Tunneling Nanotubes-Mediated Protection of Mesenchymal Stem Cells: An Update from Preclinical Studies


Abstract
1. Introduction
2. Preclinical Studies
2.1. Ischemia/Reperfusion Model
2.2. Chemotherapy and Other Stress-Induced Models
2.3. Stress Model from MELAS Patients
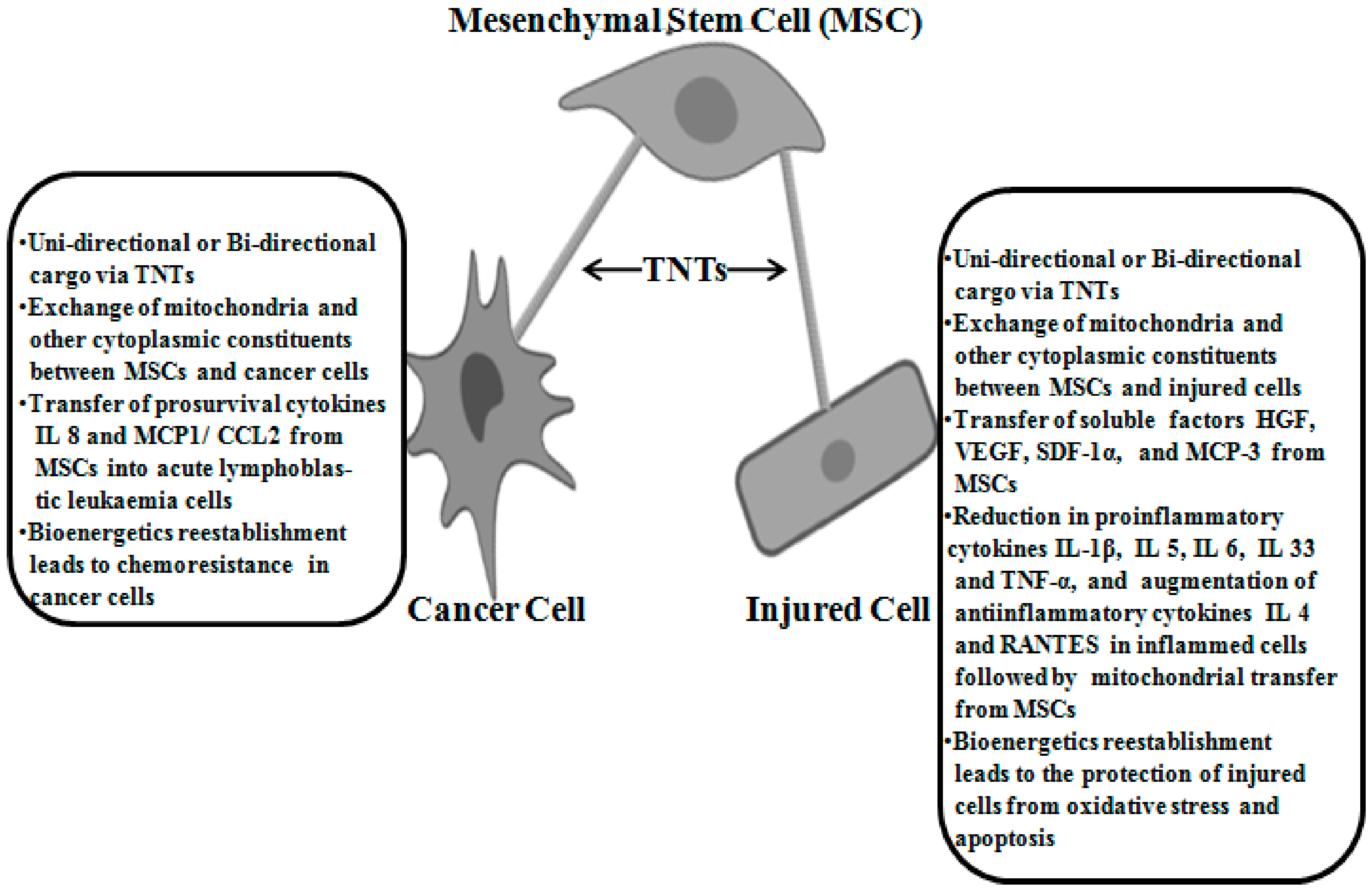
2.4. Inflammatory and Respiratory Disease Models
2.5. Other Non-Cancer Models
2.6. Cancer Models
3. Future Directions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNTs | Tunneling nanotubes |
| MSCs | Mesenchymal stem cells |
| MHCII | Major histocompatibility complex II |
| HGF | Hepatocyte growth factor |
| VEGF | Vascular endothelial growth factor |
| SDF-1α | Stromal cell-derived factor-1α |
| MCP-3 | Monocyte chemotactic protein-3 |
| Miro1 | Mitochondrial Rho-GTPase 1 |
| iPSC | Induced pluripotent stem cells |
| NAC | N-acetyl-L-cysteine |
| AAP | L-ascorbic acid 2-phosphate |
| DRP1 | Dynamin-related protein 1 |
| MELAS | Mitochondrial myopathy, Encephalopathy, Lactic acidosis, and Stroke-like episodes |
| WJMSCs | Wharton’s jelly mesenchymal stem cells |
| VSMCs | Vascular smooth muscle cells |
| TNF-α | Tumor necrosis factor-alpha |
| CoCl2 | Cobalt (II) chloride |
| BCP-ALL | B-cell precursor acute lymphoblastic leukemia |
| AML | Acute myeloid leukemia |
| T-ALL | T cell acute lymphoblastic leukemia |
References
- Drab, M.; Stopar, D.; Kralj-Iglic, V.; Iglic, A. Inception Mechanisms of Tunneling Nanotubes. Cells 2019, 8. [Google Scholar] [CrossRef]
- Dupont, M.; Souriant, S.; Lugo-Villarino, G.; Maridonneau-Parini, I.; Verollet, C. Tunneling Nanotubes: Intimate Communication between Myeloid Cells. Front. Immunol. 2018, 9, 43. [Google Scholar] [CrossRef]
- Musial-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transplant. 2019, 28, 801–812. [Google Scholar] [CrossRef]
- Machado Cde, V.; Telles, P.D.; Nascimento, I.L. Immunological characteristics of mesenchymal stem cells. Rev. Bras. De Hematol. E Hemoter. 2013, 35, 62–67. [Google Scholar] [CrossRef]
- Spees, J.L.; Lee, R.H.; Gregory, C.A. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res. Ther. 2016, 7, 125. [Google Scholar] [CrossRef]
- Liu, K.; Ji, K.; Guo, L.; Wu, W.; Lu, H.; Shan, P.; Yan, C. Mesenchymal stem cells rescue injured endothelial cells in an in vitro ischemia-reperfusion model via tunneling nanotube like structure-mediated mitochondrial transfer. Microvasc. Res. 2014, 92, 10–18. [Google Scholar] [CrossRef]
- Han, H.; Hu, J.; Yan, Q.; Zhu, J.; Zhu, Z.; Chen, Y.; Sun, J.; Zhang, R. Bone marrow-derived mesenchymal stem cells rescue injured H9c2 cells via transferring intact mitochondria through tunneling nanotubes in an in vitro simulated ischemia/reperfusion model. Mol. Med. Rep. 2016, 13, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Figeac, F.; Lesault, P.F.; Le Coz, O.; Damy, T.; Souktani, R.; Trebeau, C.; Schmitt, A.; Ribot, J.; Mounier, R.; Guguin, A.; et al. Nanotubular crosstalk with distressed cardiomyocytes stimulates the paracrine repair function of mesenchymal stem cells. Stem Cells 2014, 32, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Babenko, V.A.; Silachev, D.N.; Popkov, V.A.; Zorova, L.D.; Pevzner, I.B.; Plotnikov, E.Y.; Sukhikh, G.T.; Zorov, D.B. Miro1 Enhances Mitochondria Transfer from Multipotent Mesenchymal Stem Cells (MMSC) to Neural Cells and Improves the Efficacy of Cell Recovery. