Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders


Abstract
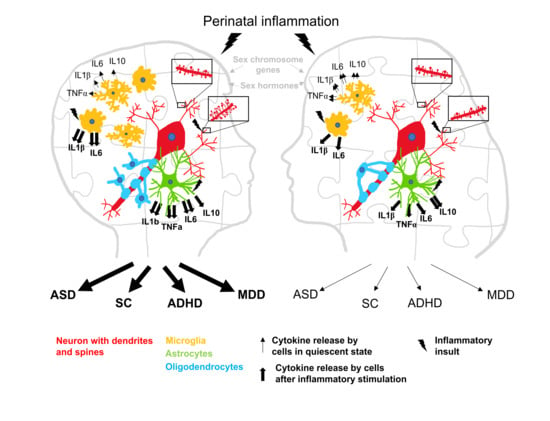
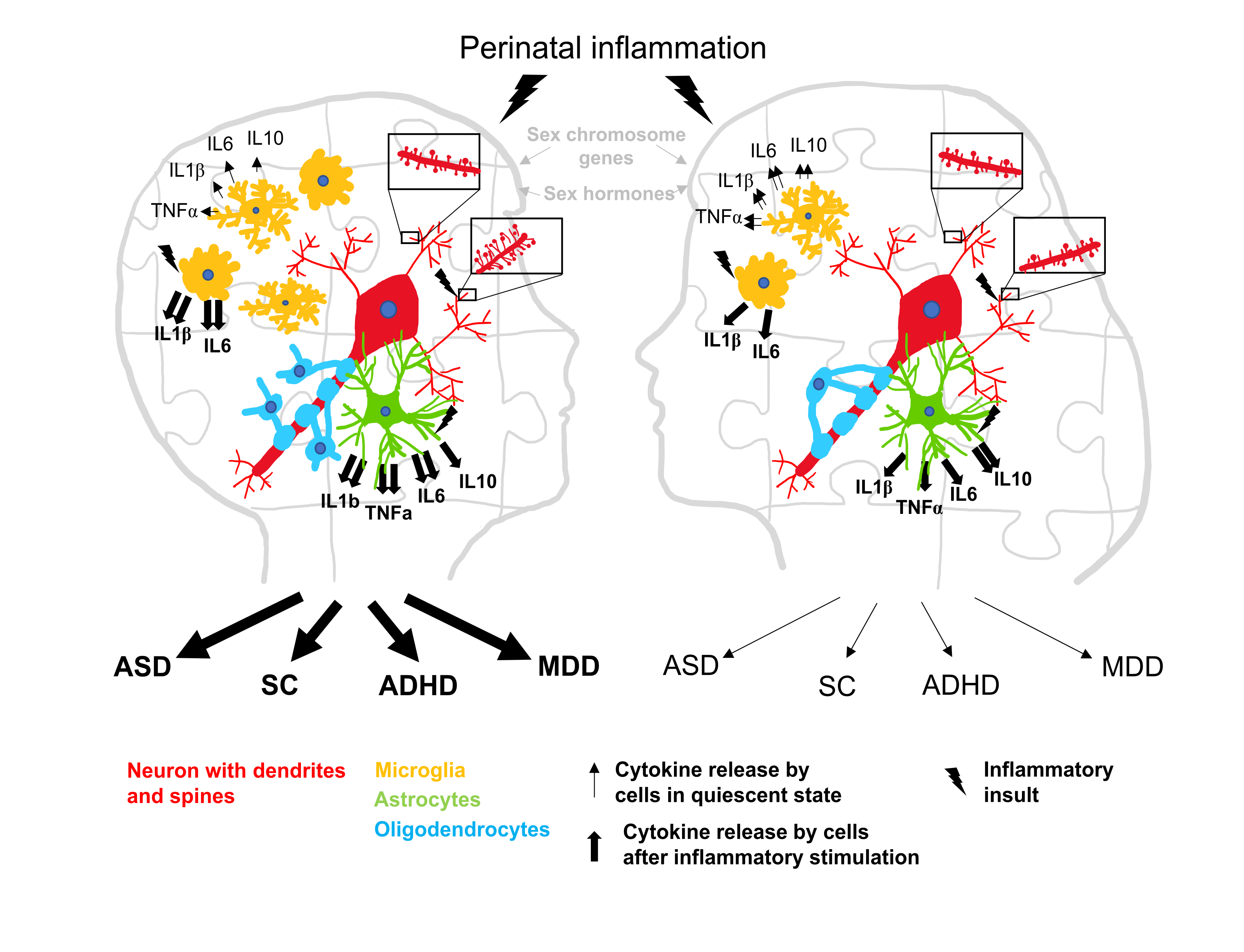
1. Introduction
2. Sex-Dependent Effects on Physiological Brain Development
3. Effects of Sex on Glial Cell Development
3.1. Microglia
3.2. Astrocytes
3.3. Oligodendrocytes
4. Neuropsychiatric Disorders: Clinical Evidence of a Role for Infection/Inflammation
4.1. Autism Spectrum Disorder (ASD)
4.2. Attention-Deficit/Hyperactivity Disorder (ADHD)
4.3. Schizophrenia
4.4. Depression
5. Animal Models of Perinatal Inflammation: Behavioral Abnormalities Related to Neurodevelopmental Disorders
6. Pre-Clinical Evidence of Pathogenesis in Neurodevelopmental Disorders
7. Sex-Dependent Effects of Perinatal Inflammation on Glial Cells and Brain Inflammation
7.1. Microglia Activation
7.2. Astrocyte Activation
7.3. Effects on Oligodendrocytes
7.4. Inflammatory Responses
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| + | Positive |
| ADHD | Attention deficit/hyperactivity disorder |
| ADNP | Activity-dependent neuroprotective protein |
| AMPA | Α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| AS | Asperger’s syndrome |
| ASD | Autistic spectrum disorder |
| Atg | Autophagy related |
| BDNF | Brain-derived neurotrophic factor |
| BTBR | BTBR T+ tf/J mouse strain |
| CA | Cornu ammonis |
| CNS | Central nervous system |
| Cntnap2 | Contactin-associated protein-like 2 |
| COX | Cyclooxygenase enzyme |
| CRP | C-reactive protein |
| Cx3cr1 | C-x3-c motif chemokine receptor 1 |
| E | Embryonic day |
| E. coli. | Escherichia coli |
| GABA | γ-aminobutyric acid |
| GAD | Glutamic acid decarboxylase |
| GBS | Group B streptococcus |
| GFAP | Glial fibrillary acidic protein |
| Iba-1 | Ionized calcium binding adaptor molecule 1 |
| IDO | Indoleamine 2,3-dioxygenase |
| IFN | Interferon |
| IgG | Immunoglobulin G |
| IL | Interleukin |
| IP | Interferon gamma-induced protein |
| LPS | Lipopolysaccharide |
| MAFB | V-maf avian musculoaponeurotic fibrosarcoma oncogene homolog B |
| MDD | Major depressive disorder |
| MECP2 | Methyl CpG binding protein 2 |
| mhDISC1 | Mutant human disrupted-in-schizophrenia 1 |
| miRNA | Micro ribonucleic acid |
| NG2 | Neural/glial antigen 2 |
| NSAID | Nonsteroidal anti-inflammatory drug |
| O4 | Oligodendrocyte marker |
| OPCs | Oligodendrocyte precursor cells |
| P | Postnatal day |
| PARP-1 | Poly [ADP-ribose] polymerase 1 |
| PDGF | Platelet-derived growth factor |
| PG | Prostaglandin |
| POA | Preoptic area |
| Poly:IC | Polyinosinic-polycytidylic acid |
| R | Receptor |
| SC | Schizophrenia |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
| TSPO | Translocator protein |
| TX | Thromboxane |
| US | United States |
References
- Hagberg, H.; David Edwards, A.; Groenendaal, F. Perinatal brain damage: The term infant. Neurobiol. Dis. 2016, 92, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Strunk, T.; Inder, T.; Wang, X.; Burgner, D.; Mallard, C.; Levy, O. Infection-induced inflammation and cerebral injury in preterm infants. Lancet Infect. Dis. 2014, 14, 751–762. [Google Scholar] [CrossRef]
- Bale, T.L. Neuroendocrine and immune influences on the CNS: it’s a matter of sex. Neuron 2009, 64, 13–16. [Google Scholar] [CrossRef]
- Careaga, M.; Murai, T.; Bauman, M.D. Maternal Immune Activation and Autism Spectrum Disorder: From Rodents to Nonhuman and Human Primates. Biol. Psychiatry 2017, 81, 391–401. [Google Scholar] [CrossRef]
- Knuesel, I.; Chicha, L.; Britschgi, M.; Schobel, S.A.; Bodmer, M.; Hellings, J.A.; Toovey, S.; Prinssen, E.P. Maternal immune activation and abnormal brain development across CNS disorders. Nat. Rev. Neurol. 2014, 10, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.C.; Tanskanen, A.; Huttunen, M.; Whittaker, J.C.; Cannon, M. Evidence for an interaction between familial liability and prenatal exposure to infection in the causation of schizophrenia. Am. J. Psychiatry 2009, 166, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Banik, A.; Kandilya, D.; Ramya, S.; Stunkel, W.; Chong, Y.S.; Dheen, S.T. Maternal factors that induce epigenetic changes contribute to neurological disorders in offspring. Genes 2017, 8, 150. [Google Scholar] [CrossRef]
- Basil, P.; Li, Q.; Dempster, E.L.; Mill, J.; Sham, P.C.; Wong, C.C.; McAlonan, G.M. Prenatal maternal immune activation causes epigenetic differences in adolescent mouse brain. Transl. Psychiatry 2014, 4, e434. [Google Scholar] [CrossRef]
- Kundakovic, M.; Jaric, I. The epigenetic link between prenatal adverse environments and neurodevelopmental disorders. Genes (Basel) 2017, 8, 104. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Lombardo, M.V.; Auyeung, B.; Ashwin, E.; Chakrabarti, B.; Knickmeyer, R. Why are autism spectrum conditions more prevalent in males? PLoS Biol. 2011, 9, e1001081. [Google Scholar] [CrossRef] [PubMed]
- Rutter, M.; Caspi, A.; Moffitt, T.E. Using sex differences in psychopathology to study causal mechanisms: Unifying issues and research strategies. J. Child. Psychol. Psychiatry 2003, 44, 1092–1115. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Kang, H.J.; Kawasawa, Y.I.; Cheng, F.; Zhu, Y.; Xu, X.; Li, M.; Sousa, A.M.; Pletikos, M.; Meyer, K.A.; Sedmak, G.; et al. Spatio-temporal transcriptome of the human brain. Nature 2011, 478, 483–489. [Google Scholar] [CrossRef]
- Arnold, A.P.; Breedlove, S.M. Organizational and activational effects of sex steroids on brain and behavior: A reanalysis. Horm. Behav. 1985, 19, 469–498. [Google Scholar] [CrossRef]
- Arnold, A.P.; Chen, X. What does the “four core genotypes” mouse model tell us about sex differences in the brain and other tissues? Front. Neuroendocrinol. 2009, 30, 1–9. [Google Scholar] [CrossRef]
- Lenz, K.M.; Nugent, B.M.; Haliyur, R.; McCarthy, M.M. Microglia are essential to masculinization of brain and behavior. J. Neurosci. 2013, 33, 2761–2772. [Google Scholar] [CrossRef]
- McCarthy, M.M. Multifaceted origins of sex differences in the brain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150106. [Google Scholar] [CrossRef]
- Stiles, J.; Jernigan, T.L. The basics of brain development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef]
- Brooks, C.E.; Clayton, J.A. Sex/gender influences on the nervous system: Basic steps toward clinical progress. J. Neurosci. Res. 2017, 95, 14–16. [Google Scholar] [CrossRef]
- Ruigrok, A.N.; Salimi-Khorshidi, G.; Lai, M.C.; Baron-Cohen, S.; Lombardo, M.V.; Tait, R.J.; Suckling, J. A meta-analysis of sex differences in human brain structure. Neurosci. Biobehav. Rev. 2014, 39, 34–50. [Google Scholar] [CrossRef]
- Konkle, A.T.; McCarthy, M.M. Developmental time course of estradiol, testosterone, and dihydrotestosterone levels in discrete regions of male and female rat brain. Endocrinology 2011, 152, 223–235. [Google Scholar] [CrossRef]
- McCarthy, M.M. Estradiol and the developing brain. Physiol. Rev. 2008, 88, 91–124. [Google Scholar] [CrossRef]
- Ritchie, S.J.; Cox, S.R.; Shen, X.; Lombardo, M.V.; Reus, L.M.; Alloza, C.; Harris, M.A.; Alderson, H.L.; Hunter, S.; Neilson, E.; et al. Sex Differences in the Adult Human Brain: Evidence from 5216 UK Biobank Participants. Cereb. Cortex 2018, 28, 2959–2975. [Google Scholar] [CrossRef]
- Caviness, V.S., Jr.; Kennedy, D.N.; Richelme, C.; Rademacher, J.; Filipek, P.A. The human brain age 7–11 years: A volumetric analysis based on magnetic resonance images. Cereb. Cortex 1996, 6, 726–736. [Google Scholar] [CrossRef]
- Lim, S.; Han, C.E.; Uhlhaas, P.J.; Kaiser, M. Preferential detachment during human brain development: Age- and sex-specific structural connectivity in diffusion tensor imaging (DTI) data. Cereb. Cortex 2015, 25, 1477–1489. [Google Scholar] [CrossRef]
- Uematsu, A.; Matsui, M.; Tanaka, C.; Takahashi, T.; Noguchi, K.; Suzuki, M.; Nishijo, H. Developmental trajectories of amygdala and hippocampus from infancy to early adulthood in healthy individuals. PLoS ONE 2012, 7, e46970. [Google Scholar] [CrossRef]
- Greimel, E.; Nehrkorn, B.; Schulte-Ruther, M.; Fink, G.R.; Nickl-Jockschat, T.; Herpertz-Dahlmann, B.; Konrad, K.; Eickhoff, S.B. Changes in grey matter development in autism spectrum disorder. Brain Struct. Funct. 2013, 218, 929–942. [Google Scholar] [CrossRef]
- Sowell, E.R.; Trauner, D.A.; Gamst, A.; Jernigan, T.L. Development of cortical and subcortical brain structures in childhood and adolescence: A structural MRI study. Dev. Med. Child. Neurol. 2002, 44, 4–16. [Google Scholar] [CrossRef]
- Koolschijn, P.C.; Crone, E.A. Sex differences and structural brain maturation from childhood to early adulthood. Dev. Cogn. Neurosci. 2013, 5, 106–118. [Google Scholar] [CrossRef]
- Qin, X.Y.; Feng, J.C.; Cao, C.; Wu, H.T.; Loh, Y.P.; Cheng, Y. Association of Peripheral Blood Levels of Brain-Derived Neurotrophic Factor With Autism Spectrum Disorder in Children: A Systematic Review and Meta-analysis. JAMA Pediatr. 2016, 170, 1079–1086. [Google Scholar] [CrossRef]
