Expression of Musashi-1 During Osteogenic Differentiation of Oral MSC: An In Vitro Study

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
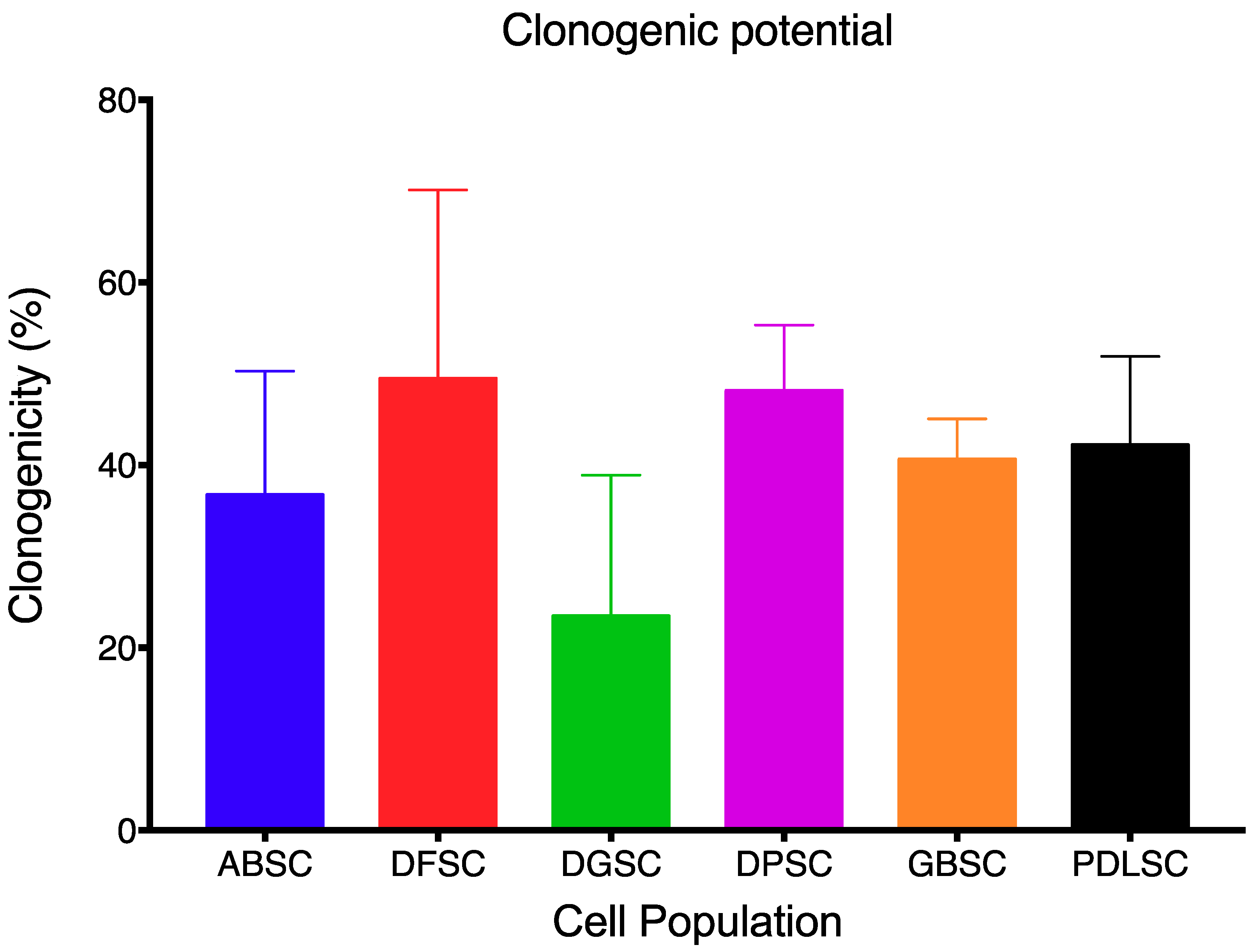
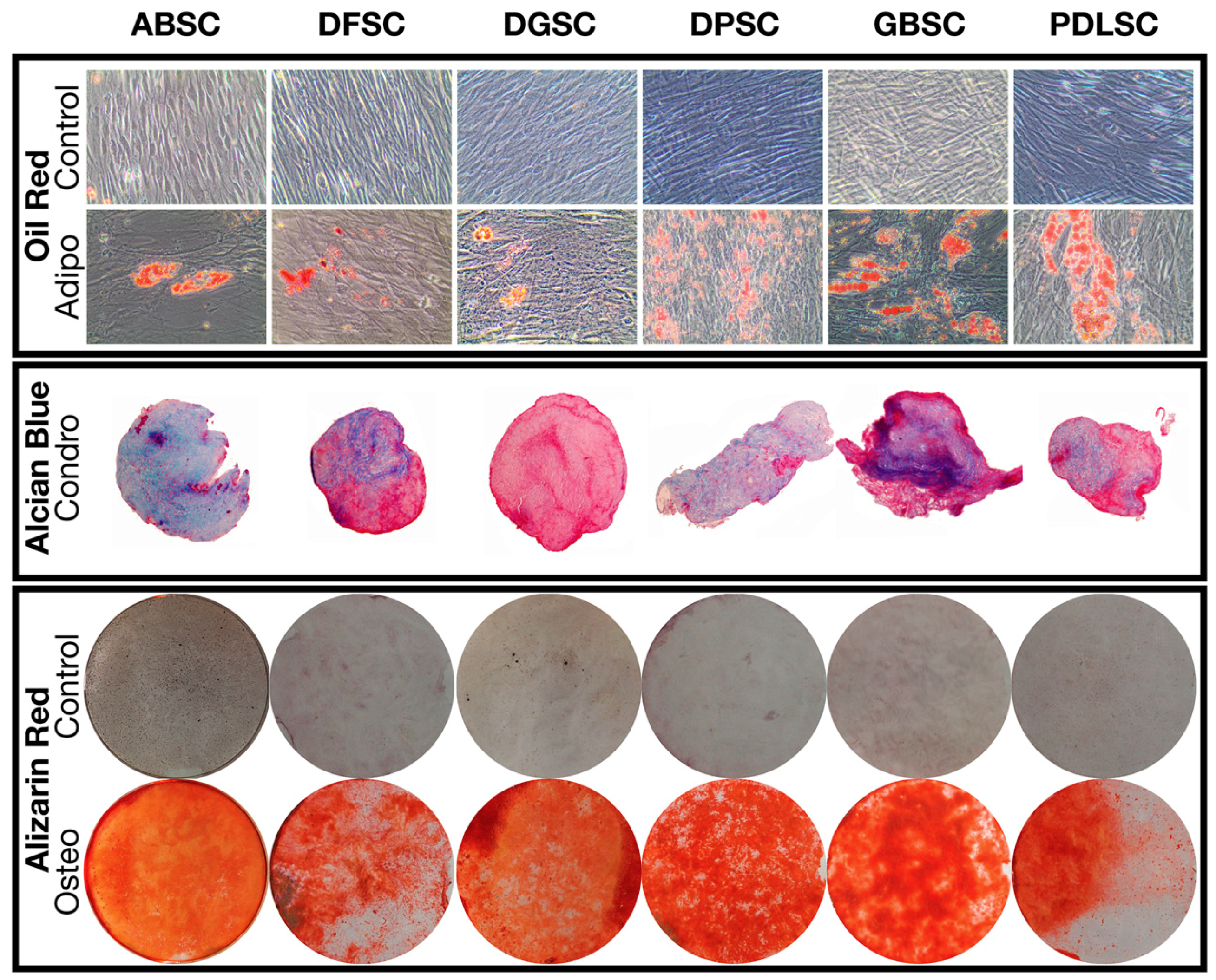
2.1. MSC Characterization
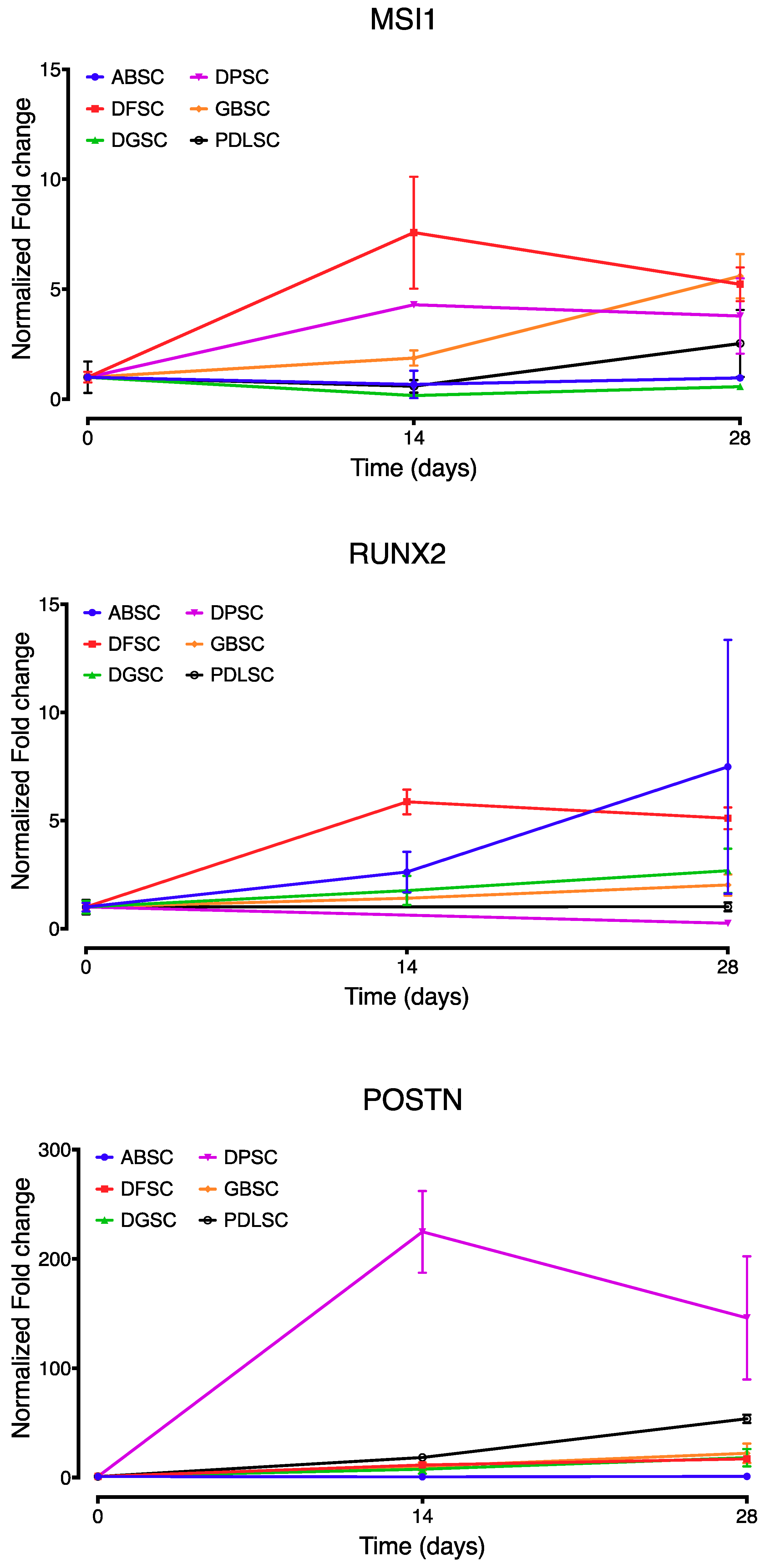
2.2. Co-Expression of MSI1, RUNX2, and POSTN During Osteogenic Differentiation
3. Discussion
4. Material and Methods
4.1. MSC Collection and Isolation
4.2. Population Doubling, Growth Kinetic and Senescence
4.3. Clonogenicity Potential
4.4. Phenotype
4.5. Stemness
4.6. Trilineage Differentiation Potential
4.6.1. Adipogenic Differentiation
4.6.2. Chondrogenic Differentiation
4.6.3. Osteogenic Differentiation
4.7. Temporal Expression of Musashi-1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical aspects
References
- Nakamura, M.; Okano, H.; Blendy, J.A.; Montell, C. Musashi, a neural RNA-binding protein required for drosophila adult external sensory organ development. Neuron 1994, 13, 67–81. [Google Scholar] [CrossRef]
- Sakakibara, S.-I.; Nakamura, Y.; Yoshida, T.; Shibata, S.; Koike, M.; Takano, H.; Ueda, S.; Uchiyama, Y.; Noda, T.; Okano, H. RNA-binding protein Musashi family: Roles for CNS stem cells and a subpopulation of ependymal cells revealed by targeted disruption and antisense ablation. Proc. Natl. Acad. Sci. USA 2002, 99, 15194–15199. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Kanno, R.; Kurihara, Y.; Uesugi, S.; Imai, T.; Sakakibara, S.; Okano, H.; Katahira, M. Structure, backbone dynamics and interactions with RNA of the C-terminal RNA-binding domain of a mouse neural RNA-binding protein, Musashi1. J. Mol. Biol. 1999, 287, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Horisawa, K.; Imai, T.; Okano, H.; Yanagawa, H. The Musashi family RNA-binding proteins in stem cells. Biomol. Concepts 2010, 1, 59–66. [Google Scholar] [CrossRef]
