Acute Effects of Single Doses of Bonito Fish Peptides and Vitamin D on Whole Blood Gene Expression Levels: A Randomized Controlled Trial

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Participant Baseline Characteristics
2.2. Changes in Glucose, Insulin, C-Peptide, and TG during Post-Oral Glucose Tolerance Test (OGTT)
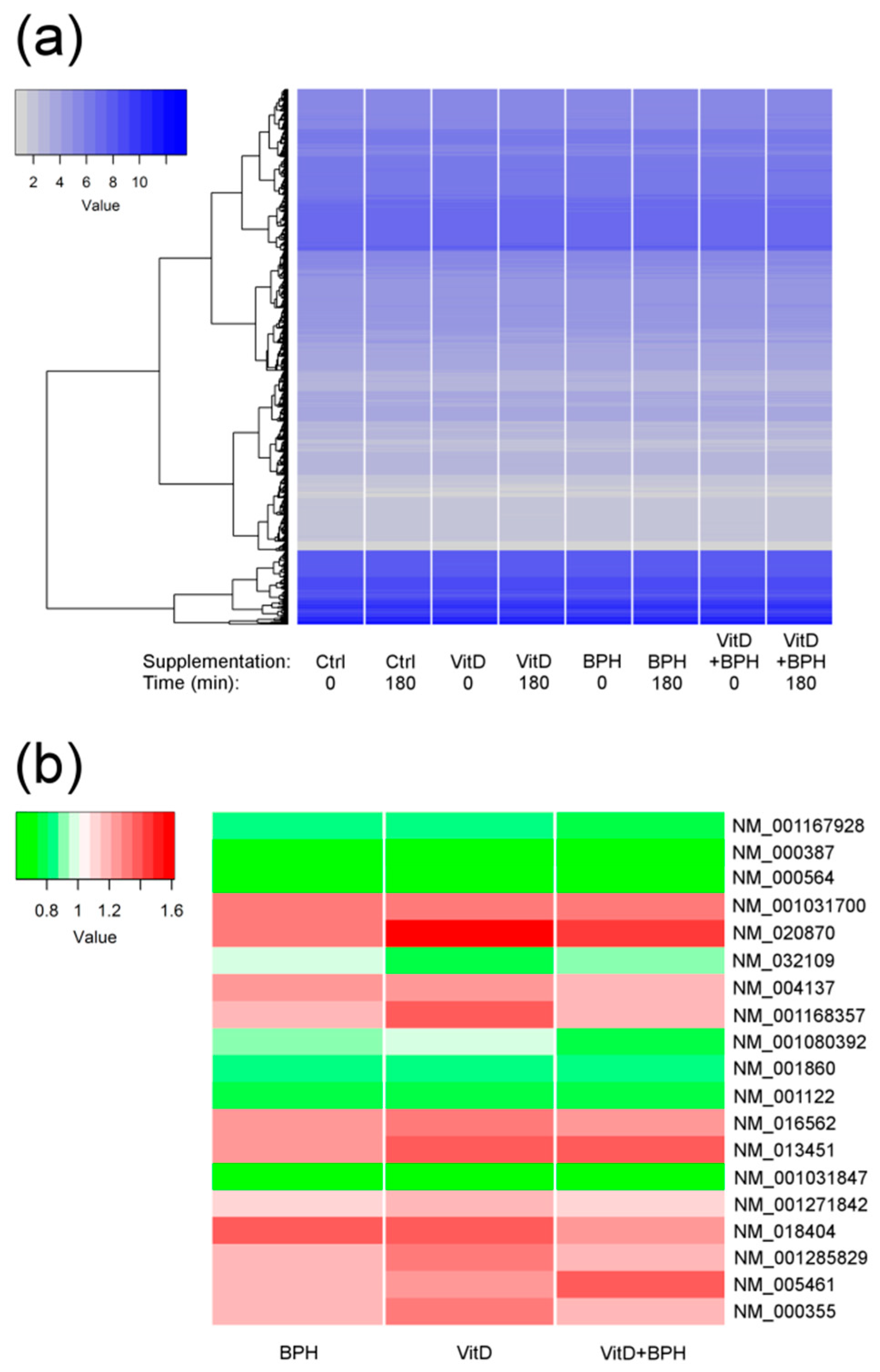
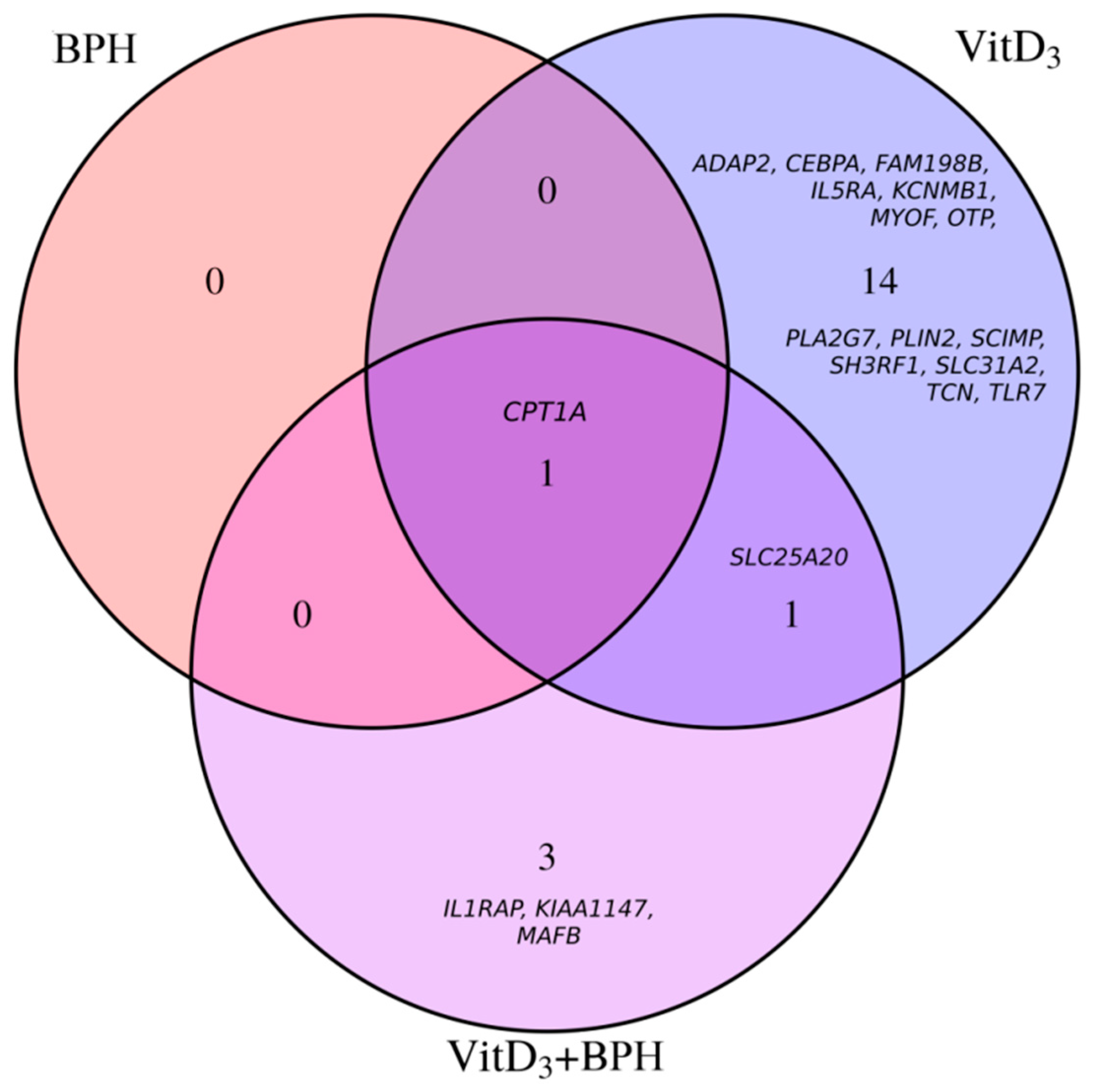
2.3. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Study Design
4.3. VitD3 and BPH Treatment
4.4. Blood Sampling and Biochemical Measures
4.5. Gene Expression Analysis
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 25(OH)D | 25-hydroxyvitamin D |
| ANOVA | Analysis of variance |
| AUC | Areas under the curve |
| BMI | Body mass index |
| BPH | Bonito fish peptide hydrolysate |
| C | Cholesterol |
| CACT | Carnitine acylcarnitine translocase |
| CEBPA | CCAAT enhancer binding protein alpha |
| CPT1A | Carnitine palmitoyltransferase 1A |
| CRP | C-reactive protein |
| CVD | Cardiovascular disease |
| DBP | Diastolic blood pressure |
| DHA | Docosahexaenoic acid |
| EPA | Eicosapentaenoic acid |
| FDR | False discovery rate |
| HDL-C | High-density lipoprotein cholesterol |
| iAUC | Incremental areas under the curve |
| INAF | Institute of Nutrition and Functional Food |
| LDL-C | Low-density lipoprotein cholesterol |
| OGTT | Oral glucose tolerance test |
| PLA2G7 | Phospholipase A2 group VII |
| PLIN2 | Perilipin 2 |
| RCT | Randomized controlled trial |
| SBP | Systolic blood pressure |
| SCIMP | SLP adaptor and CSK interacting membrane protein |
| SLC25A20 | Solute carrier family 25 member 20 |
| T2D | Type 2 diabetes |
| TG | Triglyceride |
| TF | Transcription factor |
| Total-C | Total cholesterol |
| VitD3 | cholecalciferol |
References
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.O.; Dyerberg, J.; Nielsen, A.B. Plasma Lipid and Lipoprotein Pattern in Greelandic West-Coast Eskimos. Nutr. Rev. 1986, 44, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.O.; Dyerberg, J.; Sinclair, H.M. The composition of the Eskimo food in north Western Greenland. Am. J. Clin. Nutr. 1980, 33, 2657–2661. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.M.; Hennekens, C.H.; O’Donnell, C.J.; Ajani, U.A.; Carey, V.J.; Willett, W.C.; Ruskin, J.N.; Manson, J.A.E. Fish consumption and risk of sudden cardiac death. J. Am. Med. Assoc. 1998, 279, 23–28. [Google Scholar] [CrossRef]
- Zhao, L.-G.; Sun, J.-W.; Yang, Y.; Ma, X.; Wang, Y.-Y.; Xiang, Y.-B. Fish consumption and all-cause mortality: A meta-analysis of cohort studies. Eur. J. Clin. Nutr. 2016, 70, 155–161. [Google Scholar] [CrossRef]
