG00000134698 | |
| AKT1 | Akt | AKT Serine/Threonine Kinase 1 | ENSG00000142208 |
| APC | APC Regulator Of WNT Signaling Pathway | ENSG00000134982 | |
| BAX | BCL2 associated X, apoptosis regulator | ENSG00000087088 | |
| BCL2 | Bcl-2 | BCL2 apoptosis regulator | ENSG00000171791 |
| BCL2L1 | Bcl-xl | BCL2 Like 1 | ENSG00000171552 |
| BECN1 | Beclin1 | ENSG00000126581 | |
| BIRC5 | survivin | Baculoviral IAP Repeat Containing 5 | ENSG00000089685 |
| BRCA1 | BRCA1 DNA Repair Associated | ENSG00000012048 | |
| CASC9 | Cancer Susceptibility 9 | ENSG00000249395 | |
| CASP3 | caspase-3 | Caspase 3 | ENSG00000164305 |
| CAV1 | CAV-1 | Caveolin 1 | ENSG00000105974 |
| CCND1 | Cyclin D1 | ENSG00000110092 | |
| CELF1 | CUGBP Elav-Like Family Member 1 | ENSG00000149187 | |
| CD274 | PD-L1 | CD274 Molecule | ENSG00000120217 |
| CDH1 | E-cadherin | Cadherin 1 | ENSG00000039068 |
| CDH13 | Cadherin 13 | ENSG00000140945 | |
| CDH2 | N-cadherin | Cadherin 2 | ENSG00000170558 |
| CDKN1A | p21 | CDK Inhibitor 1A | ENSG00000124762 |
| CDKN1B | p27 | CDK Inhibitor 1B | ENSG00000111276 |
| CDKN2A | p14, p16, p19 | CDK Inhibitor 2A | ENSG00000147889 |
| CDKN2B | p15 | CDK Inhibitor 2B | ENSG00000147883 |
| COL1A1 | Collagen type I 1 chain | ENSG00000108821 | |
| CBX7 | Chromobox 7 | ENSG00000100307 | |
| CTNNB1 | -catenin | Catenin 1 | ENSG00000168036 |
| CTNND1 | Catenin 1 | ENSG00000198561 | |
| CXCL5 | C-X-C Motif Chemokine Ligand 5 | ENSG00000163735 | |
| CXCL8 | IL8 | C-X-C Motif Chemokine Ligand 8 | ENSG00000169429 |
| DACH1 | Dachshund Family Transcription Factor 1 | ENSG00000276644 | |
| DAPK1 | DAPK | Death Associated Protein Kinase 1 | ENSG00000196730 |
| DGCR8 | DGCR8 Microprocessor Complex Subunit | ENSG00000128191 | |
| DLK1 | Like Non-Canonical Notch Ligand 1 | ENSG00000185559 | |
| DIO3 | Iodothyronine Deiodinase 3 | ENSG00000197406 | |
| DNMT1 | DNA Methyltransferase 1 | ENSG00000130816 | |
| DNMT3B | DNMT3b | DNA Methyltransferase 3 | ENSG00000088305 |
| DUSP6 | Dual Specificity Phosphatase 6 | ENSG00000139318 | |
| E2F1 | E2F Transcription Factor 1 | ENSG00000101412 | |
| E2F3 | E2F Transcription Factor 1 | ENSG00000112242 | |
| EDNRB | Endothelin Receptor Type B | ENSG00000136160 | |
| EED | Embryonic Ectoderm Development | ENSG00000074266 | |
| EGFR | Epidermal Growth Factor Receptor | ENSG00000146648 | |
| EIF4A3 | eIF4AIII | Eukaryotic Translation Initiation Factor 4A3 | ENSG00000141543 |
| ELAVL1 | HuR | ELAV Like RNA Binding Protein 1 | ENSG00000066044 |
| EPAS1 | Endothelial PAS Domain Protein 1 | ENSG00000116016 | |
| ESR2 | ER | Estrogen Receptor 2 | ENSG00000140009 |
| EWSR1 | EWS RNA Binding Protein 1 | ENSG00000182944 | |
| EZH1 | Enhancer Of Zeste 1 PRC2 Subunit | ENSG00000108799 | |
| EZH2 | Enhancer Of Zeste 2 PRC2 Subunit | ENSG00000106462 | |
| FAS | Fas Cell Surface Death Receptor | ENSG00000026103 | |
| Fgf1 | Fibroblast growth factor 1 | ENSRNOG00000013867 | |
| FMR1 | FMRP | FMRP Translational Regulator 1 | ENSG00000102081 |
| FXR1 | FMR1 Autosomal Homolog 1 | ENSG00000114416 | |
| FN1 | Fibronectin 1 | ENSG00000115414 | |
| FOXA1 | FoxA1 | Forkhead Box A1 | ENSG00000129514 |
| FOXA2 | FoxA2 | Forkhead Box A2 | ENSG00000125798 |
| FOXF1 | Forkhead Box F1 | ENSG00000103241 | |
| FOXF2 | Forkhead Box F2 | ENSG00000137273 | |
| FOXN1 | Forkhead Box N1 | ENSG00000109101 | |
| FOXP3 | Forkhead Box P3 | ENSG00000049768 | |
| FUS | FUS RNA Binding Protein | ENSG00000089280 | |
| HELLS | LSH | Helicase, Lymphoid Specific | ENSG00000119969 |
| HES1 | hes family bHLH transcription factor 1 | ENSG00000114315 | |
| HEY1 | hes related family bHLH transcription factor | ||
| with YRPW motif 1 | ENSG00000164683 | ||
| HIF1A | HIF-1 | Hypoxia Inducible Factor 1 Subunit | ENSG00000100644 |
| HOXA1 | Homeobox A1 | ENSG00000105991 | |
| HOXA10 | Homeobox A10 | ENSG00000253293 | |
| HOXA11 | Homeobox A11 | ENSG00000005073 | |
| HOXB7 | Homeobox B7 | ENSG00000260027 | |
| HNRNPA2B1 | hnRNPA2B1 | Heterogeneous Nuclear Ribonucleoprotein A2/B1 | ENSG00000122566 |
| HNRNPC | Heterogeneous Nuclear Ribonucleoprotein C | ENSG00000092199 | |
| HNRNPL | Heterogeneous Nuclear Ribonucleoprotein L | ENSG00000104824 | |
| IGF1R | IGF-IR | Insulin Like Growth Factor 1 Receptor | ENSG00000140443 |
| IGF2 | Insulin Like Growth Factor 2 | ENSG00000167244 | |
| IGF2BP1 | IGF2 MRNA Binding Protein 1 | ENSG00000159217 | |
| IGF2BP2 | IGF2 MRNA Binding Protein 2 | ENSG00000073792 | |
| IGF2BP3 | IGF2 MRNA Binding Protein 3 | ENSG00000136231 | |
| IL1A | IL1 | Interleukin 1 | ENSG00000115008 |
| IL1B | IL1 | inflammatory cytokines Interleukin 1 | ENSG00000125538 |
| IL6 | Interleukin 6 | ENSG00000136244 | |
| IL17A | IL17 | Interleukin 17A | ENSG00000112115 |
| IL22 | Interleukin 22 | ENSG00000127318 | |
| JARID2 | Jumonji And AT-Rich Interaction Domain Containing 2 | ENSG00000008083 | |
| JUN | c-Jun | Jun Proto-Oncogene, AP-1 Transcription Factor Subunit | ENSG00000177606 |
| KDM1A | LSD1 | Lysine Demethylase 1A | ENSG00000004487 |
| KLF2 | Kruppel Like Factor 2 | ENSG00000127528 | |
| KLF4 | Kruppel Like Factor 4 | ENSG00000136826 | |
| KRT14 | Keratin 14 | ENSG00000186847 | |
| KRT19 | Cyfra21-1 | Keratin 19 | ENSG00000171345 |
| LIMK2 | LIMK2b | LIM Domain Kinase 2 | ENSG00000182541 |
| LIN28B | Lin-28 Homolog B | ENSG00000187772 | |
| LRP1 | LDL receptor-related protein-1 | ENSG00000123384 | |
| MACC1 | MET Transcriptional Regulator MACC1 | ENSG00000183742 | |
| MAP1LC3A | LC3I | Microtubule Associated Protein 1 Light Chain 3 | ENSG00000101460 |
| MAP1LC3B | LC3II | Microtubule Associated Protein 1 Light Chain 3 | ENSG00000140941 |
| MAPK1 | ERK, ERK2, MAPK | Mitogen-Activated Protein Kinase 1 | ENSG00000100030 |
| MAPK3 | ERK1 | Mitogen-Activated Protein Kinase 3 | ENSG00000102882 |
| MAPK8 | JNK | Mitogen-Activated Protein Kinase 8 | ENSG00000107643 |
| MAPK9 | Mitogen-Activated Protein Kinase 9 | ENSG00000050748 | |
| MAP2K1 | MEK1 | Mitogen-Activated Protein Kinase Kinase 1 | ENSG00000169032 |
| MAP2K2 | MEK2 | Mitogen-Activated Protein Kinase Kinase 2 | ENSG00000126934 |
| MCL1 | MCL1 Apoptosis Regulator, BCL2 Family Member | ENSG00000143384 | |
| MDM2 | MDM2 Proto-Oncogene | ENSG00000135679 | |
| MDK | MK | Midkine | ENSG00000110492 |
| MIAT | Myocardial Infarction Associated Transcript | ENSG00000225783 | |
| MGMT | O-6-Methylguanine-DNA Methyltransferase | MGMT | |
| MMP2 | MMP-2 | Matrix Metallopeptidase 2 | ENSG00000087245 |
| MMP9 | MMP-9 | Matrix Metallopeptidase 9 | ENSG00000100985 |
| MOV10 | Mov10 RISC Complex RNA Helicase | ENSG00000155363 | |
| MTOR | mTOR | Mechanistic Target Of Rapamycin Kinase | ENSG00000198793 |
| MYC | c-myc | MYC Proto-Oncogene, BHLH Transcription Factor | ENSG00000136997 |
| MYOD1 | MYOD | Myogenic differentiation 1 | ENSG00000129152 |
| NANOG | Nanog | Nanog Homeobox | ENSG00000111704 |
| NEDD9 | Neural Precursor Cell Expressed, | ||
| Developmentally Down-Regulated 9 | ENSG00000111859 | ||
| NF1 | Neurofibromin 1 | ENSG00000196712 | |
| NFKB1 | NF-B | Nuclear Factor Kappa B Subunit 1 | ENSG00000109320 |
| PARP1 | PARP | Poly(ADP-Ribose) Polymerase 1 | ENSG00000143799 |
| PCNA | Proliferating Cell Nuclear Antigen | ENSG00000132646 | |
| PDK1 | Pyruvate Dehydrogenase Kinase 1 | ENSG00000152256 | |
| PDPK1 | 3-Phosphoinositide Dependent Protein Kinase 1 | ENSG00000140992 | |
| PHOX2A | Phox2a | Paired Like Homeobox 2A | ENSG00000165462 |
| PIK3CA | PI3K | Phosphatidylinositol-4,5-Bisphosphate | |
| 3-Kinase Catalytic Subunit | ENSG00000121879 | ||
| POU5F1 | Oct3, Oct4 | POU Class 5 Homeobox 1 | ENSG00000204531 |
| PRC1 | Polycomb Repressive Complex 1 | ||
| PRC2 | Polycomb Repressive Complex 2 | ||
| PTBP1 | PTB | Polypyrimidine Tract Binding Protein 1 | ENSG00000011304 |
| PTEN | Phosphatase And Tensin Homolog | ENSG00000171862 | |
| PUM2 | Pumilio RNA Binding Family Member 2 | ENSG00000055917 | |
| RB1 | Rb | RB Transcriptional Corepressor 1 | ENSG00000139687 |
| RELA | p65 | RELA Proto-Oncogene, NF-KB Subunit | ENSG00000173039 |
| REST | RE1 Silencing Transcription Factor | ENSG00000084093 | |
| RCOR1 | CoREST | REST Corepressor 1 | ENSG00000089902 |
| RIOX2 | mdig | Ribosomal Oxygenase 2 | ENSG00000170854 |
| RORA | RAR Related Orphan Receptor A | ENSG00000069667 | |
| ROCK2 | Rho Associated Coiled-Coil | ||
| Containing Protein Kinase 2 | ENSG00000134318 | ||
| RORC | RORt | RAR related orphan receptor C | ENSG00000143365 |
| RPS6KB1 | S6K1 | Ribosomal Protein S6 Kinase B1 | ENSG00000108443 |
| RTL1 | Retrotransposon Gag Like 1 | ENSG00000254656 | |
| SFN | 14-3-3 | Stratifin | ENSG00000175793 |
| SKP2 | S-Phase Kinase Associated Protein 2 | ENSG00000145604 | |
| SLC34A2 | Solute Carrier Family 34 Member 2 | ENSG00000157765 | |
| SMAD3 | SMAD family member 3 | ENSG00000166949 | |
| SNAI1 | Snail Family Transcriptional Repressor 1 | ENSG00000124216 | |
| SNAI2 | SLUG | Snail Family Transcriptional Repressor 2 | ENSG00000019549 |
| SOX2 | SRY-box transcription factor 2 | ENSG00000181449 | |
| SOX7 | SRY-Box Transcription Factor 7 | ENSG00000012048 | |
| SOX9 | SRY-Box Transcription Factor 9 | ENSG00000125398 | |
| SP1 | Sp1 Transcription Factor | ENSG00000185591 | |
| SRSF1 | SFRS1 | Serine And Arginine Rich Splicing Factor 1 | ENSG00000136450 |
| STAT3 | Signal transducer and activator of transcription 3 | ENSG00000168610 | |
| SUZ12 | SUZ12 Polycomb Repressive Complex 2 Subunit | ENSG00000178691 | |
| TARDBP | TDP43 | TAR DNA Binding Protein | ENSG00000120948 |
| TBILA | TGF-Beta Induced LncRNA | ENSG00000261488 | |
| TCF7 | Transcription Factor 7 | ENSG00000081059 | |
| TFAP2C | Transcription Factor AP-2 | ENSG00000087510 | |
| TGFB1 | TGF1 | transforming growth factor 1 | ENSG00000105329 |
| TIA1 | TIA1 Cytotoxic Granule Associated RNA Binding Protein | ENSG00000116001 | |
| TIAL1 | TIA1 Cytotoxic Granule Associated RNA Binding Protein Like 1 | ENSG00000151923 | |
| TIMP2 | TIMP Metallopeptidase Inhibitor 2 | ENSG00000035862 | |
| TNF | TNF | Tumor Necrosis Factor | ENSG00000232810 |
| TNRC6A | TNRC6 | Trinucleotide Repeat Containing Adaptor 6A | ENSG00000090905 |
| TP53 | tumor protein p53 | ENSG00000141510 | |
| TP63 | tumor protein p63 | ENSG00000073282 | |
| TP73 | tumor protein p73 | ENSG00000078900 | |
| U2AF2 | U2AF65 | U2 Small Nuclear RNA Auxiliary Factor 2 | ENSG00000063244 |