- Imai, T.; Tokunaga, A.; Yoshida, T.; Hashimoto, M.; Mikoshiba, K.; Weinmaster, G.; Nakafuku, M.; Okano, H. The Neural RNA-Binding Protein Musashi1 Translationally Regulates Mammalian numb Gene Expression by Interacting with Its mRNA. Mol. Cell. Biol. 2001, 21, 3888–3900. [Google Scholar] [CrossRef] [PubMed]
- Ratti, A. A role for the ELAV RNA-binding proteins in neural stem cells: Stabilization of Msi1 mRNA. J. Cell Sci. 2006, 119, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- O’Valle, F.; de Buitrago, J.G.; Hernández-Cortés, P.; Padial-Molina, M.; Crespo-Lora, V.; Cobo, M.; Aguilar, D.; Galindo-Moreno, P. Increased expression of Musashi-1 evidences mesenchymal repair in maxillary sinus floor elevation. Sci. Rep. 2018, 8, 12243. [Google Scholar] [CrossRef]
- Carbonare, L.D.; Innamorati, G.; Valenti, M.T. Transcription Factor Runx2 and its Application to Bone Tissue Engineering. Stem Cell Rev. Rep. 2012, 8, 891–897. [Google Scholar] [CrossRef]
- Farias, V.A.; Linares-Fernández, J.L.; Peñalver, J.L.; Payá Colmenero, J.A.; Ferrón, G.O.; Duran, E.L.; Fernández, R.M.; Olivares, E.G.; O’Valle, F.; Puertas, A.; et al. Human umbilical cord stromal stem cell express CD10 and exert contractile properties. Placenta 2011, 32, 86–95. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and invivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Mark Bartold, P.; Batouli, S.; Brahim, J.; Young, M.; Gehron Robey, P.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Padial-Molina, M.; O’Valle, F.; Lanis, A.; Mesa, F.; Dohan Ehrenfest, D.M.; Wang, H.-L.; Galindo-Moreno, P. Clinical Application of Mesenchymal Stem Cells and Novel Supportive Therapies for Oral Bone Regeneration. BioMed Res. Int. 2015, 2015, 1–16. [Google Scholar] [CrossRef]