- Jayedi, A.; Shab-Bidar, S.; Eimeri, S.; Djafarian, K. Fish consumption and risk of all-cause and cardiovascular mortality: A dose-response meta-analysis of prospective observational studies. Public Health Nutr. 2018, 21, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Wallin, A.; Di Giuseppe, D.; Orsini, N.; Patel, P.S.; Forouhi, N.G.; Wolk, A. Fish consumption, dietary long-chain n-3 fatty acids, and risk of type 2 diabetes: Systematic review and meta-analysis of prospective studies. Diabetes Care 2012, 35, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, C.; Botella-Carretero, J.I.; Corella, D.; Fiol, M.; Lage, M.; Lurbe, E.; Richart, C.; Fernández-Real, J.M.; Fuentes, F.; Ordóñez, A.; et al. White fish reduces cardiovascular risk factors in patients with metabolic syndrome: The WISH-CARE study, a multicenter randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, G.; Busnelli, M.; Manzini, S.; Parolini, C. Nutraceuticals and bioactive components from fish for dyslipidemia and cardiovascular risk reduction. Mar. Drugs 2016, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; McLerran, D.F.; Rolland, B.; Chen, Y.; Grant, E.J.; Vedanthan, R.; Inoue, M.; Tsugane, S.; Gao, Y.T.; Tsuji, I.; et al. Meat intake and cause-specific mortality: A pooled analysis of Asian prospective cohort studies. Am. J. Clin. Nutr. 2013, 98, 1032–1041. [Google Scholar] [CrossRef]
- Wang, M.P.; Thomas, G.N.; Ho, S.Y.; Lai, H.K.; Mak, K.H.; Lam, T.H. Fish Consumption and Mortality in Hong Kong Chinese-the LIMOR Study. Ann. Epidemiol. 2011, 21, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Osler, M.; Andreasen, A.H.; Hoidrup, S. No inverse association between fish consumption and risk of death from all-causes, and incidence of coronary heart disease in middle-aged, Danish adults. J. Clin. Epidemiol. 2003, 56, 274–279. [Google Scholar] [CrossRef]
- Olsen, A.; Egeberg, R.; Halkjær, J.; Christensen, J.; Overvad, K.; Tjønneland, A. Healthy aspects of the Nordic diet are related to lower total mortality. J. Nutr. 2011, 141, 639–644. [Google Scholar] [CrossRef]
- Ouellet, V.; Weisnagel, S.J.; Marois, J.; Bergeron, J.; Julien, P.; Gougeon, R.; Tchernof, A.; Holub, B.J.; Jacques, H. Dietary Cod Protein Reduces Plasma C-Reactive Protein in Insulin-Resistant Men and Women1. J. Nutr. 2008, 138, 2386–2391. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, V.; Marois, J.; Weisnagel, S.J.; Jacques, H. Dietary cod protein improves insulin sensitivity in insulin-resistant men and women: A randomized controlled trial. Diabetes Care 2007, 30, 2816–2821. [Google Scholar] [CrossRef]