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Guo, L.; Zhou, Z.; Pan, M.; Yan, C. Mesenchymal stem cells transfer mitochondria into cerebral microvasculature and promote recovery from ischemic stroke. Microvasc. Res. 2019, 123, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, J.; Zhao, L.; Xin, Y.; Liu, S.; Cui, W. Differential roles of microtubules in the two formation stages of membrane nanotubes between human mesenchymal stem cells and neonatal mouse cardiomyocytes. Biochem. Biophys. Res. Commun. 2019, 512, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Borg, T.K.; Ma, Z.; Xu, M.; Wetzel, G.; Saraf, L.V.; Markwald, R.; Runyan, R.B.; Gao, B.Z. Biochip-based study of unidirectional mitochondrial transfer from stem cells to myocytes via tunneling nanotubes. Biofabrication 2016, 8, 015012. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhu, R.; Shen, J.; Wu, J.; Lu, W.; Zhang, J.; Zhang, J.; Liu, K. Human Bone Marrow Mesenchymal Stem Cells Rescue Endothelial Cells Experiencing Chemotherapy Stress by Mitochondrial Transfer Via Tunneling Nanotubes. Stem Cells Dev. 2019, 28, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Gao, F.; Zhang, Y.; Wong, D.S.; Li, Q.; Tse, H.F.; Xu, G.; Yu, Z.; Lian, Q. Mitochondrial transfer of mesenchymal stem cells effectively protects corneal epithelial cells from mitochondrial damage. Cell Death Dis. 2016, 7, e2467. [Google Scholar] [CrossRef]
- Li, C.J.; Chen, P.K.; Sun, L.Y.; Pang, C.Y. Enhancement of Mitochondrial Transfer by Antioxidants in Human Mesenchymal Stem Cells. Oxidative Med. Cell. Longev. 2017, 2017, 8510805. [Google Scholar] [CrossRef]
- Lin, T.K.; Chen, S.D.; Chuang, Y.C.; Lan, M.Y.; Chuang, J.H.; Wang, P.W.; Hsu, T.Y.; Wang, F.S.; Tsai, M.H.; Huang, S.T.; et al. Mitochondrial Transfer of Wharton’s Jelly Mesenchymal Stem Cells Eliminates Mutation Burden and Rescues Mitochondrial Bioenergetics in Rotenone-Stressed MELAS Fibroblasts. Oxidative Med. Cell. Longev. 2019, 2019, 9537504. [Google Scholar] [CrossRef]
- Hu, J.; Deng, G.; Tian, Y.; Pu, Y.; Cao, P.; Yuan, W. An in vitro investigation into the role of bone marrowderived mesenchymal stem cells in the control of disc degeneration. Mol. Med. Rep. 2015, 12, 5701–5708. [Google Scholar] [CrossRef]
- Lehmann, T.P.; Filipiak, K.; Juzwa, W.; Sujka-Kordowska, P.; Jagodzinski, P.P.; Zabel, M.; Glowacki, J.; Misterska, E.; Walczak, M.; Glowacki, M. Coculture of human nucleus pulposus cells with multipotent mesenchymal stromal cells from human bone marrow reveals formation of tunnelling nanotubes. Mol. Med. Rep. 2014, 9, 574–582. [Google Scholar] [CrossRef]
- Vallabhaneni, K.C.; Haller, H.; Dumler, I. Vascular smooth muscle cells initiate proliferation of mesenchymal stem cells by mitochondrial transfer via tunneling nanotubes. Stem Cells Dev. 2012, 21, 3104–3113. [Google Scholar] [CrossRef]