- Padial-Molina, M.; Marchesan, J.T.; Taut, A.D.; Jin, Q.; Giannobile, W.V.; Rios, H.F. Methods to validate tooth-supporting regenerative therapies. Methods Mol. Biol. 2012, 887, 135–148. [Google Scholar] [PubMed]
- Kaigler, D.; Avila-Ortiz, G.; Travan, S.; Taut, A.D.; Padial-Molina, M.; Rudek, I.; Wang, F.; Lanis, A.; Giannobile, W.V. Bone Engineering of Maxillary Sinus Bone Deficiencies Using Enriched CD90+ Stem Cell Therapy: A Randomized Clinical Trial. J. Bone Miner. Res. 2015, 30, 1206–1216. [Google Scholar] [CrossRef] [PubMed]
- Sonoyama, W.; Liu, Y.; Fang, D.; Yamaza, T.; Seo, B.-M.; Zhang, C.; Liu, H.; Gronthos, S.; Wang, C.-Y.; Shi, S.; et al. Mesenchymal Stem Cell-Mediated Functional Tooth Regeneration in Swine. PLoS ONE 2006, 1, 79. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, L.F.; Kingham, P.J.; Wiberg, M.; Kelk, P. In Vitro Osteogenic Differentiation of Human Mesenchymal Stem Cells from Jawbone Compared with Dental Tissue. Tissue Eng. Regen. Med. 2017, 14, 763–774. [Google Scholar] [CrossRef]
- Fox, R.G.; Park, F.D.; Koechlein, C.S.; Kritzik, M.; Reya, T. Musashi Signaling in Stem Cells and Cancer. Annu. Rev. Cell Dev. Biol. 2015, 31, 249–267. [Google Scholar] [CrossRef]
- Hong, I.S.; Kang, K.S. The Effects of Hedgehog on the RNA-Binding Protein Msi1 in the Proliferation and Apoptosis of Mesenchymal Stem Cells. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Niu, J.; Zhao, X.; Liu, Q.; Yang, J. Knockdown of MSI1 inhibited the cell proliferation of human osteosarcoma cells by targeting p21 and p27. Oncol. Lett. 2017, 14, 5271–5278. [Google Scholar] [CrossRef]