| ULK1 | Unc-51 Like Autophagy Activating Kinase 1 | ENSG00000177169 | |
| UPF1 | UPF1 RNA Helicase And ATPase | ENSG00000005007 | |
| VIM | vimentin | ENSG00000026025 | |
| WNT10A | Wnt 10a | Wnt Family Member 10A | ENSG00000135925 |
| WNT2 | Wnt 2 | Wnt Family Member 2 | ENSG00000105989 |
| WNT3A | Wnt Family Member 3A | ENSG00000154342 | |
| WNT5A | Wnt 5a | Wnt Family Member 5A | ENSG00000114251 |
| WNT6 | Wnt 6 | Wnt Family Member 6 | ENSG00000115596 |
| YAP1 | Yes associated protein 1 | ENSG00000137693 | |
| ZC3H7B | Zinc Finger CCCH-Type Containing 7B | ENSG00000100403 | |
| ZEB1 | Zinc Finger E-Box Binding Homeobox 1 | ENSG00000148516 | |
| ZEB2 | Zinc Finger E-Box Binding Homeobox 2 | ENSG00000169554 |
References
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.F.; Flaherty, K.R.; Lasky, J.A.; et al. An Official ATS/ERS/JRS/ALAT Statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef] [PubMed]
- Guiot, J.; Corhay, J.; Louis, R. Idiopathic pulmonary fibrosis. Rev. Med. Liege 2014, 69, 605–610. [Google Scholar]
- Guiot, J.; Duysinx, B.; Seidel, L.; Henket, M.; Gester, F.; Bonhomme, O.; Corhay, J.L.; Louis, R. Clinical experience in idiopathic pulmonary fibrosis: A retrospective study. Acta Clin. Belg. Int. J. Clin. Lab. Med. 2018, 73, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Guiot, J.; Henket, M.; Corhay, J.L.; Moermans, C.; Louis, R. Sputum biomarkers in IPF: Evidence for raised gene expression and protein level of IGFBP-2, IL-8 and MMP-7. PLoS ONE 2017, 12, e0171344. [Google Scholar] [CrossRef] [PubMed]
- Nalysnyk, L.; Cid-Ruzafa, J.; Rotella, P.; Esser, D. Incidence and prevalence of idiopathic pulmonary fibrosis: Review of the literature. Eur. Respir. Rev. 2012, 21, 355–361. [Google Scholar] [CrossRef]
- Guiot, J.; Moermans, C.; Henket, M.; Corhay, J.L.; Louis, R. Blood Biomarkers in Idiopathic Pulmonary Fibrosis. Lung 2017, 195, 273–280. [Google Scholar] [CrossRef]
- Bonhomme, O.; André, B.; Gester, F.; De Seny, D.; Moermans, C.; Struman, I.; Louis, R.; Malaise, M.; Guiot, J. Biomarkers in systemic sclerosis-associated interstitial lung disease: Review of the literature. Rheumatology 2019, 58, 1534–1546. [Google Scholar] [CrossRef]
- Adeloye, D.; Chua, S.; Lee, C.; Basquill, C.; Papana, A.; Theodoratou, E.; Nair, H.; Gasevic, D.; Sridhar, D.; Campbell, H.; et al. Global and regional estimates of COPD prevalence: Systematic review and meta-analysis. J. Glob. Health 2015, 5. [Google Scholar] [CrossRef]
- Godinas, L.; Corhay, J.L.; Henket, M.; Guiot, J.; Louis, R.; Moermans, C. Increased production of TGF-β1 from sputum cells of COPD: Relationship with airway obstruction. Cytokine 2017, 99, 1–8. [Google Scholar] [CrossRef]
- Schleich, F.N.; Zanella, D.; Stefanuto, P.H.; Bessonov, K.; Smolinska, A.; Dallinga, J.W.; Henket, M.; Paulus, V.; Guissard, F.; Graff, S.; et al. Exhaled volatile organic compounds are able to discriminate between neutrophilic and eosinophilic asthma. Am. J. Respir. Crit. Care Med. 2019, 200, 444–453. [Google Scholar] [CrossRef]
- De Groot, P.M.; Wu, C.C.; Carter, B.W.; Munden, R.F. The epidemiology of lung cancer. Transl. Lung Cancer Res. 2018, 7, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.R.; Mainz, J.; Overgaard, J. Impact of delay on diagnosis and treatment of primary lung cancer. Acta Oncol. 2002, 41, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Trajkovic, K. Ceramide triggers budding of exosome vesicles into multivesicular endosomes (Science (1244)). Science 2008, 320, 179. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef]
- Njock, M.S.; Guiot, J.; Henket, M.A.; Nivelles, O.; Thiry, M.; Dequiedt, F.; Corhay, J.L.; Louis, R.E.; Struman, I. Sputum exosomes: Promising biomarkers for idiopathic pulmonary fibrosis. Thorax 2018, 74, 309–312. [Google Scholar] [CrossRef]
- Guiot, J.; Struman, I.; Louis, E.; Louis, R.; Malaise, M.; Njock, M.S. Exosomal miRNAs in Lung Diseases: From Biologic Function to Therapeutic Targets. J. Clin. Med. 2019, 8, 1345. [Google Scholar] [CrossRef]
- Guiot, J.; Struman, I.; Chavez, V.; Henket, M.; Herzog, M.; Scoubeau, K.; Hardat, N.; Bondue, B.; Corhay, J.L.; Moermans, C.; et al. Altered epigenetic features in circulating nucleosomes in idiopathic pulmonary fibrosis. Clin. Epigenet. 2017, 9, 1–7. [Google Scholar] [CrossRef]
- Njock, M.S.; Cheng, H.S.; Dang, L.T.; Nazari-Jahantigh, M.; Lau, A.C.; Boudreau, E.; Roufaiel, M.; Cybulsky, M.I.; Schober, A.; Fish, J.E. Endothelial cells suppress monocyte activation through secretion of extracellular vesicles containing antiinflammatory microRNAs. Blood 2015, 125, 3202–3212. [Google Scholar] [CrossRef]
- Maes, T.; Cobos, F.A.; Schleich, F.; Sorbello, V.; Henket, M.; De Preter, K.; Bracke, K.R.; Conickx, G.; Mesnil, C.; Vandesompele, J.; et al. Asthma inflammatory phenotypes show differential microRNA expression in sputum. J. Allergy Clin. Immunol. 2016, 137, 1433–1446. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ling, M.; Xue, J.; Dai, X.; Sun, Q.; Chen, C.; Liu, Y.; Zhou, L.; Liu, J.; Luo, F.; et al. Exosomal microRNA-21 derived from bronchial epithelial cells is involved in aberrant epithelium-fibroblast cross-talk in COPD induced by cigarette smoking. Theranostics 2018, 8, 5419–5433. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Araya, J.; Ito, S.; Kobayashi, K.; Kosaka, N.; Yoshioka, Y.; Kadota, T.; Hara, H.; Kuwano, K.; Ochiya, T. Suppression of autophagy by extracellular vesicles promotes myofibroblast differentiation in COPD pathogenesis. J. Extracell. Vesicles 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Konno, S.; Makita, H.; Shimizu, K.; Kimura, H.; Kimura, H.; Nishimura, M. Altered circulating exosomal RNA profiles detected by next-generation sequencing in patients with severe asthma. Eur. Respir. J. 2016, 48. [Google Scholar] [CrossRef]
- Zhan, C.; Zhang, Y.; Ma, J.; Wang, L.; Jiang, W.; Shi, Y.; Wang, Q. Identification of reference genes for qRT-PCR in human lung squamous-cell carcinoma by RNA-Seq. Acta Biochim. Biophys. Sin. 2014, 46, 330–337. [Google Scholar] [CrossRef]
- Pereira-Fantini, P.M.; Rajapaksa, A.E.; Oakley, R.; Tingay, D.G. Selection of reference genes for gene expression studies related to lung injury in a preterm lamb model. Sci. Rep. 2016, 6, 26476. [Google Scholar] [CrossRef]
- Moermans, C.; Deliege, E.; Pirottin, D.; Poulet, C.; Guiot, J.; Henket, M.; da Silva, J.; Louis, R. Suitable reference genes determination for real time PCR using induced sputum samples. Eur. Respir. J. 2019, 54, 1800644. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Feng, T.; Zhang, P.; Sun, Y.; Wang, Y.; Tong, J.; Dai, H.; Hua, Z. High throughput sequencing identifies breast cancer-secreted exosomal LncRNAs initiating pulmonary pre-metastatic niche formation. Gene 2019, 710, 258–264. [Google Scholar] [CrossRef]
- Segura, E.; Nicco, C.; Lombard, B.; Véron, P.; Raposo, G.; Batteux, F.; Amigorena, S.; Théry, C. ICAM-1 on exosomes from mature dendritic cells is critical for efficient naive T-cell priming. Blood 2005, 106, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Admyre, C.; Johansson, S.M.; Paulie, S.; Gabrielsson, S. Direct exosome stimulation of peripheral human T cells detected by ELISPOT. Eur. J. Immunol. 2006, 36, 1772–1781. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhu, Y.L.; Zhou, Y.Y.; Liang, G.F.; Wang, Y.Y.; Hu, F.H.; Xiao, Z.D. Exosome uptake through clathrin-mediated endocytosis and macropinocytosis and mediating miR-21 delivery. J. Biol. Chem. 2014, 289, 22258–22267. [Google Scholar] [CrossRef] [PubMed]
- Commisso, C.; Davidson, S.M.; Soydaner-Azeloglu, R.G.; Parker, S.J.; Kamphorst, J.J.; Hackett, S.; Grabocka, E.; Nofal, M.; Drebin, J.A.; Thompson, C.B.; et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 2013, 497, 633–637. [Google Scholar] [CrossRef]
- Fitzner, D.; Schnaars, M.; Van Rossum, D.; Krishnamoorthy, G.; Dibaj, P.; Bakhti, M.; Regen, T.; Hanisch, U.K.; Simons, M. Selective transfer of exosomes from oligodendrocytes to microglia by macropinocytosis. J. Cell Sci. 2011, 124, 447–458. [Google Scholar] [CrossRef]
- Parolini, I.; Federici, C.; Raggi, C.; Lugini, L.; Palleschi, S.; De Milito, A.; Coscia, C.; Iessi, E.; Logozzi, M.; Molinari, A.; et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 2009, 284, 34211–34222. [Google Scholar] [CrossRef]
- Kesimer, M.; Scull, M.; Brighton, B.; DeMaria, G.; Burns, K.; O’Neal, W.; Pickles, R.J.; Sheehan, J.K. Characterization of exosome-like vesicles released from human tracheobronchial ciliated epithelium: A possible role in innate defense. FASEB J. 2009, 23, 1858–1868. [Google Scholar] [CrossRef]
- Gupta, R.; Radicioni, G.; Abdelwahab, S.; Dang, H.; Carpenter, J.; Chua, M.; Mieczkowski, P.A.; Sheridan, J.T.; Randell, S.H.; Kesimer, M. Intercellular communication between airway epithelial cells is mediated by exosome-like vesicles. Am. J. Respir. Cell Mol. Biol. 2019, 60, 209–220. [Google Scholar] [CrossRef]
- Bourdonnay, E.; Zasłona, Z.; Penke, L.R.K.; Speth, J.M.; Schneider, D.J.; Przybranowski, S.; Swanson, J.A.; Mancuso, P.; Freeman, C.M.; Curtis, J.L.; et al. Transcellular delivery of vesicular SOCS proteins from macrophages to epithelial cells blunts inflammatory signaling. J. Exp. Med. 2015, 212, 729–742. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef]
- Kartha, R.V.; Subramanian, S. Competing endogenous RNAs (ceRNAs): New entrants to the intricacies of gene regulation. Front. Genet. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Latgé, G.; Poulet, C.; Bours, V.; Josse, C.; Jerusalem, G. Natural antisense transcripts: Molecular mechanisms and implications in breast cancers. Int. J. Mol. Sci. 2018, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.; Yu, K.; Sun, S.; Xie, C.; Skogerbø, G.; Miao, R.; Xiao, H.; Liao, Q.; Luo, H.; Zhao, G.; et al. NONCODE v3.0: Integrative annotation of long noncoding RNAs. Nucleic Acids Res. 2012, 40, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, H.; Maheronnaghsh, M.; Molaei, F.; Mashouri, L.; Reza Aref, A.; Momeny, M.; Alahari, S.K. Long noncoding RNAs and exosomal lncRNAs: Classification, and mechanisms in breast cancer metastasis and drug resistance. Oncogene 2019, 953–974. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Banks, I.R.; Zhang, Y.; Wiggins, B.E.; Heck, G.R.; Ivashuta, S. RNA decoys. Plant Signal. Behav. 2012, 7, 1188–1193. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Qu, R.; Chen, X.; Zhang, C. LncRNA ZEB1-AS1/miR-409–3p/ZEB1 feedback loop is involved in the progression of non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2018, 507, 450–456. [Google Scholar] [CrossRef]