- Hagberg, H. Mitochondrial impairment in the developing brain after hypoxia-ischemia. J. Bioenerg. Biomembr. 2004, 36, 369–373. [Google Scholar] [CrossRef]
- Amateau, S.K.; McCarthy, M.M. A novel mechanism of dendritic spine plasticity involving estradiol induction of prostaglandin-E2. J. Neurosci. 2002, 22, 8586–8596. [Google Scholar] [CrossRef]
- Wright, C.L.; McCarthy, M.M. Prostaglandin E2-induced masculinization of brain and behavior requires protein kinase A, AMPA/kainate, and metabotropic glutamate receptor signaling. J. Neurosci. 2009, 29, 13274–13282. [Google Scholar] [CrossRef]
- Lenz, K.M.; Wright, C.L.; Martin, R.C.; McCarthy, M.M. Prostaglandin E(2) regulates AMPA receptor phosphorylation and promotes membrane insertion in preoptic area neurons and glia during sexual differentiation. PLoS ONE 2011, 6, e18500. [Google Scholar] [CrossRef]
- Amateau, S.K.; McCarthy, M.M. Induction of PGE2 by estradiol mediates developmental masculinization of sex behavior. Nat. Neurosci. 2004, 7, 643–650. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Pickett, L.A.; VanRyzin, J.W.; Kight, K.E. Surprising origins of sex differences in the brain. Horm. Behav. 2015, 76, 3–10. [Google Scholar] [CrossRef]
- Amateau, S.K.; Alt, J.J.; Stamps, C.L.; McCarthy, M.M. Brain estradiol content in newborn rats: Sex differences, regional heterogeneity, and possible de novo synthesis by the female telencephalon. Endocrinology 2004, 145, 2906–2917. [Google Scholar] [CrossRef]
- Hammond, T.R.; Robinton, D.; Stevens, B. Microglia and the Brain: Complementary Partners in Development and Disease. Annu Rev. Cell Dev. Biol. 2018, 34, 523–544. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic pruning by microglia is necessary for normal brain development. Science 2011, 333, 1456–1458. [Google Scholar] [CrossRef]
- Yirmiya, R.; Rimmerman, N.; Reshef, R. Depression as a microglial disease. Trends Neurosci. 2015, 38, 637–658. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Gonzalez-Rueda, A.; Sampaio-Baptista, C.; Paulsen, O.; Rodriguez-Moreno, A. Distinct mechanisms of spike timing-dependent LTD at vertical and horizontal inputs onto L2/3 pyramidal neurons in mouse barrel cortex. Physiol. Rep. 2014, 2, e00271. [Google Scholar] [CrossRef] [PubMed]
- Sloan, S.A.; Barres, B.A. Mechanisms of astrocyte development and their contributions to neurodevelopmental disorders. Curr. Opin. Neurobiol. 2014, 27, 75–81. [Google Scholar] [CrossRef]
- Koyama, R.; Ikegaya, Y. Microglia in the pathogenesis of autism spectrum disorders. Neurosci. Res. 2015, 100, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Schafer, D.P.; Lehrman, E.K.; Stevens, B. The “quad-partite” synapse: Microglia-synapse interactions in the developing and mature CNS. Glia 2013, 61, 24–36. [Google Scholar] [CrossRef]
- Lenz, K.M.; Nelson, L.H. Microglia and Beyond: Innate Immune Cells As Regulators of Brain Development and Behavioral Function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Makinson, R.; Lloyd, K.; Rayasam, A.; McKee, S.; Brown, A.; Barila, G.; Grissom, N.; George, R.; Marini, M.; Fabry, Z.; et al. Intrauterine inflammation induces sex-specific effects on neuroinflammation, white matter, and behavior. Brain Behav. Immun. 2017, 66, 277–288. [Google Scholar] [CrossRef]
- Thion, M.S.; Low, D.; Silvin, A.; Chen, J.M.; Grisel, P.; Schulte-Schrepping, J.; Blecher, R.; Ulas, T.; Squarzoni, P.; Hoeffel, G.; et al. Microbiome Influences Prenatal and Adult Microglia in a Sex-Specific Manner. Cell 2018, 172, 500–516. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Sholar, P.W.; Bilbo, S.D. Sex differences in microglial colonization of the developing rat brain. J. Neurochem. 2012, 120, 948–963. [Google Scholar] [CrossRef]
- Hanamsagar, R.; Alter, M.D.; Block, C.S.; Sullivan, H.; Bolton, J.L.; Bilbo, S.D. Generation of a microglial developmental index in mice and in humans reveals a sex difference in maturation and immune reactivity. Glia 2018, 66, 460. [Google Scholar] [CrossRef]
- Ruggiero, M.J.; Boschen, K.E.; Roth, T.L.; Klintsova, A.Y. Sex Differences in Early Postnatal Microglial Colonization of the Developing Rat Hippocampus Following a Single-Day Alcohol Exposure. J. Neuroimmune Pharmacol. 2018, 13, 189–203. [Google Scholar] [CrossRef]
- Nelson, L.H.; Warden, S.; Lenz, K.M. Sex differences in microglial phagocytosis in the neonatal hippocampus. Brain Behav. Immun. 2017, 64, 11–22. [Google Scholar] [CrossRef]
- Schafer, D.P.; Stevens, B. Phagocytic glial cells: Sculpting synaptic circuits in the developing nervous system. Curr. Opin. Neurobiol. 2013, 23, 1034–1040. [Google Scholar] [CrossRef]
- Turano, A.; Lawrence, J.H.; Schwarz, J.M. Activation of neonatal microglia can be influenced by other neural cells. Neurosci. Lett. 2017, 657, 32–37. [Google Scholar] [CrossRef]
- Crain, J.M.; Nikodemova, M.; Watters, J.J. Microglia express distinct M1 and M2 phenotypic markers in the postnatal and adult central nervous system in male and female mice. J. Neurosci. Res. 2013, 91, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Osborne, B.F.; Turano, A.; Caulfield, J.I.; Schwarz, J.M. Sex- and region-specific differences in microglia phenotype and characterization of the peripheral immune response following early-life infection in neonatal male and female rats. Neurosci. Lett. 2018, 692, 1–9. [Google Scholar] [CrossRef]
- Ohsawa, K.; Imai, Y.; Kanazawa, H.; Sasaki, Y.; Kohsaka, S. Involvement of Iba1 in membrane ruffling and phagocytosis of macrophages/microglia. J. Cell Sci. 2000, 113, 3073–3084. [Google Scholar]
- Hagberg, H.; Wilson, M.A.; Matsushita, H.; Zhu, C.; Lange, M.; Gustavsson, M.; Poitras, M.F.; Dawson, T.M.; Dawson, V.L.; Northington, F.; et al. PARP-1 gene disruption in mice preferentially protects males from perinatal brain injury. J. Neurochem. 2004, 90, 1068–1075. [Google Scholar] [CrossRef]
- Zhu, C.; Xu, F.; Wang, X.; Shibata, M.; Uchiyama, Y.; Blomgren, K.; Hagberg, H. Different apoptotic mechanisms are activated in male and female brains after neonatal hypoxia-ischaemia. J. Neurochem. 2006, 96, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pouchoulen, M.; VanRyzin, J.W.; McCarthy, M.M. Morphological and Phagocytic Profile of Microglia in the Developing Rat Cerebellum. eNeuro 2015, 2, ENEURO-0036. [Google Scholar] [CrossRef] [PubMed]
- Prilutsky, D.; Kho, A.T.; Feiglin, A.; Hammond, T.; Stevens, B.; Kohane, I.S. Sexual dimorphism of complement-dependent microglial synaptic pruning and other immune pathways in the developing brain. bioRxiv 2017, 204412. [Google Scholar]
- Osborne, B.F.; Turano, A.; Schwarz, J.M. Sex Differences in the Neuroimmune System. Curr. Opin. Behav. Sci. 2018, 23, 118–123. [Google Scholar] [CrossRef]
- Pekny, M.; Pekna, M. Astrocyte reactivity and reactive astrogliosis: Costs and benefits. Physiol. Rev. 2014, 94, 1077–1098. [Google Scholar] [CrossRef]
- Reemst, K.; Noctor, S.C.; Lucassen, P.J.; Hol, E.M. The Indispensable Roles of Microglia and Astrocytes during Brain Development. Front. Hum. Neurosci. 2016, 10, 566. [Google Scholar] [CrossRef]
- Ge, W.P.; Miyawaki, A.; Gage, F.H.; Jan, Y.N.; Jan, L.Y. Local generation of glia is a major astrocyte source in postnatal cortex. Nature 2012, 484, 376–380. [Google Scholar] [CrossRef]
- Andriezen, W.L. The Neuroglia Elements in the Human Brain. Br. Med. J. 1893, 2, 227–230. [Google Scholar] [CrossRef]
- Miller, R.H.; Raff, M.C. Fibrous and protoplasmic astrocytes are biochemically and developmentally distinct. J. Neurosci. 1984, 4, 585–592. [Google Scholar] [CrossRef]
- Arnold, A.P. The organizational-activational hypothesis as the foundation for a unified theory of sexual differentiation of all mammalian tissues. Horm. Behav. 2009, 55, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Segura, L.M.; Duenas, M.; Busiguina, S.; Naftolin, F.; Chowen, J.A. Gonadal hormone regulation of neuronal-glial interactions in the developing neuroendocrine hypothalamus. J. Steroid Biochem. Mol. Biol. 1995, 53, 293–298. [Google Scholar] [CrossRef]
- Mong, J.A.; Kurzweil, R.L.; Davis, A.M.; Rocca, M.S.; McCarthy, M.M. Evidence for sexual differentiation of glia in rat brain. Horm. Behav. 1996, 30, 553–562. [Google Scholar] [CrossRef]
- Piechota, M.; Korostynski, M.; Golda, S.; Ficek, J.; Jantas, D.; Barbara, Z.; Przewlocki, R. Transcriptional signatures of steroid hormones in the striatal neurons and astrocytes. BMC Neurosci. 2017, 18, 37. [Google Scholar] [CrossRef]
- Chowen, J.A.; Azcoitia, I.; Cardona-Gomez, G.P.; Garcia-Segura, L.M. Sex steroids and the brain: Lessons from animal studies. J. Pediatr. Endocrinol. Metab. 2000, 13, 1045–1066. [Google Scholar] [CrossRef]