- Hong, I.S.; Lee, H.Y.; Choi, S.W.; Kim, H.S.; Yu, K.R.; Seo, Y.; Jung, J.W.; Kang, K.S. The effects of hedgehog on RNA binding protein Msi1 during the osteogenic differentiation of human cord blood-derived mesenchymal stem cells. Bone 2013, 56, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Magan-Fernandez, A.; Fernandez-Barbero, J.E.; O’Valle, F.; Ortiz, R.; Galindo-Moreno, P.; Mesa, F. Simvastatin exerts antiproliferative and differentiating effects on MG63 osteoblast-like cells: Morphological and immunocytochemical study. J. Periodontal Res. 2018, 53, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Kidwai, F.K.; Kopher, R.A.; Motl, J.; Kellum, C.A.; Westendorf, J.J.; Kaufman, D.S. Use of RUNX2 Expression to Identify Osteogenic Progenitor Cells Derived from Human Embryonic Stem Cells. Stem Cell Rep. 2015, 4, 190–198. [Google Scholar] [CrossRef]
- Olaechea, A.; Mendoza-Azpur, G.; O′Valle, F.; Padial-Molina, M.; Martin-Morales, N.; Galindo-Moreno, P. Biphasic hydroxyapatite and ß-Tricalcium phosphate biomaterial behavior in a case-series of maxillary sinus augmentation in humans. Clin. Oral Implant. Res. 2019, 30, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Flichy-Fernández, A.J.; Blaya-Tárraga, J.A.; O’Valle, F.; Padial-Molina, M.; Peñarrocha-Diago, M.; Galindo-Moreno, P. Sinus Floor Elevation Using Particulate PLGA-Coated Biphasic Calcium Phosphate Bone Graft Substitutes: A Prospective Histological and Radiological Study. Clin. Implant Dent. Relat. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Moreno, P.; de Buitrago, J.G.; Padial-Molina, M.; Fernández-Barbero, J.E.; Ata-Ali, J.; O′Valle, F. Histopathological comparison of healing after maxillary sinus augmentation using xenograft mixed with autogenous bone versus allograft mixed with autogenous bone. Clin. Oral Implant. Res. 2018, 29, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Rosselli-Murai, L.K.; Almeida, L.O.; Zagni, C.; Galindo-Moreno, P.; Padial-Molina, M.; Volk, S.L.; Murai, M.J.; Rios, H.F.; Squarize, C.H.; Castilho, R.M. Periostin responds to mechanical stress and tension by activating the MTOR signaling pathway. PLoS ONE 2013, 8, e83580. [Google Scholar] [CrossRef]
