Clinical and Biological Significance of ESR1 Gene Alteration and Estrogen Receptors Isoforms Expression in Breast Cancer Patients

 ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results
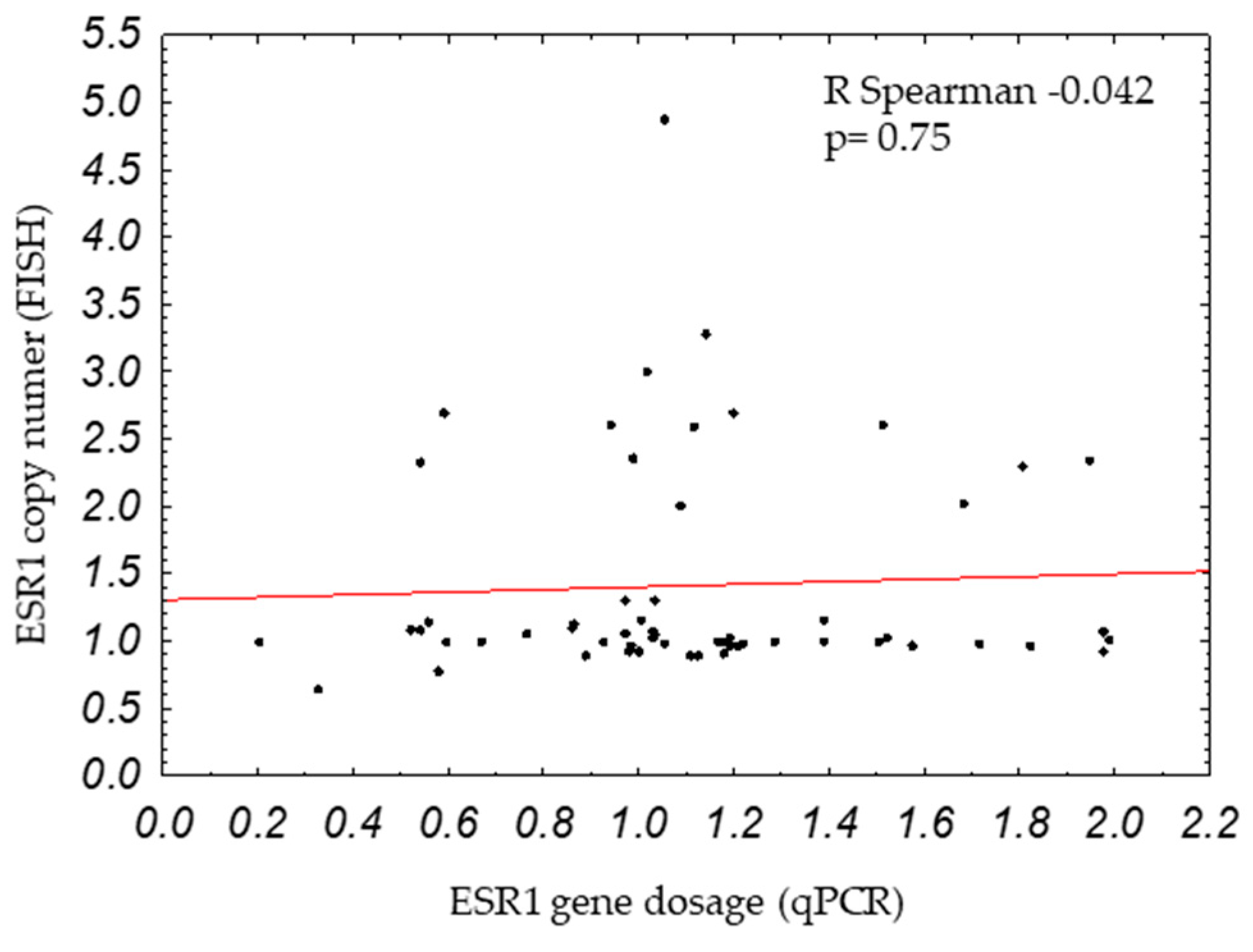
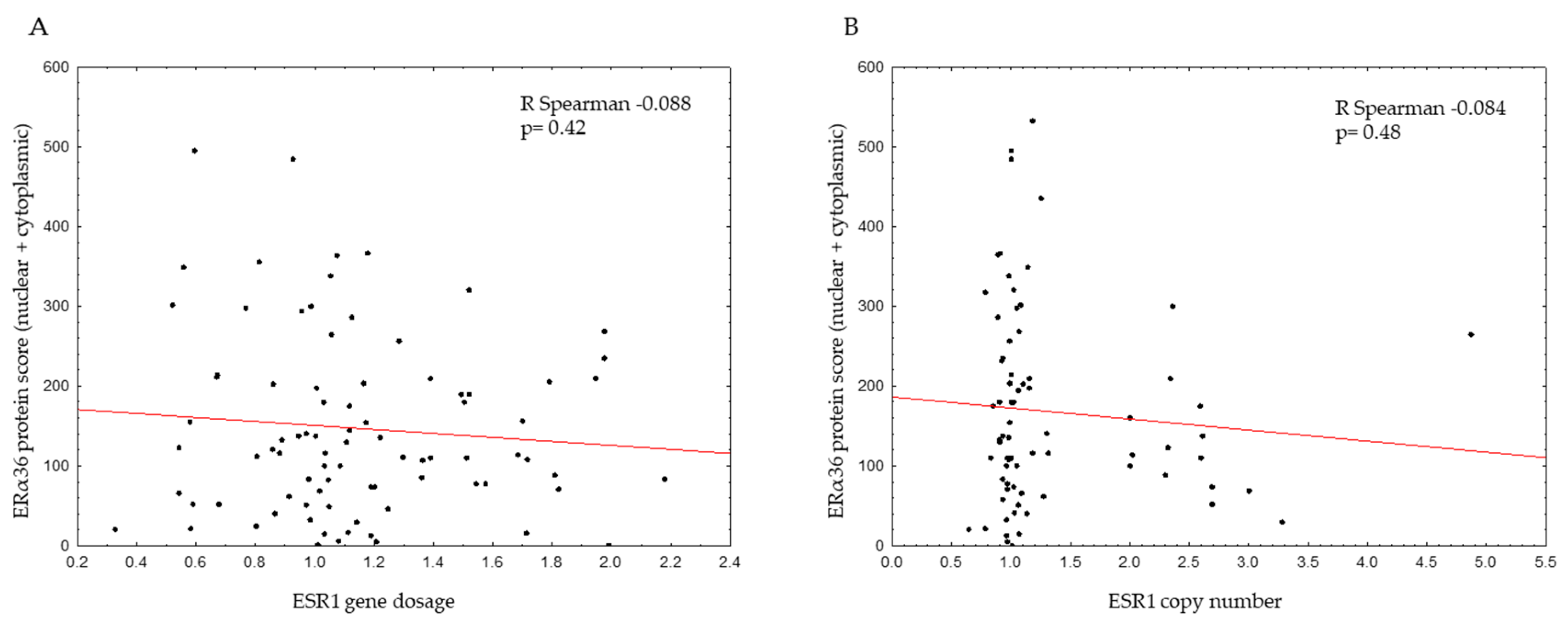
2.1. ESR1 Gene Dosage and Copy Number in Breast Cancers
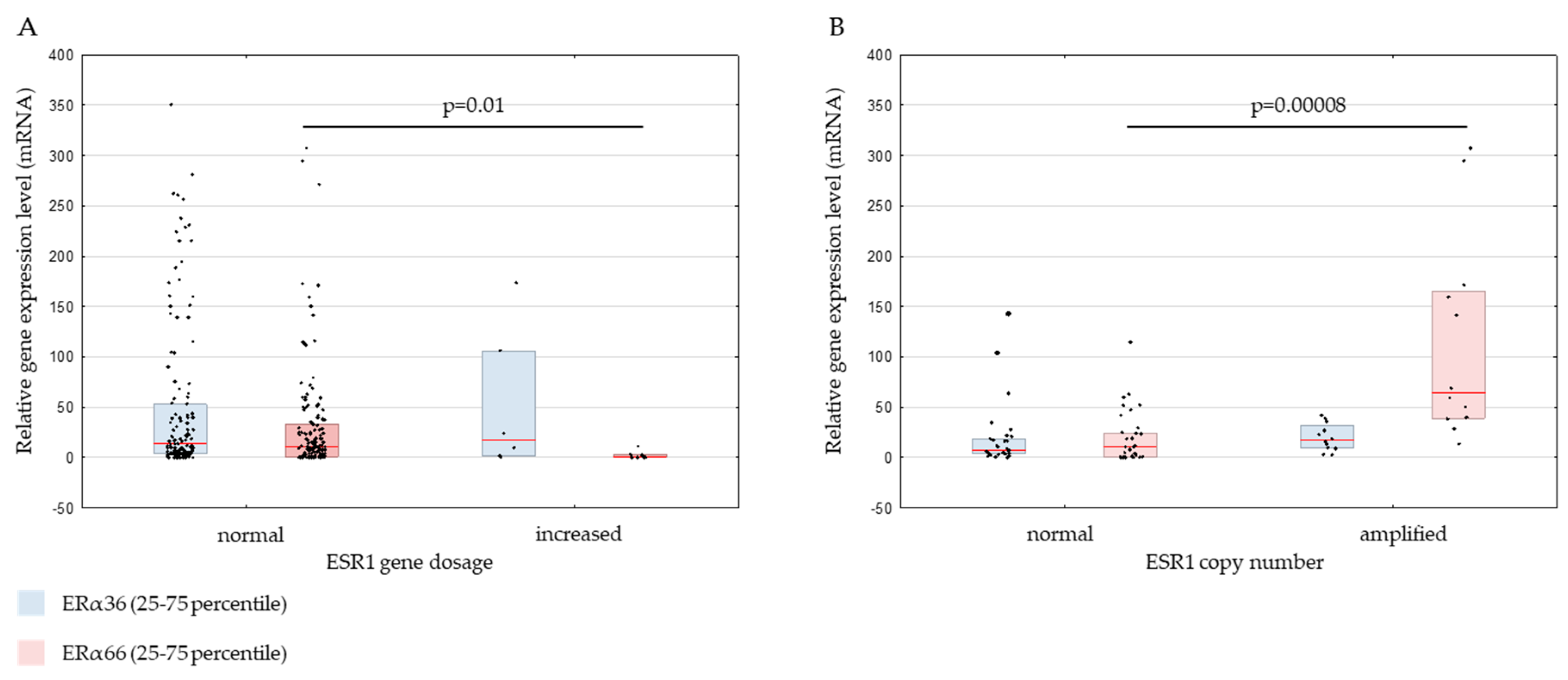
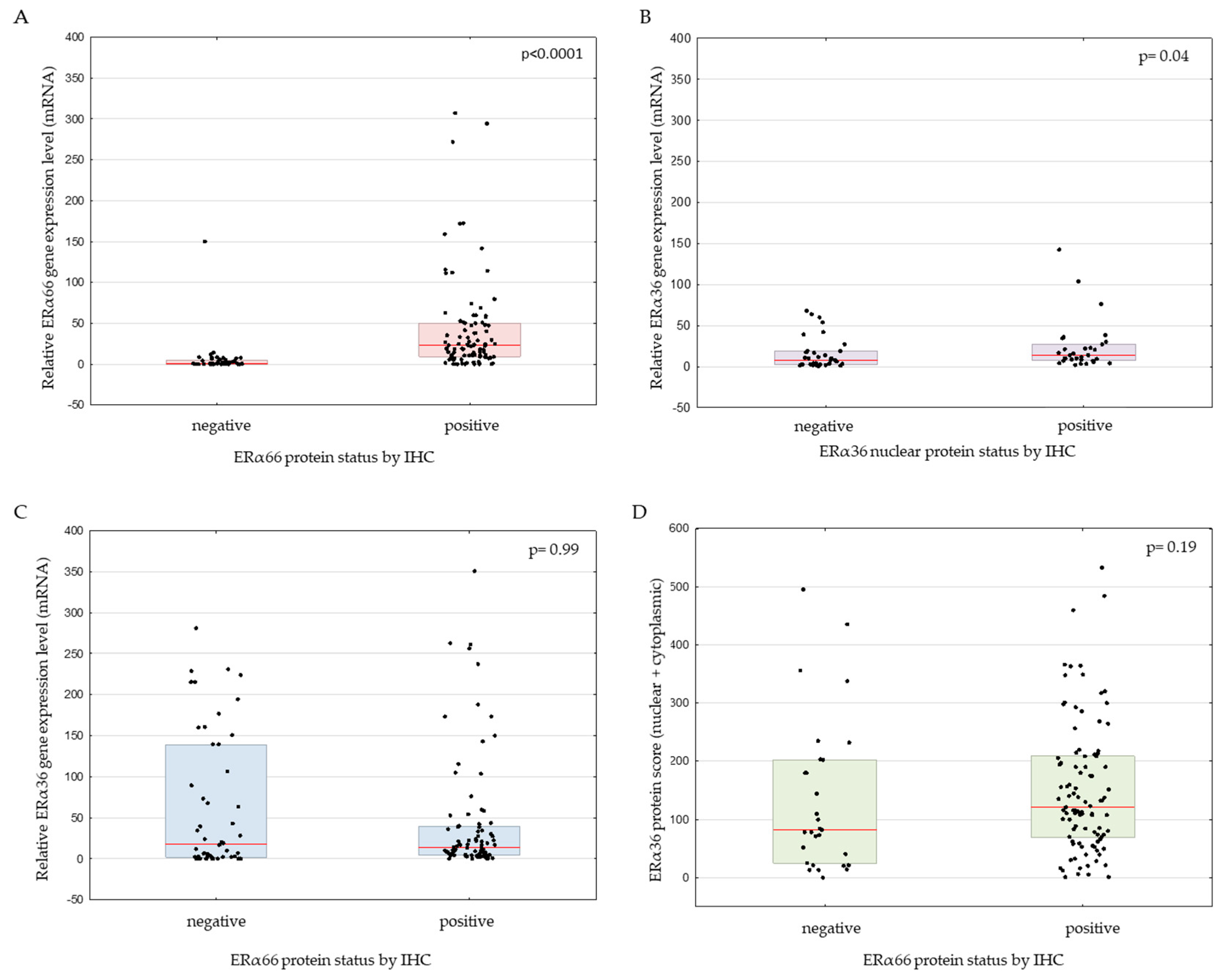