- Vikoren, L.A.; Nygård, O.K.; Lied, E.; Rostrup, E.; Gudbrandsen, O.A. A randomised study on the effects of fish protein supplement on glucose tolerance, lipids and body composition in overweight adults. Br. J. Nutr. 2013, 109, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Watanabe, K.; Ma, M.; Hirayama, M.; Kobayashi, T.; Oyama, H.; Sakaguchi, Y.; Kanda, M.; Kodama, M.; Aizawa, Y. The Effects of γ-Aminobutyric Acid, Vinegar, and Dried Bonito on Blood Pressure in Normotensive and Mildly or Moderately Hypertensive Volunteers. J. Clin. Biochem. Nutr. 2009, 45, 93–100. [Google Scholar] [CrossRef]
- Tremblay, F.; Lavigne, C.; Jacques, H.; Marette, A. Role of Dietary Proteins and Amino Acids in the Pathogenesis of Insulin Resistance. Annu. Rev. Nutr. 2007, 27, 293–310. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, D.; Anderson, G.H. Recent advances in dietary proteins and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Chevrier, G.; Mitchell, P.L.; Rioux, L.-E.; Hasan, F.; Jin, T.; Roblet, C.R.; Doyen, A.; Pilon, G.; St-Pierre, P.; Lavigne, C.; et al. Low-Molecular-Weight Peptides from Salmon Protein Prevent Obesity-Linked Glucose Intolerance, Inflammation, and Dyslipidemia in LDLR-/-/ApoB100/100 Mice. J. Nutr. 2015, 145, 1415–1422. [Google Scholar] [CrossRef]
- Pilon, G.; Ruzzin, J.; Rioux, L.E.; Lavigne, C.; White, P.J.; Frøyland, L.; Jacques, H.; Bryl, P.; Beaulieu, L.; Marette, A. Differential effects of various fish proteins in altering body weight, adiposity, inflammatory status, and insulin sensitivity in high-fat-fed rats. Metabolism 2011, 60, 1122–1130. [Google Scholar] [CrossRef]
- Pilz, S.; Verheyen, N.; Grübler, M.R.; Tomaschitz, A.; März, W. Vitamin D and cardiovascular disease prevention. Nat. Rev. Cardiol. 2016, 13, 404–417. [Google Scholar] [CrossRef]
- Jorde, R.; Grimnes, G. Vitamin D and metabolic health with special reference to the effect of vitamin D on serum lipids. Prog. Lipid Res. 2011, 50, 303–312. [Google Scholar] [CrossRef]
- Gagnon, C.; Lu, Z.X.; Magliano, D.J.; Dunstan, D.W.; Shaw, J.E.; Zimmet, P.Z.; Sikaris, K.; Grantham, N.; Ebeling, P.R.; Daly, R.M. Serum 25-Hydroxyvitamin D, Calcium Intake, and Risk of Type 2 Diabetes After 5 Years. Diabetes Care 2011, 34, 1133–1138. [Google Scholar] [CrossRef]
- Gagnon, C.; Lu, Z.X.; Magliano, D.J.; Dunstan, D.W.; Shaw, J.E.; Zimmet, P.Z.; Sikaris, K.; Ebeling, P.R.; Daly, R.M. Low serum 25-hydroxyvitamin D is associated with increased risk of the development of the metabolic syndrome at five years: Results from a national, population-based prospective study (The Australian Diabetes, Obesity and Lifestyle Study: AusDiab). J. Clin. Endocrinol. Metab. 2012, 97, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Prevention and Management of Osteoporosis: Report of a WHO Scientific Group; World Health Organization: Geneva, Switzerland, 2003; ISBN 9241209216. Available online: https://apps.who.int/iris/bitstream/handle/10665/42841/WHO_TRS_921.pdf (accessed on 18 December 2018).