- Balbin, O.A.; Malik, R.; Dhanasekaran, S.M.; Prensner, J.R.; Cao, X.; Wu, Y.M.; Robinson, D.; Wang, R.; Chen, G.; Beer, D.G.; et al. The landscape of antisense gene expression in human cancers. Genome Res. 2015, 25, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.; Fiscon, G.; Conte, F.; Rizzo, M.; Paci, P.; Pellegrini, M. Interplay between long noncoding RNAs and microRNAs in cancer. Methods Mol. Biol. 2018, 1819, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Veneti, Z.; Gkouskou, K.K.; Eliopoulos, A.G. Polycomb repressor complex 2 in genomic instability and cancer. Int. J. Mol. Sci. 2017, 18, 1657. [Google Scholar] [CrossRef]
- Davidovich, C.; Cech, T.R. The recruitment of chromatin modifiers by long noncoding RNAs: Lessons from PRC2. Rna 2015, 21, 2007–2022. [Google Scholar] [CrossRef] [PubMed]
- Portoso, M.; Ragazzini, R.; Brenčič, Ž.; Moiani, A.; Michaud, A.; Vassilev, I.; Wassef, M.; Servant, N.; Sargueil, B.; Margueron, R. PRC 2 is dispensable for HOTAIR -mediated transcriptional repression. EMBO J. 2017, 36, 981–994. [Google Scholar] [CrossRef]
- Kornienko, A.E.; Dotter, C.P.; Guenzl, P.M.; Gisslinger, H.; Gisslinger, B.; Cleary, C.; Kralovics, R.; Pauler, F.M.; Barlow, D.P. Long non-coding RNAs display higher natural expression variation than protein-coding genes in healthy humans. Genome Biol. 2016, 17, 1–23. [Google Scholar] [CrossRef]
- Darbellay, F.; Necsulea, A. Comparative Transcriptomics Analyses across Species, Organs, and Developmental Stages Reveal Functionally Constrained lncRNAs. Mol. Biol. Evol. 2020, 37, 240–259. [Google Scholar] [CrossRef]
- Ye, S.; Zhu, S.; Feng, L. LncRNA ANRIL/miR-125a axis exhibits potential as a biomarker for disease exacerbation, severity, and inflammation in bronchial asthma. J. Clin. Lab. Anal. 2020, 34, e23092. [Google Scholar] [CrossRef]
- Qiu, Y.Y.; Wu, Y.; Lin, M.J.; Bian, T.; Xiao, Y.L.; Qin, C. LncRNA-MEG3 functions as a competing endogenous RNA to regulate Treg/Th17 balance in patients with asthma by targeting microRNA-17/RORγt. Biomed. Pharmacother. 2019, 111, 386–394. [Google Scholar] [CrossRef]
- Keenan, C.R.; Schuliga, M.J.; Stewart, A.G. Pro-inflammatory mediators increase levels of the noncoding RNA GAS5 in airway smooth muscle and epithelial cells. Can. J. Physiol. Pharmacol. 2015, 93, 203–206. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Mao, D.; Gao, W.; Hu, H. Peripheral whole blood lncRNA expression analysis in patients with eosinophilic asthma. Medicine 2018, 97, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Feng, X.; Zhang, J.; Tong, Z. Long noncoding RNA TUG1 promotes airway smooth muscle cells proliferation and migration via sponging miR-590-5p/FGF1 in asthma. Am. J. Transl. Res. 2019, 11, 3159–3166. [Google Scholar] [PubMed]
- Du, Y.; Hao, X.; Liu, X. Low expression of long noncoding RNA CDKN2B-AS1 in patients with idiopathic pulmonary fibrosis predicts lung cancer by regulating the p53-signaling pathway. Oncol. Lett. 2018, 15, 4912–4918. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liang, Y.; Zeng, X.; Yang, X.; Xu, D. lncRNA FENDRR Exhibits Anti-Fibrotic Activity in Pulmonary Fibrosis. Am. J. Respir. Cell Mol. Biol. 2019, 1–69. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, Z.; Dai, L.; Jiang, T.; Jia, L.; Jing, X.; An, L.; Wang, H.; Liu, M. Knockdown of Long Noncoding RNA H19 Represses the Progress of Pulmonary Fibrosis through the Transforming Growth Factor β/Smad3 Pathway by Regulating MicroRNA 140. Mol. Cell. Biol. 2019, 39, 1–12. [Google Scholar] [CrossRef]
- Gokey, J.J.; Snowball, J.; Sridharan, A.; Speth, J.P.; Black, K.E.; Hariri, L.P.; Perl, A.K.T.; Xu, Y.; Whitsett, J.A. MEG3 is increased in idiopathic pulmonary fibrosis and regulates epithelial cell differentiation. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Ge, J.; Geng, S.; Jiang, H. Long noncoding RNAs antisense noncoding RNA in the INK4 locus (ANRIL) correlates with lower acute exacerbation risk, decreased inflammatory cytokines, and mild GOLD stage in patients with chronic obstructive pulmonary disease. J. Clin. Lab. Anal. 2019, 33, 1–8. [Google Scholar] [CrossRef]
- Lewis, A.; Lee, J.Y.; Donaldson, A.V.; Natanek, S.A.; Vaidyanathan, S.; Man, W.D.; Hopkinson, N.S.; Sayer, A.A.; Patel, H.P.; Cooper, C.; et al. Increased expression of H19/miR-675 is associated with a low fat-free mass index in patients with COPD. J. Cachexia Sarcopenia Muscle 2016, 7, 330–344. [Google Scholar] [CrossRef]
- Xia, H.; Xue, J.; Xu, H.; Lin, M.; Shi, M.; Sun, Q.; Xiao, T.; Dai, X.; Wu, L.; Li, J.; et al. Andrographolide antagonizes the cigarette smoke-induced epithelial-mesenchymal transition and pulmonary dysfunction through anti-inflammatory inhibiting HOTAIR. Toxicology 2019, 422, 84–94. [Google Scholar] [CrossRef]
- Song, B.; Ye, L.; Wu, S.; Jing, Z. Long non-coding RNA MEG3 regulates CSE-induced apoptosis and inflammation via regulating miR-218 in 16HBE cells. Biochem. Biophys. Res. Commun. 2019, 521, 368–374. [Google Scholar] [CrossRef]
- Gu, W.; Yuan, Y.; Wang, L.; Yang, H.; Li, S.; Tang, Z.; Li, Q. Long non-coding RNA TUG1 promotes airway remodelling by suppressing the miR-145-5p/DUSP6 axis in cigarette smoke-induced COPD. J. Cell. Mol. Med. 2019, 23, 7200–7209. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Shen, Z.; Guo, J.; Sun, S. Screening of long non-coding RNA and TUG1 inhibits proliferation with TGF-β induction in patients with COPD. Int. J. COPD 2016, 11, 2951–2964. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Chen, B.; Li, D.; Wang, Q.; Zhu, Y.; Li, M.; Wang, Y.; Fang, S.; Guo, L. HOTAIR contributes to chemoresistance by activating NF-κB signaling in small-cell lung cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 2997–3004. [Google Scholar] [PubMed]
- Liu, F.; Killian, J.K.; Yang, M.; Walker, R.L.; Hong, J.A.; Zhang, M.; Davis, S.; Zhang, Y.; Hussain, M.; Xi, S.; et al. Epigenomic alterations and gene expression profiles in respiratory epithelia exposed to cigarette smoke condensate. Oncogene 2010, 29, 3650–3664. [Google Scholar] [CrossRef]
- Liu, J.; Tian, W.; Zhang, W.; Jia, Y.; Yang, X.; Wang, Y.; Zhang, J. MicroRNA-142-3p/MALAT1 inhibits lung cancer progression through repressing β-catenin expression. Biomed. Pharmacother. 2019, 114, 108847. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, W.; Zhang, L.; Wang, L.; Wang, J.; Wan, Z.; Hong, Y.; Yu, L. MALAT1-miR-101-SOX9 feedback loop modulates the chemoresistance of lung cancer cell to DDP via Wnt signaling pathway. Oncotarget 2017, 8, 94317–94329. [Google Scholar] [CrossRef]
- Zhang, X.; He, X.; Liu, Y.; Zhang, H.; Chen, H.; Guo, S.; Liang, Y. MiR-101-3p inhibits the growth and metastasis of non-small cell lung cancer through blocking PI3K/AKT signal pathway by targeting MALAT-1. Biomed. Pharmacother. 2017, 93, 1065–1073. [Google Scholar] [CrossRef]
- Li, S.; Mei, Z.; Hu, H.B.; Zhang, X. The lncRNA MALAT1 contributes to non-small cell lung cancer development via modulating miR-124/STAT3 axis. J. Cell. Physiol. 2018, 233, 6679–6688. [Google Scholar] [CrossRef]
- Tang, Y.; Xiao, G.; Chen, Y.; Deng, Y. LncRNA MALAT1 promotes migration and invasion of non-small-cell lung cancer by targeting miR-206 and activating Akt/mTOR signaling. Anti Cancer Drugs 2018, 29, 725–735. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, H.; Li, Y.; Wang, R.; Li, Y.; Zhang, H.; Ren, D.; Liu, H.; Kang, C.; Chen, J. HOTAIR, a long noncoding RNA, is a marker of abnormal cell cycle regulation in lung cancer. Cancer Sci. 2018, 109, 2717–2733. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, C.; Yang, Y.; Guo, L.; Huang, J.; Liu, X.; Wu, C.; Zou, J. Silencing of LncRNA-HOTAIR decreases drug resistance of Non-Small Cell Lung Cancer cells by inactivating autophagy via suppressing the phosphorylation of ULK1. Biochem. Biophys. Res. Commun. 2018, 497, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Chen, J.; Ou, B.; Liu, C.; Zou, Y.; Chen, Q. GAS5 knockdown reduces the chemo-sensitivity of non-small cell lung cancer (NSCLC) cell to cisplatin (DDP) through regulating miR-21/PTEN axis. Biomed. Pharmacother. 2017, 93, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, L.; Zhang, Y.; Wu, Z.; He, D.; Li, X.; Wang, Z. Long non-coding RNA TUG1 enhances chemosensitivity in non-small cell lung cancer by impairing microRNA-221-dependent PTEN inhibition. Aging 2019, 11, 7553–7569. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Moazeni-Roodi, A.; Sarabandi, S.; Karami, S.; Ghavami, S. Association between genetic polymorphisms of long noncoding RNA H19 and cancer risk: A meta-analysis. J. Genet. 2019, 98. [Google Scholar] [CrossRef]
- Yang, Z.; Li, H.; Li, J.; Lv, X.; Gao, M.; Bi, Y.; Zhang, Z.; Wang, S.; Li, S.; Li, N.; et al. Association Between Long Noncoding RNA MEG3 Polymorphisms and Lung Cancer Susceptibility in Chinese Northeast Population. DNA Cell Biol. 2018, 37, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, G.; Fu, X.; Cui, H.; Pu, G.; Xiao, Y.; Sun, W.; Dong, X.; Zhang, L.; Cao, S.; et al. Long noncoding RNA TUG1 is a diagnostic factor in lung adenocarcinoma and suppresses apoptosis via epigenetic silencing of BAX. Oncotarget 2017, 8, 101899–101910. [Google Scholar] [CrossRef]
- Xu, R.; Mao, Y.; Chen, K.; He, W.; Shi, W.; Han, Y. The long noncoding RNA ANRIL acts as an oncogene and contributes to paclitaxel resistance of lung adenocarcinoma A549 cells. Oncotarget 2017, 8, 39177–39184. [Google Scholar] [CrossRef]
- Tian, Z.; Wen, S.; Zhang, Y.; Shi, X.; Zhu, Y.; Xu, Y.; Lv, H.; Wang, G. Identification of dysregulated long non-coding RNAs/microRNAs/mRNAs in TNM I stage lung adenocarcinoma. Oncotarget 2017, 8, 51703–51718. [Google Scholar] [CrossRef]
- Luo, J.; Li, Q.; Pan, J.; Li, L.; Fang, L.; Zhang, Y. Expression level of long noncoding RNA H19 in plasma of patients with nonsmall cell lung cancer and its clinical significance. J. Cancer Res. Ther. 2018, 14, 860. [Google Scholar] [CrossRef]