- Mong, J.A.; McCarthy, M.M. Ontogeny of sexually dimorphic astrocytes in the neonatal rat arcuate. Brain Res. Dev. Brain Res. 2002, 139, 151–158. [Google Scholar] [CrossRef]
- Catalani, A.; Sabbatini, M.; Consoli, C.; Cinque, C.; Tomassoni, D.; Azmitia, E.; Angelucci, L.; Amenta, F. Glial fibrillary acidic protein immunoreactive astrocytes in developing rat hippocampus. Mech. Ageing Dev. 2002, 123, 481–490. [Google Scholar] [CrossRef]
- Suarez, I.; Bodega, G.; Rubio, M.; Fernandez, B. Sexual dimorphism in the hamster cerebellum demonstrated by glial fibrillary acidic protein (GFAP) and vimentin immunoreactivity. Glia 1992, 5, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Krebs-Kraft, D.L.; Hill, M.N.; Hillard, C.J.; McCarthy, M.M. Sex difference in cell proliferation in developing rat amygdala mediated by endocannabinoids has implications for social behavior. Proc. Natl. Acad. Sci. USA 2010, 107, 20535–20540. [Google Scholar] [CrossRef]
- Brown, G.C.; Neher, J.J. Microglial phagocytosis of live neurons. Nat. Rev. Neurosci. 2014, 15, 209–216. [Google Scholar] [CrossRef]
- Santos-Galindo, M.; Acaz-Fonseca, E.; Bellini, M.J.; Garcia-Segura, L.M. Sex differences in the inflammatory response of primary astrocytes to lipopolysaccharide. Biol. Sex Differ. 2011, 2, 7. [Google Scholar] [CrossRef]
- Chistyakov, D.V.; Azbukina, N.V.; Astakhova, A.A.; Goriainov, S.V.; Chistyakov, V.V.; Sergeeva, M.G. Sex-Mediated Differences in LPS Induced Alterations of TNFalpha, IL-10 Expression, and Prostaglandin Synthesis in Primary Astrocytes. Int. J. Mol. Sci. 2018, 19, 2793. [Google Scholar] [CrossRef]
- Elbaz, B.; Popko, B. Molecular Control of Oligodendrocyte Development. Trends Neurosci. 2019, 42, 263–277. [Google Scholar] [CrossRef]
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Volpe, J.J.; Kinney, H.C. Arrested oligodendrocyte lineage progression during human cerebral white matter development: Dissociation between the timing of progenitor differentiation and myelinogenesis. J. Neuropathol. Exp. Neurol. 2002, 61, 197–211. [Google Scholar] [CrossRef]
- Cerghet, M.; Skoff, R.P.; Bessert, D.; Zhang, Z.; Mullins, C.; Ghandour, M.S. Proliferation and death of oligodendrocytes and myelin proteins are differentially regulated in male and female rodents. J. Neurosci. 2006, 26, 1439–1447. [Google Scholar] [CrossRef]
- Ghoumari, A.M.; Baulieu, E.E.; Schumacher, M. Progesterone increases oligodendroglial cell proliferation in rat cerebellar slice cultures. Neuroscience 2005, 135, 47–58. [Google Scholar] [CrossRef]
- Swamydas, M.; Bessert, D.; Skoff, R. Sexual dimorphism of oligodendrocytes is mediated by differential regulation of signaling pathways. J. Neurosci. Res. 2009, 87, 3306–3319. [Google Scholar] [CrossRef] [PubMed]
- Atladottir, H.O.; Thorsen, P.; Ostergaard, L.; Schendel, D.E.; Lemcke, S.; Abdallah, M.; Parner, E.T. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J. Autism Dev. Disord 2010, 40, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Hisle-Gorman, E.; Susi, A.; Stokes, T.; Gorman, G.; Erdie-Lalena, C.; Nylund, C.M. Prenatal, perinatal, and neonatal risk factors of autism spectrum disorder. Pediatr. Res. 2018, 84, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Sourander, A.; Hinkka-Yli-Salomaki, S.; McKeague, I.W.; Sundvall, J.; Surcel, H.M. Elevated maternal C-reactive protein and autism in a national birth cohort. Mol. Psychiatry 2014, 19, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Atladottir, H.O.; Thorsen, P.; Schendel, D.E.; Ostergaard, L.; Lemcke, S.; Parner, E.T. Association of hospitalization for infection in childhood with diagnosis of autism spectrum disorders: A Danish cohort study. Arch. Pediatr. Adolesc Med. 2010, 164, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Strickland, A.D. Prevention of cerebral palsy, autism spectrum disorder, and attention deficit-hyperactivity disorder. Med. Hypotheses 2014, 82, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.K.; Fineberg, A.M.; Maxwell, S.D.; Alloy, L.B.; Zimmermann, L.; Krigbaum, N.Y.; Cohn, B.A.; Drabick, D.A.G.; Ellman, L.M. Maternal infection and stress during pregnancy and depressive symptoms in adolescent offspring. Psychiatry Res. 2017, 257, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Gilman, S.E.; Cherkerzian, S.; Buka, S.L.; Hahn, J.; Hornig, M.; Goldstein, J.M. Prenatal immune programming of the sex-dependent risk for major depression. Transl. Psychiatry 2016, 6, e822. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Pedersen, C.B.; Hougaard, D.M.; Norgaard-Petersen, B.; Mors, O.; Borglum, A.D.; Yolken, R.H. A Danish National Birth Cohort study of maternal HSV-2 antibodies as a risk factor for schizophrenia in their offspring. Schizophr. Res. 2010, 122, 257–263. [Google Scholar] [CrossRef]
- Mednick, S.A.; Huttunen, M.O.; Machon, R.A. Prenatal influenza infections and adult schizophrenia. Schizophr. Bull. 1994, 20, 263–267. [Google Scholar] [CrossRef]
- Limosin, F.; Rouillon, F.; Payan, C.; Cohen, J.M.; Strub, N. Prenatal exposure to influenza as a risk factor for adult schizophrenia. Acta Psychiatr. Scand. 2003, 107, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Machon, R.A.; Mednick, S.A.; Huttunen, M.O. Adult major affective disorder after prenatal exposure to an influenza epidemic. Arch. Gen. Psychiatry 1997, 54, 322–328. [Google Scholar] [CrossRef]
- Sorensen, H.J.; Mortensen, E.L.; Reinisch, J.M.; Mednick, S.A. Association between prenatal exposure to bacterial infection and risk of schizophrenia. Schizophr. Bull. 2009, 35, 631–637. [Google Scholar] [CrossRef]
- Takei, N.; Mortensen, P.B.; Klaening, U.; Murray, R.M.; Sham, P.C.; O’Callaghan, E.; Munk-Jorgensen, P. Relationship between in utero exposure to influenza epidemics and risk of schizophrenia in Denmark. Biol. Psychiatry 1996, 40, 817–824. [Google Scholar] [CrossRef]
- Goldstein, J.M.; Cherkerzian, S.; Seidman, L.J.; Donatelli, J.A.; Remington, A.G.; Tsuang, M.T.; Hornig, M.; Buka, S.L. Prenatal maternal immune disruption and sex-dependent risk for psychoses. Psychol. Med. 2014, 44, 3249–3261. [Google Scholar] [CrossRef] [PubMed]
- Masi, A.; Breen, E.J.; Alvares, G.A.; Glozier, N.; Hickie, I.B.; Hunt, A.; Hui, J.; Beilby, J.; Ravine, D.; Wray, J.; et al. Cytokine levels and associations with symptom severity in male and female children with autism spectrum disorder. Mol. Autism 2017, 8, 63. [Google Scholar] [CrossRef]
- Xu, G.; Strathearn, L.; Liu, B.; Bao, W. Prevalence of Autism Spectrum Disorder Among US Children and Adolescents, 2014–2016. JAMA 2018, 319, 81–82. [Google Scholar] [CrossRef]
- Atladottir, H.O.; Schendel, D.E.; Henriksen, T.B.; Hjort, L.; Parner, E.T. Gestational Age and Autism Spectrum Disorder: Trends in Risk Over Time. Autism Res. 2016, 9, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Rao, S.C.; Bulsara, M.K.; Patole, S.K. Prevalence of Autism Spectrum Disorder in Preterm Infants: A Meta-analysis. Pediatrics 2018, 142, e20180134. [Google Scholar] [CrossRef]
- O’Shea, T.M.; Joseph, R.M.; Kuban, K.C.; Allred, E.N.; Ware, J.; Coster, T.; Fichorova, R.N.; Dammann, O.; Leviton, A.; Investigators, E.S. Elevated blood levels of inflammation-related proteins are associated with an attention problem at age 24 mo in extremely preterm infants. Pediatr. Res. 2014, 75, 781–787. [Google Scholar] [CrossRef]
- Limperopoulos, C.; Bassan, H.; Sullivan, N.R.; Soul, J.S.; Robertson, R.L., Jr.; Moore, M.; Ringer, S.A.; Volpe, J.J.; du Plessis, A.J. Positive screening for autism in ex-preterm infants: Prevalence and risk factors. Pediatrics 2008, 121, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Schendel, D.; Bhasin, T.K. Birth weight and gestational age characteristics of children with autism, including a comparison with other developmental disabilities. Pediatrics 2008, 121, 1155–1164. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef]
- Patterson, P.H. Maternal infection and immune involvement in autism. Trends Mol. Med. 2011, 17, 389–394. [Google Scholar] [CrossRef]
- Lammert, C.R.; Frost, E.L.; Bolte, A.C.; Paysour, M.J.; Shaw, M.E.; Bellinger, C.E.; Weigel, T.K.; Zunder, E.R.; Lukens, J.R. Cutting Edge: Critical Roles for Microbiota-Mediated Regulation of the Immune System in a Prenatal Immune Activation Model of Autism. J. Immunol. 2018, 201, 845–850. [Google Scholar] [CrossRef]
- Parracho, H.M.; Bingham, M.O.; Gibson, G.R.; McCartney, A.L. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J. Med. Microbiol 2005, 54, 987–991. [Google Scholar] [CrossRef]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; Youn, E.; Summanen, P.H.; Granpeesheh, D.; Dixon, D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Garbett, K.; Ebert, P.J.; Mitchell, A.; Lintas, C.; Manzi, B.; Mirnics, K.; Persico, A.M. Immune transcriptome alterations in the temporal cortex of subjects with autism. Neurobiol. Dis. 2008, 30, 303–311. [Google Scholar] [CrossRef]