- Pilz, S.; Gaksch, M.; Kienreich, K.; Grübler, M.; Verheyen, N.; Fahrleitner-Pammer, A.; Treiber, G.; Drechsler, C.; Ó Hartaigh, B.; Obermayer-Pietsch, B.; et al. Effects of Vitamin D on Blood Pressure and Cardiovascular Risk Factors: A Randomized Controlled Trial. Hypertension 2015, 65, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Ponda, M.P.; Dowd, K.; Finkielstein, D.; Holt, P.R.; Breslow, J.L. The short-term effects of vitamin d repletion on cholesterol: A randomized, placebo-controlled trial. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2510–2515. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xia, N.; Yang, Y.; Peng, D.-Q. Influence of vitamin D supplementation on plasma lipid profiles: A meta-analysis of randomized controlled trials. Lipids Health Dis. 2012, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Z.B.; Tanisawa, K.; Ito, T.; Oshima, S.; Higuchi, M. Vitamin D supplementation reduces insulin resistance in Japanese adults: A secondary analysis of a double-blind, randomized, placebo-controlled trial. Nutr. Res. 2016, 36, 1121–1129. [Google Scholar] [CrossRef]
- El Hajj, C.; Chardigny, J.M.; Boirie, Y.; Yammine, K.; Helou, M.; Walrand, S. Effect of Vitamin D Treatment on Glucose Homeostasis and Metabolism in Lebanese Older Adults: A Randomized Controlled Trial. J. Nutr. Heal. Aging 2018, 22, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Gulseth, H.L.; Wium, C.; Angel, K.; Eriksen, E.F.; Birkeland, K.I. Effects of Vitamin D supplementation on insulin sensitivity and insulin secretion in subjectswith type 2 diabetes and Vitamin D deficiency: A randomized controlled trial. Diabetes Care 2017, 40, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Muscogiuri, G.; Annweiler, C.; Duval, G.; Karras, S.; Tirabassi, G.; Salvio, G.; Balercia, G.; Kimball, S.; Kotsa, K.; Mascitelli, L.; et al. Vitamin D and cardiovascular disease: From atherosclerosis to myocardial infarction and stroke. Int. J. Cardiol. 2017, 230, 577–584. [Google Scholar] [CrossRef]
- Berlanga-Taylor, A.J.; Plant, K.; Dahl, A.; Lau, E.; Hill, M.; Sims, D.; Heger, A.; Emberson, J.; Armitage, J.; Clarke, R.; et al. Genomic Response to Vitamin D Supplementation in the Setting of a Randomized, Placebo-Controlled Trial. EBioMedicine 2018. [Google Scholar] [CrossRef] [PubMed]
- Einarsdottir, E.; Pekkinen, M.; Krjutskov, K.; Katayama, S.; Kere, J.; Makitie, O.; Viljakainen, H. A preliminary transcriptome analysis suggests a transitory effect of vitamin D on mitochondrial function in obese young Finnish subjects. Endocr. Connect. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hossein-nezhad, A.; Spira, A.; Holick, M.F. Influence of Vitamin D Status and Vitamin D3 Supplementation on Genome Wide Expression of White Blood Cells: A Randomized Double-Blind Clinical Trial. PLoS ONE 2013, 8, e58725. [Google Scholar] [CrossRef] [PubMed]
- Pasing, Y.; Fenton, C.G.; Jorde, R.; Paulssen, R.H. Changes in the human transcriptome upon vitamin D supplementation. J. Steroid Biochem. Mol. Biol. 2017, 173, 93–99. [Google Scholar] [CrossRef]
- Punthakee, Z.; Goldenberg, R.; Katz, P. Definition, Classification and Diagnosis of Diabetes, Prediabetes and Metabolic Syndrome. Can. J. Diabetes 2018, 42, S10–S15. [Google Scholar] [CrossRef]