- Pan, J.; Bian, Y.; Cao, Z.; Lei, L.; Pan, J.; Huang, J.; Cai, X.; Lan, X.; Zheng, H. Long noncoding RNA MALAT1 as a candidate serological biomarker for the diagnosis of non-small cell lung cancer: A meta-analysis. Thorac. Cancer 2020, 11, 329–335. [Google Scholar] [CrossRef]
- Peng, H.; Wang, J.; Li, J.; Zhao, M.; Huang, S.K.; Gu, Y.Y.; Li, Y.; Sun, X.J.; Yang, L.; Luo, Q.; et al. A circulating non-coding RNA panel as an early detection predictor of non-small cell lung cancer. Life Sci. 2016, 151, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Yu, F.; Wang, J.; Li, Y.; Li, Y.; Li, Z.; Zhou, Q. Expression of MALAT1 in the peripheral whole blood of patients with lung cancer. Biomed. Rep. 2015, 3, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.G.; Johnen, G.; Casjens, S.; Bryk, O.; Pesch, B.; Jöckel, K.H.; Kollmeier, J.; Brüning, T. Evaluation of long noncoding RNA MALAT1 as a candidate blood-based biomarker for the diagnosis of non-small cell lung cancer. BMC Res. Notes 2013, 6, 518. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Gu, Z.T.; Chen, W.H.; Cao, K.J. Increased expression of the long non-coding RNA ANRIL promotes lung cancer cell metastasis and correlates with poor prognosis. Diagn. Pathol. 2015, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Esfandi, F.; Taheri, M.; Omrani, M.D.; Shadmehr, M.B.; Arsang-Jang, S.; Shams, R.; Ghafouri-Fard, S. Expression of long non-coding RNAs (lncRNAs) has been dysregulated in non-small cell lung cancer tissues. BMC Cancer 2019, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cheng, N.; Li, X.; Pan, H.; Li, C.; Ren, S.; Su, C.; Cai, W.; Zhao, C.; Zhang, L.; et al. Correlation of long non-coding RNA H19 expression with cisplatin-resistance and clinical outcome in lung adenocarcinoma. Oncotarget 2017, 8, 2558–2567. [Google Scholar] [CrossRef]
- Zhou, Y.; Sheng, B.; Xia, Q.; Guan, X.; Zhang, Y. Association of long non-coding RNA H19 and microRNA-21 expression with the biological features and prognosis of non-small cell lung cancer. Cancer Gene Ther. 2017, 24, 317–324. [Google Scholar] [CrossRef]
- Nakagawa, T.; Endo, H.; Yokoyama, M.; Abe, J.; Tamai, K.; Tanaka, N.; Sato, I.; Takahashi, S.; Kondo, T.; Satoh, K. Large noncoding RNA HOTAIR enhances aggressive biological behavior and is associated with short disease-free survival in human non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2013, 436, 319–324. [Google Scholar] [CrossRef]
- Wang, X.; Yin, H.; Zhang, L.; Zheng, D.; Yang, Y.; Zhang, J.; Jiang, H.; Ling, X.; Xin, Y.; Liang, H.; et al. The construction and analysis of the aberrant lncRNA-miRNA-mRNA network in non-small cell lung cancer. J. Thorac. Dis. 2019, 11, 1772–1778. [Google Scholar] [CrossRef]
- Sun, Y.J.; Li, J.; Chen, C.H. Effects of miR-221 on the apoptosis of non-small cell lung cancer cells by lncRNA HOTAIR. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 4226–4233. [Google Scholar] [CrossRef]
- Schmidt, L.H.; Spieker, T.; Koschmieder, S.; Humberg, J.; Jungen, D.; Bulk, E.; Hascher, A.; Wittmer, D.; Marra, A.; Hillejan, L.; et al. The long noncoding MALAT-1 RNA indicates a poor prognosis in non-small cell lung cancer and induces migration and tumor growth. J. Thorac. Oncol. 2011, 6, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Chen, L.; Wang, Y.; Jiang, X.; Xia, H.; Zhuang, Z. Long noncoding RNA MALAT1 promotes brain metastasis by inducing epithelial-mesenchymal transition in lung cancer. J. Neuro Oncol. 2015, 121, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, L.; Zhang, G.; Lu, C.; Chu, H.; Yang, R.; Zhao, G. MALAT1/miR-101-3p/MCL1 axis mediates cisplatin resistance in lung cancer. Oncotarget 2018, 9, 7501–7512. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhao, W.; Chen, S.; Zhang, L.; Guo, Z.; Wang, L.; Wang, J.; Wan, Z.; Hong, Y.; Yu, L. Expression and correlation of MALAT1 and SOX9 in non-small cell lung cancer. Clin. Respir. J. 2018, 12, 2284–2291. [Google Scholar] [CrossRef]
- Zhang, C.G.; Yin, D.D.; Sun, S.Y.; Han, L. The use of lncRNA analysis for stratification management of prognostic risk in patients with NSCLC. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 115–119. [Google Scholar]
- Zhang, Z.; Liu, T.; Wang, K.; Qu, X.; Pang, Z.; Liu, S.; Liu, Q.; Du, J. Down-regulation of long non-coding RNA MEG3 indicates an unfavorable prognosis in non-small cell lung cancer: Evidence from the GEO database. Gene 2017, 630, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.B.; Yin, D.D.; Sun, M.; Kong, R.; Liu, X.H.; You, L.H.; Han, L.; Xia, R.; Wang, K.M.; Yang, J.S.; et al. P53-regulated long non-coding RNA TUG1 affects cell proliferation in human non-small cell lung cancer, partly through epigenetically regulating HOXB7 expression. Cell Death Dis. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Lin, P.C.; Huang, H.D.; Chang, C.C.; Chang, Y.S.; Yen, J.C.; Lee, C.C.; Chang, W.H.; Liu, T.C.; Chang, J.G. Long noncoding RNA TUG1 is downregulated in non-small cell lung cancer and can regulate CELF1 on binding to PRC2. BMC Cancer 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Gupta, C.; Su, J.; Zhan, M.; Stass, S.A.; Jiang, F. Sputum long non-coding RNA biomarkers for diagnosis of lung cancer. Cancer Biomark. 2019, 26, 219–227. [Google Scholar] [CrossRef]
- Dong, S.; Qu, X.; Li, W.; Zhong, X.; Li, P.; Yang, S.; Chen, X.; Shao, M.; Zhang, L. The long non-coding RNA, GAS5, enhances gefitinib-induced cell death in innate EGFR tyrosine kinase inhibitor-resistant lung adenocarcinoma cells with wide-type EGFR via downregulation of the IGF-1R expression. J. Hematol. Oncol. 2015, 8, 1–13. [Google Scholar] [CrossRef]
- Chen, L.; Ren, P.; Zhang, Y.; Gong, B.; Yu, D.; Sun, X. Long non-coding RNA GAS5 increases the radiosensitivity of A549 cells through interaction with the miR-21/PTEN/Akt axis. Oncol. Rep. 2020, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, M.; Lu, K.; Liu, J.; Zhang, M.; Wu, W.; De, W.; Wang, Z.; Wang, R. The Long Noncoding RNA HOTAIR Contributes to Cisplatin Resistance of Human Lung Adenocarcinoma Cells via downregualtion of p21WAF1/CIP1 Expression. PLoS ONE 2013, 8, e77293. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Li, X.Q.; Gao, T.H.; Cui, Y.; Ma, N.; Zhou, Y.; Zhang, G.J. Elevated HOTAIR expression associated with cisplatin resistance in non-small cell lung cancer patients. J. Thorac. Dis. 2016, 8, 3314–3322. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Zheng, F.; Tang, Q.; Wu, J.J.; Xie, J.; Huang, H.D.; Yang, X.B.; Hann, S.S. Repression of PDK1-and LncRNA HOTAIR-Mediated EZH2 Gene Expression Contributes to the Enhancement of Atractylenolide 1 and Erlotinib in the Inhibition of Human Lung Cancer Cells. Cell. Physiol. Biochem. 2018, 49, 1615–1632. [Google Scholar] [CrossRef]
- Fang, S.; Gao, H.; Tong, Y.; Yang, J.; Tang, R.; Niu, Y.; Li, M.; Guo, L. Long noncoding RNA-HOTAIR affects chemoresistance by regulating HOXA1 methylation in small cell lung cancer cells. Lab. Investig. 2016, 96, 60–68. [Google Scholar] [CrossRef]
- Xu, R.; Han, Y. Long non-coding RNA FOXF1 adjacent non-coding developmental regulatory RNA inhibits growth and chemotherapy resistance in non-small cell lung cancer. Arch. Med. Sci. 2017, 15, 1539–1546. [Google Scholar] [CrossRef]
- Wang, P.; Chen, D.; Ma, H.; Li, Y. LncRNA MEG3 enhances cisplatin sensitivity in non-small cell lung cancer by regulating miR-21-5p/SOX7 axis. OncoTargets Ther. 2017, 10, 5137–5149. [Google Scholar] [CrossRef]
- Xia, Y.; He, Z.; Liu, B.; Wang, P.; Chen, Y. Downregulation of Meg3 enhances cisplatin resistance of lung cancer cells through activation of the WNT/β-catenin signaling pathway. Mol. Med. Rep. 2015, 12, 4530–4537. [Google Scholar] [CrossRef]
- Liu, J.; Wan, L.; Lu, K.; Sun, M.; Pan, X.; Zhang, P.; Lu, B.; Liu, G.; Wang, Z. The long noncoding RNA MEG3 contributes to cisplatin resistance of human lung adenocarcinoma. PLoS ONE 2015, 10, e0114586. [Google Scholar] [CrossRef]
- Xia, H.; Qu, X.L.; Liu, L.Y.; Qian, D.H.; Jing, H.Y. LncRNA MEG3 promotes the sensitivity of vincristine by inhibiting autophagy in lung cancer chemotherapy. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1020–1027. [Google Scholar] [CrossRef]
- Zhang, D.; Ding, L.; Li, Y.; Ren, J.; Shi, G.; Wang, Y.; Zhao, S.; Ni, Y.; Hou, Y. Midkine derived from cancer-associated fibroblasts promotes cisplatin-resistance via up-regulation of the expression of lncRNA ANRIL in tumour cells. Sci. Rep. 2017, 7, 16231. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res. 2019, 47, D745–D751. [Google Scholar] [CrossRef] [PubMed]
- Yates, B.; Braschi, B.; Gray, K.A.; Seal, R.L.; Tweedie, S.; Bruford, E.A. Genenames.org: The HGNC and VGNC resources in 2017. Nucleic Acids Res. 2017, 45, D619–D625. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Iny Stein, T.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef]
- Gao, Z.H.; Suppola, S.; Liu, J.; Heikkilä, P.; Jänne, J.; Voutilainen, R. Association of H19 promoter methylation with the expression of H19 and IGF-II genes in adrenocortical tumors. J. Clin. Endocrinol. Metab. 2002, 87, 1170–1176. [Google Scholar] [CrossRef]
- Lu, Q.; Guo, Z.; Xie, W.; Jin, W.; Zhu, D.; Chen, S.; Ren, T. The lncRNA H19 Mediates Pulmonary Fibrosis by Regulating the miR-196a/COL1A1 Axis. Inflammation 2018, 41, 896–903. [Google Scholar] [CrossRef]
- Austin, P.J.; Tsitsiou, E.; Boardman, C.; Jones, S.W.; Lindsay, M.A.; Adcock, I.M.; Chung, K.F.; Perry, M.M. Transcriptional profiling identifies the long noncoding RNA plasmacytoma variant translocation ( PVT1 ) as a novel regulator of the asthmatic phenotype in human airway smooth muscle. J. Allergy Clin. Immunol. 2017, 139, 780–789. [Google Scholar] [CrossRef]
- Qian, B.; Wang, D.M.; Gu, X.S.; Zhou, K.; Wu, J.; Zhang, C.Y.; He, X.Y. LncRNA H19 serves as a ceRNA and participates in non-small cell lung cancer development by regulating microRNA-107. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5946–5953. [Google Scholar] [CrossRef]
- Huang, T.; Wen, Y.; Peng, B.; Ding, G.; Yang, L.; Wang, Z. Upregulated lncRNA H19 promotes non-small cell lung cancer cell proliferation through miR-138/PDK1 axis. Int. J. Clin. Exp. Pathol. 2017, 10, 9012–9020. [Google Scholar]
