Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa


 ,
,
Abstract
1. Introduction
2. Results
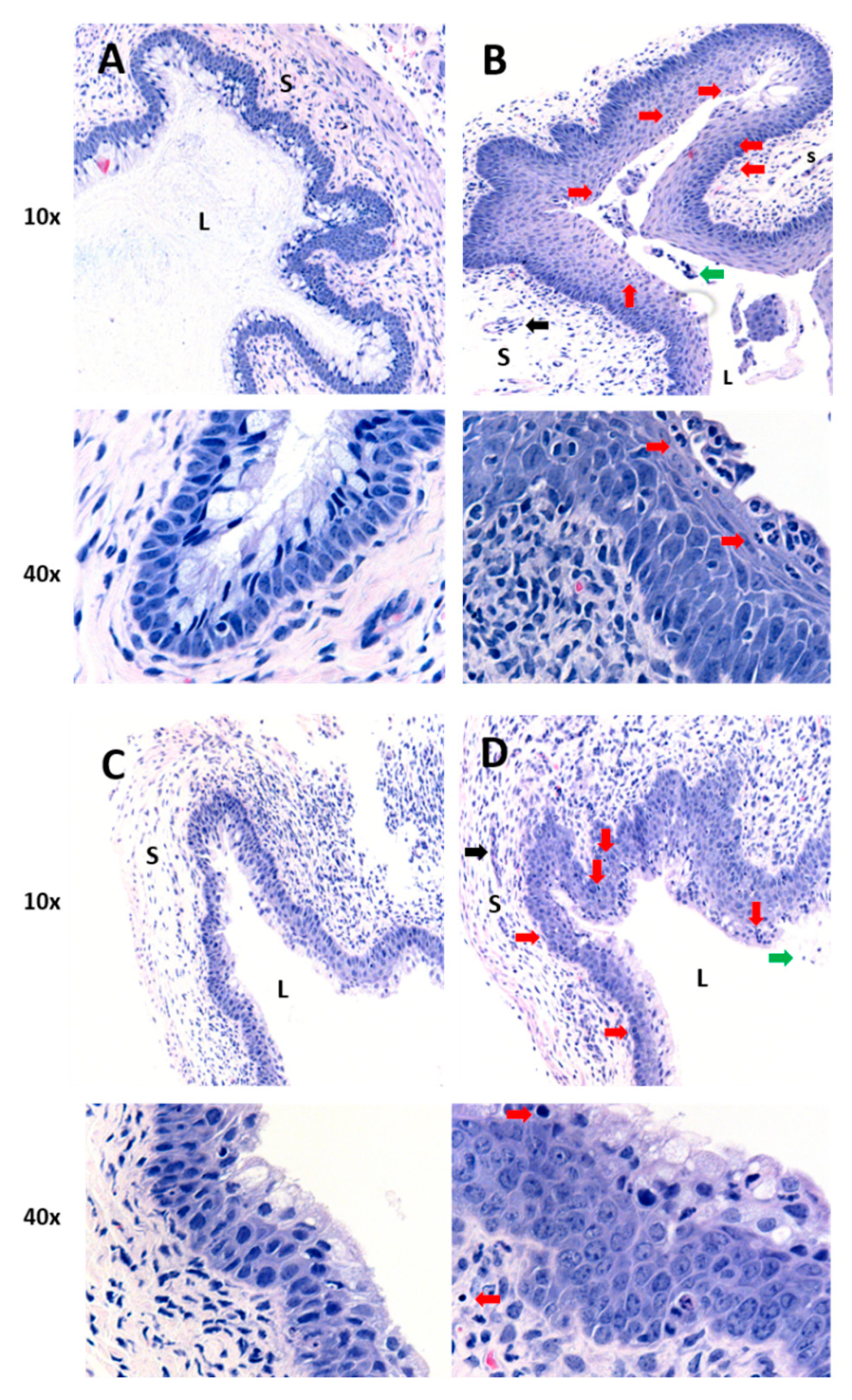
2.1. Oral or IP Administration of Curcumin Nanoparticles Does Not Demonstrate Anti-Inflammatory Activity in the Genital Tract
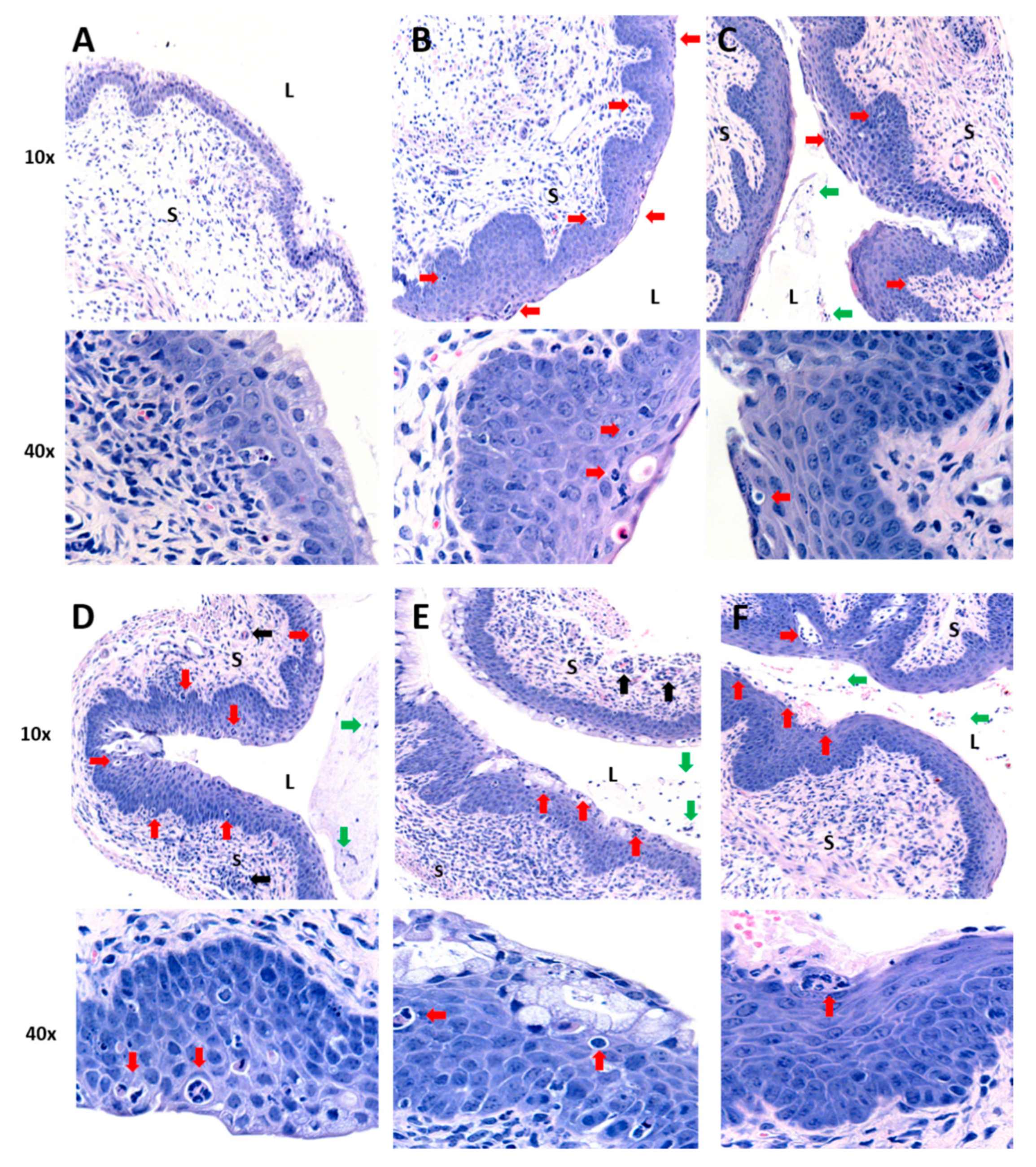
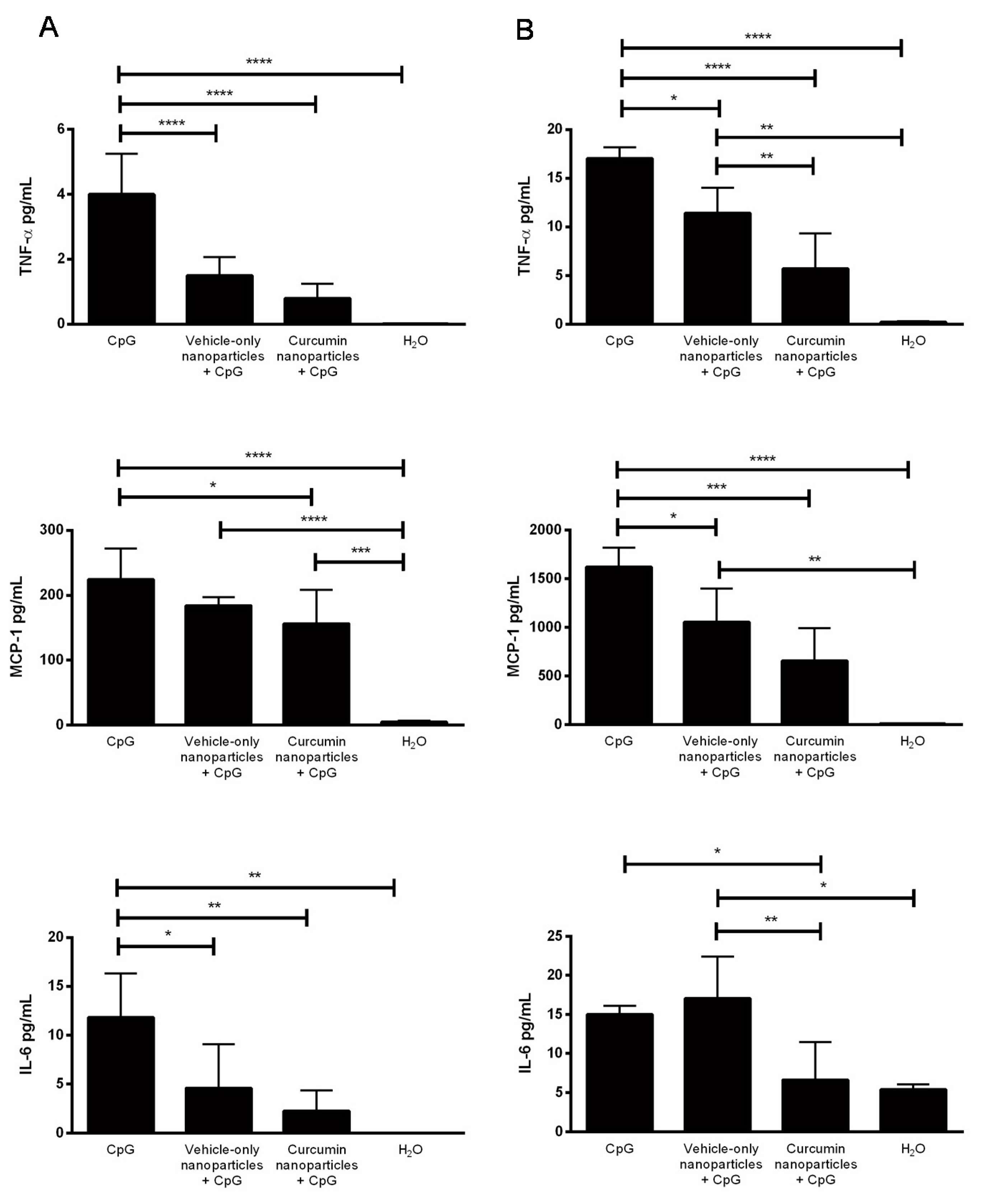
2.2. IVAG Delivery of Curcumin Nanoparticles Reduces Tissue Inflammation and Inflammatory Cytokines in the Genital Tract
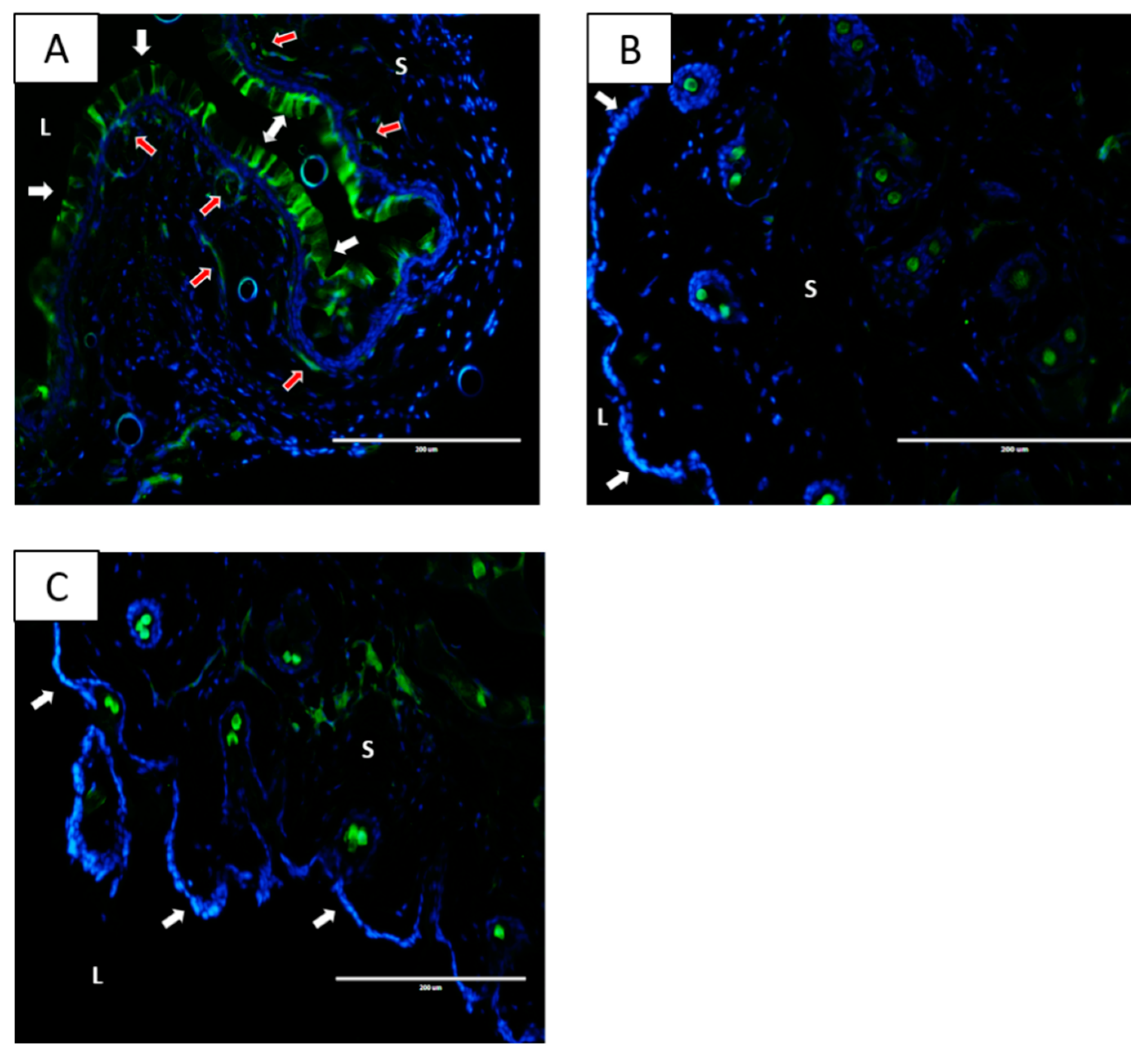
2.3. Nanoparticles Localize within the Vaginal Tissue Following IVAG Administration but Not after IP Delivery
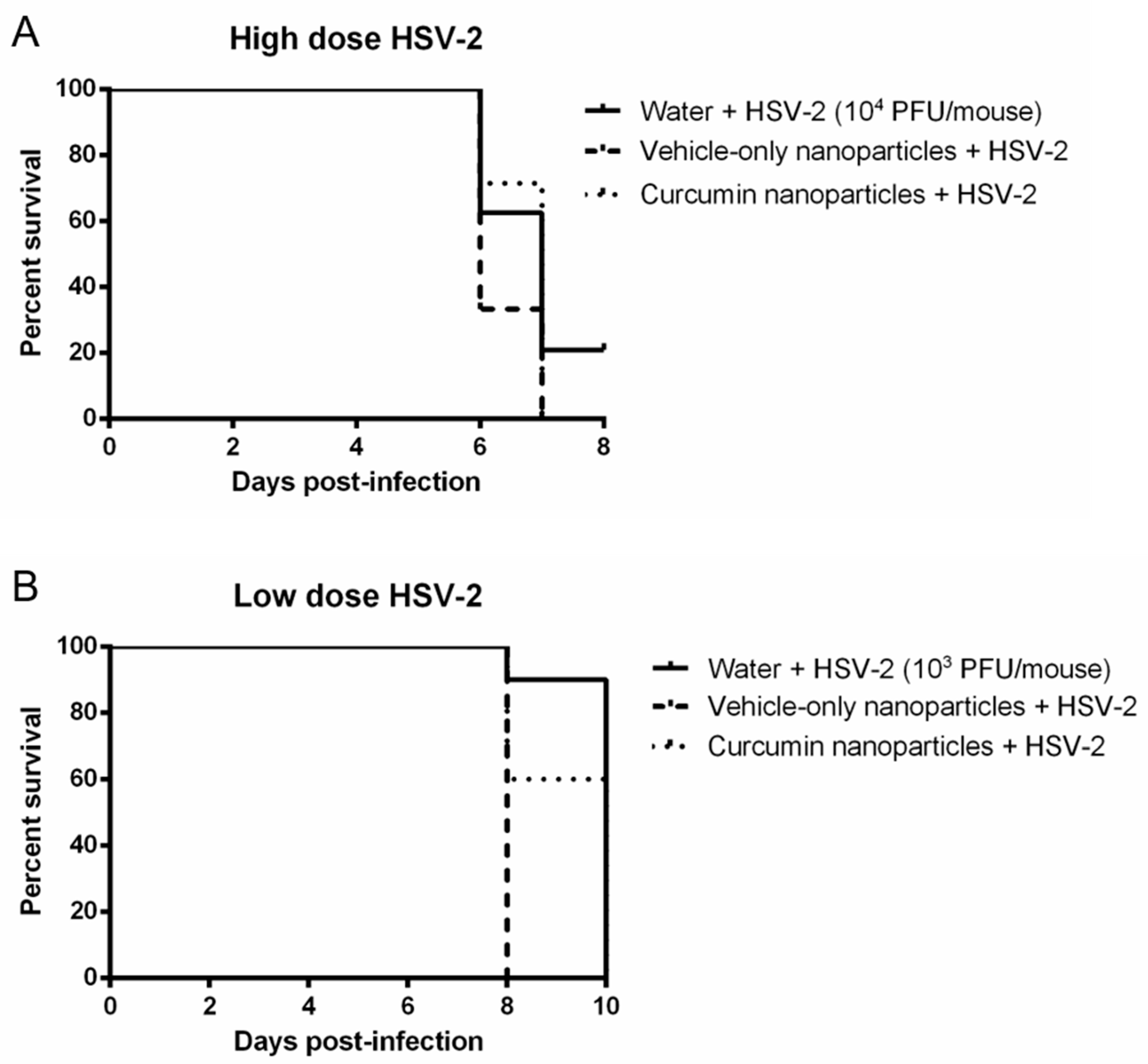
2.4. Curcumin Nanoparticles Do Not Confer Protection against High- or Low-Dose IVAG Challenge with HSV-2
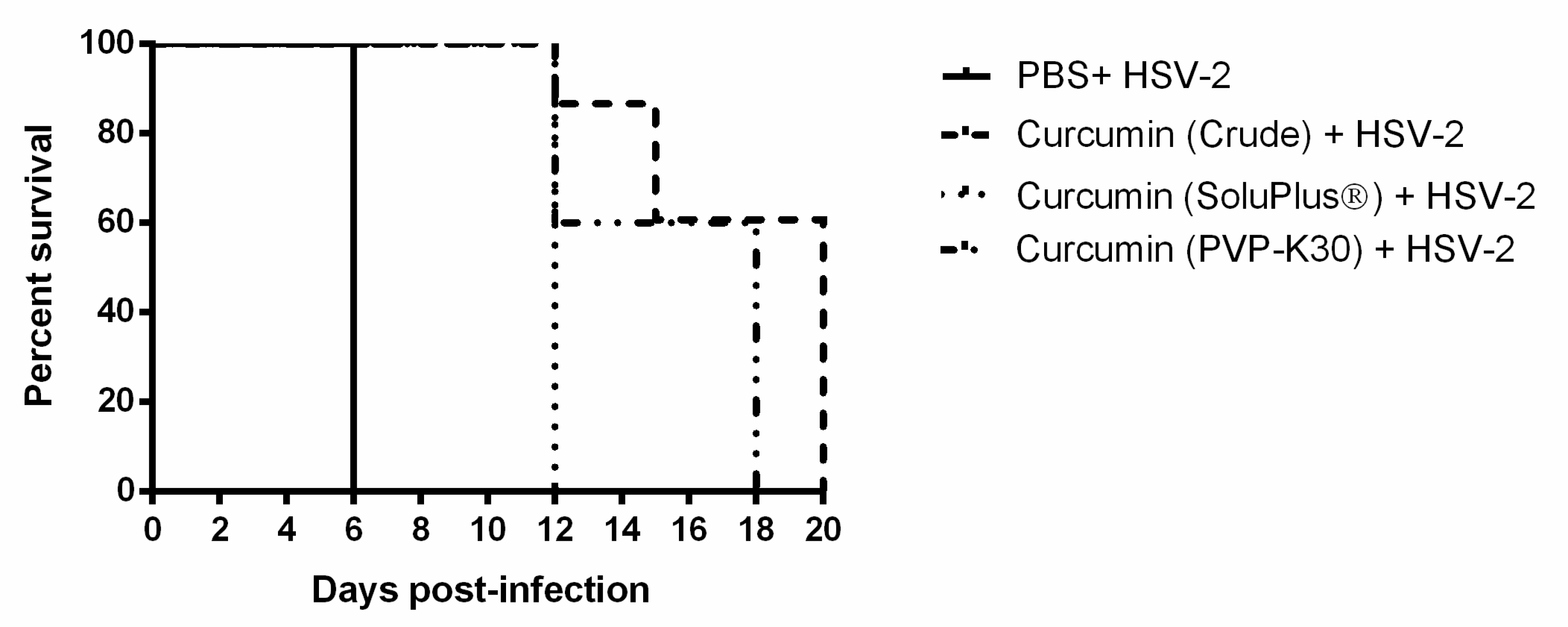
2.5. IVAG Pretreatment with Crude and Solubilized Curcumin Increases Survival Time for HSV-2 Infection and Reduces Pathology
3. Discussion
4. Materials and Methods
4.1. Preparation of Curcumin Nanoparticles
4.2. Mice
4.3. Depo-Provera Injection and Staging
4.4. Histomorphology of the Vaginal Tract Following CpG-ODN Treatment
4.5. Multiplex Cytokine Assay
4.6. HSV-2 Primary Inoculation
4.7. Genital Pathology Assessment during HSV-2 Infection
4.8. Collection of Vaginal Washes
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HSV-2 | Herpes Simplex Virus—Type 2 |
| FGT | Female Genital Tract |
| HIV | Human Immunodeficiency Virus |
| TNF-α | Tumor Necrosis Factor Alpha |
| NFκB | Nuclear Factor Kappa-B |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| IL-6 | Interleukin 6 |
| PLGA | Poly(Lactic-Co-Glycolic Acid) |
| ODN | Oligodeoxynucleotides |
| IVAG | Intravaginal |
| IP | Intraperitoneal |
| H&E | Hematoxylin and Eosin |
| WT | Wild Type |
| PFU | Plaque-Forming Unit |
| PVP-K30 | Polyvinylpyroline K30 |
References
- UNAIDS. Global AIDS Update 2016; UNAIDS: Geneva, Switzerland, 2016. [Google Scholar]
- Wira, C.R.; Fahey, J.V. A new strategy to understand how HIV infects women: Identification of a window of vulnerability during the menstrual cycle. AIDS 2008, 22, 1909–1917. [Google Scholar] [CrossRef]
- Hladik, F.; McElrath, M.J. Setting the stage: Host invasion by HIV. Nat. Rev. Immunol. 2008, 8, 447–457. [Google Scholar] [CrossRef]