- Schwetz, V.; Scharnagl, H.; Trummer, C.; Stojakovic, T.; Pandis, M.; Grübler, M.R.; Verheyen, N.; Gaksch, M.; Zittermann, A.; Aberer, F.; et al. Vitamin D supplementation and lipoprotein metabolism: A randomized controlled trial. J. Clin. Lipidol. 2018, 12, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Jafari, T.; Fallah, A.A.; Barani, A. Effects of vitamin D on serum lipid profile in patients with type 2 diabetes: A meta-analysis of randomized controlled trials. Clin. Nutr. 2016, 35, 1259–1268. [Google Scholar] [CrossRef]
- Larriba, M.J.; González-Sancho, J.M.; Bonilla, F.; Muñoz, A. Interaction of vitamin D with membrane-based signaling pathways. Front. Physiol. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Drynan, L.; Quant, P.A.; Zammit, V.A. Flux control exerted by mitochondrial outer membrane carnitine palmitoyltransferase over beta-oxidation, ketogenesis and tricarboxylic acid cycle activity in hepatocytes isolated from rats in different metabolic states. Biochem.J. 1996, 317, 791–795. [Google Scholar] [CrossRef]
- Bonnefont, J.P.; Djouadi, F.; Prip-Buus, C.; Gobin, S.; Munnich, A.; Bastin, J. Carnitine palmitoyltransferases 1 and 2: Biochemical, molecular and medical aspects. Mol. Aspects Med. 2004, 25, 495–520. [Google Scholar] [CrossRef] [PubMed]
- Caimari, A.; Oliver, P.; Rodenburg, W.; Keijer, J.; Palou, A. Slc27a2 expression in peripheral blood mononuclear cells as a molecular marker for overweight development. Int. J. Obes. 2010, 34, 831–839. [Google Scholar] [CrossRef]
- Reynés, B.; García-Ruiz, E.; Palou, A.; Oliver, P. The intake of high-fat diets induces an obesogenic-like gene expression profile in peripheral blood mononuclear cells, which is reverted by dieting. Br. J. Nutr. 2016, 115, 1887–1895. [Google Scholar] [CrossRef] [PubMed]
- Saneei, P.; Salehi-Abargouei, A.; Esmaillzadeh, A. Serum 25-hydroxy vitamin D levels in relation to body mass index: A systematic review and meta-analysis. Obes. Rev. 2013, 14, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Neme, A.; Seuter, S.; Malinen, M.; Nurmi, T.; Tuomainen, T.P.; Virtanen, J.K.; Carlberg, C. In vivo transcriptome changes of human white blood cells in response to vitamin D. J. Steroid Biochem. Mol. Biol. 2019, 188, 71–76. [Google Scholar] [CrossRef]
- Larrick, B.M.; Kim, K.H.; Donkin, S.S.; Teegarden, D. 1,25-Dihydroxyvitamin D regulates lipid metabolism and glucose utilization in differentiated 3T3-L1 adipocytes. Nutr. Res. 2018, 58, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.K.F.; Brandão-Lima, P.N.; Tete, R.M.D.D.; Freire, A.R.S.; Pires, L.V. Vitamin D ratio and glycaemic control in individuals with type 2 diabetes mellitus: A systematic review. Diabetes. Metab. Res. Rev. 2018, 34, e2969. [Google Scholar] [CrossRef]
- Sulaiman Al-Shoumera, K.A.A.; Al-Asoosib, A.A.; Ali, A.H.; Nair, V.S. Does insulin resistance in type 2 diabetes alter vitamin D status? Prim. Care Diabetes 2013, 7, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Mackawy, A.M.H.; Badawi, M.E.H. Association of vitamin D and vitamin D receptor gene polymorphisms with chronic inflammation, insulin resistance and metabolic syndrome components in type 2 diabetic Egyptian patients. Meta Gene 2014, 2, 540–556. [Google Scholar] [CrossRef] [PubMed]
- Pittas, A.G.; Harris, S.S.; Stark, P.C.; Dawson-Hughes, B. The effects of calcium and vitamin D supplementation on blood glucose and markers of inflammation in nondiabetic adults. Diabetes Care 2007, 30, 980–986. [Google Scholar] [CrossRef]