- Zhao, Y.; Feng, C.; Li, Y.; Ma, Y.; Cai, R. LncRNA H19 promotes lung cancer proliferation and metastasis by inhibiting miR-200a function. Mol. Cell. Biochem. 2019, 460, 1–8. [Google Scholar] [CrossRef]
- Liu, L.; Liu, L.; Lu, S. LncRNA H19 promotes viability and epithelial-mesenchymal transition of lung adenocarcinoma cells by targeting miR-29b-3p and modifying STAT3. Int. J. Oncol. 2019, 54, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Lei, W.; Hu, H.B.; Zhang, H.; Zhu, Y. H19 promotes non-small-cell lung cancer (NSCLC) development through STAT3 signaling via sponging miR-17. J. Cell. Physiol. 2018, 233, 6768–6776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, X.; Li, X.; Li, X.; Chen, Z. LncRNA H19 promotes epithelial-mesenchymal transition (EMT) by targeting miR-484 in human lung cancer cells. J. Cell. Biochem. 2018, 119, 4447–4457. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Yu, C.; Liu, H.; Zhang, C.; Li, Y.; Zhong, X. Long non-coding RNA H19 promotes the proliferation and invasion of lung cancer cells and regulates the expression of E-cadherin, N-cadherin, and vimentin. OncoTargets Ther. 2019, 12, 4099–4107. [Google Scholar] [CrossRef]
- Xu, J.L.; Hua, T.; Ding, J.; Fan, Y.; Liu, Z.J.; Lian, J.W. FOXF2 aggravates the progression of non-small cell lung cancer through targeting lncRNA H19 to downregulate PTEN. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10796–10802. [Google Scholar] [CrossRef]
- Zhang, E.; Li, W.; Yin, D.; De, W.; Zhu, L.; Sun, S.; Han, L. c-Myc-regulated long non-coding RNA H19 indicates a poor prognosis and affects cell proliferation in non-small-cell lung cancer. Tumor Biol. 2016, 37, 4007–4015. [Google Scholar] [CrossRef]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res. 2006, 66, 5330–5337. [Google Scholar] [CrossRef]
- Shahdoust, M.; Hajizadeh, E.; Mozdarani, H.; Chehrei, A. Finding genes discriminating smokers from non-smokers by applying a growing self-organizing clustering method to large airway epithelium cell microarray data. Asian Pac. J. Cancer Prev. 2013, 14, 111–116. [Google Scholar] [CrossRef][Green Version]
- Chen, B.; Yu, M.; Chang, Q.; Lu, Y.; Thakur, C.; Ma, D.; Yi, Z.; Chen, F. Mdig de-represses H19 large intergenic non-coding RNA (lincRNA) by down-regulating H3K9me3 and heterochromatin. Oncotarget 2013, 4, 1427–1437. [Google Scholar] [CrossRef]
- Gao, L.M.; Xu, S.F.; Zheng, Y.; Wang, P.; Zhang, L.; Shi, S.S.; Wu, T.; Li, Y.; Zhao, J.; Tian, Q.; et al. Long non-coding RNA H19 is responsible for the progression of lung adenocarcinoma by mediating methylation-dependent repression of CDH1 promoter. J. Cell. Mol. Med. 2019, 23, 6411–6428. [Google Scholar] [CrossRef]
- Wang, L.; Sun, Y.; Yi, J.; Wang, X.; Liang, J.; Pan, Z.; Li, L.; Jiang, G. Targeting H19 by lentivirus-mediated RNA interference increases A549 cell migration and invasion. Exp. Lung Res. 2016, 42, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Fu, J.; Ma, T.; Yan, B.; Gao, R.; An, Z.; Wang, D. LncRNA H19-elevated LIN28B promotes lung cancer progression through sequestering miR-196b. Cell Cycle 2018, 17, 1372–1380. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.J.; Zheng, L.M.; Feng, Z.X.; Li, M.Y.; Liu, L.; Zhao, Y.J.; Jiang, J.Y. H19 contributes to poor clinical features in NSCLC patients and leads to enhanced invasion in A549 cells through regulating miRNA-203-mediated epithelial-mesenchymal transition. Oncol. Lett. 2018, 16, 4480–4488. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.H.; Wu, D.M.; Fan, S.H.; Zhang, Z.F.; Chen, G.Q.; Lu, J. Upregulation of miR-675-5p induced by lncRNA H19 was associated with tumor progression and development by targeting tumor suppressor p53 in non–small cell lung cancer. J. Cell. Biochem. 2019, 120, 18724–18735. [Google Scholar] [CrossRef]
- Kondo, M.; Suzuki, H.; Ueda, R.; Osada, H.; Takagi, K.; Takahashi, T.; Takahashi, T. Frequent loss of imprinting of the H19 gene is often associated with its overexpression in human lung cancers. Oncogene 1995. [Google Scholar] [CrossRef]
- Booton, R.; Lindsay, M.A. Emerging role of microRNAs and long noncoding RNAs in respiratory disease. Chest 2014, 146, 193–204. [Google Scholar] [CrossRef]
- Xu, Y.Z.; Lin, J.; Jin, Y.Y.; Chen, M.; Zheng, H.H.; Feng, J.X. The miRNA hsa-miR-6515-3p potentially contributes to lncRNA H19-mediated-lung cancer metastasis. J. Cell. Biochem. 2019, 120, 17413–17421. [Google Scholar] [CrossRef]
- Lei, Y.; Guo, W.; Chen, B.; Chen, L.; Gong, J.; Li, W. Tumor-released lncRNA H19 promotes gefitinib resistance via packaging into exosomes in non-small cell lung cancer. Oncol. Rep. 2018, 40, 3438–3446. [Google Scholar] [CrossRef]
- Lu, K.-H.; Li, W.; Liu, X.H.; Sun, M.; Zhang, M.-L.; Wu, W.-Q.; Xie, W.P.; Hou, Y.Y. Long non-coding RNA MEG3 inhibits NSCLC cells proliferation and induces apoptosis by affecting p53 expression. BMC Cancer 2013, 13. [Google Scholar] [CrossRef]
- Molina-Pinelo, S.; Salinas, A.; Moreno-Mata, N.; Ferrer, I.; Suarez, R.; Andrés-León, E.; Rodríguez-Paredes, M.; Gutekunst, J.; Jantus-Lewintre, E.; Camps, C.; et al. Impact of DLK1-DIO3 imprinted cluster hypomethylation in smoker patients with lung cancer. Oncotarget 2018, 9, 4395–4410. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Zhou, Y.; Ansell, P.J.; Klibanski, A. Cyclic AMP stimulates MEG3 gene expression in cells through a cAMP-response element (CRE) in the MEG3 proximal promoter region. Int. J. Biochem. Cell Biol. 2006, 38, 1808–1820. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; O’Sullivan, M.J.; Green, A.J.; Watabe, Y.; Arisaka, O.; Masawa, N.; Matsuoka, K.; Fukami, M.; Matsubara, K.; Kato, F.; et al. The IG-DMR and the MEG3-DMR at human chromosome 14q32.2: Hierarchical interaction and distinct functional properties as imprinting control centers. PLoS Genet. 2010, 6, e1000992. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Han, D.; Wu, J.; Huo, X. Skp2 regulates non-small cell lung cancer cell growth by Meg3 and miR-3163. Tumor Biol. 2016, 37, 3925–3931. [Google Scholar] [CrossRef] [PubMed]
- Al-Rugeebah, A.; Alanazi, M.; Parine, N.R. MEG3: An Oncogenic Long Non-coding RNA in Different Cancers. Pathol. Oncol. Res. 2019, 25, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Kruer, T.L.; Dougherty, S.M.; Reynolds, L.; Long, E.; De Silva, T.; Lockwood, W.W.; Clem, B.F. Expression of the lncRNA maternally expressed gene 3 (MEG3) contributes to the control of lung cancer cell proliferation by the Rb pathway. PLoS ONE 2016, 11, e0166363. [Google Scholar] [CrossRef]
- Wu, J.L.; Meng, F.M.; Li, H.J. High expression of lncRNA MEG3 participates in non-small cell lung cancer by regulating microRNA-7-5p. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5938–5945. [Google Scholar] [CrossRef]
- Li, X.; Zheng, M.; Pu, J.; Zhou, Y.; Hong, W.; Fu, X.; Peng, Y.; Zhou, W.; Pan, H.; Li, B.; et al. Identification of abnormally expressed lncRNAs induced by PM2.5 in human bronchial epithelial cells. Biosci. Rep. 2018, 38, 1–14. [Google Scholar] [CrossRef]
- Terashima, M.; Tange, S.; Ishimura, A.; Suzuki, T. MEG3 long noncoding RNA contributes to the epigenetic regulation of epithelial-mesenchymal transition in lung cancer cell lines. J. Biol. Chem. 2017, 292, 82–99. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, Z.; Shi, S.; Wang, J.; Li, N. Long non-coding RNA MEG3 regulates migration and invasion of lung cancer stem cells via miR-650/SLC34A2 axis. Biomed. Pharmacother. 2019, 120, 109457. [Google Scholar] [CrossRef]
- Wang, P.; Chen, D.; Ma, H.; Li, Y. Long non-coding RNA MEG3 regulates proliferation and apoptosis in non-small cell lung cancer: Via the miR-205-5p/LRP1 pathway. RSC Adv. 2017, 7, 49710–49719. [Google Scholar] [CrossRef]
- Li, D.S.; Ainiwaer, J.L.; Sheyhiding, I.; Zhang, Z.; Zhang, L.W. Identification of key long non-coding RNAs as competing endogenous RNAs for miRNA-mRNA in lung adenocarcinoma. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2285–2295. [Google Scholar] [PubMed]
- Xu, J.; Su, C.; Zhao, F.; Tao, J.; Hu, D.; Shi, A.; Pan, J.; Zhang, Y. Paclitaxel promotes lung cancer cell apoptosis via MEG3-P53 pathway activation. Biochem. Biophys. Res. Commun. 2018, 504, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Filippov-Levy, N.; Cohen-Schussheim, H.; Tropé, C.G.; Hetland Falkenthal, T.E.; Smith, Y.; Davidson, B.; Reich, R. Expression and clinical role of long non-coding RNA in high-grade serous carcinoma. Gynecol. Oncol. 2018, 148, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.C.; Luo, X.H.; Tao, G.X.; Guan, M.; Yuan, H.; Hu, D.K. Exosomal Long Noncoding RNAs are Differentially Expressed in the Cervicovaginal Lavage Samples of Cervical Cancer Patients. J. Clin. Lab. Anal. 2016, 30, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, Y.; Liu, R.; Chen, B.; Liu, C.; Xing, J. Long noncoding RNA (MEG3) in urinal exosomes functions as a biomarker for the diagnosis of Hunner-type interstitial cystitis (HIC). J. Cell. Biochem. 2020, 121, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- He, R.Z.; Luo, D.X.; Mo, Y.Y. Emerging roles of lncRNAs in the post-transcriptional regulation in cancer. Genes Dis. 2019, 6, 6–15. [Google Scholar] [CrossRef]
- Parker, M.M.; Chase, R.P.; Lamb, A.; Reyes, A.; Saferali, A.; Yun, J.H.; Himes, B.E.; Silverman, E.K.; Hersh, C.P.; Castaldi, P.J. RNA sequencing identifies novel non-coding RNA and exon-specific effects associated with cigarette smoking. BMC Med. Genom. 2017, 10, 58. [Google Scholar] [CrossRef]
- Gutschner, T.; Hämmerle, M.; Eißmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Groß, M.; et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef]
- Guo, F.; Jiao, F.; Song, Z.; Li, S.; Liu, B.; Yang, H.; Zhou, Q.; Li, Z. Regulation of MALAT1 expression by TDP43 controls the migration and invasion of non-small cell lung cancer cells in vitro. Biochem. Biophys. Res. Commun. 2015, 465, 293–298. [Google Scholar] [CrossRef]