- Looker, K.J.; Magaret, A.S.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global estimates of prevalent and incident herpes simplex virus type 2 infections in 2012. PLoS ONE 2015, 10, e114989. [Google Scholar] [CrossRef]
- Masson, L.; Passmore, J.A.; Liebenberg, L.J.; Werner, L.; Baxter, C.; Arnold, K.B.; Williamson, C.; Little, F.; Mansoor, L.E.; Naranbhai, V.; et al. Genital inflammation and the risk of HIV acquisition in women. Clin. Infect. Dis. 2015, 61, 260–269. [Google Scholar] [CrossRef]
- Passmore, J.A.; Jaspan, H.B.; Masson, L. Genital inflammation, immune activation and risk of sexual HIV acquisition. Curr. Opin. HIV AIDS 2016, 11, 156–162. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, M.; Patel, M.V.; Wira, C.R. Innate and adaptive anti-HIV immune responses in the female reproductive tract. J. Reprod. Immunol. 2013, 97, 74–84. [Google Scholar] [CrossRef]
- Ferreira, V.H.; Nazli, A.; Khan, G.; Mian, M.F.; Ashkar, A.A.; Gray-Owen, S.; Kaul, R.; Kaushic, C. Endometrial Epithelial Cell Responses to Coinfecting Viral and Bacterial Pathogens in the Genital Tract Can Activate the HIV-1 LTR in an NFκB-and AP-1–Dependent Manner. J. Infect. Dis. 2011, 204, 299–308. [Google Scholar] [CrossRef]
- Nazli, A.; Chan, O.; Dobson-Belaire, W.N.; Ouellet, M.; Tremblay, M.J.; Gray-Owen, S.D.; Arsenault, A.L.; Kaushic, C. Exposure to HIV-1 directly impairs mucosal epithelial barrier integrity allowing microbial translocation. PLoS Pathog. 2010, 6, e1000852. [Google Scholar] [CrossRef]
- Nazli, A.; Kafka, J.K.; Ferreira, V.H.; Anipindi, V.; Mueller, K.; Osborne, B.J.; Dizzell, S.; Chauvin, S.; Mian, M.F.; Ouellet, M.; et al. HIV-1 gp120 Induces TLR2- and TLR4-Mediated Innate Immune Activation in Human Female Genital Epithelium. J. Immunol. 2013, 191, 4246–4258. [Google Scholar] [CrossRef]
- Martinelli, E.; Tharinger, H.; Frank, I.; Arthos, J.; Piatak, M., Jr.; Lifson, J.D.; Blanchard, J.; Gettie, A.; Robbiani, M. HSV-2 infection of dendritic cells amplifies a highly susceptible HIV-1 cell target. PLoS Pathog. 2011, 7, e1002109. [Google Scholar] [CrossRef]