- Tsukada, J.; Yoshida, Y.; Kominato, Y.; Auron, P.E. The CCAAT/enhancer (C/EBP) family of basic-leucine zipper (bZIP) transcription factors is a multifaceted highly-regulated system for gene regulation. Cytokine 2011, 54, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Robitaille, J.; Després, J.-P.; Pérusse, L.; Vohl, M.-C. The PPAR-gamma P12A polymorphism modulates the relationship between dietary fat intake and components of the metabolic syndrome: Results from the Québec Family Study. Clin. Genet. 2003, 63, 109–116. [Google Scholar] [CrossRef]
- Taylor, H.L.; Jacobs, D.R.; Schucker, B.; Knudsen, J.; Leon, A.S.; Debacker, G. A questionnaire for the assessment of leisure time physical activities. J. Chronic Dis. 1978, 31, 741–755. [Google Scholar] [CrossRef]
- Goulet, J.; Nadeau, G.; Lapointe, A.; Lamarche, B.; Lemieux, S. Validity and reproducibility of an interviewer-administered food frequency questionnaire for healthy French-Canadian men and women. Nutr. J. 2004, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Ledue, T.B.; Weiner, D.L.; Sipe, J.D.; Poulin, S.E.; Collins, M.F.; Rifai, N. Analytical evaluation of particle-enhanced immunonephelometric assays for C-reactive protein, serum amyloid A and mannose-binding protein in human serum. Ann. Clin. Biochem. 1998, 35, 745–753. [Google Scholar] [CrossRef]
- Vossen, M.; Tödter, K.; Altenburg, C.; Beisiegel, U.; Scheja, L. Plasma triglycerides after oral glucose load specifically associate with metabolic risk markers in healthy type 2 diabetes offspring. Atherosclerosis 2011, 217, 214–219. [Google Scholar] [CrossRef]
- Buchanan, J.R.; Santen, R.; Cauffman, S.; Cavaliere, A.; Greer, R.B.; Demers, L.M. The effect of endogenous estrogen fluctuation on metabolism of 25-hydroxyvitamin D. Calcif. Tissue Int. 1986, 39, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Gaujoux, R.; Seoighe, C. CellMix: A comprehensive toolbox for gene expression deconvolution. Bioinformatics 2013, 29, 2211–2212. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Lee, S.; Majewski, I.J.; Alexander, W.S.; Smyth, G.K. Robust hyperparameter estimation protects against hypervariable genes and improves power to detect differential expression. Ann. Appl. Stat. 2016, 10, 946–963. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Characteristics | Control | BPH | VitD3 | VitD3 + BPH | p Value † |
|---|---|---|---|---|---|
| Number of subjects (males/females) | 22 (11/11) | 22 (11/11) | 22 (11/11) | 22 (11/11) | --- |
| BMI (kg/m2) | 31.5 ± 4.9 | 31.4 ± 4.9 | 31.4 ± 4.8 | 31.4 ± 4.8 | 1.00 |
| Waist girth (cm) | 103.8 ± 14.5 | 103.5 ± 14.5 | 103.7 ± 14.4 | 103.6 ± 14.3 | 1.00 |
| Lipid profile | |||||
| Total-C (mmol/L) | 4.93 ± 0.98 | 5.09 ± 0.85 | 5.18 ± 0.86 | 5.17 ± 0.89 | 0.78 |
| LDL-C (mmol/L) | 2.95 ± 0.74 | 3.10 ± 0.76 | 3.04 ± 0.61 | 3.13 ± 0.72 | 0.84 |
| HDL-C (mmol/L) | 1.26 ± 0.28 | 1.30 ± 0.33 | 1.35 ± 0.38 | 1.27 ± 0.34 | 0.83 |
| TG (mmol/L) | 1.56 ± 0.67 | 1.50 ± 0.59 | 1.74 ± 1.07 | 1.81 ± 1.21 | 0.89 |
| Total-C/HDL-C | 4.07 ± 1.16 | 4.13 ± 1.19 | 4.14 ± 1.39 | 4.32 ± 1.32 | 0.92 |
| Blood pressure (mm Hg) | |||||
| SBP | 114.1 ± 8.6 | 112.2 ± 8.0 | 112.9 ± 7.3 | 112.5 ± 8.3 | 0.88 |
| DBP | 71.3 ± 5.0 | 69.4 ± 7.0 | 71.0 ± 6.7 | 70.4 ± 7.6 | 0.77 |
| Glucose homeostasis | |||||
| Fasting glucose (mmol/L) | 5.13 ± 0.46 | 5.16 ± 0.42 | 5.25 ± 0.43 | 5.19 ± 0.46 | 0.84 |
| Fasting insulin (pmol/L) | 114.0 ± 54.3 | 118.8 ± 61.5 | 115.8 ± 42.9 | 116.0 ± 53.5 | 0.99 |
| HOMA-IR | 3.74 ± 1.83 | 3.91 ± 2.05 | 3.87 ± 1.44 | 3.85 ± 1.80 | 0.99 |
| Diabetes status (Nondiabetic/prediabetic/de novo diabetes) | |||||
| Fasting glucose | 21/1/0 | 21/1/0 | 21/1/0 | 21/1/0 | 1.00 |
| 2 h, 75 g OGTT | 16/6/0 | --- | --- | --- | --- |
| CRP (mg/L) | 2.86 ± 3.58 | 3.27 ± 3.52 | 2.58 ± 2.65 | 2.76 ± 4.03 | 0.95 * |