- Tano, K.; Onoguchi-Mizutani, R.; Yeasmin, F.; Uchiumi, F.; Suzuki, Y.; Yada, T.; Akimitsu, N. Identification of minimal p53 promoter region regulated by MALAT1 in human lung adenocarcinoma cells. Front. Genet. 2018, 9, 208. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, W.; Xu, Y.; Lv, X.; Zhang, M.; Jiang, H. Polyphyllin I modulates MALAT1/STAT3 signaling to induce apoptosis in gefitinib-resistant non-small cell lung cancer. Toxicol. Appl. Pharmacol. 2018, 356, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Ding, J.; He, M.; Chen, Y.; Wang, R.; Han, Z.; Xing, E.Z.; Zhang, C.; Yeh, S. Estrogen receptor β promotes the vasculogenic mimicry (VM) and cell invasion via altering the lncRNA-MALAT1/miR-145-5p/NEDD9 signals in lung cancer. Oncogene 2018, 38, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wu, K.; Liu, K.; Miao, R. Effects of MALAT1 on proliferation and apoptosis of human non-small cell lung cancer A549 cells in vitro and tumor xenograft growth in vivo by modulating autophagy. Cancer Biomarkers 2018, 22, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xia, Y.; Wang, Z.; Zheng, J.; Chen, Y.; Li, X.; Wang, Y.; Ming, H. Serum long non coding RNA MALAT-1 protected by exosomes is up-regulated and promotes cell proliferation and migration in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2017, 490, 406–414. [Google Scholar] [CrossRef]
- Lin, L.; Li, H.; Zhu, Y.; He, S.; Ge, H. Expression of metastasis-associated lung adenocarcinoma transcript 1 long non-coding RNA in vitro and in patients with non-small cell lung cancer. Oncol. Lett. 2018, 15, 9443–9449. [Google Scholar] [CrossRef]
- Guo, F.; Guo, L.; Li, Y.; Zhou, Q.; Li, Z. MALAT1 is an oncogenic long non-coding RNA associated with tumor invasion in non-small cell lung cancer regulated by DNA methylation. Int. J. Clin. Exp. Pathol. 2015, 8, 15903–15910. [Google Scholar]
- Liu, M.; Sun, W.; Liu, Y.; Dong, X. The role of lncRNA MALAT1 in bone metastasis in patients with non-small cell lung cancer. Oncol. Rep. 2016, 36, 1679–1685. [Google Scholar] [CrossRef]
- Jen, J.; Tang, Y.A.; Lu, Y.H.; Lin, C.C.; Lai, W.W.; Wang, Y.C. Oct4 transcriptionally regulates the expression of long non-coding RNAs NEAT1 and MALAT1 to promote lung cancer progression. Mol. Cancer 2017, 16, 1–12. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Ashrafi Hafez, A.; Taheri, M. Metastasis Associated Lung Adenocarcinoma Transcript 1: An update on expression pattern and functions in carcinogenesis. Exp. Mol. Pathol. 2020, 112, 104330. [Google Scholar] [CrossRef]
- Li, S.; Wang, Q.; Qiang, Q.; Shan, H.; Shi, M.; Chen, B.; Zhao, S.; Yuan, L. Sp1-mediated transcriptional regulation of MALAT1 plays a critical role in tumor. J. Cancer Res. Clin. Oncol. 2015, 141, 1909–1920. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Xu, Y.; Zhang, K.; Huang, J.; Pan, B.; Chen, D.; Cui, S.; Song, H.; Wang, R.; et al. TFAP2C-Activated MALAT1 Modulates the Chemoresistance of Docetaxel-Resistant Lung Adenocarcinoma Cells. Mol. Ther. Nucleic Acids 2019, 14, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Chen, W.; Yuan, Z.; Liu, X.; Jiang, H. LncRNA-MALAT1 contributes to the cisplatin-resistance of lung cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed. Pharmacother. 2018, 101, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, H.; Jia, J.; Ruan, X.; Liu, Y.; Zhang, X. High metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) expression promotes proliferation, migration, and invasion of non-small cell lung cancer via ERK/mitogen-activated protein kinase (MAPK) signaling pathway. Med. Sci. Monit. 2019, 25, 5143–5149. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Li, G.; Zhang, X.; Dai, F.; Zhang, R. Increased MALAT1 expression contributes to cisplatin resistance in non-small cell lung cancer. Oncol. Lett. 2018, 16, 4821–4828. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Chen, Y.; Li, S.; Jin, M.; Wang, H.; Chen, Z.; Yu, W. LncRNA MALAT1 exerts oncogenic functions in lung adenocarcinoma by targeting miR-204. Am. J. Cancer Res. 2016, 6, 1099–1107. [Google Scholar]
- Wei, S.; Wang, K.; Huang, X.; Zhao, Z.; Zhao, Z. LncRNA malat1 contributes to non-small cell lung cancer progression via modulating mir-200a-3p/programmed death-ligand 1 axis. Int. J. Immunopathol. Pharmacol. 2019, 33. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Zhu, Q.; Zhou, J. Long non-coding RNA MALAT1 interaction with miR-429 regulates the proliferation and EMT of lung adenocarcinoma cells through RhoA. Int. J. Clin. Exp. Pathol. 2019, 12, 419–430. [Google Scholar]
- Yang, T.; Li, H.; Chen, T.; Ren, H.; Shi, P.; Chen, M. LncRNA MALAT1 depressed chemo-sensitivity of NSCLC cells through directly functioning on miR-197-3p/p120 catenin axis. Mol. Cells 2019, 42, 270–283. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Zhang, Z.L.; Cui, H.X.; Wang, R.K.; Fu, L. Long non-coding RNA FENDRR inhibits NSCLC cell growth and aggressiveness by sponging miR-761. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8324–8332. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, S.; Li, X.; Feng, C.; Huang, Q.; Wang, S.; Wang, S.; Xia, W.; Yang, F.; Yin, R.; et al. Profiling expression of coding genes, long noncoding RNA, and circular RNA in lung adenocarcinoma by ribosomal RNA-depleted RNA sequencing. FEBS Open Bio 2018, 8, 544–555. [Google Scholar] [CrossRef]
- Chen, W.J.; Tang, R.X.; He, R.Q.; Li, D.Y.; Liang, L.; Zeng, J.H.; Hu, X.H.; Ma, J.; Li, S.K.; Chen, G. Clinical roles of the aberrantly expressed lncRNAs in lung squamous cell carcinoma: A study based on RNA sequencing and microarray data mining. Oncotarget 2017, 8, 61282–61304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, S.; Choi, Y.L.; Lee, J.; Gong, Z.; Liu, X.; Pei, Y.; Jiang, A.; Ye, M.; Mao, M.; et al. Systematic identification of cancer-related long noncoding RNAs and aberrant alternative splicing of quintuple-negative lung adenocarcinoma through RNA-Seq. Lung Cancer 2017, 109, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Acha-Sagredo, A.; Uko, B.; Pantazi, P.; Bediaga, N.; Moschandrea, C.; Rainbow, L.; Marcus, M.; Davies, M.; Field, J.; Liloglou, T. Long non-coding RNA dysregulation is a frequent event in non-small cell lung carcinoma pathogenesis. Br. J. Cancer 2020, 122, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Huang, Z.; Zengli, Z.; Li, H.; Chen, Q.; Yao, C.; Cai, H.; Xiao, Y.; Xia, H.; Wang, Y. Loss of long noncoding RNA FOXF1-AS1 regulates epithelialmesenchymal transition, stemness and metastasis of non-small cell lung cancer cells. Oncotarget 2016, 7, 68339–68349. [Google Scholar] [CrossRef]
- Gong, F.; Dong, D.; Zhang, T.; Xu, W. Long non-coding RNA FENDRR attenuates the stemness of non-small cell lung cancer cells via decreasing multidrug resistance gene 1 (MDR1) expression through competitively binding with RNA binding protein HuR. Eur. J. Pharmacol. 2019, 853, 345–352. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, Q.; Zhang, X.; Ding, Z.; Liu, R. LncRNA FENDRR suppresses the progression of NSCLC via regulating miR-761/TIMP2 axis. Biomed. Pharmacother. 2019, 118, 109309. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, D.; He, Z.; Zheng, L. Integrated analysis reveals five potential ceRNA biomarkers in human lung adenocarcinoma. PeerJ 2019, 2019. [Google Scholar] [CrossRef]
- Ghaforui-Fard, S.; Vafaee, R.; Taheri, M. Taurine-upregulated gene 1: A functional long noncoding RNA in tumorigenesis. J. Cell. Physiol. 2019, 234, 17100–17112. [Google Scholar] [CrossRef]
- Ma, P.J.; Guan, Q.K.; Meng, L.; Qin, N.; Zhao, J.; Jin, B.Z. Long non-coding RNA TUG1 as a potential prognostic biomarker in human cancers: A meta-analysis. Oncotarget 2017, 8, 62454–62462. [Google Scholar] [CrossRef]
- Niu, Y.; Ma, F.; Huang, W.; Fang, S.; Li, M.; Wei, T.; Guo, L. Long non-coding RNA TUG1 is involved in cell growth and chemoresistance of small cell lung cancer by regulating LIMK2b via EZH2. Mol. Cancer 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Scalia, M.; Magnano, A.; Caltabiano, R.; Barbagallo, D.; Biondi, A.; Cappellani, A.; et al. LncRNA UCA1, Upregulated in CRC Biopsies and Downregulated in Serum Exosomes, Controls mRNA Expression by RNA-RNA Interactions. Mol. Ther. Nucleic Acids 2018, 12, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Burd, C.E.; Jeck, W.R.; Liu, Y.; Sanoff, H.K.; Wang, Z.; Sharpless, N.E. Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk. PLoS Genet. 2010, 6, e1001165. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular Interplay of the Noncoding RNA ANRIL and Methylated Histone H3 Lysine 27 by Polycomb CBX7 in Transcriptional Silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15 INK4B tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Busch, S.E.; Moser, R.D.; Gurley, K.E.; Kelly-Spratt, K.S.; Liggitt, H.D.; Kemp, C.J. ARF inhibits the growth and malignant progression of non-small-cell lung carcinoma. Oncogene 2014, 33, 2665–2673. [Google Scholar] [CrossRef]
- Alsibai, K.D.; Vacher, S.; Meseure, D.; Nicolas, A.; Lae, M.; Schnitzler, A.; Chemlali, W.; Cros, J.; Longchampt, E.; Cacheux, W.; et al. High positive correlations between ANRIL and p16-CDKN2A/p15-CDKN2B/p14-ARF gene cluster overexpression in multi-tumor types suggest deregulated activation of an ANRIL-ARF bidirectional promoter. Non-Coding RNA 2019, 5, 44. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, X.H.; Xu, L.; Rong, C.H.; Shen, C.; Bian, W. Long noncoding RNA ANRIL could be transactivated by c-Myc and promote tumor progression of non-small-cell lung cancer. OncoTargets Ther. 2016, 9, 3077–3084. [Google Scholar] [CrossRef][Green Version]
- Nie, F.Q.; Sun, M.; Yang, J.S.; Xie, M.; Xu, T.P.; Xia, R.; Liu, Y.W.; Liu, X.H.; Zhang, E.B.; Lu, K.H.; et al. Long noncoding RNA ANRIL promotes non-small cell lung cancer cell proliferation and inhibits apoptosis by silencing KLF2 and P21 expression. Mol. Cancer Ther. 2015, 14, 268–277. [Google Scholar] [CrossRef]
- Fehringer, G.; Kraft, P.; Pharoah, P.D.; Eeles, R.A.; Chatterjee, N.; Schumacher, F.R.; Schildkraut, J.M.; Lindström, S.; Brennan, P.; Bickeböller, H.; et al. Cross-cancer genome-wide analysis of lung, ovary, breast, prostate, and colorectal cancer reveals novel pleiotropic associations. Cancer Res. 2016, 76, 5103–5114. [Google Scholar] [CrossRef]