- Stefanidou, M.; Ramos, I.; Mas Casullo, V.; Trépanier, J.B.; Rosenbaum, S.; Fernandez-Sesma, A.; Herold, B.C. Herpes simplex virus 2 (HSV-2) prevents dendritic cell maturation, induces apoptosis, and triggers release of proinflammatory cytokines: Potential links to HSV-HIV synergy. J. Virol. 2013, 87, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Marino-Merlo, F.; Papaianni, E.; Medici, M.A.; Macchi, B.; Grelli, S.; Mosca, C.; Borner, C.; Mastino, A. HSV-1-induced activation of NF-kappaB protects U937 monocytic cells against both virus replication and apoptosis. Cell Death Dis. 2016, 7, e2354. [Google Scholar] [CrossRef] [PubMed]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Goel, A.; Jhurani, S.; Aggarwal, B.B. Multi-targeted therapy by curcumin: How spicy is it? Mol. Nutr. Food Res. 2008, 52, 1010–1030. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Hashimoto, S.H.U.; Horie, T. Curcumin inhibition of inflammatory cytokine production by peripheral blood monocytes and alveolar maccrophages. Pharmacol. Res. 1999, 39, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, C.; Bright, J.J. Curcumin Inhibits Experimental Allergic Encephalomyelitis by Blocking IL-12 Signaling Through Janus Kinase-STAT Pathway in T Lymphocytes. J. Immunol. 2002, 168, 6506–6513. [Google Scholar] [CrossRef] [PubMed]
- Siwak, D.R.; Shishodia, S.; Aggarwal, B.B.; Kurzrock, R. Curcumin-induced antiproliferative and proapoptotic effects in melanoma cells are associated with suppression of IκB kinase and nuclear factor κB activity and are independent of the B-Raf/mitogen-activated/extracellular signal-regulated protein kinase pathway and the Akt pathway. Cancer 2005, 104, 879–890. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Chun, K.-S.; Cha, H.-H.; Han, S.S.; Keum, Y.-S.; Park, K.-K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Ferreira, V.H.; Nazli, A.; Dizzell, S.E.; Mueller, K.; Kaushic, C. The anti-inflammatory activity of curcumin protects the genital mucosal epithelial barrier from disruption and blocks replication of HIV-1 and HSV-2. PLoS ONE 2015, 10, e0124903. [Google Scholar] [CrossRef]