- Wu, Z.; Dai, W.; Wang, P.; Zhang, X.; Tang, Y.; Liu, L.; Wang, Q.; Li, M.; Tang, C. Periostin promotes migration, proliferation, and differentiation of human periodontal ligament mesenchymal stem cells. Connect. Tissue Res. 2018, 59, 108–119. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, L.; Wang, P.; Chen, D.; Wu, Z.; Tang, C. Periostin promotes migration and osteogenic differentiation of human periodontal ligament mesenchymal stem cells via the Jun amino-terminal kinases (JNK) pathway under inflammatory conditions. Cell Prolif. 2017, 50, 12369. [Google Scholar] [CrossRef]
- Cha, Y.; Jeon, M.; Lee, H.S.; Kim, S.; Kim, S.O.; Lee, J.H.; Song, J.S. Effects of In Vitro Osteogenic Induction on In Vivo Tissue Regeneration by Dental Pulp and Periodontal Ligament Stem Cells. J. Endod. 2015, 41, 1462–1468. [Google Scholar] [CrossRef]
- Padial-Molina, M.; Volk, S.L.; Rios, H.F. Preliminary insight into the periostin leverage during periodontal tissue healing. J. Clin. Periodontol. 2015, 42, 764–772. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ABSC | DFSC | DGSC | DPSC | GBSC | PDLSC | |
|---|---|---|---|---|---|---|
| Best-fit values | ||||||
| B0 | 1.142 | −0.8773 | 0.6326 | 0.3618 | 0.8496 | −0.8511 |
| B1 | 0.5771 | 0.7993 | 0.6278 | 0.837 | 0.5941 | 0.7096 |
| B2 | −0.003228 | −0.004974 | −0.002007 | −0.004444 | −0.00446 | −0.002893 |
| CPD | 27.50 ± 4.36 | 32.86 ± 0.35 | 39.04 ± 12.48 | 42.45 ± 0.34 | 20.85 ± 4.24 | 37.52 ± 10.94 |
| Number of passages | 14.40 ± 0.52 | 13.13 ± 0.99 | 15.00 ± 1.10 | 14.55 ± 1.29 | 13.00 ± 0.85 | 16.00 ± 3.27 |