- Matula, Z.; Nemeth, A.; Lorincz, P.; Szepesi, A.; Brozik, A.; Buzas, E.I.; Low, P.; Nemet, K.; Uher, F.; Urban, V.S. The Role of Extracellular Vesicle and Tunneling Nanotube-Mediated Intercellular Cross-Talk Between Mesenchymal Stem Cells and Human Peripheral T Cells. Stem Cells Dev. 2016, 25, 1818–1832. [Google Scholar] [CrossRef]
- Islam, M.N.; Das, S.R.; Emin, M.T.; Wei, M.; Sun, L.; Westphalen, K.; Rowlands, D.J.; Quadri, S.K.; Bhattacharya, S.; Bhattacharya, J. Mitochondrial transfer from bone-marrow-derived stromal cells to pulmonary alveoli protects against acute lung injury. Nat. Med. 2012, 18, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.A.; Yerkovich, S.T.; Hopkins, P.M.; Chambers, D.C. Characterization of intercellular communication and mitochondrial donation by mesenchymal stromal cells derived from the human lung. Stem Cell Res. Ther. 2016, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Mukherjee, S.; Pattnaik, B.; Kumar, M.; Singh, S.; Kumar, M.; Rehman, R.; Tiwari, B.K.; Jha, K.A.; Barhanpurkar, A.P.; et al. Miro1 regulates intercellular mitochondrial transport & enhances mesenchymal stem cell rescue efficacy. EMBO J. 2014, 33, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Yeung, S.C.; Liang, Y.; Liang, X.; Ding, Y.; Ip, M.S.; Tse, H.F.; Mak, J.C.; Lian, Q. Mitochondrial transfer of induced pluripotent stem cell-derived mesenchymal stem cells to airway epithelial cells attenuates cigarette smoke-induced damage. Am. J. Respir. Cell Mol. Biol. 2014, 51, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.V.; Morrison, T.J.; Doherty, D.F.; McAuley, D.F.; Matthay, M.A.; Kissenpfennig, A.; O’Kane, C.M.; Krasnodembskaya, A.D. Mitochondrial Transfer via Tunneling Nanotubes is an Important Mechanism by Which Mesenchymal Stem Cells Enhance Macrophage Phagocytosis in the In Vitro and In Vivo Models of ARDS. Stem Cells 2016, 34, 2210–2223. [Google Scholar] [CrossRef]
- Montgomery, M.K. Mitochondrial Dysfunction and Diabetes: Is Mitochondrial Transfer a Friend or Foe? Biology 2019, 8. [Google Scholar] [CrossRef]
- Hough, K.P.; Deshane, J.S. Exosomes in Allergic Airway Diseases. Curr. Allergy Asthma Rep. 2019, 19, 26. [Google Scholar] [CrossRef]
- Yao, Y.; Fan, X.L.; Jiang, D.; Zhang, Y.; Li, X.; Xu, Z.B.; Fang, S.B.; Chiu, S.; Tse, H.F.; Lian, Q.; et al. Connexin 43-Mediated Mitochondrial Transfer of iPSC-MSCs Alleviates Asthma Inflammation. Stem Cell Rep. 2018, 11, 1120–1135. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Khryapenkova, T.G.; Galkina, S.I.; Sukhikh, G.T.; Zorov, D.B. Cytoplasm and organelle transfer between mesenchymal multipotent stromal cells and renal tubular cells in co-culture. Exp. Cell Res. 2010, 316, 2447–2455. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, G.; Zhang, Y.L.; He, H.W.; Yu, B.Q.; Hong, Y.M.; You, W.; Li, X. Transfer of mitochondria from mesenchymal stem cells derived from induced pluripotent stem cells attenuates hypoxia-ischemia-induced mitochondrial dysfunction in PC12 cells. Neural Regen. Res. 2020, 15, 464–472. [Google Scholar] [CrossRef]