2.2. Genomic ESR1 Level versus mRNA and Protein Isoforms Expression
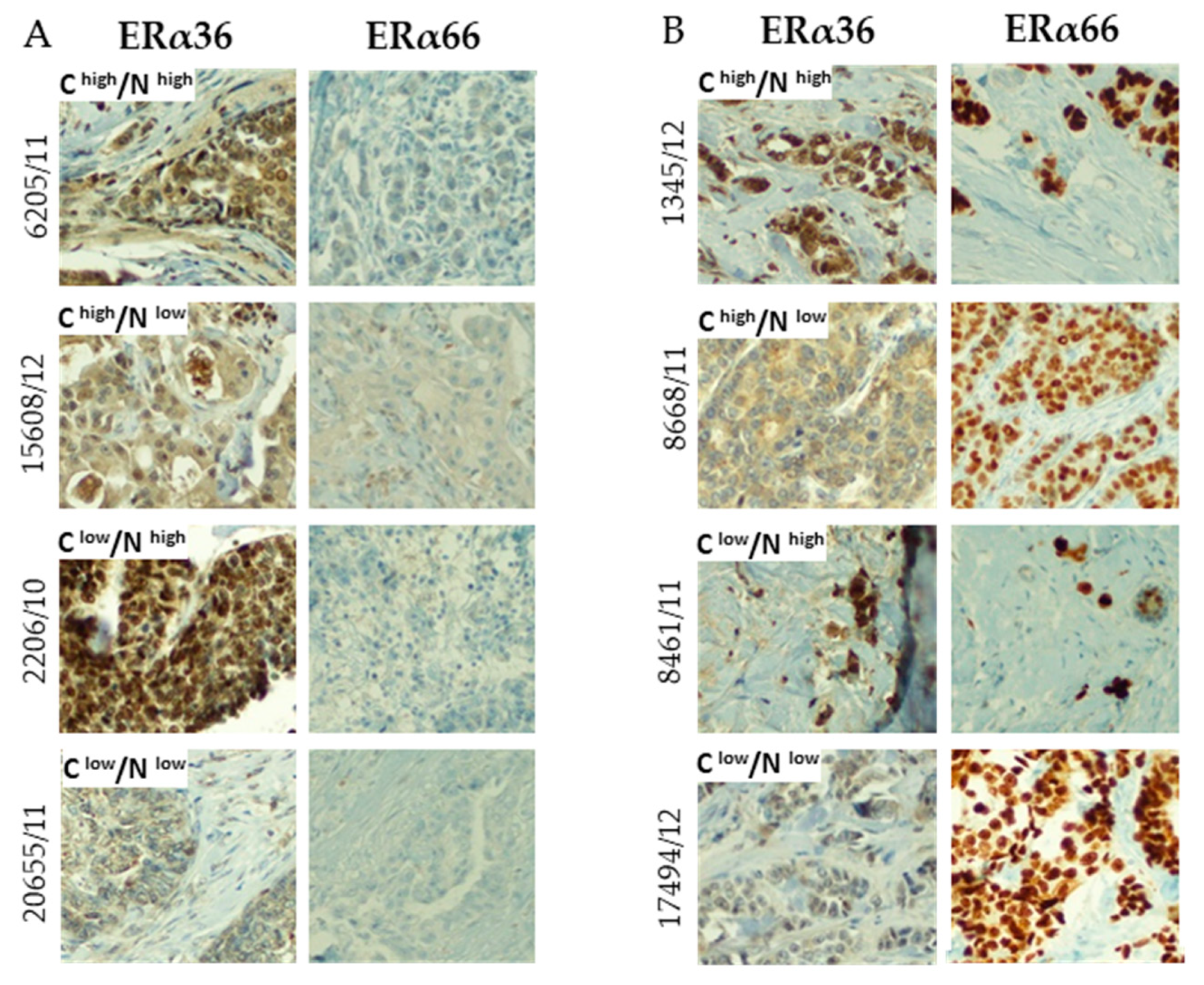
2.3. Correlation between ERα66 and ERα36 Isoforms on mRNA and Protein Level
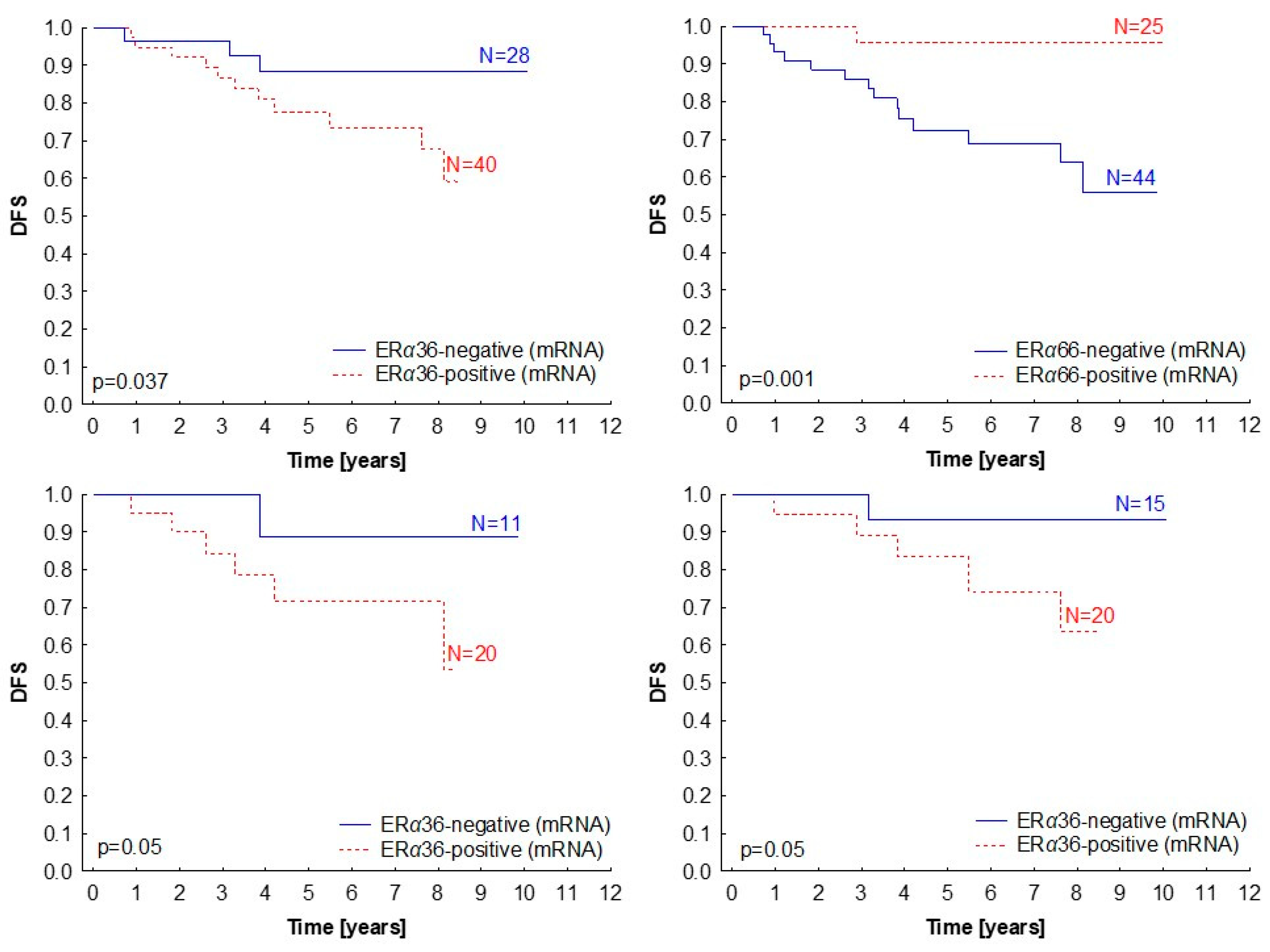
2.4. Clinical Significance of ERα Isoforms Expression
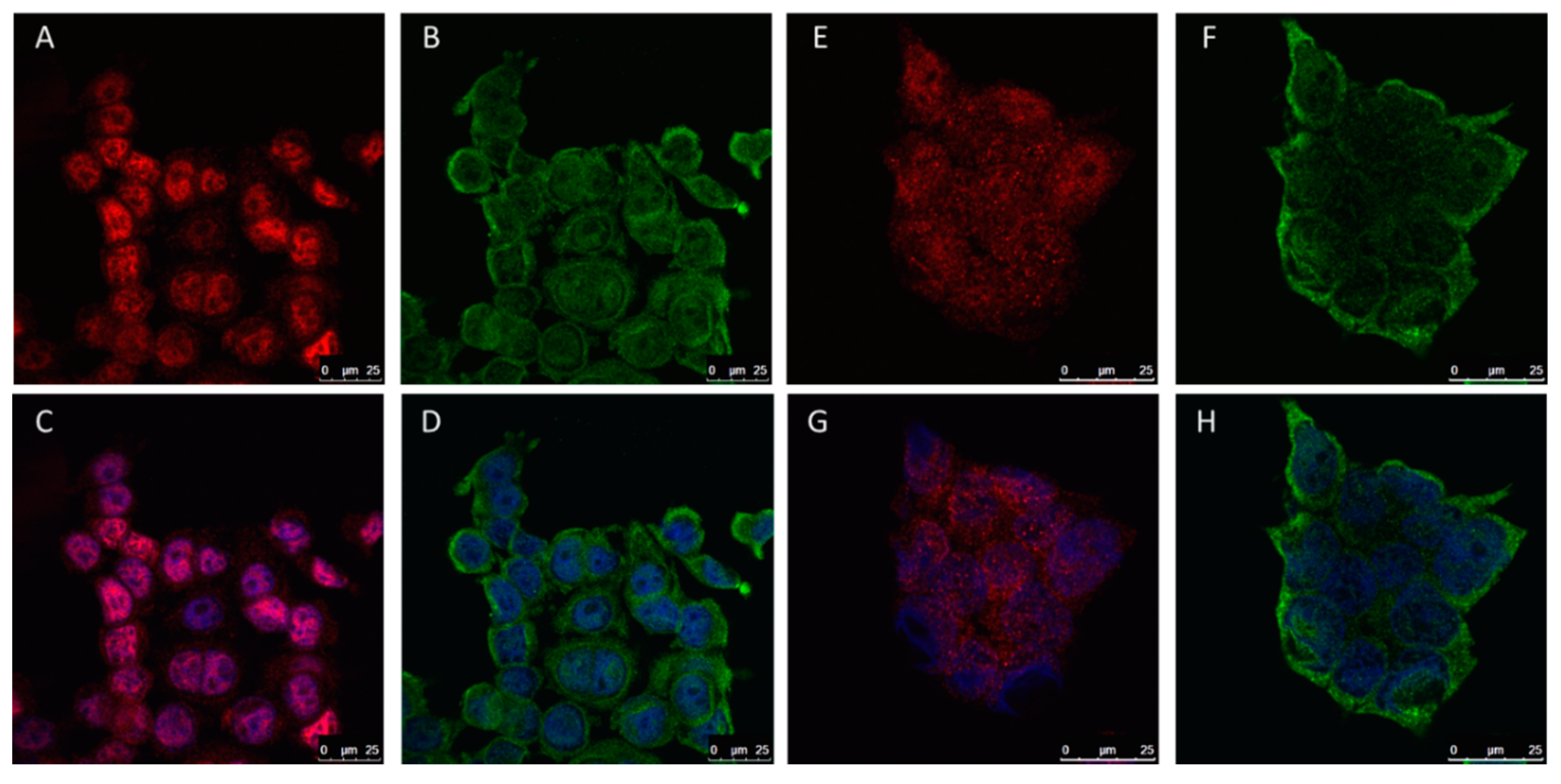
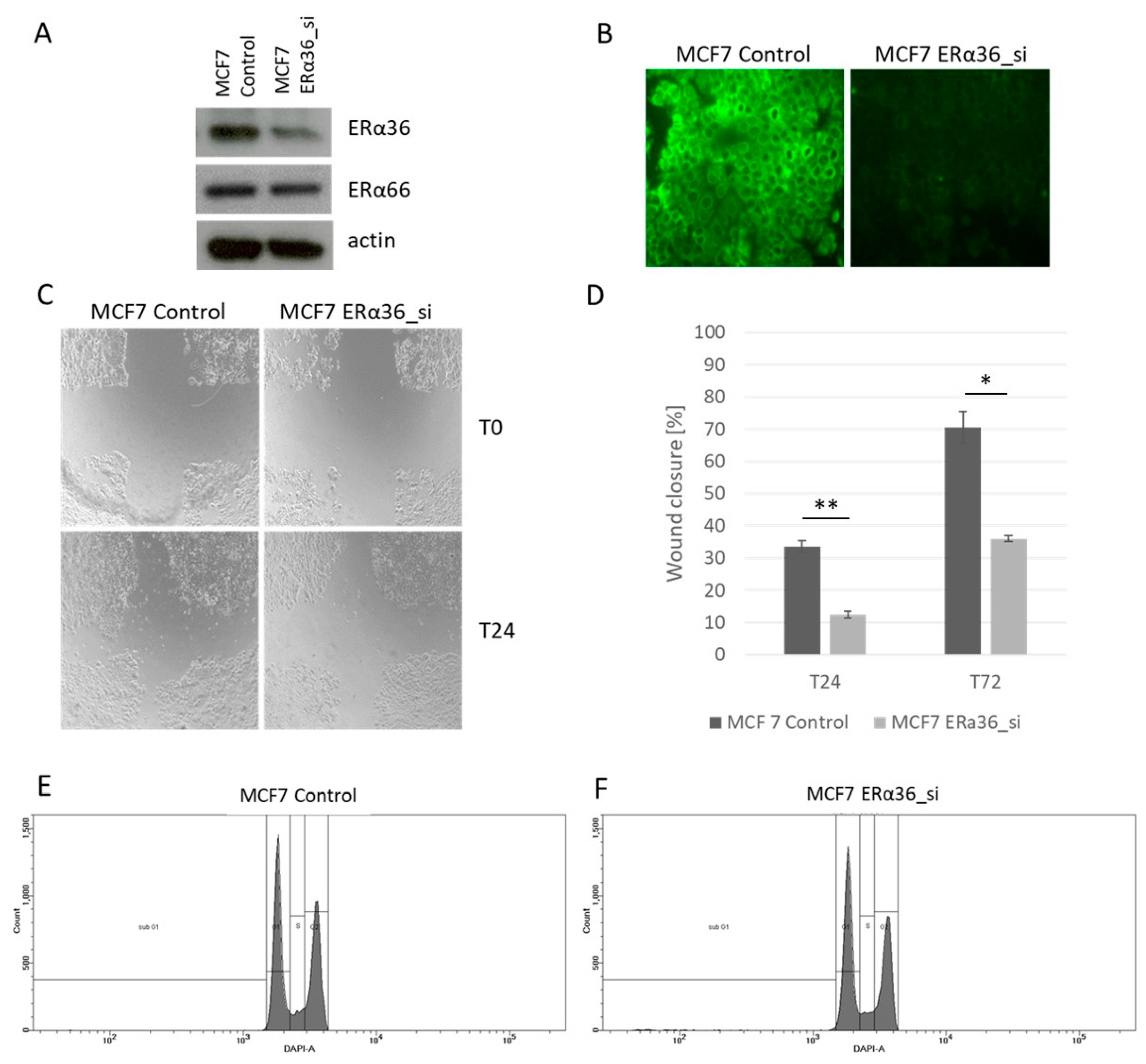