- Ashwood, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Pessah, I.; Van de Water, J. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain Behav. Immun. 2011, 25, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Nadeem, A.; Ansari, M.A.; Bakheet, S.A.; Al-Ayadhi, L.Y.; Attia, S.M. Upregulation of IL-9 and JAK-STAT signaling pathway in children with autism. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79 Pt B, 472–480. [Google Scholar] [CrossRef]
- Krakowiak, P.; Goines, P.E.; Tancredi, D.J.; Ashwood, P.; Hansen, R.L.; Hertz-Picciotto, I.; Van de Water, J. Neonatal Cytokine Profiles Associated With Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 442–451. [Google Scholar] [CrossRef]
- Heuer, L.; Ashwood, P.; Schauer, J.; Goines, P.; Krakowiak, P.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.A.; Pessah, I.N.; Van de Water, J. Reduced levels of immunoglobulin in children with autism correlates with behavioral symptoms. Autism Res. 2008, 1, 275–283. [Google Scholar] [CrossRef]
- Croen, L.A.; Goines, P.; Braunschweig, D.; Yolken, R.; Yoshida, C.K.; Grether, J.K.; Fireman, B.; Kharrazi, M.; Hansen, R.L.; Van de Water, J. Brain-derived neurotrophic factor and autism: Maternal and infant peripheral blood levels in the Early Markers for Autism (EMA) Study. Autism Res. 2008, 1, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Wills, S.; Cabanlit, M.; Bennett, J.; Ashwood, P.; Amaral, D.G.; Van de Water, J. Detection of autoantibodies to neural cells of the cerebellum in the plasma of subjects with autism spectrum disorders. Brain Behav. Immun. 2009, 23, 64–74. [Google Scholar] [CrossRef]
- Schwarz, E.; Guest, P.C.; Rahmoune, H.; Wang, L.; Levin, Y.; Ingudomnukul, E.; Ruta, L.; Kent, L.; Spain, M.; Baron-Cohen, S.; et al. Sex-specific serum biomarker patterns in adults with Asperger’s syndrome. Mol. Psychiatry 2011, 16, 1213–1220. [Google Scholar] [CrossRef]
- Goines, P.E.; Croen, L.A.; Braunschweig, D.; Yoshida, C.K.; Grether, J.; Hansen, R.; Kharrazi, M.; Ashwood, P.; Van de Water, J. Increased midgestational IFN-gamma, IL-4 and IL-5 in women bearing a child with autism: A case-control study. Mol. Autism 2011, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.W.; Larsen, N.; Grove, J.; Norgaard-Pedersen, B.; Thorsen, P.; Mortensen, E.L.; Hougaard, D.M. Amniotic fluid inflammatory cytokines: Potential markers of immunologic dysfunction in autism spectrum disorders. World J. Biol. Psychiatry 2013, 14, 528–538. [Google Scholar] [CrossRef]
- Hornig, M.; Bresnahan, M.A.; Che, X.; Schultz, A.F.; Ukaigwe, J.E.; Eddy, M.L.; Hirtz, D.; Gunnes, N.; Lie, K.K.; Magnus, P.; et al. Reichborn-Kjennerud, T.; Schjolberg, S.; Oyen, A.S.; Levin, B.; Susser, E.S.; Stoltenberg, C.; Lipkin, W.I. Prenatal fever and autism risk. Mol. Psychiatry 2018, 23, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Zerbo, O.; Iosif, A.M.; Walker, C.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Is maternal influenza or fever during pregnancy associated with autism or developmental delays? Results from the CHARGE (CHildhood Autism Risks from Genetics and Environment) study. J. Autism Dev. Disord 2013, 43, 25–33. [Google Scholar] [CrossRef]
- Avella-Garcia, C.B.; Julvez, J.; Fortuny, J.; Rebordosa, C.; Garcia-Esteban, R.; Galan, I.R.; Tardon, A.; Rodriguez-Bernal, C.L.; Iniguez, C.; Andiarena, A.; et al. Acetaminophen use in pregnancy and neurodevelopment: Attention function and autism spectrum symptoms. Int. J. Epidemiol. 2016, 45, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Enstrom, A.; Onore, C.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.; Van de Water, J.; Ashwood, P. Detection of IL-17 and IL-23 in Plasma Samples of Children with Autism. Am. J. Biochem. Biotechnol. 2008, 4, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Enstrom, A.; Onore, C.; Tarver, A.; Hertz-Picciotto, I.; Hansen, R.; Croen, L.; Van de Water, J.; Ashwood, P. Peripheral Blood Leukocyte Production of BDNF following Mitogen Stimulation in Early Onset and Regressive Autism. Am. J. Biochem. Biotechnol. 2008, 4, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H.; Geng, L.; Cushing-Ruby, A.; Quraishi, H. Impact of innate immunity in a subset of children with autism spectrum disorders: A case control study. J. Neuroinflamm. 2008, 5, 52. [Google Scholar] [CrossRef]
- Enstrom, A.M.; Onore, C.E.; Van de Water, J.A.; Ashwood, P. Differential monocyte responses to TLR ligands in children with autism spectrum disorders. Brain Behav. Immun. 2010, 24, 64–71. [Google Scholar] [CrossRef]
- Mallard, C. Innate immune regulation by toll-like receptors in the brain. ISRN Neurol. 2012, 2012, 701950. [Google Scholar] [CrossRef]
- Du, X.; Fleiss, B.; Li, H.; D’Angelo, B.; Sun, Y.; Zhu, C.; Hagberg, H.; Levy, O.; Mallard, C.; Wang, X. Systemic stimulation of TLR2 impairs neonatal mouse brain development. PLoS ONE 2011, 6, e19583. [Google Scholar] [CrossRef]
- Polanczyk, G.; de Lima, M.S.; Horta, B.L.; Biederman, J.; Rohde, L.A. The worldwide prevalence of ADHD: A systematic review and metaregression analysis. Am. J. Psychiatry 2007, 164, 942–948. [Google Scholar] [CrossRef]
- Asherson, P.; Buitelaar, J.; Faraone, S.V.; Rohde, L.A. Adult attention-deficit hyperactivity disorder: Key conceptual issues. Lancet Psychiatry 2016, 3, 568–578. [Google Scholar] [CrossRef]
- Rucklidge, J.J. Gender differences in attention-deficit/hyperactivity disorder. Psychiatr. Clin. North. Am. 2010, 33, 357–373. [Google Scholar] [CrossRef] [PubMed]
- Leviton, A.; Hooper, S.R.; Hunter, S.J.; Scott, M.N.; Allred, E.N.; Joseph, R.M.; O’Shea, T.M.; Kuban, K.; Investigators, E.S. Antecedents of Screening Positive for Attention Deficit Hyperactivity Disorder in Ten-Year-Old Children Born Extremely Preterm. Pediatr. Neurol. 2018, 81, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Allred, E.N.; Dammann, O.; Fichorova, R.N.; Hooper, S.R.; Hunter, S.J.; Joseph, R.M.; Kuban, K.; Leviton, A.; O’Shea, T.M.; Scott, M.N.; et al. Systemic Inflammation during the First Postnatal Month and the Risk of Attention Deficit Hyperactivity Disorder Characteristics among 10 year-old Children Born Extremely Preterm. J. Neuroimmune Pharmacol. 2017, 12, 531–543. [Google Scholar] [CrossRef]
- Instanes, J.T.; Halmoy, A.; Engeland, A.; Haavik, J.; Furu, K.; Klungsoyr, K. Attention-Deficit/Hyperactivity Disorder in Offspring of Mothers With Inflammatory and Immune System Diseases. Biol. Psychiatry 2017, 81, 452–459. [Google Scholar] [CrossRef]
- Giana, G.; Romano, E.; Porfirio, M.C.; D’Ambrosio, R.; Giovinazzo, S.; Troianiello, M.; Barlocci, E.; Travaglini, D.; Granstrem, O.; Pascale, E.; et al. Detection of auto-antibodies to DAT in the serum: Interactions with DAT genotype and psycho-stimulant therapy for ADHD. J. Neuroimmunol. 2015, 278, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Wittchen, H.U.; Jacobi, F.; Rehm, J.; Gustavsson, A.; Svensson, M.; Jonsson, B.; Olesen, J.; Allgulander, C.; Alonso, J.; Faravelli, C.; et al. The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur. Neuropsychopharmacol. 2011, 21, 655–679. [Google Scholar] [CrossRef]
- Fasseeh, A.; Nemeth, B.; Molnar, A.; Fricke, F.U.; Horvath, M.; Koczian, K.; Gotze, A.; Kalo, Z. A systematic review of the indirect costs of schizophrenia in Europe. Eur. J. Public Health 2018, 28, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Begg, M.D.; Gravenstein, S.; Schaefer, C.A.; Wyatt, R.J.; Bresnahan, M.; Babulas, V.P.; Susser, E.S. Serologic evidence of prenatal influenza in the etiology of schizophrenia. Arch. Gen. Psychiatry 2004, 61, 774–780. [Google Scholar] [CrossRef]
- Blomstrom, A.; Karlsson, H.; Wicks, S.; Yang, S.; Yolken, R.H.; Dalman, C. Maternal antibodies to infectious agents and risk for non-affective psychoses in the offspring--a matched case-control study. Schizophr Res. 2012, 140, 25–30. [Google Scholar] [CrossRef]
- Takei, N.; O’Callaghan, E.; Sham, P.C.; Glover, G.; Murray, R.M. Does prenatal influenza divert susceptible females from later affective psychosis to schizophrenia? Acta Psychiatr. Scand. 1993, 88, 328–336. [Google Scholar] [CrossRef]
- Nahmias, A.J.; Nahmias, S.B.; Danielsson, D. The possible role of transplacentally-acquired antibodies to infectious agents, with molecular mimicry to nervous system sialic acid epitopes, as causes of neuromental disorders: Prevention and vaccine implications. Clin. Dev. Immunol. 2006, 13, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.M.; Schwarz, E.; Guest, P.C.; van Beveren, N.J.; Leweke, F.M.; Rothermundt, M.; Bogerts, B.; Steiner, J.; Bahn, S. Distinct molecular phenotypes in male and female schizophrenia patients. PLoS ONE 2013, 8, e78729. [Google Scholar] [CrossRef] [PubMed]