- Pasquier, J.; Guerrouahen, B.S.; Al Thawadi, H.; Ghiabi, P.; Maleki, M.; Abu-Kaoud, N.; Jacob, A.; Mirshahi, M.; Galas, L.; Rafii, S.; et al. Preferential transfer of mitochondria from endothelial to cancer cells through tunneling nanotubes modulates chemoresistance. J. Transl. Med. 2013, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Polak, R.; de Rooij, B.; Pieters, R.; den Boer, M.L. B-cell precursor acute lymphoblastic leukemia cells use tunneling nanotubes to orchestrate their microenvironment. Blood 2015, 126, 2404–2414. [Google Scholar] [CrossRef] [PubMed]
- Marlein, C.R.; Zaitseva, L.; Piddock, R.E.; Robinson, S.D.; Edwards, D.R.; Shafat, M.S.; Zhou, Z.; Lawes, M.; Bowles, K.M.; Rushworth, S.A. NADPH oxidase-2 derived superoxide drives mitochondrial transfer from bone marrow stromal cells to leukemic blasts. Blood 2017, 130, 1649–1660. [Google Scholar] [CrossRef]
- Marlein, C.R.; Piddock, R.E.; Mistry, J.J.; Zaitseva, L.; Hellmich, C.; Horton, R.H.; Zhou, Z.; Auger, M.J.; Bowles, K.M.; Rushworth, S.A. CD38-Driven Mitochondrial Trafficking Promotes Bioenergetic Plasticity in Multiple Myeloma. Cancer Res. 2019, 79, 2285–2297. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Qiu, Y.; Shi, Y.; Cai, J.; Wang, B.; Wei, X.; Ke, Q.; Sui, X.; Wang, Y.; et al. Cell adhesion-mediated mitochondria transfer contributes to mesenchymal stem cell-induced chemoresistance on T cell acute lymphoblastic leukemia cells. J. Hematol. Oncol. 2018, 11, 11. [Google Scholar] [CrossRef]
- Burt, R.; Dey, A.; Aref, S.; Aguiar, M.; Akarca, A.; Bailey, K.; Day, W.; Hooper, S.; Kirkwood, A.; Kirschner, K.; et al. Activated stromal cells transfer mitochondria to rescue acute lymphoblastic leukemia cells from oxidative stress. Blood 2019, 134, 1415–1429. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Peault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef]
- Agnati, L.F.; Guidolin, D.; Guescini, M.; Genedani, S.; Fuxe, K. Understanding wiring and volume transmission. Brain Res. Rev. 2010, 64, 137–159. [Google Scholar] [CrossRef]
- Babenko, V.A.; Silachev, D.N.; Zorova, L.D.; Pevzner, I.B.; Khutornenko, A.A.; Plotnikov, E.Y.; Sukhikh, G.T.; Zorov, D.B. Improving the Post-Stroke Therapeutic Potency of Mesenchymal Multipotent Stromal Cells by Cocultivation With Cortical Neurons: The Role of Crosstalk Between Cells. Stem Cells Transl. Med. 2015, 4, 1011–1020. [Google Scholar] [CrossRef]
- Lee, H.Y.; Hong, I.S. Double-edged sword of mesenchymal stem cells: Cancer-promoting versus therapeutic potential. Cancer Sci. 2017, 108, 1939–1946. [Google Scholar] [CrossRef]


{kind=link}
| Field of Study | MSCs Source | Experimental Model | Results | Ref. |
|---|---|---|---|---|
| Ischemia/reperfusion | Human bone marrow MSCs | Human umbilical vein endothelial cell exposed to OGD | Reduced apoptosis | [6] |
| Ischemia/reperfusion | Rat bone marrow MSCs | H9c2 rat ventricular cell line in vitro model of SI/R | Reduced apoptosis | [7] |
| Ischemia/reperfusion | Human adipose MSCs | In vitro: Mouse cardiomyocyte; In vivo: mouse subjected to MI | In vitro: increased release of cardioprotective soluble factors by MSCs; In vivo: greater cardiac function recovery associated with higher angiogenesis | [8] |