- Timmers, P.R.; Mounier, N.; Lall, K.; Fischer, K.; Ning, Z.; Feng, X.; Bretherick, A.D.; Clark, D.W.; Agbessi, M.; Ahsan, H.; et al. Genomics of 1 million parent lifespans implicates novel pathways and common diseases and distinguishes survival chances. eLife 2019, 8, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Pfeiffer, R.M.; Hyland, P.L.; Shi, J.; Gu, F.; Wang, Z.; Bhattacharjee, S.; Luo, J.; Xiong, X.; Yeager, M.; et al. Genetic polymorphisms in the 9p21 region associated with risk of multiple cancers. Carcinogenesis 2014, 35, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, M.N.; Hung, R.J.; Rafnar, T.; Christiani, D.C.; Field, J.K.; Bickeböller, H.; Risch, A.; Mckay, J.D.; Wang, Y.; Dai, J.; et al. Influence of common genetic variation on lung cancer risk: Meta-analysis of 14 900 cases and 29 485 controls. Hum. Mol. Genet. 2012, 21, 4980–4995. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Cui, Z.; Li, H.; Li, J.; Yang, Z.; Bi, Y.; Gao, M.; Zhang, Z.; Wang, S.; Zhou, B.; et al. Association between polymorphism in CDKN2B-AS1 gene and its interaction with smoking on the risk of lung cancer in a Chinese population. Hum. Genom. 2019, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Abbastabar, M.; Sarfi, M.; Golestani, A.; Karimi, A.; Pourmand, G.; Khalili, E. Tumor-derived urinary exosomal long non-coding RNAs as diagnostic biomarkers for bladder cancer. EXCLI J. 2020, 19, 301–310. [Google Scholar] [CrossRef]
- Zhou, C.; Ye, L.; Jiang, C.; Bai, J.; Chi, Y.; Zhang, H. Long noncoding RNA HOTAIR, a hypoxia-inducible factor-1α activated driver of malignancy, enhances hypoxic cancer cell proliferation, migration, and invasion in non-small cell lung cancer. Tumor Biol. 2015, 36, 9179–9188. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, F.; Xu, Y.; Wang, B.; Zhao, Y.; Xu, W.; Shi, L.; Lu, X.; Liu, Q. Epithelial-mesenchymal transition and cancer stem cells, mediated by a long non-coding RNA, HOTAIR, are involved in cell malignant transformation induced by cigarette smoke extract. Toxicol. Appl. Pharmacol. 2015, 282, 9–19. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Song, G.; Zhang, X.; Gao, S.; Liu, H. HOX cluster-embedded antisense long non-coding RNAs in lung cancer. Cancer Lett. 2019, 450, 14–21. [Google Scholar] [CrossRef]
- Liu, W.; Yin, N.C.; Liu, H.; Nan, K.J. Cav-1 promote lung cancer cell proliferation and invasion through lncRNA HOTAIR. Gene 2018, 641, 335–340. [Google Scholar] [CrossRef]
- Zhai, N.; Xia, Y.; Yin, R.; Liu, J.; Gao, F. A negative regulation loop of long noncoding RNA HOTAIR and p53 in non-small-cell lung cancer. OncoTargets Ther. 2016, 9, 5713–5720. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, X.; Huang, Y.; Tang, Q.; Ma, C.; Zheng, F.; Wu, W.; Hann, S.S. Interaction of C-Jun and HOTAIR- increased expression of p21 converge in Polyphyllin I-inhibited growth of human lung cancer cells. OncoTargets Ther. 2019, 12, 10115–10127. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Yan, B.; Li, Z.; Jiang, Y.; Mao, C.; Wang, X.; Zhou, X. Long non-coding RNA HOX transcript antisense RNA promotes expression of 14-3-3σ in non-small cell lung cancer. Exp. Ther. Med. 2017, 14, 4503–4508. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Cao, Z.; Guo, H.; Li, S. The action mechanism of lncRNA-HOTAIR on the drug resistance of non-small cell lung cancer by regulating Wnt signaling pathway. Exp. Ther. Med. 2018, 15, 4885–4889. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Shi, Y.; Chen, L.; Jiang, Y.; Mao, C.; Yan, B.; Liu, S.; Shan, B.; Tao, Y.; Wang, X. The ratio of FoxA1 to FoxA2 in lung adenocarcinoma is regulated by LncRNA HOTAIR and chromatin remodeling factor LSH. Sci. Rep. 2015, 5, 17826. [Google Scholar] [CrossRef]
- Fang, S.; Shen, Y.; Chen, B.; Wu, Y.; Jia, L.; Li, Y.; Zhu, Y.; Yan, Y.; Li, M.; Chen, R.; et al. H3K27me3 induces multidrug resistance in small cell lung cancer by affecting HOXA1 DNA methylation via regulation of the lncRNA HOTAIR. Ann. Transl. Med. 2018, 6, 440. [Google Scholar] [CrossRef]
- Tang, Q.; Zheng, F.; Liu, Z.; Wu, J.J.; Chai, X.S.; He, C.X.; Li, L.; Hann, S.S. Novel reciprocal interaction of lncRNA HOTAIR and miR-214-3p contribute to the solamargine-inhibited PDPK1 gene expression in human lung cancer. J. Cell. Mol. Med. 2019, 23, 7749–7761. [Google Scholar] [CrossRef]
- Chen, S.S.; Peng, M.; Zhou, G.Z.; Pu, Y.C.; Yi, M.C.; Zhu, Y.; Jiang, B. Long non-coding RNA HOTAIR regulates the development of non-small cell lung cancer through miR-217/DACH1 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 670–678. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Chen, Z.; Wang, J.; Chen, Y.; Xu, Z.; Jin, M.; Yu, W. miR-326 reverses chemoresistance in human lung adenocarcinoma cells by targeting specificity protein 1. Tumor Biol. 2016, 37, 13287–13294. [Google Scholar] [CrossRef]
- Wang, R.; Chen, X.; Xu, T.; Xia, R.; Han, L.; Chen, W.; De, W.; Shu, Y. MiR-326 regulates cell proliferation and migration in lung cancer by targeting phox2a and is regulated by HOTAIR. Am. J. Cancer Res. 2016, 6, 173–186. [Google Scholar]
- Jiang, C.; Yang, Y.; Yang, Y.; Guo, L.; Huang, J.; Liu, X.; Wu, C.; Zou, J. Long noncoding RNA (lncRNA) HOTAIR affects tumorigenesis and metastasis of non-small cell lung cancer by upregulating miR-613. Oncol. Res. 2018, 26, 725–734. [Google Scholar] [CrossRef]
- Sassenberg, M.; Droop, J.; Schulz, W.A.; Dietrich, D.; Loick, S.M.; Wiek, C.; Scheckenbach, K.; Gaisa, N.T.; Hoffmann, M.J. Upregulation of the long non-coding RNA CASC9 as a biomarker for squamous cell carcinoma. BMC Cancer 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Yin, Z.; Yang, L.; Fan, J.; Xu, J.; Jin, Y.; Yu, J.; Zhang, D.; Yang, G. Smoking induced extracellular vesicles release and their distinct properties in non-small cell lung cancer. J. Cancer 2019, 10, 3435–3443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, L.; Deng, G.; Ding, Y.; Bi, K.; Jin, H.; Shu, J.; Yang, J.; Deng, H.; Wang, Z.; et al. Exosomal HOTAIR promotes proliferation, migration and invasion of lung cancer by sponging miR-203. Sci. China Life Sci. 2020, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Kulkarni, P.; Majid, S.; Shahryari, V.; Hashimoto, Y.; Bhat, N.S.; Shiina, M.; Deng, G.; Saini, S.; Laura Tabatabai, Z.; et al. MicroRNA-203 inhibits long noncoding RNA hotair and regulates tumorigenesis through epithelial-to-mesenchymal transition pathway in renal cell carcinoma. Mol. Cancer Ther. 2018, 17, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, G.; Li, Y.; Zhu, Z. Upregulation of long noncoding RNA GAS5 inhibits lung cancer cell proliferation and metastasis via miR-205/PTEN axis. Med. Sci. Monit. 2019, 25, 2311–2319. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ni, T.; Jiang, Y.; Li, Y. Long noncoding RNA GAS5 inhibits tumorigenesis and enhances radiosensitivity by suppressing miR-135b expression in non-small cell lung cancer. Oncol. Res. 2017, 25, 1305–1316. [Google Scholar] [CrossRef]
- Mei, Y.; Si, J.; Wang, Y.; Huang, Z.; Zhu, H.; Feng, S.; Wu, X.; Wu, L. Long noncoding RNA GAS5 suppresses tumorigenesis by inhibiting miR-23a expression in non-small cell lung cancer. Oncol. Res. 2017, 25, 1027–1037. [Google Scholar] [CrossRef]
- Cheng, Y.; Dai, X.; Yang, T.; Zhang, N.; Liu, Z.; Jiang, Y. Low Long Noncoding RNA Growth Arrest-Specific Transcript 5 Expression in the Exosomes of Lung Cancer Cells Promotes Tumor Angiogenesis. J. Oncol. 2019, 2019. [Google Scholar] [CrossRef]
- Li, C.; Lv, Y.; Shao, C.; Chen, C.; Zhang, T.; Wei, Y.; Fan, H.; Lv, T.; Liu, H.; Song, Y. Tumor-derived exosomal lncRNA GAS5 as a biomarker for early-stage non-small-cell lung cancer diagnosis. J. Cell. Physiol. 2019, 234, 20721–20727. [Google Scholar] [CrossRef]
- Qian, Y.; Mao, Z.D.; Shi, Y.J.; Liu, Z.G.; Cao, Q.; Zhang, Q. Comprehensive Analysis of miRNA-mRNA-lncRNA Networks in Non-Smoking and Smoking Patients with Chronic Obstructive Pulmonary Disease. Cell. Physiol. Biochem. 2018, 50, 1154–1163. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, Z.D.; Shi, Y.J.; Qian, Y.; Liu, Z.G.; Yin, X.W.; Zhang, Q. Comprehensive analysis of miRNA–mRNA–lncRNA networks in severe asthma. Epigenomics 2019, 11, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Dang, X.; Wang, W.; Li, Y.; Xu, D.; Shang, D.; Chang, Y. Long Noncoding RNAs and mRNA Regulation in Peripheral Blood Mononuclear Cells of Patients with Chronic Obstructive Pulmonary Disease. Mediat. Inflamm. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Tong, F.; Dong, X.; Wu, G.; Zhang, R. lncRNA UCA1 Promotes Gefitinib Resistance as a ceRNA to Target FOSL2 by Sponging miR-143 in Non-small Cell Lung Cancer. Mol. Ther. Nucleic Acids 2020, 19, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Kwok, H.H.; Ning, Z.; Chong, P.W.C.; Wan, T.S.K.; Ng, M.H.L.; Ho, G.Y.; Ip, M.S.M.; Lam, D.C.L. Transfer of extracellular vesicle-associated-RNAs induces drug resistance in ALK-translocated lung adenocarcinoma. Cancers 2019, 11, 104. [Google Scholar] [CrossRef]
- Wu, D.-M.; Deng, S.-H.; Liu, T.; Han, R.; Zhang, T.; Xu, Y. TGF-β-mediated exosomal lnc-MMP2-2 regulates migration and invasion of lung cancer cells to the vasculature by promoting MMP2 expression. Cancer Med. 2018, 7, 5118–5129. [Google Scholar] [CrossRef]
- Zhang, W.; Cai, X.; Yu, J.; Lu, X.; Qian, Q.; Qian, W. Exosome-mediated transfer of lncRNA RP11-838N2.4 promotes erlotinib resistance in non-small cell lung cancer. Int. J. Oncol. 2018, 53, 527–538. [Google Scholar] [CrossRef]
- Tao, Y.; Tang, Y.; Yang, Z.; Wu, F.; Wang, L.; Yang, L.; Lei, L.; Jing, Y.; Jiang, X.; Jin, H.; et al. Exploration of Serum Exosomal LncRNA TBILA and AGAP2-AS1 as Promising Biomarkers for Diagnosis of Non-Small Cell Lung Cancer. Int. J. Biol. Sci. 2020, 16, 471–482. [Google Scholar] [CrossRef]
- Teng, Y.; Kang, H.; Chu, Y. Identification of an Exosomal Long Noncoding RNA SOX2-OT in Plasma as a Promising Biomarker for Lung Squamous Cell Carcinoma. Genet. Test. Mol. Biomarkers 2019, 23, 235–240. [Google Scholar] [CrossRef]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [CrossRef]
- Yu, X.; Li, Z. Long non-coding RNA HOTAIR: A novel oncogene (Review). Mol. Med. Rep. 2015, 12, 5611–5618. [Google Scholar] [CrossRef]