- Vandeleur, C.L.; Fassassi, S.; Castelao, E.; Glaus, J.; Strippoli, M.F.; Lasserre, A.M.; Rudaz, D.; Gebreab, S.; Pistis, G.; Aubry, J.M.; et al. Prevalence and correlates of DSM-5 major depressive and related disorders in the community. Psychiatry Res. 2017, 250, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, R.; Weidenfeld, J.; Pollak, Y.; Morag, M.; Morag, A.; Avitsur, R.; Barak, O.; Reichenberg, A.; Cohen, E.; Shavit, Y.; et al. Cytokines, “depression due to a general medical condition,” and antidepressant drugs. Adv. Exp. Med. Biol. 1999, 461, 283–316. [Google Scholar]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctot, K.L. A meta-analysis of cytokines in major depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Bendix, I.; Hadamitzky, M.; Herz, J.; Felderhoff-Muser, U. Adverse neuropsychiatric development following perinatal brain injury: From a preclinical perspective. Pediatr. Res. 2018. [Google Scholar] [CrossRef]
- Connors, E.J.; Shaik, A.N.; Migliore, M.M.; Kentner, A.C. Environmental enrichment mitigates the sex-specific effects of gestational inflammation on social engagement and the hypothalamic pituitary adrenal axis-feedback system. Brain Behav. Immun. 2014, 42, 178–190. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Gagnidze, K.; Reyes, A.; Norstedt, N.; Mansson, K.; Francis, K.; Pfaff, D.W. Sex-specific gene-environment interactions underlying ASD-like behaviors. Proc. Natl. Acad. Sci. USA 2017, 114, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Xuan, I.C.; Hampson, D.R. Gender-dependent effects of maternal immune activation on the behavior of mouse offspring. PLoS ONE 2014, 9, e104433. [Google Scholar] [CrossRef]
- Schwartzer, J.J.; Careaga, M.; Onore, C.E.; Rushakoff, J.A.; Berman, R.F.; Ashwood, P. Maternal immune activation and strain specific interactions in the development of autism-like behaviors in mice. Transl. Psychiatry 2013, 3, e240. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Cossio, L.; Guzman, A.; van der Veldt, S.; Luheshi, G.N. Prenatal infection leads to ASD-like behavior and altered synaptic pruning in the mouse offspring. Brain Behav. Immun. 2017, 63, 88–98. [Google Scholar] [CrossRef]
- Basta-Kaim, A.; Fijal, K.; Slusarczyk, J.; Trojan, E.; Glombik, K.; Budziszewska, B.; Leskiewicz, M.; Regulska, M.; Kubera, M.; Lason, W.; et al. Prenatal administration of lipopolysaccharide induces sex-dependent changes in glutamic acid decarboxylase and parvalbumin in the adult rat brain. Neuroscience 2015, 287, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Basta-Kaim, A.; Budziszewska, B.; Leskiewicz, M.; Fijal, K.; Regulska, M.; Kubera, M.; Wedzony, K.; Lason, W. Hyperactivity of the hypothalamus-pituitary-adrenal axis in lipopolysaccharide-induced neurodevelopmental model of schizophrenia in rats: Effects of antipsychotic drugs. Eur. J. Pharmacol. 2011, 650, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Harvey, L.; Boksa, P. A stereological comparison of GAD67 and reelin expression in the hippocampal stratum oriens of offspring from two mouse models of maternal inflammation during pregnancy. Neuropharmacology 2012, 62, 1767–1776. [Google Scholar] [CrossRef]
- Wischhof, L.; Irrsack, E.; Osorio, C.; Koch, M. Prenatal LPS-exposure--a neurodevelopmental rat model of schizophrenia--differentially affects cognitive functions, myelination and parvalbumin expression in male and female offspring. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 57, 17–30. [Google Scholar] [CrossRef]
- Howland, J.G.; Cazakoff, B.N.; Zhang, Y. Altered object-in-place recognition memory, prepulse inhibition, and locomotor activity in the offspring of rats exposed to a viral mimetic during pregnancy. Neuroscience 2012, 201, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cazakoff, B.N.; Thai, C.A.; Howland, J.G. Prenatal exposure to a viral mimetic alters behavioural flexibility in male, but not female, rats. Neuropharmacology 2012, 62, 1299–1307. [Google Scholar] [CrossRef]
- Kentner, A.C.; McLeod, S.A.; Field, E.F.; Pittman, Q.J. Sex-dependent effects of neonatal inflammation on adult inflammatory markers and behavior. Endocrinology 2010, 151, 2689–2699. [Google Scholar] [CrossRef]
- Lante, F.; Meunier, J.; Guiramand, J.; Maurice, T.; Cavalier, M.; Ferreira, M.C.D.; Aimar, R.; Cohen-Solal, C.; Vignes, M.; Barbanel, G. Neurodevelopmental damage after prenatal infection: Role of oxidative stress in the fetal brain. Free Radical Bio Med. 2007, 42, 1231–1245. [Google Scholar] [CrossRef]
- Bitanihirwe, B.K.; Peleg-Raibstein, D.; Mouttet, F.; Feldon, J.; Meyer, U. Late prenatal immune activation in mice leads to behavioral and neurochemical abnormalities relevant to the negative symptoms of schizophrenia. Neuropsychopharmacology 2010, 35, 2462–2478. [Google Scholar] [CrossRef]
- Bronson, S.L.; Bale, T.L. Prenatal stress-induced increases in placental inflammation and offspring hyperactivity are male-specific and ameliorated by maternal antiinflammatory treatment. Endocrinology 2014, 155, 2635–2646. [Google Scholar] [CrossRef]
- Taylor, P.V.; Veenema, A.H.; Paul, M.J.; Bredewold, R.; Isaacs, S.; de Vries, G.J. Sexually dimorphic effects of a prenatal immune challenge on social play and vasopressin expression in juvenile rats. Biol. Sex Differ. 2012, 3, 15. [Google Scholar] [CrossRef]
- Borrell, J.; Vela, J.M.; Arevalo-Martin, A.; Molina-Holgado, E.; Guaza, C. Prenatal immune challenge disrupts sensorimotor gating in adult rats. Implications for the etiopathogenesis of schizophrenia. Neuropsychopharmacology 2002, 26, 204–215. [Google Scholar] [CrossRef]
- Custodio, C.S.; Mello, B.S.F.; Filho, A.; de Carvalho Lima, C.N.; Cordeiro, R.C.; Miyajima, F.; Reus, G.Z.; Vasconcelos, S.M.M.; Barichello, T.; Quevedo, J.; et al. Neonatal Immune Challenge with Lipopolysaccharide Triggers Long-lasting Sex- and Age-related Behavioral and Immune/Neurotrophic Alterations in Mice: Relevance to Autism Spectrum Disorders. Mol. Neurobiol. 2018, 55, 3775–3788. [Google Scholar] [CrossRef]
- Patrich, E.; Piontkewitz, Y.; Peretz, A.; Weiner, I.; Attali, B. Maturation- and sex-sensitive depression of hippocampal excitatory transmission in a rat schizophrenia model. Brain Behav. Immun. 2016, 51, 240–251. [Google Scholar] [CrossRef]
- Bauman, M.D.; Iosif, A.M.; Ashwood, P.; Braunschweig, D.; Lee, A.; Schumann, C.M.; Van de Water, J.; Amaral, D.G. Maternal antibodies from mothers of children with autism alter brain growth and social behavior development in the rhesus monkey. Transl. Psychiatry 2013, 3, e278. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Valdez, R.; Mottahedin, A.; Stridh, L.; Yellowhair, T.R.; Jantzie, L.L.; Northington, F.J.; Mallard, C. Evidence for Sexual Dimorphism in the Response to TLR3 Activation in the Developing Neonatal Mouse Brain: A Pilot Study. Front. Physiol. 2019, 10, 306. [Google Scholar] [CrossRef]
- Naviaux, R.K.; Zolkipli, Z.; Wang, L.; Nakayama, T.; Naviaux, J.C.; Le, T.P.; Schuchbauer, M.A.; Rogac, M.; Tang, Q.; Dugan, L.L.; et al. Antipurinergic therapy corrects the autism-like features in the poly(IC) mouse model. PLoS ONE 2013, 8, e57380. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Napoli, E.; Schwartzer, J.; Careaga, M.; Ashwood, P. Gestational exposure to a viral mimetic poly(i:C) results in long-lasting changes in mitochondrial function by leucocytes in the adult offspring. Mediators Inflamm. 2013, 2013, 609602. [Google Scholar] [CrossRef]
- Bauman, M.D.; Iosif, A.M.; Smith, S.E.; Bregere, C.; Amaral, D.G.; Patterson, P.H. Activation of the maternal immune system during pregnancy alters behavioral development of rhesus monkey offspring. Biol. Psychiatry 2014, 75, 332–341. [Google Scholar] [CrossRef]
- Malkova, N.V.; Yu, C.Z.; Hsiao, E.Y.; Moore, M.J.; Patterson, P.H. Maternal immune activation yields offspring displaying mouse versions of the three core symptoms of autism. Brain Behav. Immun. 2012, 26, 607–616. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Bilbo, S.D. Sex, glia, and development: Interactions in health and disease. Horm. Behav. 2012, 62, 243–253. [Google Scholar] [CrossRef]
- Meyer, U.; Feldon, J.; Schedlowski, M.; Yee, B.K. Immunological stress at the maternal-foetal interface: A link between neurodevelopment and adult psychopathology. Brain Behav. Immun. 2006, 20, 378–388. [Google Scholar] [CrossRef]
- Meyer, U.; Nyffeler, M.; Engler, A.; Urwyler, A.; Schedlowski, M.; Knuesel, I.; Yee, B.K.; Feldon, J. The time of prenatal immune challenge determines the specificity of inflammation-mediated brain and behavioral pathology. J. Neurosci. 2006, 26, 4752–4762. [Google Scholar] [CrossRef]
- Schwendener, S.; Meyer, U.; Feldon, J. Deficient maternal care resulting from immunological stress during pregnancy is associated with a sex-dependent enhancement of conditioned fear in the offspring. J. Neurodev. Disord. 2009, 1, 15–32. [Google Scholar] [CrossRef]