- Bansal, S.S.; Goel, M.; Aqil, F.; Vadhanam, M.V.; Gupta, R.C. Advanced drug delivery systems of curcumin for cancer chemoprevention. Cancer Prev. Res. (Phila) 2011, 4, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Kharat, M.; Du, Z.; Zhang, G.; McClements, D.J. Physical and Chemical Stability of Curcumin in Aqueous Solutions and Emulsions: Impact of pH, Temperature, and Molecular Environment. J. Agric. Food Chem. 2017, 65, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Gordon, O.N.; Edwards, R.L.; Luis, P.B. Degradation of Curcumin: From Mechanism to Biological Implications. J. Agric. Food Chem. 2015, 63, 7606–7614. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, J.; Ankola, D.D.; Beniwal, V.; Singh, D.; Kumar, M.N.V.R. Nanoparticle encapsulation improves oral bioavailability of curcumin by at least 9-fold when compared to curcumin administered with piperine as absorption enhancer. Eur. J. Pharma. Sci. 2009, 37, 223–230. [Google Scholar] [CrossRef]
- Grama, C.N.; Suryanarayana, P.; Patil, M.A.; Raghu, G.; Balakrishna, N.; Kumar, M.N.V.R.; Reddy, G.B. Efficacy of biodegradable curcumin nanoparticles in delaying cataract in diabetic rat model. PLoS ONE 2013, 8, e78217. [Google Scholar] [CrossRef]
- Devadasu, V.R.; Wadsworth, R.M.; Kumar, M.N. Protective effects of nanoparticulate coenzyme Q10 and curcumin on inflammatory markers and lipid metabolism in streptozotocin-induced diabetic rats: A possible remedy to diabetic complications. Drug Deliv. Transl. Res. 2011, 1, 448–455. [Google Scholar] [CrossRef]
- Parikh, A.; Kathawala, K.; Jintao, L.; Chen, C.; Shan, Z.; Cao, X.; Zhou, X.-F.; Garg, S. Curcumin-loaded self-nanomicellizing solid dispersion system: Part II: In vivo safety and efficacy assessment against behavior deficit in Alzheimer disease. Drug Deliv. Transl. Res. 2018, 8, 1406–1420. [Google Scholar] [CrossRef]
- Parikh, A.; Kathawala, K.; Song, Y.; Zhou, X.-F.; Garg, S. Curcumin-loaded self-nanomicellizing solid dispersion system: Part I: Development, optimization, characterization, and oral bioavailability. Drug Deliv. Transl. Res. 2018, 8, 1389–1405. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Yao, X.-D.; Gill, N.; Sajic, D.; Patrick, A.J.; Rosenthal, K.L. Toll-Like Receptor (TLR)-3, but Not TLR4, Agonist Protects against Genital Herpes Infection in the Absence of Inflammation Seen with CpG DNA. J. Infect. Dis. 2004, 190, 1841–1849. [Google Scholar] [CrossRef]