| 25-hydroxyvitamin D (nmol/L) | 56.6 ± 19.8 | 57.3 ± 21.9 | 56.8 ± 20.3 | 57.1 ± 19.1 | 1.00 |
| Deficiency/sufficiency | 8/14 | 9/13 | 9/13 | 7/15 | 0.91 ‡ |
| Treatment Metabolic Parameter | 0 | 15 | 30 | 60 | 120 | 180 | p Value † |
|---|---|---|---|---|---|---|---|
| Control | |||||||
| Glucose (mmol/L) | 5.13 ± 0.46 | 6.86 ± 0.97 | 7.99 ± 1.60 | 7.72 ± 2.58 | 6.14 ± 1.64 | 4.40 ± 1.35 | 0.98 |
| Insulin (pmol/L) | 114.0 ± 54.3 | 524.0 ± 286.5 | 858.6 ± 446.3 | 977.5 ± 502.9 | 733.6 ± 402.5 | 286.5 ± 258.7 | 0.21 |
| C-peptide (pmol/L) | 894.7 ± 279.9 | 2028.3 ± 659.5 | 3016.9 ± 944.6 | 3811.3 ± 1047.0 | 3624.4 ± 1169.0 | 2171.6 ± 944.1 | 0.41 |
| TG (mmol/L) | 1.43 ± 0.67 | 1.40 ± 0.60 | 1.47 ± 0.63 | 1.52 ± 0.64 | 1.41 ± 0.67 | 1.37 ± 0.65 | 0.54 |
| BPH | |||||||
| Glucose (mmol/L) | 5.16 ± 0.42 | 7.06 ± 1.11 | 8.08 ± 1.79 | 7.61 ± 2.31 | 6.51 ± 1.87 | 4.39 ± 1.80 | |
| Insulin (pmol/L) | 118.8 ± 61.5 | 592.2 ± 398.1 | 1025.9 ± 565.4 | 1151.6 ± 872.5 | 947.8 ± 506.2 | 313.5 ± 276.5 | |
| C-peptide (pmol/L) | 960.0 ± 287.7 | 2284.0 ± 889.1 | 3382.1 ± 1091.7 | 4163.0 ± 1467.1 | 4189.3 ± 1163.2 | 2487.0 ± 1034.6 | |
| TG (mmol/L) | 1.36 ± 0.56 | 1.38 ± 0.56 | 1.40 ± 0.54 | 1.42 ± 0.57 | 1.36 ± 0.55 | 1.36 ± 0.56 | |
| VitD3 | |||||||
| Glucose (mmol/L) | 5.25 ± 0.43 | 7.00 ± 0.94 | 8.08 ± 1.42 | 8.03 ± 2.67 | 6.49 ± 1.97 | 4.41 ± 1.61 | |
| Insulin (pmol/L) | 115.8 ± 42.9 | 562.0 ± 309.1 | 898.6 ± 469.9 | 1052.1 ± 495.2 | 914.6 ± 527.0 | 300.9 ± 252.6 | |
| C-peptide (pmol/L) | 934.4 ± 225.8 | 2132.8 ± 682.2 | 3169.7 ± 957.7 | 4083.3 ± 899.8 | 4016.3 ± 1448.1 | 2282.7 ± 1075.2 | |
| TG (mmol/L) | 1.59 ± 0.99 | 1.58 ± 0.98 | 1.58 ± 0.96 | 1.59 ± 0.94 | 1.53 ± 0.97 | 1.54 ± 0.99 | |
| VitD3 + BPH | |||||||
| Glucose (mmol/L) | 5.19 ± 0.46 | 7.21 ± 1.13 | 7.90 ± 1.65 | 8.12 ± 2.49 | 6.42 ± 1.78 | 4.39 ± 1.48 | |
| Insulin (pmol/L) | 116.0 ± 53.5 | 624.5 ± 351.7 | 889.6 ± 595.9 | 1114.2 ± 772.1 | 1035.7 ± 508.3 | 393.7 ± 372.3 | |
| C-peptide (pmol/L) | 957.9 ± 292.1 | 2295.3 ± 783.2 | 3179.8 ± 1071.9 | 4177.2 ± 1195.0 | 4229.2 ± 1106.9 | 2478.3 ± 1264.5 | |
| TG (mmol/L) | 1.61 ± 0.99 | 1.62 ± 0.97 | 1.59 ± 0.85 | 1.60 ± 0.83 | 1.45 ± 0.81 | 1.46 ± 0.69 | |
| Glucose p value †† | 0.81 | 0.71 | 0.98 | 0.88 | 0.89 | 1.00 | |
| Insulin p value †† | 0.99 | 0.78 | 0.70 | 0.84 | 0.23 | 0.63 | |
| C-peptide p value †† | 0.83 | 0.58 | 0.68 | 0.71 | 0.35 | 0.70 | |
| TG p value †† | 0.69 | 0.72 | 0.80 | 0.84 | 0.90 | 0.82 |
| Characteristics | Control | BPH | VitD3 | VitD3 + BPH | p Value † |
|---|---|---|---|---|---|
| Number of subjects (males/females) | 18 (10/8) | 18 (10/8) | 18 (10/8) | 18 (10/8) | --- |
| BMI (kg/m2) | 31.4 ± 4.9 | 31.3 ± 4.9 | 31.3 ± 4.9 | 31.3 ± 4.8 | 1.00 |
| Waist girth (cm) | 105.7 ± 14.9 | 105.3 ± 15.0 | 105.6 ± 14.9 | 105.5 ± 14.8 | 1.00 |
| Lipid profile | |||||
| Total-C (mmol/L) | 4.90 ± 0.95 | 5.14 ± 0.89 | 5.25 ± 0.89 | 5.18 ± 0.94 | 0.68 |
| LDL-C (mmol/L) | 2.95 ± 0.68 | 3.17 ± 0.77 | 3.15 ± 0.58 | 3.18 ± 0.72 | 0.70 |
| HDL-C (mmol/L) | 1.23 ± 0.25 | 1.26 ± 0.27 | 1.33 ± 0.35 | 1.27 ± 0.29 | 0.76 |
| TG (mmol/L) | 1.56 ± 0.70 | 1.54 ± 0.62 | 1.66 ± 1.06 | 1.59 ± 0.82 | 0.98 |
| Total-C/HDL-C | 4.09 ± 1.05 | 4.24 ± 1.14 | 4.18 ± 1.38 | 4.24 ± 1.12 | 0.98 |
| Blood pressure (mm Hg) | |||||
| SBP | 115.5 ± 8.7 | 113.2 ± 8.2 | 114.6 ± 6.9 | 113.6 ± 8.6 | 0.83 |
| DBP | 70.9 ± 4.9 | 68.5 ± 7.1 | 70.7 ± 7.1 | 69.1 ± 7.2 | 0.64 |
| Glucose homeostasis | |||||