- Enayati, M.; Solati, J.; Hosseini, M.H.; Shahi, H.R.; Saki, G.; Salari, A.A. Maternal infection during late pregnancy increases anxiety- and depression-like behaviors with increasing age in male offspring. Brain Res. Bull. 2012, 87, 295–302. [Google Scholar] [CrossRef]
- Lin, Y.L.; Wang, S. Prenatal lipopolysaccharide exposure increases depression-like behaviors and reduces hippocampal neurogenesis in adult rats. Behav. Brain Res. 2014, 259, 24–34. [Google Scholar] [CrossRef]
- Kohman, R.A.; Tarr, A.J.; Sparkman, N.L.; Bogale, T.M.; Boehm, G.W. Neonatal endotoxin exposure impairs avoidance learning and attenuates endotoxin-induced sickness behavior and central IL-1beta gene transcription in adulthood. Behav. Brain Res. 2008, 194, 25–31. [Google Scholar] [CrossRef]
- Missig, G.; Robbins, J.O.; Mokler, E.L.; McCullough, K.M.; Bilbo, S.D.; McDougle, C.J.; Carlezon, W.A., Jr. Sex-dependent neurobiological features of prenatal immune activation via TLR7. Mol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Allard, M.J.; Brochu, M.E.; Bergeron, J.D.; Sebire, G. Hyperactive behavior in female rats in utero-exposed to group B Streptococcus-induced inflammation. Int. J. Dev. Neurosci. 2018, 69, 17–22. [Google Scholar] [CrossRef]
- Gantert, M.; Kreczmanski, P.; Kuypers, E.; Jellema, R.; Strackx, E.; Bastian, N.; Gavilanes, A.W.; Zimmermann, L.J.; Garnier, Y.; Schmitz, C.; et al. Effects of in utero endotoxemia on the ovine fetal brain: A model for schizophrenia? Front. Biosci. 2012, 4, 2745–2753. [Google Scholar]
- Rose, D.R.; Careaga, M.; Van de Water, J.; McAllister, K.; Bauman, M.D.; Ashwood, P. Long-term altered immune responses following fetal priming in a non-human primate model of maternal immune activation. Brain Behav. Immun. 2017, 63, 60–70. [Google Scholar] [CrossRef]
- Machado, C.J.; Whitaker, A.M.; Smith, S.E.; Patterson, P.H.; Bauman, M.D. Maternal immune activation in nonhuman primates alters social attention in juvenile offspring. Biol. Psychiatry 2015, 77, 823–832. [Google Scholar] [CrossRef]
- Mooney, M.A.; McWeeney, S.K.; Faraone, S.V.; Hinney, A.; Hebebrand, J.; Consortium, I.; German, A.G.G.; Nigg, J.T.; Wilmot, B. Pathway analysis in attention deficit hyperactivity disorder: An ensemble approach. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171, 815–826. [Google Scholar] [CrossRef]
- Gallo, E.F.; Posner, J. Moving towards causality in attention-deficit hyperactivity disorder: Overview of neural and genetic mechanisms. Lancet Psychiatry 2016, 3, 555–567. [Google Scholar] [CrossRef]
- Abazyan, B.; Nomura, J.; Kannan, G.; Ishizuka, K.; Tamashiro, K.L.; Nucifora, F.; Pogorelov, V.; Ladenheim, B.; Yang, C.; Krasnova, I.N.; et al. Prenatal interaction of mutant DISC1 and immune activation produces adult psychopathology. Biol. Psychiatry 2010, 68, 1172–1181. [Google Scholar] [CrossRef]
- Goldstein, J.M. Sex, hormones and affective arousal circuitry dysfunction in schizophrenia. Horm. Behav. 2006, 50, 612–622. [Google Scholar] [CrossRef]
- Goldstein, J.M.; Seidman, L.J.; Makris, N.; Ahern, T.; O’Brien, L.M.; Caviness, V.S., Jr.; Kennedy, D.N.; Faraone, S.V.; Tsuang, M.T. Hypothalamic abnormalities in schizophrenia: Sex effects and genetic vulnerability. Biol. Psychiatry 2007, 61, 935–945. [Google Scholar] [CrossRef]
- Labouesse, M.A.; Dong, E.; Grayson, D.R.; Guidotti, A.; Meyer, U. Maternal immune activation induces GAD1 and GAD2 promoter remodeling in the offspring prefrontal cortex. Epigenetics 2015, 10, 1143–1155. [Google Scholar] [CrossRef]
- Goeden, N.; Velasquez, J.; Arnold, K.A.; Chan, Y.; Lund, B.T.; Anderson, G.M.; Bonnin, A. Maternal Inflammation Disrupts Fetal Neurodevelopment via Increased Placental Output of Serotonin to the Fetal Brain. J. Neurosci. 2016, 36, 6041–6049. [Google Scholar] [CrossRef]
- Foley, K.A.; Ossenkopp, K.P.; Kavaliers, M.; Macfabe, D.F. Pre- and neonatal exposure to lipopolysaccharide or the enteric metabolite, propionic acid, alters development and behavior in adolescent rats in a sexually dimorphic manner. PLoS ONE 2014, 9, e87072. [Google Scholar] [CrossRef]
- Stephenson, D.T.; O’Neill, S.M.; Narayan, S.; Tiwari, A.; Arnold, E.; Samaroo, H.D.; Du, F.; Ring, R.H.; Campbell, B.; Pletcher, M.; et al. Histopathologic characterization of the BTBR mouse model of autistic-like behavior reveals selective changes in neurodevelopmental proteins and adult hippocampal neurogenesis. Mol. Autism 2011, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Coretti, L.; Cristiano, C.; Florio, E.; Scala, G.; Lama, A.; Keller, S.; Cuomo, M.; Russo, R.; Pero, R.; Paciello, O.; et al. Sex-related alterations of gut microbiota composition in the BTBR mouse model of autism spectrum disorder. Sci. Rep. 2017, 7, 45356. [Google Scholar] [CrossRef]
- Schwartzer, J.J.; Onore, C.E.; Rose, D.; Ashwood, P. C57BL/6J bone marrow transplant increases sociability in BTBR T(+) Itpr3(tf)/J mice. Brain Behav. Immun. 2017, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Matcovitch-Natan, O.; Winter, D.R.; Giladi, A.; Vargas Aguilar, S.; Spinrad, A.; Sarrazin, S.; Ben-Yehuda, H.; David, E.; Zelada Gonzalez, F.; Perrin, P.; et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science 2016, 353, aad8670. [Google Scholar] [CrossRef]
- Nelson, L.H.; Lenz, K.M. Microglia depletion in early life programs persistent changes in social, mood-related, and locomotor behavior in male and female rats. Behav. Brain Res. 2017, 316, 279–293. [Google Scholar] [CrossRef]
- Kim, H.J.; Cho, M.H.; Shim, W.H.; Kim, J.K.; Jeon, E.Y.; Kim, D.H.; Yoon, S.Y. Deficient autophagy in microglia impairs synaptic pruning and causes social behavioral defects. Mol. Psychiatry 2017, 22, 1576–1584. [Google Scholar] [CrossRef]
- Loram, L.C.; Sholar, P.W.; Taylor, F.R.; Wiesler, J.L.; Babb, J.A.; Strand, K.A.; Berkelhammer, D.; Day, H.E.; Maier, S.F.; Watkins, L.R. Sex and estradiol influence glial pro-inflammatory responses to lipopolysaccharide in rats. Psychoneuroendocrinology 2012, 37, 1688–1699. [Google Scholar] [CrossRef] [PubMed]
- Patisaul, H.B.; Sullivan, A.W.; Radford, M.E.; Walker, D.M.; Adewale, H.B.; Winnik, B.; Coughlin, J.L.; Buckley, B.; Gore, A.C. Anxiogenic effects of developmental bisphenol A exposure are associated with gene expression changes in the juvenile rat amygdala and mitigated by soy. PLoS ONE 2012, 7, e43890. [Google Scholar] [CrossRef]
- Rebuli, M.E.; Gibson, P.; Rhodes, C.L.; Cushing, B.S.; Patisaul, H.B. Sex differences in microglial colonization and vulnerabilities to endocrine disruption in the social brain. Gen. Comp. Endocrinol. 2016, 238, 39–46. [Google Scholar] [CrossRef]
- Zhan, Y.; Paolicelli, R.C.; Sforazzini, F.; Weinhard, L.; Bolasco, G.; Pagani, F.; Vyssotski, A.L.; Bifone, A.; Gozzi, A.; Ragozzino, D.; et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 2014, 17, 400–406. [Google Scholar] [CrossRef]
- Derecki, N.C.; Cronk, J.C.; Lu, Z.; Xu, E.; Abbott, S.B.; Guyenet, P.G.; Kipnis, J. Wild-type microglia arrest pathology in a mouse model of Rett syndrome. Nature 2012, 484, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Nance, E.; Kambhampati, S.P.; Smith, E.S.; Zhang, Z.; Zhang, F.; Singh, S.; Johnston, M.V.; Kannan, R.M.; Blue, M.E.; Kannan, S. Dendrimer-mediated delivery of N-acetyl cysteine to microglia in a mouse model of Rett syndrome. J. Neuroinflamm. 2017, 14, 252. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Lawson, M.A.; Andre, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol. Psychiatry 2009, 14, 511–522. [Google Scholar] [CrossRef]
- Campbell, B.M.; Charych, E.; Lee, A.W.; Moller, T. Kynurenines in CNS disease: Regulation by inflammatory cytokines. Front. Neurosci. 2014, 8, 12. [Google Scholar] [CrossRef]
- Forrest, C.M.; Khalil, O.S.; Pisar, M.; Darlington, L.G.; Stone, T.W. Prenatal inhibition of the tryptophan-kynurenine pathway alters synaptic plasticity and protein expression in the rat hippocampus. Brain Res. 2013, 1504, 1–15. [Google Scholar] [CrossRef]
- O’Roak, B.J.; Vives, L.; Fu, W.; Egertson, J.D.; Stanaway, I.B.; Phelps, I.G.; Carvill, G.; Kumar, A.; Lee, C.; Ankenman, K.; et al. Multiplex targeted sequencing identifies recurrently mutated genes in autism spectrum disorders. Science 2012, 338, 1619–1622. [Google Scholar] [CrossRef]
- Malishkevich, A.; Amram, N.; Hacohen-Kleiman, G.; Magen, I.; Giladi, E.; Gozes, I. Activity-dependent neuroprotective protein (ADNP) exhibits striking sexual dichotomy impacting on autistic and Alzheimer’s pathologies. Transl. Psychiatry 2015, 5, e501. [Google Scholar] [CrossRef]
- Quesseveur, G.; David, D.J.; Gaillard, M.C.; Pla, P.; Wu, M.V.; Nguyen, H.T.; Nicolas, V.; Auregan, G.; David, I.; Dranovsky, A.; et al. BDNF overexpression in mouse hippocampal astrocytes promotes local neurogenesis and elicits anxiolytic-like activities. Transl. Psychiatry 2013, 3, e253. [Google Scholar] [CrossRef]