- Sajic, D.; Ashkar, A.A.; Patrick, A.J.; McCluskie, M.J.; Davis, H.L.; Levine, K.L.; Holl, R.; Rosenthal, K.L. Parameters of CpG oligodeoxynucleotide-induced protection against intravaginal HSV-2 challenge. J. Med. Virol. 2003, 71, 561–568. [Google Scholar] [CrossRef]
- Alcaide, M.L.; Rodriguez, V.J.; Brown, M.R.; Pallikkuth, S.; Arheart, K.; Martinez, O.; Roach, M.; Fichorova, R.N.; Jones, D.L.; Pahwa, S.; et al. High Levels of Inflammatory Cytokines in the Reproductive Tract of Women with BV and Engaging in Intravaginal Douching: A Cross-Sectional Study of Participants in the Women Interagency HIV Study. AIDS Res. Hum. Retrovir. 2017, 33, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.B.; Burgener, A.; Birse, K.; Romas, L.; Dunphy, L.J.; Shahabi, K.; Abou, M.; Westmacott, G.R.; McCorrister, S.; Kwatampora, J.; et al. Increased levels of inflammatory cytokines in the female reproductive tract are associated with altered expression of proteases, mucosal barrier proteins, and an influx of HIV-susceptible target cells. Mucosal Immunol. 2016, 9, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Estes, J.D.; Schlievert, P.M.; Duan, L.; Brosnahan, A.J.; Southern, P.J.; Reilly, C.S.; Peterson, M.L.; Schultz-Darken, N.; Brunner, K.G.; et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature 2009, 458, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, Y.S.; Kumar, P.; Kishore, G.; Bhaskar, C.; Kondapi, A.K. Triple combination MPT vaginal microbicide using curcumin and efavirenz loaded lactoferrin nanoparticles. Sci. Rep. 2016, 6, 25479. [Google Scholar] [CrossRef] [PubMed]
- Bourne, K.Z.; Bourne, N.; Reising, S.F.; Stanberry, L.R. Plant products as topical microbicide candidates: Assessment of in vitro and in vivo activity against herpes simplex virus type 2. Antivir. Res. 1999, 42, 219–226. [Google Scholar] [CrossRef]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, R.; Xie, Q.; Li, A.; Xiao, Y.; Li, K.; Liu, H.; Cui, D.; Chen, Y.; Wang, S. Enhanced bioavailability and efficiency of curcumin for the treatment of asthma by its formulation in solid lipid nanoparticles. Int. J. Nanomed. 2012, 7, 3667–3677. [Google Scholar] [CrossRef]