| Fasting glucose (mmol/L) | 5.18 ± 0.45 | 5.22 ± 0.43 | 5.29 ± 0.42 | 5.24 ± 0.48 | 0.91 |
| Fasting insulin (pmol/L) | 110.9 ± 58.6 | 111.0 ± 64.1 | 113.3 ± 45.0 | 108.8 ± 55.2 | 1.00 |
| HOMA-IR | 3.71 ± 2.02 | 3.73 ± 2.20 | 3.82 ± 1.49 | 3.67 ± 1.91 | 1.00 |
| Diabetes status (Nondiabetic/prediabetic/de novo diabetes) | |||||
| Fasting glucose | 18/1/0 | 18/1/0 | 18/1/0 | 18/1/0 | 1.00 |
| 2 h, 75 g OGTT | 13/6/0 | --- | --- | --- | --- |
| CRP (mg/L) | 3.16 ± 3.99 | 3.14 ± 3.42 | 2.71 ± 2.92 | 2.98 ± 4.40 | 0.97 * |
| 25-hydroxyvitamin D (nmol/L) | 61.0 ± 18.5 | 62.2 ± 20.3 | 61.4 ± 18.6 | 61.8 ± 17.0 | 0.98 |
| Deficiency/sufficiency | 5/13 | 6/12 | 6/12 | 4/14 | 0.74 ‡ |
| Transcript | Gene Symbol | Probe ID | Treatment | Fold Change | FDR p Value |
|---|---|---|---|---|---|
| NM_001031847 | CPT1A | TC1100011395.hg.1 | VitD3 | 0.67 | 2.6 × 10−6 |
| NM_001031847 | CPT1A | TC1100011395.hg.1 | BPH | 0.67 | 0.0005 |
| NM_001031847 | CPT1A | TC1100011395.hg.1 | VitD3 + BPH | 0.60 | 0.002 |
| NM_000387 | SLC25A20 | TC0300011038.hg.1 | VitD3 + BPH | 0.61 | 0.0002 |
| NM_000387 | SLC25A20 | TC0300011038.hg.1 | VitD3 | 0.63 | 0.009 |
| NM_001167928 | IL1RAP | TC0300009855.hg.1 | VitD3 + BPH | 0.78 | 0.04 |
| NM_001080392 | KIAA1147 | TC0700012836.hg.1 | VitD3 + BPH | 0.81 | 0.04 |
| NM_005461 | MAFB | TC2000009116.hg.1 | VitD3 + BPH | 1.40 | 0.04 |
| NM_020870 | SH3RF1 | TC0400012378.hg.1 | VitD3 | 1.62 | 0.008 |
| NM_001031700 | FAM198B | TC0400012245.hg.1 | VitD3 | 1.34 | 0.008 |
| NM_018404 | ADAP2 | TC1700012226.hg.1 | VitD3 | 1.37 | 0.008 |
| NM_000564 | IL5RA | TC0300013923.hg.1 | VitD3 | 0.74 | 0.008 |
| NM_001285829 | CEBPA | TC1900010386.hg.1 | VitD3 | 1.34 | 0.009 |
| NM_001860 | SLC31A2 | TC0900008482.hg.1 | VitD3 | 0.84 | 0.02 |
| NM_001271842 | SCIMP | TC1700009556.hg.1 | VitD3 | 1.20 | 0.02 |
| NM_016562 | TLR7 | TC0X00006625.hg.1 | VitD3 | 1.30 | 0.03 |
| NM_013451 | MYOF | TC1000011445.hg.1 | VitD3 | 1.40 | 0.03 |
| NM_004137 | KCNMB1 | TC0500012791.hg.1 | VitD3 | 1.24 | 0.03 |
| NM_001168357 | PLA2G7 | TC0600011953.hg.1 | VitD3 | 1.39 | 0.04 |
| NM_032109 | OTP | TC0500011211.hg.1 | VitD3 | 0.79 | 0.04 |
| NM_001122 | PLIN2 | TC0900012212.hg.1 | VitD3 | 0.76 | 0.04 |
| NM_000355 | TCN | TC2200009257.hg.1 | VitD3 | 1.30 | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guénard, F.; Jacques, H.; Gagnon, C.; Marette, A.; Vohl, M.-C. Acute Effects of Single Doses of Bonito Fish Peptides and Vitamin D on Whole Blood Gene Expression Levels: A Randomized Controlled Trial. Int. J. Mol. Sci. 2019, 20, 1944. https://doi.org/10.3390/ijms20081944
Guénard F, Jacques H, Gagnon C, Marette A, Vohl M-C. Acute Effects of Single Doses of Bonito Fish Peptides and Vitamin D on Whole Blood Gene Expression Levels: A Randomized Controlled Trial. International Journal of Molecular Sciences. 2019; 20(8):1944. https://doi.org/10.3390/ijms20081944
Chicago/Turabian StyleGuénard, Frédéric, Hélène Jacques, Claudia Gagnon, André Marette, and Marie-Claude Vohl. 2019. "Acute Effects of Single Doses of Bonito Fish Peptides and Vitamin D on Whole Blood Gene Expression Levels: A Randomized Controlled Trial" International Journal of Molecular Sciences 20, no. 8: 1944. https://doi.org/10.3390/ijms20081944
APA StyleGuénard, F., Jacques, H., Gagnon, C., Marette, A., & Vohl, M.-C. (2019). Acute Effects of Single Doses of Bonito Fish Peptides and Vitamin D on Whole Blood Gene Expression Levels: A Randomized Controlled Trial. International Journal of Molecular Sciences, 20(8), 1944. https://doi.org/10.3390/ijms20081944