- Liu, M.; Hurn, P.D.; Roselli, C.E.; Alkayed, N.J. Role of P450 aromatase in sex-specific astrocytic cell death. J. Cereb. Blood Flow Metab. 2007, 27, 135–141. [Google Scholar] [CrossRef]
- Garcia-Segura, L.M.; Melcangi, R.C. Steroids and glial cell function. Glia 2006, 54, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Veiga, S.; Carrero, P.; Pernia, O.; Azcoitia, I.; Garcia-Segura, L.M. Translocator protein 18 kDa is involved in the regulation of reactive gliosis. Glia 2007, 55, 1426–1436. [Google Scholar] [CrossRef]
- Veenman, L.; Papadopoulos, V.; Gavish, M. Channel-like functions of the 18-kDa translocator protein (TSPO): Regulation of apoptosis and steroidogenesis as part of the host-defense response. Curr. Pharm. Des. 2007, 13, 2385–2405. [Google Scholar] [CrossRef]
- Sugai, T.; Kawamura, M.; Iritani, S.; Araki, K.; Makifuchi, T.; Imai, C.; Nakamura, R.; Kakita, A.; Takahashi, H.; Nawa, H. Prefrontal abnormality of schizophrenia revealed by DNA microarray: Impact on glial and neurotrophic gene expression. Ann. N. Y. Acad. Sci. 2004, 1025, 84–91. [Google Scholar] [CrossRef]
- Foong, J.; Symms, M.R.; Barker, G.J.; Maier, M.; Miller, D.H.; Ron, M.A. Investigating regional white matter in schizophrenia using diffusion tensor imaging. Neuroreport 2002, 13, 333–336. [Google Scholar] [CrossRef]
- Uranova, N.; Orlovskaya, D.; Vikhreva, O.; Zimina, I.; Kolomeets, N.; Vostrikov, V.; Rachmanova, V. Electron microscopy of oligodendroglia in severe mental illness. Brain Res. Bull. 2001, 55, 597–610. [Google Scholar] [CrossRef]
- van Tilborg, E.; Heijnen, C.J.; Benders, M.J.; van Bel, F.; Fleiss, B.; Gressens, P.; Nijboer, C.H. Impaired oligodendrocyte maturation in preterm infants: Potential therapeutic targets. Prog. Neurobiol. 2016, 136, 28–49. [Google Scholar] [CrossRef]
- Bansal, R.; Lakhina, V.; Remedios, R.; Tole, S. Expression of FGF receptors 1, 2, 3 in the embryonic and postnatal mouse brain compared with Pdgfralpha, Olig2 and Plp/dm20: Implications for oligodendrocyte development. Dev. Neurosci. 2003, 25, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Makinodan, M.; Tatsumi, K.; Manabe, T.; Yamauchi, T.; Makinodan, E.; Matsuyoshi, H.; Shimoda, S.; Noriyama, Y.; Kishimoto, T.; Wanaka, A. Maternal immune activation in mice delays myelination and axonal development in the hippocampus of the offspring. J. Neurosci. Res. 2008, 86, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, J.D.; Deslauriers, J.; Grignon, S.; Fortier, L.C.; Lepage, M.; Stroh, T.; Poyart, C.; Sebire, G. White matter injury and autistic-like behavior predominantly affecting male rat offspring exposed to group B streptococcal maternal inflammation. Dev. Neurosci. 2013, 35, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Rousset, C.I.; Chalon, S.; Cantagrel, S.; Bodard, S.; Andres, C.; Gressens, P.; Saliba, E. Maternal exposure to LPS induces hypomyelination in the internal capsule and programmed cell death in the deep gray matter in newborn rats. Pediatr. Res. 2006, 59, 428–433. [Google Scholar] [CrossRef]
- Pang, Y.; Cai, Z.; Rhodes, P.G. Effects of lipopolysaccharide on oligodendrocyte progenitor cells are mediated by astrocytes and microglia. J. Neurosci. Res. 2000, 62, 510–520. [Google Scholar] [CrossRef]
- Boksa, P. Effects of prenatal infection on brain development and behavior: A review of findings from animal models. Brain Behav. Immun. 2010, 24, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.B.; Sheng, G.Q. Interleukin-1beta with learning and memory. Neurosci. Bull. 2010, 26, 455–468. [Google Scholar] [CrossRef]
- Williamson, L.L.; Sholar, P.W.; Mistry, R.S.; Smith, S.H.; Bilbo, S.D. Microglia and memory: Modulation by early-life infection. J. Neurosci. 2011, 31, 15511–15521. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Zou, H.; Sheikh, A.M.; Malik, M.; Dobkin, C.; Brown, W.T.; Li, X. IL-6 is increased in the cerebellum of autistic brain and alters neural cell adhesion, migration and synaptic formation. J. Neuroinflamm. 2011, 8, 52. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; Patterson, P.H. Activation of the maternal immune system induces endocrine changes in the placenta via IL-6. Brain Behav. Immun. 2011, 25, 604–615. [Google Scholar] [CrossRef]
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef]
- Gilmore, J.H.; Fredrik Jarskog, L.; Vadlamudi, S.; Lauder, J.M. Prenatal infection and risk for schizophrenia: IL-1beta, IL-6, and TNFalpha inhibit cortical neuron dendrite development. Neuropsychopharmacology 2004, 29, 1221–1229. [Google Scholar] [CrossRef]
- Samuelsson, A.M.; Jennische, E.; Hansson, H.A.; Holmang, A. Prenatal exposure to interleukin-6 results in inflammatory neurodegeneration in hippocampus with NMDA/GABA(A) dysregulation and impaired spatial learning. Am. J. Physiol. Regul Integr Comp. Physiol. 2006, 290, R1345–R1356. [Google Scholar] [CrossRef]
- Mottahedin, A.; Ardalan, M.; Chumak, T.; Riebe, I.; Ek, J.; Mallard, C. Effect of Neuroinflammation on Synaptic Organization and Function in the Developing Brain: Implications for Neurodevelopmental and Neurodegenerative Disorders. Front. Cell Neurosci. 2017, 11, 190. [Google Scholar] [CrossRef]
- Veerhuis, R.; Nielsen, H.M.; Tenner, A.J. Complement in the brain. Mol. Immunol. 2011, 48, 1592–1603. [Google Scholar] [CrossRef]
- Kojouharova, M.; Reid, K.; Gadjeva, M. New insights into the molecular mechanisms of classical complement activation. Mol. Immunol. 2010, 47, 2154–2160. [Google Scholar] [CrossRef]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; Ransohoff, R.M.; Greenberg, M.E.; Barres, B.A.; Stevens, B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 2012, 74, 691–705. [Google Scholar] [CrossRef]
- Jarlestedt, K.; Rousset, C.I.; Stahlberg, A.; Sourkova, H.; Atkins, A.L.; Thornton, C.; Barnum, S.R.; Wetsel, R.A.; Dragunow, M.; Pekny, M.; et al. Receptor for complement peptide C3a: A therapeutic target for neonatal hypoxic-ischemic brain injury. FASEB J. 2013, 27, 3797–3804. [Google Scholar] [CrossRef]
- Moran, J.; Stokowska, A.; Walker, F.R.; Mallard, C.; Hagberg, H.; Pekna, M. Intranasal C3a treatment ameliorates cognitive impairment in a mouse model of neonatal hypoxic-ischemic brain injury. Exp. Neurol. 2017, 290, 74–84. [Google Scholar] [CrossRef]
- Monji, A.; Kato, T.; Kanba, S. Cytokines and schizophrenia: Microglia hypothesis of schizophrenia. Psychiatry Clin. Neurosci. 2009, 63, 257–265. [Google Scholar] [CrossRef]
- Pardo, C.A.; Vargas, D.L.; Zimmerman, A.W. Immunity, neuroglia and neuroinflammation in autism. Int. Rev. Psychiatry 2005, 17, 485–495. [Google Scholar] [CrossRef]
- Sekar, A.; Bialas, A.R.; de Rivera, H.; Davis, A.; Hammond, T.R.; Kamitaki, N.; Tooley, K.; Presumey, J.; Baum, M.; Van Doren, V.; et al. Schizophrenia Working Group of the Psychiatric Genomics, C.; Daly, M.J.; Carroll, M.C.; Stevens, B.; McCarroll, S.A. Schizophrenia risk from complex variation of complement component 4. Nature 2016, 530, 177–183. [Google Scholar] [CrossRef]
- Ma, Y.; Ramachandran, A.; Ford, N.; Parada, I.; Prince, D.A. Remodeling of dendrites and spines in the C1q knockout model of genetic epilepsy. Epilepsia 2013, 54, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Corbett, B.A.; Kantor, A.B.; Schulman, H.; Walker, W.L.; Lit, L.; Ashwood, P.; Rocke, D.M.; Sharp, F.R. A proteomic study of serum from children with autism showing differential expression of apolipoproteins and complement proteins. Mol. Psychiatry 2007, 12, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Coulthard, L.G.; Hawksworth, O.A.; Li, R.; Balachandran, A.; Lee, J.D.; Sepehrband, F.; Kurniawan, N.; Jeanes, A.; Simmons, D.G.; Wolvetang, E.; et al. Complement C5aR1 Signaling Promotes Polarization and Proliferation of Embryonic Neural Progenitor Cells through PKCzeta. J. Neurosci. 2017, 37, 5395–5407. [Google Scholar] [CrossRef]
- Arion, D.; Unger, T.; Lewis, D.A.; Levitt, P.; Mirnics, K. Molecular evidence for increased expression of genes related to immune and chaperone function in the prefrontal cortex in schizophrenia. Biol. Psychiatry 2007, 62, 711–721. [Google Scholar] [CrossRef]
- Meyer, U.; Feldon, J.; Dammann, O. Schizophrenia and autism: Both shared and disorder-specific pathogenesis via perinatal inflammation? Pediatr. Res. 2011, 69, 26R–33R. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, U.; Quinn, T.; Walker, D.W.; Dickinson, H. Cytokines and the neurodevelopmental basis of mental illness. Front. Neurosci. 2013, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Karimi, P.; Kamali, E.; Mousavi, S.M.; Karahmadi, M. Environmental factors influencing the risk of autism. J. Res. Med. Sci. 2017, 22, 27. [Google Scholar] [PubMed]

{kind=link}
| Immune Activation | Neuropsychiatric Outcome |
|---|---|