2.5. The Role of ERα36 Isoform in Breast Cancer Cell Lines
3. Discussion
4. Materials and Methods
4.1. RNA and DNA Isolation, Reverse Transcription, and Gene Expression Analysis
4.2. ESR1 Gene Dosage Analysis with qPCR
4.3. ESR1 Copy Number Analysis with Fluorescent in situ Hybridization
4.4. ERα36 Protein Level Analysis with Immunohistochemistry
4.5. ERα66, PR, and HER2 Status Analysis in Breast Cancer Samples
4.6. Cell Culture
4.7. Gene Knock-Down with siRNA and Western Blotting
4.8. Immunofluorescent Staining
4.9. Wound-Healing Assay
4.10. Cell Cycle Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ESR1 | estrogen receptor gene |
| ER | estrogen receptor alpha |
| ERα36 | estrogen receptor alpha, 36kDa isoform |
| ERα66 | estrogen receptor alpha, 66kDa isoform |
| FISH | fluorescent in situ hybridization |
| HER2 | human epidermal growth factor receptor |
| IHC | immunohistochemistry |
| PR | progesterone receptor alpha |
| OS | overall survial |
| DFS | disease free survival |
References
- Orlando, L.; Schiavone, P.; Fedele, P.; Calvani, N.; Nacci, A.; Rizzo, P.; Marino, A.; D’Amico, M.; Sponziello, F.; Mazzoni, E.; et al. Molecularly targeted endocrine therapies for breast cancer. Cancer