| ABSC | % Alive | 82.40% | ||||
| CD73/CD90/CD105 | 99.69% | 97.90% | 98.80% | |||
| CD34/CD45/CD14/CD31 | 1.79% | 1.28% | 0.03% | 1.31% | ||
| DFSC | % Alive | 71.60% | ||||
| CD73/CD90/CD105 | 99.92% | 99.16% | 97.38% | |||
| CD34/CD45/CD14/CD31 | 0.99% | 1.83% | 0.02% | 0.05% | ||
| DGSC | % Alive | 95.40% | ||||
| CD73/CD90/CD105 | 99.90% | 84.30% | 66.90% | |||
| CD34/CD45/CD14/CD31 | 2.31% | 0.31% | 0.11% | 0.05% | ||
| DPSC | % Alive | 75.30% | ||||
| CD73/CD90/CD105 | 99.95% | 96.86% | 96.75% | |||
| CD34/CD45/CD14/CD31 | 0.88% | 1.12% | 0.02% | 0.06% | ||
| GBSC | % Alive | 90.00% | ||||
| CD73/CD90/CD105 | 99.78% | 96.37% | 98.40% | |||
| CD34/CD45/CD14/CD31 | 0.45% | 1.12% | 0% | 1.05% | ||
| PDLSC | % Alive | 88.30% | ||||
| CD73/CD90/CD105 | 99.61% | 99.80% | 98.69% | |||
| CD34/CD45/CD14/CD31 | 0.43% | 0.4% | 0% | 0.09% | ||
| CONTROL | OSTEO | p Value (T-Test) | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| ABSC | 1.000 | 0.027 | 9.926 | 1.448 | 0.001 |
| DFSC | 1.000 | 0.069 | 10.817 | 0.761 | 0.000 |
| DGSC | 1.000 | 0.050 | 10.427 | 0.598 | 0.000 |
| DPSC | 1.000 | 0.049 | 31.111 | 4.955 | 0.001 |
| GBSC | 1.000 | 0.060 | 39.197 | 1.176 | 0.000 |
| PDLSC | 1.000 | 0.041 | 3.498 | 0.227 | 0.000 |
| CONTROL | OSTEO | p Value (T-Test) | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| ABSC | 1.000 | 0.073 | 1.098 | 0.283 | 0.681 |
| DFSC | 1.000 | 0.243 | 1.030 | 0.201 | 0.906 |
| DGSC | 1.000 | 0.290 | 3.531 | 0.173 | 0.008 |
| DPSC | 1.000 | 0.098 | 2.462 | 0.241 | 0.015 |
| GBSC | 1.000 | 0.049 | 2.737 | 0.268 | 0.012 |
| PDLSC | 1.000 | 0.147 | 28.125 | 6.824 | 0.030 |
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| MYC | GGAGATCCGGAGCGAATAGGG | GTTTCGTGGATGCGGCAAGG |
| NANOG | TGAATCCTTCCTCTCCCCTCC | CCTCGCTGATTAGGCTCCAAC |