| Ischemia/reperfusion | Human bone marrow MSCs | In vitro: Rat astrocytes exposed to OGD and PC12 pheochromocytoma cells exposed to ethidium bromide; In vivo: Rats subjected to MCAO | In vitro: MSCs rescued astrocytes and PC12 cells from damage; In vivo: MSCs, especially those overexpressing Miro1, ameliorated neurological impairment | [9] |
| Ischemia/reperfusion | Rat bone marrow MSCs | Rats subjected to MCAO | Increase in angiogenesis, improvements in functional recovery | [10] |
| Ischemia/reperfusion | Human umbilical cord MSCs | Neonatal mouse cardiomyocytes exposed to hypoxia/reoxygenation | Reduced apoptosis | [11] |
| Ischemia/reperfusion | Rat bone marrow MSCs | Neonatal rat cardiomyocytes | Initial TNTs formation derived from MSCs, while after 24h majority of TNTs derived from cardiomyocytes | [12] |
| Chemotherapy | Human bone marrow MSCs | Human umbilical cord vein endothelial cells treated with cytarabine | Reduced apoptosis, restored transmembrane migration ability and angiogenic capacity of endothelial cells | [13] |
| Oxidative stress | Human iPSC-derived MSCs | In vitro: Corneal epithelial cells exposed to rotenone; In vivo: Rabbit model of corneal alkaline burn induced by sodium hydroxide | In vitro: protection against rotenone oxidative stress; In vivo: beneficial effects for corneal wound recovery | [14] |
| Oxidative stress | Human adipose derived MSCs | MSCs exposed to hydrogen peroxide, N-acetyl-L-cysteine, and L-ascorbic acid 2-phosphate | Antioxidants increased mitochondrial mass and respiratory capacity | [15] |
| MELAS patients | Wharton’s jelly MSCs | Human MELAS fibroblasts treated with rotenone | Improved mitochondrial stress | [16] |
| Inflammation | Rat bone marrow MSCs | Rat nucleus pulposus cells exposed to IL-1β | Reduced apoptosis only in direct coculture | [17] |
| Human bone marrow MSCs | Human nucleus pulposus cells | TNTs formation in coculture system resulting in phenotypic changes in human nucleus pulposus cells | [18] | |
| Inflammatory disease | Human bone marrow MSCs | Human vascular smooth muscle cells | Increased MSCs proliferation, but not differentiation | [19] |
| Immunomodulation | Human adipose derived MSCs | Human peripheral T lymphocytes | Bi-directional cytoplasmic content exchanges | [20] |
| Lung injury | Mouse and human bone marrow MSCs | Mouse acute lung injury model | Mitochondria transfer by TNTs exerted protective effects | [21] |
| Lung injury | Human MSCs derived from digested parenchymal lung tissue and from lung transplant recipients’ bronchoalveolar lavage fluid | Human bronchial epithelial BEAS-2B cells | Formation of TNTs between MSCs and epithelial cells | [22] |
| Lung injury | Human bone marrow MSCs | In vivo: Mouse model of rotenone-induced airway epithelial injury and allergic airway inflammation In vitro: Mouse bronchial epithelial cells treated with conditioned medium derived from macrophages exposed to IL-13 | Protection exerted by TNTs mitochondrial transfer. MSCs overexpressing Miro1 showed increased mitochondria transfer | [23] |