- Raveh, E.; Matouk, I.J.; Gilon, M.; Hochberg, A. The H19 Long non-coding RNA in cancer initiation, progression and metastasis—A proposed unifying theory. Mol. Cancer 2015, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, X.; Klibanski, A. MEG3 noncoding RNA: A tumor suppressor. J. Mol. Endocrinol. 2012, 48, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Taheri, M. Maternally expressed gene 3 (MEG3): A tumor suppressor long non coding RNA. Biomed. Pharmacother. 2019, 118, 109129. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Shi, X.; Zhu, Q.; Li, Q.; Liu, Y.; Yao, Y.; Song, Y. The growth arrest-specific transcript 5 (GAS5): A pivotal tumor suppressor long noncoding RNA in human cancers. Tumor Biol. 2016, 37, 1437–1444. [Google Scholar] [CrossRef]
- Yu, Y.; Hann, S.S. Novel Tumor Suppressor lncRNA Growth Arrest-Specific 5 (GAS5) In Human Cancer. OncoTargets Ther. 2019, 12, 8421–8436. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shen, J.; Chan, M.T.; Wu, W.K.K. TUG1: A pivotal oncogenic long non-coding RNA of human cancers. Cell Prolif. 2016, 49, 471–475. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Kelemen, E.; Danis, J.; Göblös, A.; Bata-Csörgo, Z.; Széll, M. Exosomal long non-coding RNAs as biomarkers in human diseases. Electron. J. Int. Fed. Clin. Chem. Lab. Med. 2019, 30, 224–236. [Google Scholar]













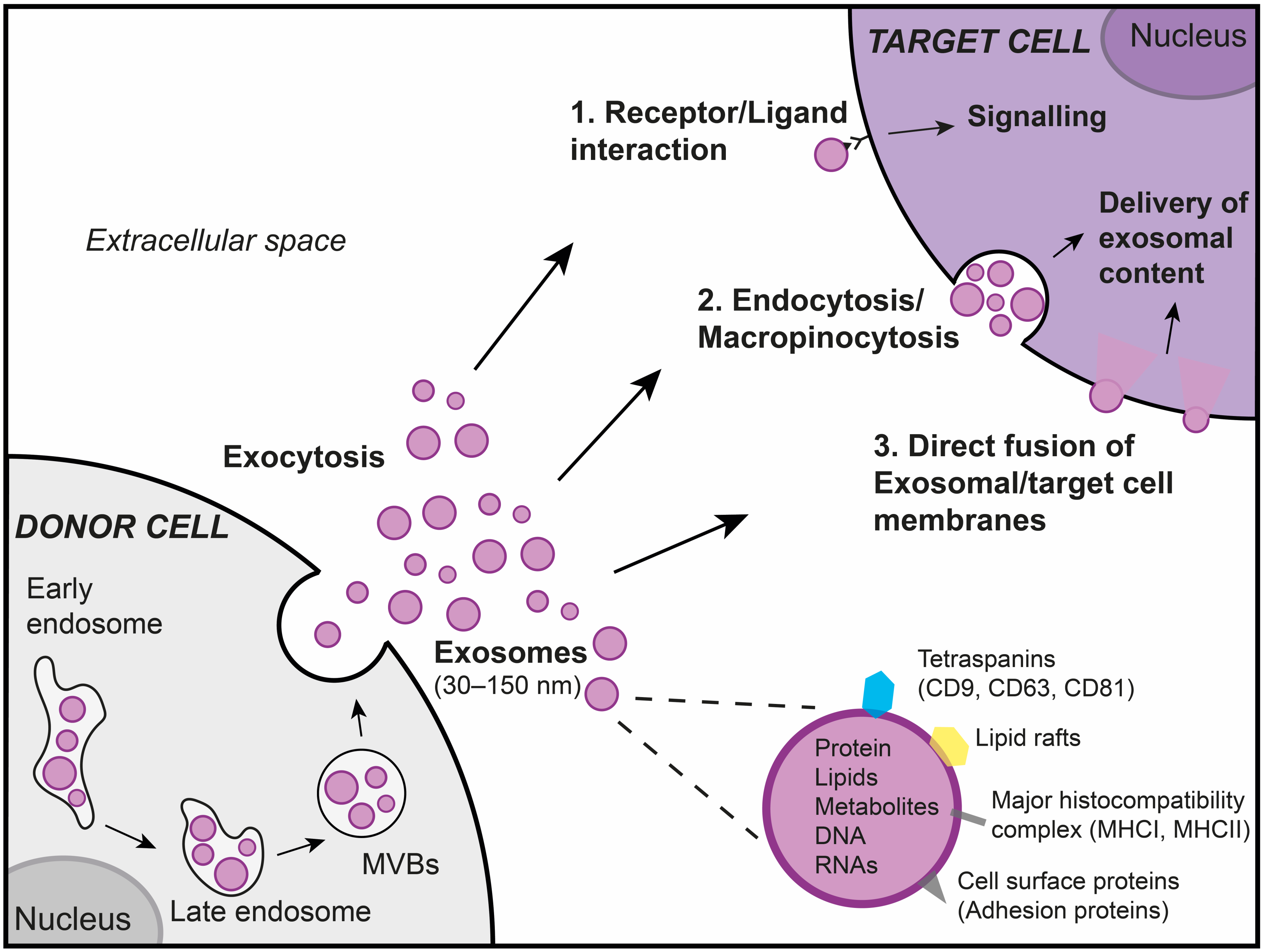{kind=link}
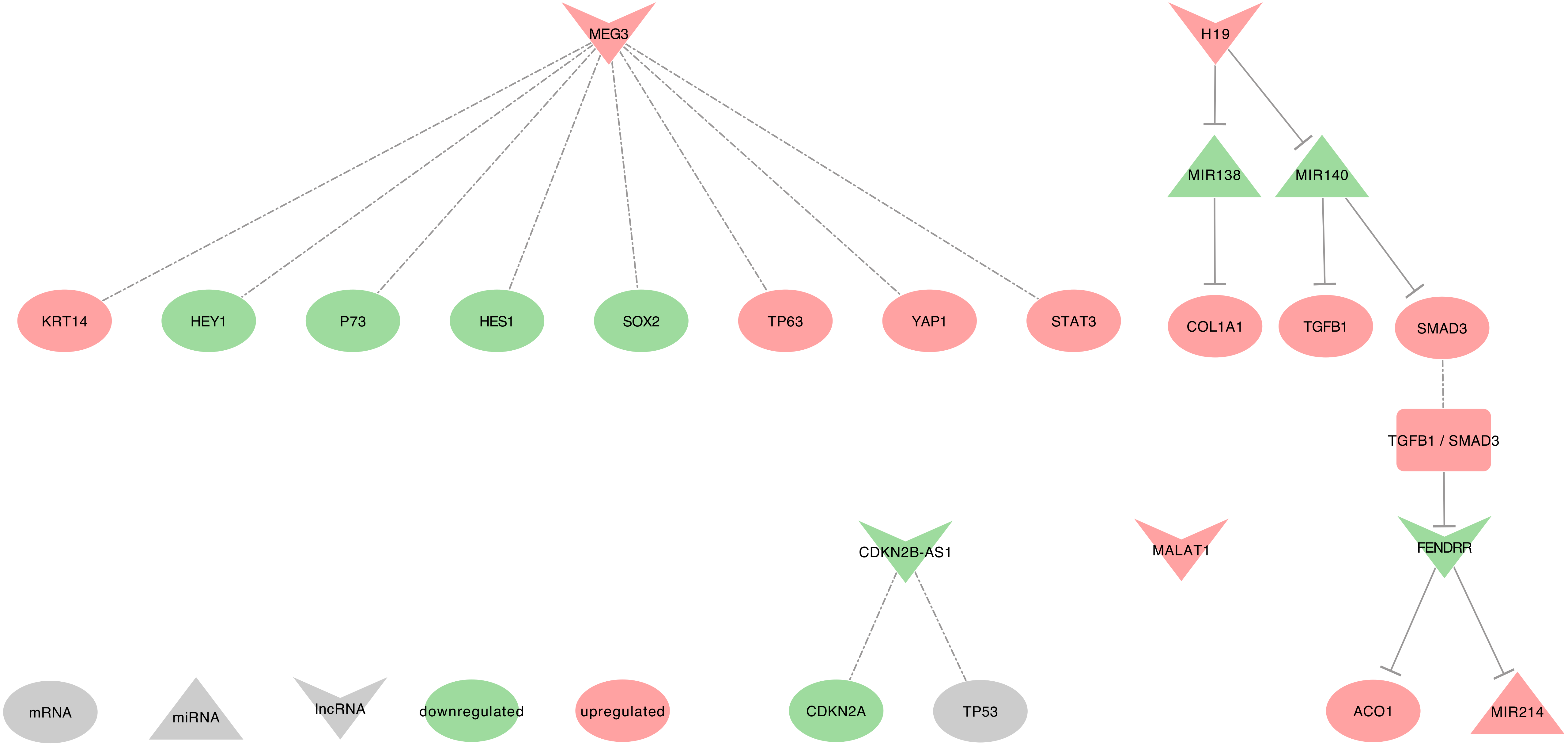{kind=link}
{kind=link}
{kind=link}
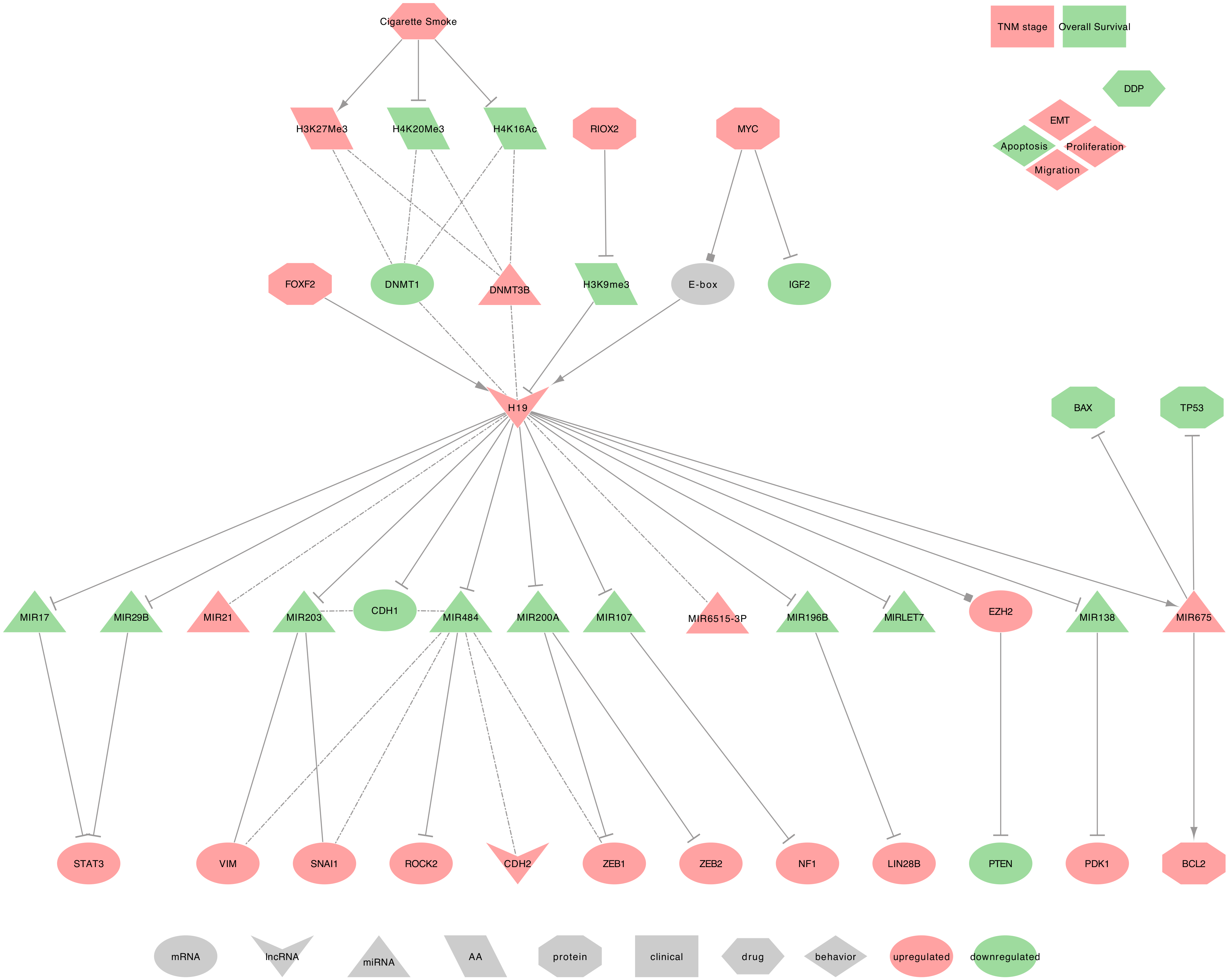{kind=link}
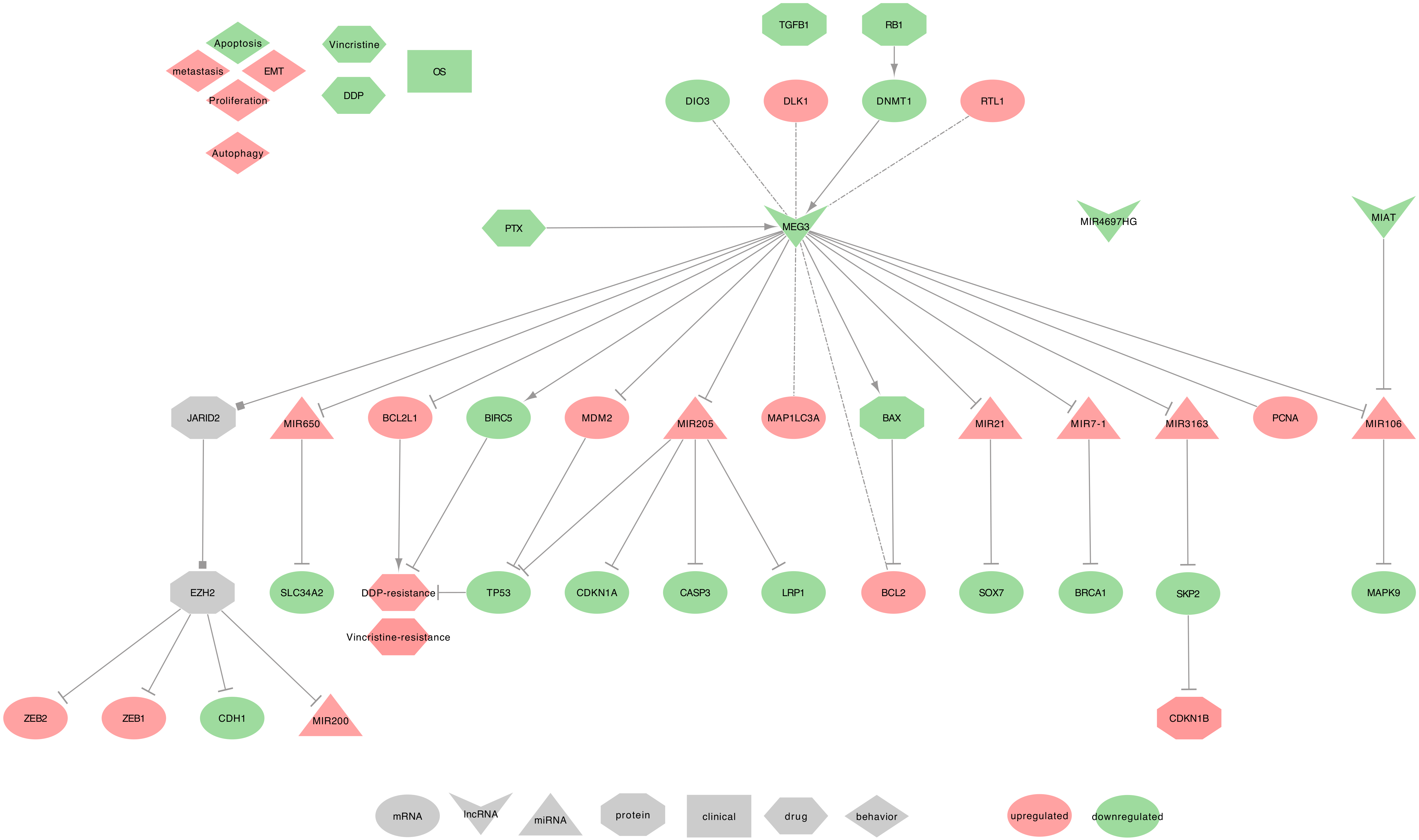{kind=link}
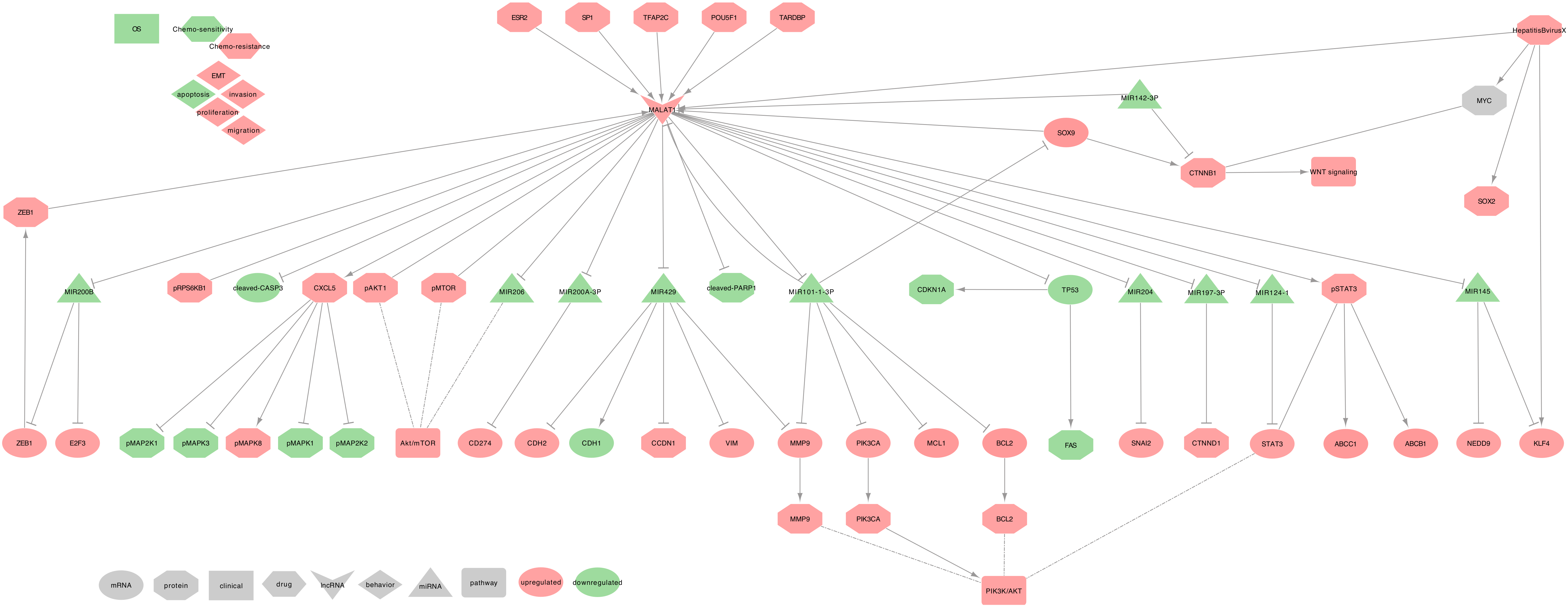{kind=link}
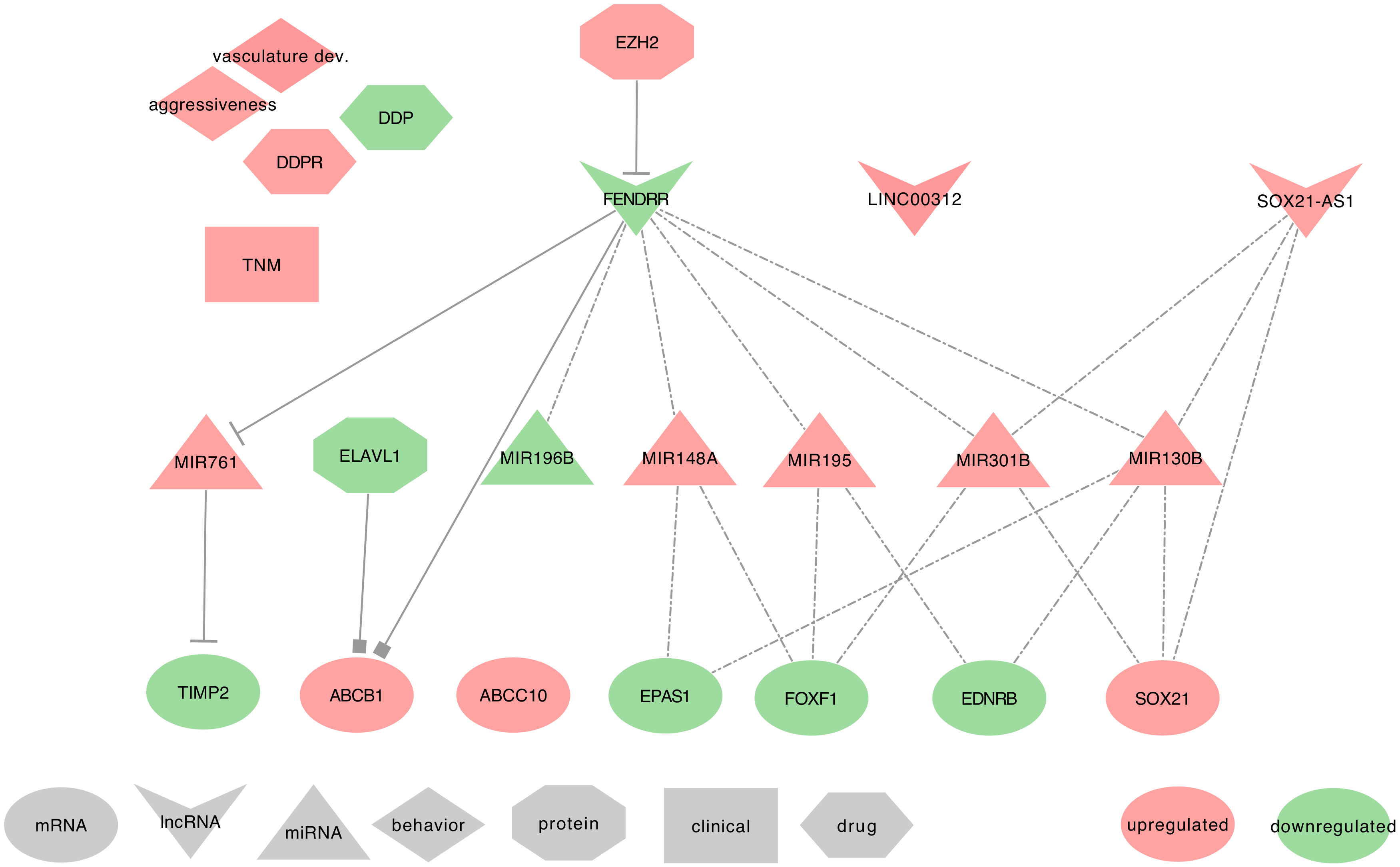{kind=link}
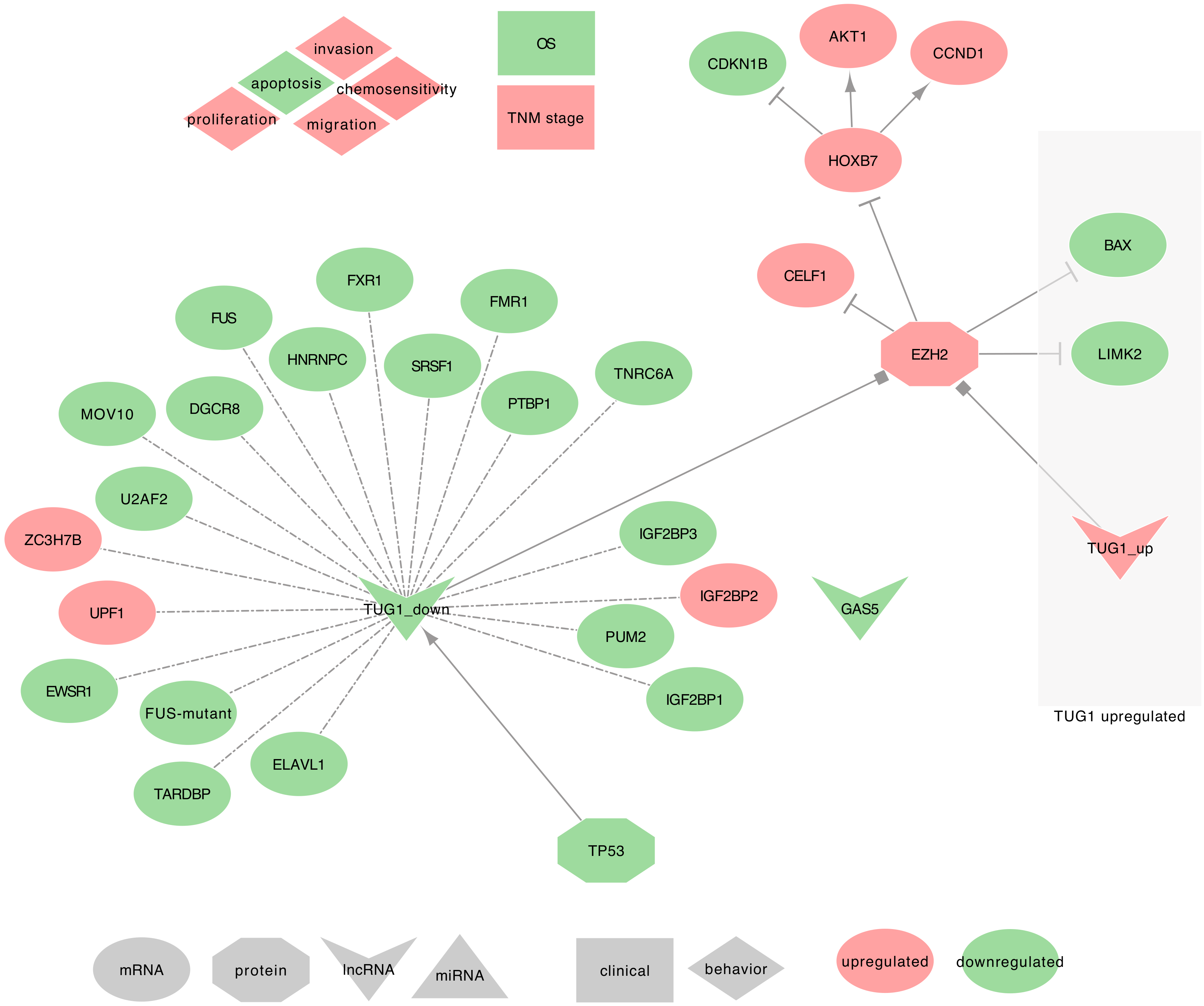{kind=link}
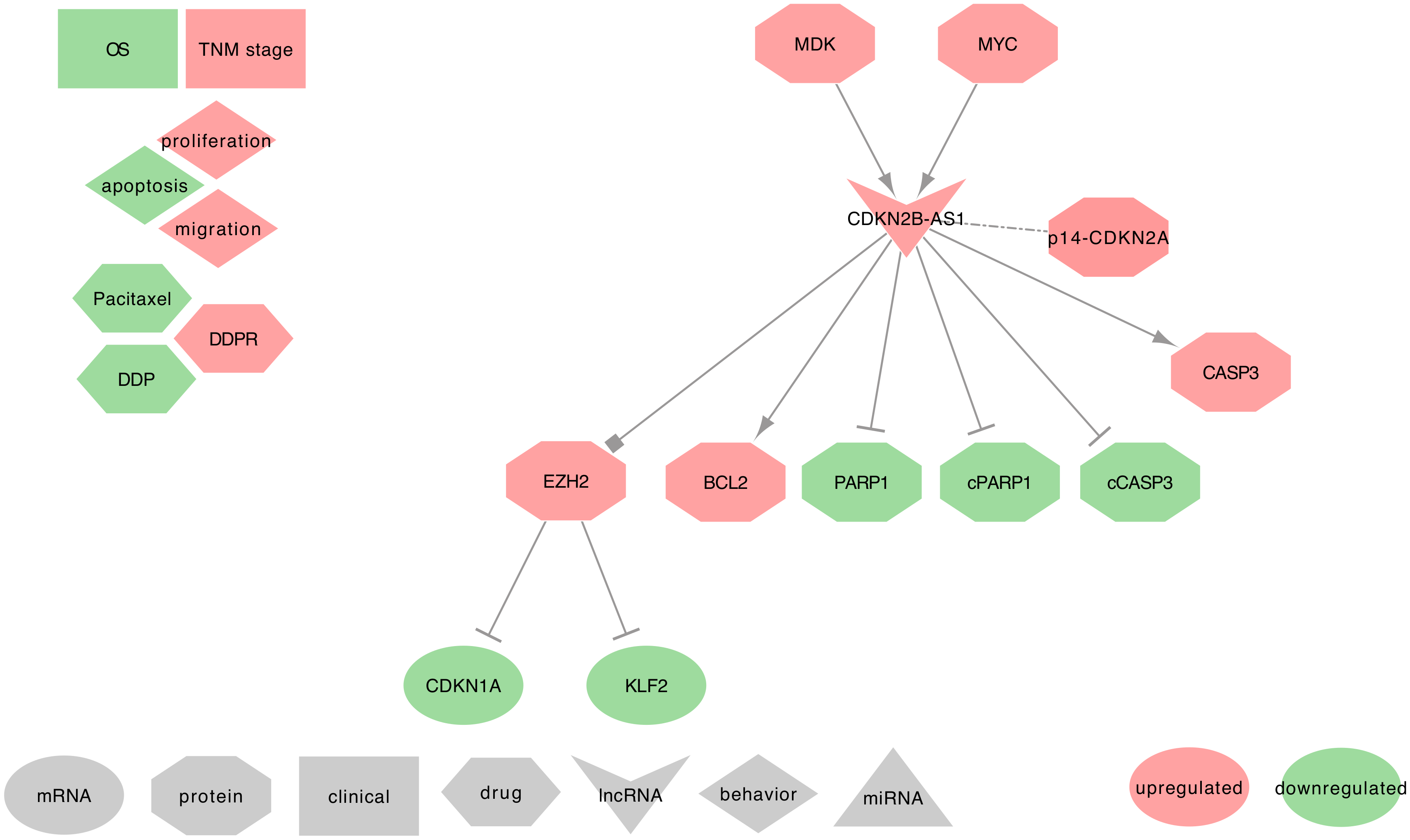{kind=link}
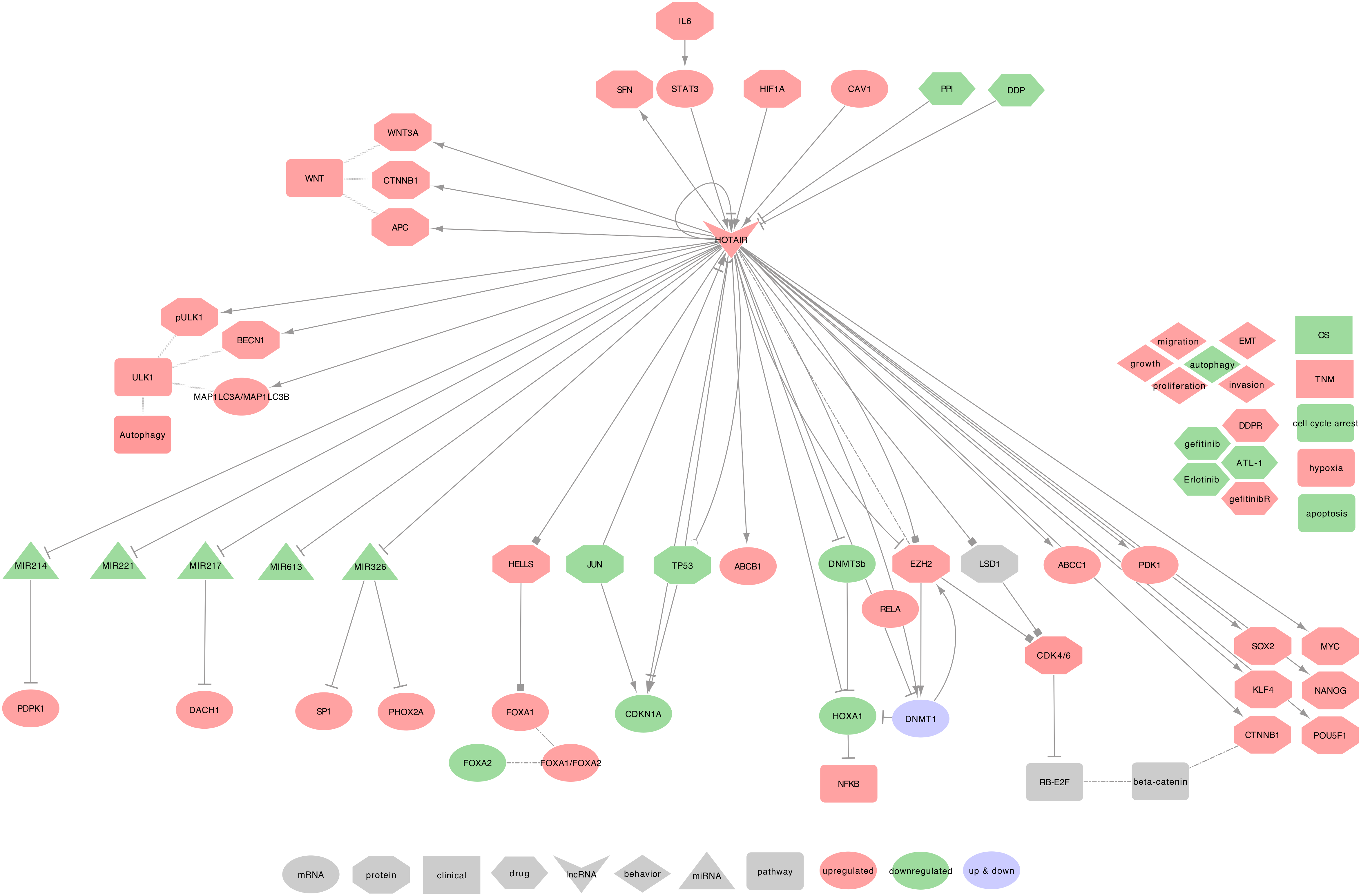{kind=link}
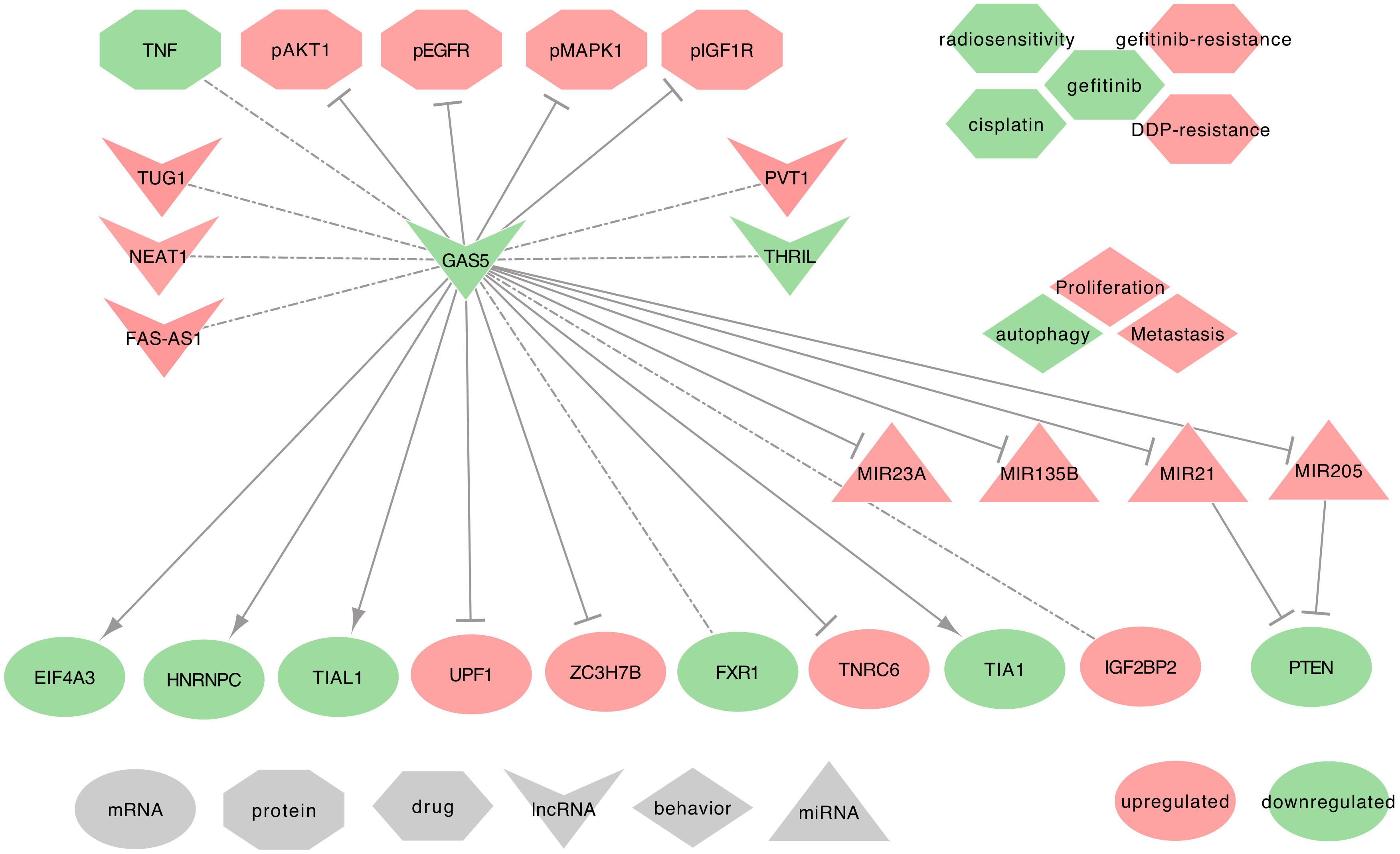{kind=link}
| Disease | LncRNA | Expression | Location | Action | Targeted Pathway | Downstream Targets | References |
|---|---|---|---|---|---|---|---|
| Asthma | CDKN2B-AS1 | up | BA-E & BA-R | Pro-inflammatory | - | TNF, IL17A | [58] |
| GAS5 | up | Severe asthma CD4T-cells | Treg/Th17 balance | - | FOXP3, RORC | [59] | |
| BEAS-2B and primary human ASM cell cultures | Glucocorticoid activity | - | - | [60] | |||
| MALAT1 | up | blood of highly-expressed IgE eosinophilic asthma | Inhibits pathway | T-cell receptor | - | [61] | |
| Severe asthma CD4T-cells | Treg/Th17 balance | - | FOXP3, RORC | [59] | |||
| MEG3 | up | Severe asthma CD4T-cells | Pro-inflammatory | Th17 cell differentiation | IL17A, IL22, RORC | [59] | |
| TUG1 | up | ASM of Sprague Dawley rats | Promotes cell proliferation and migration | - | Fgf1 | [62] | |
| IPF | CDKN2B-AS1 | down | peripheral blood | Activates cell cycle arrest | P53 | CDKN2A, TP53 | [63] |
| FENDRR | down | fibrotic human lung cells and mouse primary lung fibroblasts | Inhibits fibroblast activation & reduces pulmonary fibrosis | TGFB / SMAD3 | ACO1 | [64] | |
| H19 | up | human pulmonary fibrotic tissues | Induces fibrosis | TGFB / SMAD3 | TGB1, SMAD3 | [65] | |
| MEG3 | up | pulmonary epithelial cells from IPF lung tissue | Promotes cell migration | - | TP63, KRT14, STAT3, YAP1, TP73, SOX2, HES1, HEY1 | [66] | |
| COPD | CDKN2B-AS1 | down | plasma of AECOPD | Anti-inflammatory | - | TNF, IL1B, IL17A, CXCL8 | [67] |
| H19 | up | Quadriceps of FFMI patients with COPD | Susceptibility to low FFMI | - | MYOD1 | [68] | |
| HOTAIR | up | CS-exposed male BALB/c mice & HBE cells treated with CSE | - | - | IL6, CXCL8, CDH2, VIM, ACTA2, CDH1 | [69] | |
| MEG3 | up | lung from COPD & CSE-treated 16HBE cells | Induces apoptosis and inflammation | Apoptosis | IL1B, IL6, TNF | [70] | |
| TUG1 | up | sputum and lung from COPD smokers & non-smokers | Inhibits inflammation and airway remodelling | - | DUSP6 | [71] | |
| TGFB1 treated BEAS-2B and HFL1 cells | Inhibits cell proliferation | - | ACTA2, FN1 | [72] | |||
| SCLC | HOTAIR | up | H69 and H446 cell lines | Activates the pathway | NF-κB | HOXA1 | [73] |
| NSCLC | H19 | up | CDK-4/hTERT-immortalized HBEC | Associated with pathway activation | WNT/β-catenin | WNT2, WNT5A, WNT6, WNT10A, FOXN1, TCF7 | [74] |
| MALAT1 | up | Tumor tissues and H1299 cell line | Associated with pathway activation | WNT/β-catenin | - | [75,76] | |
| H1299 and H520 cell lines | Activates the pathway | PTEN / PI3K / AKT | BCL2, MMP9, PIK3CA, STAT3 | [77,78] | |||
| Tumor tissues & A549 and H1299 cell lines | Regulates the pathway | AKT / MTOR | - | [79] | |||
| CDKN2B-AS1 | down | Peripheral blood of IPF | Regulates the pathway | P53 | - | [63] | |
| HOTAIR | up | 95C, 95D and YTMLC-90 cell lines | Regulates the pathway | WNT/β-catenin | RB1, E2F1 | [80] | |
| A549, H460, H1299, NCI-H460 and HCC-827 cell lines | Activates the pathway | Apoptosis | pULK1 | [81] | |||
| GAS5 | down | Tumor tissues & A549, NCI-H1299, H460, SK-MES-1, H157, and H358 cell lines | Regulates the pathway | PTEN / PI3K / AKT | PTEN | [82] | |
| TUG1 | down | Tumor tissues & SPC-A1, NCI-H520, NCI-H520 and NCI-H1299 cell lines | Regulates the pathway | PTEN / PI3K / AKT | PTEN | [83] |
| Disease | Location | LncRNA | Type | Value | Relevancy | References |
|---|---|---|---|---|---|---|
| Asthma | CD4T-cells | GAS5 / MEG3 | expression | upregulated | Up in asthmatic patients vs. healthy patients | [59] |
| peripheral whole blood | MALAT1 | expression | upregulated | Up in highly-expressed IgE eosinophilic asthmatic (EA) patients vs. healthy patients | [61] | |
| plasma | CDKN2B-AS1 | expression | upregulated | Up in patients with bronchial asthma vs. healthy patients | [58] | |
| COPD | plasma | CDKN2B-AS1 | expression | downregulated | Down in patients with acute exacerbations of COPD vs. stable COPD or healthy patients | [67] |
| sputum & lung | TUG1 | expression | upregulated | Up in COPD patients with or without smoking history | [71] | |
| IPF | peripheral whole blood | CDKN2B-AS1 | expression | downregulated | Down in IPF patients, vs. healthy controls May promote the occurrence of lung cancers | [63] |
| Lung cancers | constitutive DNA | H19 | polymorphism | rs217727 C >T | Associated with increased risk of lung cancer in meta-analysis | [84] |
| MEG3 | rs4081134 G >A | Genotype [AA] associated with lung cancer risk in chinese northeast population | [85] | |||
| LUAD | serum | TUG1 | expression | upregulated | Up in LUAD patients vs. healthy patients | [86] |
| tumor | CDKN2B-AS1 | expression | upregulated | Up in cell lines & positively correlated with the differentiation grade and the TNM stages | [87] | |
| tumor | FENDRR | expression | downregulated | Strongly associated with High TNM 1 stage in LUAD patients vs. healthy patients. Predicts LUAD cancer vs. healthy state when associated with LINC00312 | [88] | |
| NSCLC | plasma | H19 | expression | upregulated | Up in NSCLC vs. begnin lung disease | [89] |
| serum | MALAT1 | expression | downregulated | Down in patients with NSCLC vs. healthy patients | [90,91,92,93] | |
| tumor | CDKN2B-AS1 | expression | upregulated | Correlated with poor patient OS | [94] | |
| GAS5 | expression | downregulated | Down in male subjects vs. corresponding ANCTs. | [95] | ||
| H19 | expression | upregulated | Up in stage III and IV vs. stage I and II & negatively correlated with OS | [96,97] | ||
| HOTAIR | expression | upregulated | Up in patients with stage III and IV vs. stage I and II. Positively correlated with greater tumor size, lymph node metastasis or lymph-vascular invasion, short disease free interval, and reduced OS. | [98,99,100] | ||
| MALAT1 | expression | upregulated | Associated with a poor prognosis and short OS. Associated with age, tumour size & TNM stage, when combined to SOX9 | [76,79,101,102,103,104,105] | ||
| MEG3 | expression | downregulated | Associated with short-term survival | [106] | ||
| TUG1 | expression | downregulated | Associated with a high TNM stage and a poor patient outcome | [105,107,108] | ||
| SCC | sputum | H19/HOTAIR | expression | - | Diagnosis of SCC vs. LUAD | [109] |
| Disease | Treatment | LncRNA | Expression | Relevancy | References |
|---|---|---|---|---|---|
| LUAD | Gefitinib | GAS5 | down | Overexpression increases sensitivity to treatment | [110] |
| Ionizing radiation | Overexpression increases radiosensitivity | [111] | |||
| Cisplatine | HOTAIR | up | Repression increases sensitivity to treatment | [112,113] | |
| Erlotinib | Repression of PDK1 and HOTAIR-mediated EZH2 gene expression increases sensitivity to treatment | [114] | |||
| Atractylenolide 1 | |||||
| SCLC | Cisplatine | HOTAIR | up | Repression increases sensitivity to treatment | [115] |
| Adriamycin | |||||
| Etoposide | |||||
| NSCLC | Cisplatine | FENDRR | down | Negatively correlated with treatment response | [116] |
| GAS5 | down | Could regulate chemo-resistance to treatment | [82] | ||
| H19 | up | Negatively correlated with treatment response | [96] | ||
| MALAT1 | up | Increases resistance to treatment through positive feedback loop with SOX9 | [76] | ||
| MEG3 | down | Overexpression increases sensitivity to treatment | [117,118,119] | ||
| TUG1 | down | Overexpression increases sensitivity to treatment | [83] | ||
| Crizotinib | HOTAIR | up | Repression increases sensitivity to treatment | [81] | |
| Paclitaxel | CDKN2B-AS1 | up | Inhibits sensitivity to treatment | [87] | |
| Vincristine | MEG3 | down | Overexpression increases sensitivity to treatment | [117,120] | |
| OSCC | Cisplatine | CDKN2B-AS1 | up | Associated with Midkine to treatment resistance | [121] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poulet, C.; Njock, M.-S.; Moermans, C.; Louis, E.; Louis, R.; Malaise, M.; Guiot, J. Exosomal Long Non-Coding RNAs in Lung Diseases. Int. J. Mol. Sci. 2020, 21, 3580. https://doi.org/10.3390/ijms21103580
Poulet C, Njock M-S, Moermans C, Louis E, Louis R, Malaise M, Guiot J. Exosomal Long Non-Coding RNAs in Lung Diseases. International Journal of Molecular Sciences. 2020; 21(10):3580. https://doi.org/10.3390/ijms21103580
Chicago/Turabian StylePoulet, Christophe, Makon-Sébastien Njock, Catherine Moermans, Edouard Louis, Renaud Louis, Michel Malaise, and Julien Guiot. 2020. "Exosomal Long Non-Coding RNAs in Lung Diseases" International Journal of Molecular Sciences 21, no. 10: 3580. https://doi.org/10.3390/ijms21103580
APA StylePoulet, C., Njock, M.-S., Moermans, C., Louis, E., Louis, R., Malaise, M., & Guiot, J. (2020). Exosomal Long Non-Coding RNAs in Lung Diseases. International Journal of Molecular Sciences, 21(10), 3580. https://doi.org/10.3390/ijms21103580