| Maternal bacterial and viral infections [85] | Relationship between maternal infection and autism without specification on sex effect |
| Maternal and infant infections [86] | Relationship between maternal, infant infection and autism without specification on sex effect |
| Maternal early gestational C-reactive protein (CRP) [87] | Relation between maternal C-reactive protein (CRP) and risk of autism in both sexes |
| Children hospitalized for infection [88] | Stronger association of ASDs/infantile autism for girls compared with boys |
| Maternal autoimmune disorders [89] | No sex difference in ADHD prevalence |
| Maternal stress-infection interactions during pregnancy [90] | Association of maternal infection during the second trimester with depressive symptoms in adulthood for both sexes |
| Maternal cytokine levels [91] | Association TNF-α to IL-10 ratio in maternal serum with depression in a sex-dependent manner |
| Maternal herpes simplex virus 2 IgG level [92] | Association with schizophrenia without specification for sex effect |
| Maternal influenza infection [93] | Association with schizophrenia without specification for sex effect |
| Maternal influenza infection [94] | Association with schizophrenia without specification for sex effect |
| Maternal influenza infection [95] | Association with major depressive disorder without specification for sex effect |
| Maternal bacterial infection [96] | First-trimester infection, upper respiratory tract and gonococcal infections were associated with elevated risk of schizophrenia without specification for sex effect |
| Maternal influenza infection [97] | Association with schizophrenia without specification for sex effect |
| Maternal serum level of cytokines [98] | Schizophrenic males with high IL-6 levels and lower TNF-α levels among schizophrenic females |
| Level of cytokines in plasma samples from children diagnosed with ASD [99] | Negative correlation of ASD symptom severity with levels of IL-1β, IL-8, MIP-1β, and VEGF in females, but not in males |
| Genetic Background | Immune Activation | Behavioural Outcome | Neuropathological Outcome |
|---|---|---|---|
| Sprague Dawley rats [148] | Maternal LPS (Escherichia coli, 026:B6, 0.1 mg/kg) at E11 | Autistic-like behavior in male | Decreased glutamate, increased corticosterone levels within the hippocampus in males, Decreased levels of NR2B protein in both male and female |
| Cntnap2 KO and WT mice [149] | Maternal LPS (0.3 mg/kg) at E7 | Autistic-like behavior in male | Lower corticotropin-releasing hormone receptor-1 (Crhr1) gene expression in the left hippocampus of male mice |
| C57BL/6J mice [150] | Maternal LPS (Escherichia coli, O127:B8, 0.075 mg/kg) at both E11.5 and E12 or Poly I:C (20 mg/kg) at E12.5 | Autistic-like behavior (from both LPS and Poly I:C-treated mothers) | Not applied |
| C57/BL6 and BTBR mice [151] | Maternal Poly I:C (20 mg/kg) at E12.5 | Autistic-like behavior only in BTBR males | Not applied |
| C57/BL6 mice [152] | Maternal LPS (Escherichia coli, 0111:B4, 0.1 mg/kg) at E15 | Autistic-like behavior in male and female | Increased number of spines, reduced expression of the fractalkine microglial receptor (CX3CR1) in the hippocampus of only male |
| Wistar rats [153] | Maternal LPS (Escherichia coli, 026:B6, 1 mg/kg) every other day from E7 until birth | Schizophrenia-related behavioral phenotypes in both male and female | Decrease in the total number of PV- and GAD67-positive neurons in the medial prefrontal cortices of adult females, but only in the hippocampus of adult males |
| Wistar rats [154] | Maternal LPS (Escherichia coli, 026:B6, 1 mg/kg) every second day of pregnancy from E7 until birth | Schizophrenia-related behavioral phenotypes in both male and female | Elevated plasma level of corticosterone and a decrease in the glucocorticoid receptor level in the hippocampus and the glucocorticoid receptor-associated proteins FKBP51 concentration in the frontal cortex in both male and female |
| Swiss-Webster mice [155] | Maternal LPS (Escherichia coli, 0111:B4, 0.2 mg/kg) or Poly I:C (20 mg/kg) at E9 | Not applied | At PD28, increased GAD67 expression in the ventral stratum oriens in male mice prenatally treated with LPS, and in female mice prenatally treated with poly I:C |
| Wistar rats [156] | Maternal LPS (Escherichia coli, 0111:B4, 0.1 mg/kg) at E15 and E16 | Schizophrenia-related behavioral phenotypes in both male and female rats | Female rats showed weaker cognitive impairments, less demyelination and only a modest loss of PV expressing cells |
| Long-Evans rats [157] | Maternal Poly I:C (4 mg/kg) at E15 | Schizophrenia-related behavioral phenotypes (Male displayed significantly impaired object-in-place recognition memory; disturbed prepulse inhibition (PPI) in both sexes) | Not applied |
| Long-Evans rats [158] | Maternal Poly I:C (4 mg/kg) at E15 | Schizophrenia-related executive function that involve the prefrontal cortex (PFC) in male rats | Not applied |
| Sprague Dawley rats [159] | Neonatal LPS (Escherichia coli, 026:B6, 0.1 mg/kg) at P14 | No anhedonia after both, neonatal and 2-hit LPS challenges | Elevated expression of hypothalamic COX-2 in male after neonatal LPS |
| Sprague Dawley rats [160] | Maternal LPS (Escherichia coli, 055:B5, 0.5 mg/kg) at E19 | Impaired spatial recognition in males | Increased protein carbonylation, decreased a-Tocopherol and reduced/oxidized glutathione ratio, impaired NMDA synaptic currents and LTP in male hippocampus |
| C57BL/6 mice [161] | Maternal Poly I:C (5 mg/kg) at E17 | Schizophrenia-related behavioral phenotypes in both male and female mice | Decreased dopamine level in prefrontal cortex and ventral hippocampus in both male and female Male: reduced glutamate, aspartate, taurine in prefrontal cortex Female: decreased g-aminobutyric acid, taurine in dorsal hippocampus, increased 5-hydroxytryptamine levels in amygdala and nucleus accumbens |
| C57/Bl6:129 [162] | Maternal stress (E1–E7) | Stress induced locomotor hyperactivity in males (ADHD) | Both sexes: increased proapoptotic factor, Fas ligand in placenta Male: increased placental cytokines, decreased D2 receptor in nucleus accumbens, increased D1 receptor in prefrontal cortex Female: decreased CCL2, dopamin receptor levels were not analyzed |
| Wistar rats [163] | Maternal LPS (Escherichia coli, 026:B6, 0.1 mg/kg) at E15 | Reduced frequency of juvenile play (social) behavior in males | Reduced arginine vasopressin mRNA expression in the medial amygdala and bed nucleus of the stria terminalis only in males |
| Wistar rats [164] | Maternal LPS (Escherichia coli, 026:B6, 1 mg/kg LPS) on alternate days during pregnancy | Schizophrenia-related behavioral phenotypes in both male mice | Astroglial and microglial reactivity in different brain regions only in male Increased blood levels of IL-6, IL-2 in male and female |
| Swiss mice [165] | Neonatal LPS (Escherichia coli, 055:B5, 0.05 mg/kg) at P5 and P7 | Depressive like, risk-taking, anxiety-like, and repetitive behaviors with working memory deficits in male Prepulse inhibition (PPI) deficits in female | Increased levels of IL-4 (PFC, hippocampus) and decreased levels of IL-6 (PFC, hippocampus) and increased BDNF levels in both male and female |
| Wistar rats [166] | Maternal Poly I:C (4 mg/kg) at E15 | Not applied | Reduced hippocampal size in both male and female |
| Rhesus monkeys [167] | Maternal purified IgG, at E30, 44, 58, 72, 86, 100 | Autistic-Like Behavior in male and female | Significant higher total brain volume in the age 3 and 6 months of life only in male IgG-ASD37/73 kDa especially in frontal and occipital lobes |
| C57BL/6 mice [168] | Neonatal Poly I:C (10 mg/kg) at P8 | Not applied | Activation of caspase-dependent pathways is earlier in female mice than male mice Higher GFAP expression 14 h after injection only in male mice Persistence of microglial activation at 7 days after poly I:C only in female mice |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ardalan, M.; Chumak, T.; Vexler, Z.; Mallard, C. Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 2270. https://doi.org/10.3390/ijms20092270
Ardalan M, Chumak T, Vexler Z, Mallard C. Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders. International Journal of Molecular Sciences. 2019; 20(9):2270. https://doi.org/10.3390/ijms20092270
Chicago/Turabian StyleArdalan, Maryam, Tetyana Chumak, Zinaida Vexler, and Carina Mallard. 2019. "Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders" International Journal of Molecular Sciences 20, no. 9: 2270. https://doi.org/10.3390/ijms20092270
APA StyleArdalan, M., Chumak, T., Vexler, Z., & Mallard, C. (2019). Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders. International Journal of Molecular Sciences, 20(9), 2270. https://doi.org/10.3390/ijms20092270