- Zunino, S.J.; Storms, D.H.; Newman, J.W.; Pedersen, T.L.; Keen, C.L.; Ducore, J.M. Oral or parenteral administration of curcumin does not prevent the growth of high-risk t(4;11) acute lymphoblastic leukemia cells engrafted into a NOD/SCID mouse model. Int. J. Oncol. 2013, 42, 741–748. [Google Scholar] [CrossRef]
- Tamaddonfard, E.; Erfanparast, A.; Hamzeh-Gooshchi, N.; Yousofizadeh, S. Effect of curcumin, the active constituent of turmeric, on penicillin-induced epileptiform activity in rats. Avicenna J. Phytomed 2012, 2, 196–205. [Google Scholar]
- Yu, W.G.; Xu, G.; Ren, G.J.; Xu, X.; Yuan, H.Q.; Qi, X.L.; Tian, K.L. Preventive action of curcumin in experimental acute pancreatitis in mouse. Indian J. Med. Res. 2011, 134, 717–724. [Google Scholar] [CrossRef]
- Young, N.A.; Bruss, M.S.; Gardner, M.; Willis, W.L.; Mo, X.; Valiente, G.R.; Cao, Y.; Liu, Z.; Jarjour, W.N.; Wu, L.C. Oral administration of nano-emulsion curcumin in mice suppresses inflammatory-induced NFkappaB signaling and macrophage migration. PLoS ONE 2014, 9, e111559. [Google Scholar] [CrossRef] [PubMed]
- Ung, V.Y.; Foshaug, R.R.; MacFarlane, S.M.; Churchill, T.A.; Doyle, J.S.; Sydora, B.C.; Fedorak, R.N. Oral administration of curcumin emulsified in carboxymethyl cellulose has a potent anti-inflammatory effect in the IL-10 gene-deficient mouse model of IBD. Dig. Dis. Sci. 2010, 55, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Betsuyaku, T.; Ito, Y.; Nagai, K.; Odajima, N.; Moriyama, C.; Nasuhara, Y.; Nishimura, M. Curcumin attenuates elastase- and cigarette smoke-induced pulmonary emphysema in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L614–L623. [Google Scholar] [CrossRef] [PubMed]
- James, J.S. Curcumin: Clinical trial finds no antiviral effect. AIDS Treat News 1996, 242, 1–2. [Google Scholar]
- Thurman, A.R.; Clark, M.R.; Hurlburt, J.A.; Doncel, G.F. Intravaginal rings as delivery systems for microbicides and multipurpose prevention technologies. Int. J. Womens Health 2013, 5, 695–708. [Google Scholar] [CrossRef]