| Respiratory disease | Human iPSC-derived MSCs and bone marrow MSCs | In vitro: Human bronchial epithelial BEAS-2B cells exposed to cigarette smoke medium In vivo: Rats injured by cigarette smoke | In vitro: attenuated mitochondrial dysfunction In vivo: improved alveolar destruction | [24] |
| Respiratory disease | Human bone marrow MSCs | In vitro: Human monocyte-derived macrophages exposed to LPS or Escherichia coli In vivo: Mice subjected to Escherichia coli pneumonia | In vitro: enhanced phagocytic capacity; In vivo: enhanced alveolar macrophages phagocytosis, decreased inflammation, and functional recovery | [25] |
| Respiratory disease | Human iPSC-derived MSCs | In vivo: Mice subjected to asthmatic inflammation by treatment with ovalbumin; In vitro: Human bronchial epithelium cell line BEAS-2B cells treated with CoCl2 | In vivo: Attenuated mitochondrial dysfunction and inflammation; In vitro: Attenuated mitochondrial dysfunction | [28] |
| Human MSCs | Rat renal tubular cells | Bi-directional cytoplasmic content exchanges and MSCs differentiation | [29] | |
| Mitochondria damage | Human iPSC-derived MSCs | PC12 cells treated with CoCl2 | Attenuated mitochondrial dysfunction | [30] |
| Cancer | MSCs | Human cancer cell lines, such as SKOV3 ovarian cancer cells and MCF7 breast cancer cells | Exchange of cytoplasmic components | [31] |
| Cancer | Human bone marrow MSCs | Primary B cell precursor acute lymphoblastic leukemia cells (BCP-ALL) | Chemoresistance of BCP-ALL to prednisolone | [32] |
| Cancer | Human bone marrow MSCs | Acute myeloid leukemia cells | NADPH oxidase-2 derived superoxide drove mitochondrial transfer from MSCs to leukemic cells | [33] |
| Cancer | Human bone marrow MSCs | Human multiple myeloma cells | CD-38 dependent TNTs formation | [34] |
| Cancer | Human bone marrow MSCs | Human T cell acute lymphoblastic leukemia cell line Jurkat exposed to methotrexate and cytarabine | Chemoresistance from drug-induced apoptosis | [35] |
| Cancer | Human bone marrow MSCs | Human acute lymphoblastic leukemia cell lines | Reduced drug-induced apoptosis | [36] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soundara Rajan, T.; Gugliandolo, A.; Bramanti, P.; Mazzon, E. Tunneling Nanotubes-Mediated Protection of Mesenchymal Stem Cells: An Update from Preclinical Studies. Int. J. Mol. Sci. 2020, 21, 3481. https://doi.org/10.3390/ijms21103481
Soundara Rajan T, Gugliandolo A, Bramanti P, Mazzon E. Tunneling Nanotubes-Mediated Protection of Mesenchymal Stem Cells: An Update from Preclinical Studies. International Journal of Molecular Sciences. 2020; 21(10):3481. https://doi.org/10.3390/ijms21103481
Chicago/Turabian StyleSoundara Rajan, Thangavelu, Agnese Gugliandolo, Placido Bramanti, and Emanuela Mazzon. 2020. "Tunneling Nanotubes-Mediated Protection of Mesenchymal Stem Cells: An Update from Preclinical Studies" International Journal of Molecular Sciences 21, no. 10: 3481. https://doi.org/10.3390/ijms21103481
APA StyleSoundara Rajan, T., Gugliandolo, A., Bramanti, P., & Mazzon, E. (2020). Tunneling Nanotubes-Mediated Protection of Mesenchymal Stem Cells: An Update from Preclinical Studies. International Journal of Molecular Sciences, 21(10), 3481. https://doi.org/10.3390/ijms21103481