| POU5F1 | AGGCCCGAAAGAGAAAGCGA | CTGATCTGCTGCAGTGTGGGT |
| SOX2 | TCAGGAGTTGTCAAGGCAGAG | CCGCCGCCGATGATTGTT |
| MSI1 | TGAGCAGTTTGGGAAGGTG | TCACACACTTTCTCCACGATG |
| POSTN | TTTCTACTGGAGGTGGAGAAAC | GTGACCTTGGTGACCTCTTC |
| SPARC | AGAACAACACCCCCATGTGCGT | TCCAGGGTGCACTTTGTGGCAA |
| RUNX2 | ACCGTCTTCACAAATCCTCCC | AGCTTCTGTCTGTGCCTTCTG |
| GAPDH | AGCTCATTTCCTGGTATGACAAC | TTACTCCTTGGAGGCCATGTG |
| RPLP0 | CAGATTGGCTACCCAACTGTT | GGCCAGGACTCGTTTGTACC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padial-Molina, M.; de Buitrago, J.G.; Sainz-Urruela, R.; Abril-Garcia, D.; Anderson, P.; O’Valle, F.; Galindo-Moreno, P. Expression of Musashi-1 During Osteogenic Differentiation of Oral MSC: An In Vitro Study. Int. J. Mol. Sci. 2019, 20, 2171. https://doi.org/10.3390/ijms20092171
Padial-Molina M, de Buitrago JG, Sainz-Urruela R, Abril-Garcia D, Anderson P, O’Valle F, Galindo-Moreno P. Expression of Musashi-1 During Osteogenic Differentiation of Oral MSC: An In Vitro Study. International Journal of Molecular Sciences. 2019; 20(9):2171. https://doi.org/10.3390/ijms20092171
Chicago/Turabian StylePadial-Molina, Miguel, Juan G. de Buitrago, Raquel Sainz-Urruela, Dario Abril-Garcia, Per Anderson, Francisco O’Valle, and Pablo Galindo-Moreno. 2019. "Expression of Musashi-1 During Osteogenic Differentiation of Oral MSC: An In Vitro Study" International Journal of Molecular Sciences 20, no. 9: 2171. https://doi.org/10.3390/ijms20092171
APA StylePadial-Molina, M., de Buitrago, J. G., Sainz-Urruela, R., Abril-Garcia, D., Anderson, P., O’Valle, F., & Galindo-Moreno, P. (2019). Expression of Musashi-1 During Osteogenic Differentiation of Oral MSC: An In Vitro Study. International Journal of Molecular Sciences, 20(9), 2171. https://doi.org/10.3390/ijms20092171