- Mogal, S.A.; Gurgja, P.N.; Yamgar, D.S.; Kamod, A.C. Solid dispersion technique for improving solubility of some poorly soluble drugs. Der Pharm. Lett. 2012, 4, 1574–1586. [Google Scholar]
- Kaewnopparat, N.; Kaewnopparat, S.; Jangwang, A.; Maneenaun, D.; Chuchome, T.; Panichayupakaranat, P. Increased solubility, dissolution and physicochemical studies ofcurcumin-polyvinylpyrrolidone K-30 solid dispersions. Int. J. Med. Health Biomed. Bioeng. Pharm. Eng. 2009, 3, 137–142. [Google Scholar]
- Kaushic, C.; Ashkar, A.A.; Reid, L.A.; Rosenthal, K.L. Progesterone increases susceptibility and decreases immune responses to genital herpes infection. J. Virol. 2003, 77, 4558–4565. [Google Scholar] [CrossRef]
- Bhavanam, S.; Snider, D.P.; Kaushic, C. Intranasal and subcutaneous immunization under the effect of estradiol leads to better protection against genital HSV-2 challenge compared to progesterone. Vaccine 2008, 26, 6165–6172. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Treatment | Dose d | CpG-ODN | Mouse | Pathology Score | Average Score per Mouse (±S.D.) |
|---|---|---|---|---|---|
| Water (Oral) | 0 | N | 1 | 0 | 0.33 (0.58) **,^ |
| 2 | 1 | ||||
| 3 | 0 | ||||
| Water (Oral) | 0 | Y | 1 | 2 | 1.67 (0.58) * |
| 2 | 1 | ||||
| 3 | 2 | ||||
| Vehicle-Only Nanoparticles (Oral) | 0 | Y | 1 | 1 | 1.17 (0.52) * |
| 2 | 2 | ||||
| 3 | 2 | ||||
| 4 | 1 | ||||
| 5 | 1 | ||||
| 6 | 0 | ||||
| Vehicle-Only Nanoparticles (IP) | 0 | Y | 1 | 1 | 1.20 (0.84) * |
| 2 | 2 | ||||
| 3 | 1 | ||||
| 4 | 2 | ||||
| 5 | 0 | ||||
| Curcumin Nanoparticles (Oral) | 1.25 | Y | 1 | 1 | 1.67 (0.52) * |
| 2 | 1 | ||||
| 3 | 2 | ||||
| 4 | 2 | ||||
| 5 | 2 | ||||
| 6 | 2 | ||||
| Curcumin Nanoparticles (IP) | 1.25 | Y | 1 | 0 | 0.80 (1.1) * |
| 2 | 2 | ||||
| 3 | 0 | ||||
| 4 | 2 | ||||
| 5 | 0 | ||||
| Curcumin Nanoparticles (IP) | 2.4 | Y | 1 | 2 | 1.16 (0.75) * |
| 2 | 1 | ||||
| 3 | 2 | ||||
| 4 | 1 | ||||
| 5 | 1 | ||||
| 6 | 0 | ||||
| Curcumin Nanoparticles (IP) | 4.8 | Y | 1 | 0.5 | 1.50 (0.87) * |
| 2 | 2 | ||||
| 3 | 2 |
| Pre-Treatment | CpG-ODN | Mouse | Pathology Score | Average Score per Mouse (± S.D.) |
|---|---|---|---|---|
| Water | N | 1 | 0 | 0.33 (0.58) **,^ |
| 2 | 0 | |||
| 3 | 1 | |||
| Water | Y | 1 | 3 | 2.67 (0.58) * |
| 2 | 3 | |||
| 3 | 2 | |||
| Vehicle-Only Nanoparticles | Y | 1 | 1 | 1.67 (0.58) * |
| 2 | 2 | |||
| 3 | 2 | |||
| Curcumin Nanoparticles | Y | 1 | 0 | 0.50 (0.58) **,^ |
| 2 | 1 | |||
| 3 | 0 | |||
| 4 | 1 |
| Pre-Treatment | HSV-2 Infection Dosage | Number of Mice | Days with No Pathology | Cumulative Pathology Score | Average Score per Mouse |
|---|---|---|---|---|---|
| Water | high | 4 | 1–3 | 75 | 18.8 |
| Vehicle-only Nanoparticles | high | 5 | 1–2 | 98 | 19.6 |
| Curcumin Nanoparticles | high | 6 | 1–3 | 95 | 15.8 |
| Vehicle-only Nanoparticles | low | 5 | 4–6 | 53 | 10.6 |
| Curcumin Nanoparticles | low | 5 | 4–6 | 53 | 10.6 |
| Pre-Treatment | Number of Mice | Mice with Pathology Score ≥3 by Day 6 | Cumulative Pathology Score | Average Score per Mouse |
|---|---|---|---|---|
| PBS | 5 | 5 | 182 | 36.4 |
| Crude Curcumin | 5 | 3 | 112 | 22.2 |
| Curcumin in PVP-K30 | 5 | 0 | 60 | 12.0 |
| Curcumin in Soluplus® | 5 | 0 | 59 | 11.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitali, D.; Bagri, P.; Wessels, J.M.; Arora, M.; Ganugula, R.; Parikh, A.; Mandur, T.; Felker, A.; Garg, S.; Kumar, M.N.V.R.; et al. Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa. Int. J. Mol. Sci. 2020, 21, 337. https://doi.org/10.3390/ijms21010337
Vitali D, Bagri P, Wessels JM, Arora M, Ganugula R, Parikh A, Mandur T, Felker A, Garg S, Kumar MNVR, et al. Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa. International Journal of Molecular Sciences. 2020; 21(1):337. https://doi.org/10.3390/ijms21010337
Chicago/Turabian StyleVitali, Danielle, Puja Bagri, Jocelyn M. Wessels, Meenakshi Arora, Raghu Ganugula, Ankit Parikh, Talveer Mandur, Allison Felker, Sanjay Garg, M.N.V. Ravi Kumar, and et al. 2020. "Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa" International Journal of Molecular Sciences 21, no. 1: 337. https://doi.org/10.3390/ijms21010337
APA StyleVitali, D., Bagri, P., Wessels, J. M., Arora, M., Ganugula, R., Parikh, A., Mandur, T., Felker, A., Garg, S., Kumar, M. N. V. R., & Kaushic, C. (2020). Curcumin Can Decrease Tissue Inflammation and the Severity of HSV-2 Infection in the Female Reproductive Mucosa. International Journal of Molecular Sciences, 21(1), 337. https://doi.org/10.3390/ijms21010337