Treat Rev. 2010, 36, 67–71. [Google Scholar] [CrossRef]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. Arch. Pathol. Lab. Med. 2010, 134, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.J.; Panel members. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef] [PubMed]
- Holst, F.; Stahl, P.R.; Ruiz, C.; Hellwinkel, O.; Jehan, Z.; Wendland, M.; Lebeau, A.; Terracciano, L.; Al-Kuraya, K.; Jänicke, F.; et al. Estrogen receptor alpha (ESR1) gene amplification is frequent in breast cancer. Nat. Genet. 2007, 39, 655–660. [Google Scholar] [CrossRef]
- Moelans, C.B.; Holst, F.; Hellwinkel, O.; Simon, R.; van Diest, P.J. ESR1 amplification in breast cancer by optimized RNase FISH: Frequent but low-level and heterogeneous. PLoS ONE 2013, 8, e84189. [Google Scholar] [CrossRef][Green Version]
- Markiewicz, A.; Welnicka-Jaskiewicz, M.; Skokowski, J.; Jaskiewicz, J.; Szade, J.; Jassem, J.; Zaczek, A.J. Prognostic significance of ESR1 amplification and ESR1 PvuII, CYP2C19*2, UGT2B15*2 polymorphisms in breast cancer patients. PLoS ONE 2013, 8, e72219. [Google Scholar] [CrossRef]
- Ooi, A.; Inokuchi, M.; Harada, S.; Inazawa, J.; Tajiri, R.; Kitamura, S.S.; Ikeda, H.; Kawashima, H.; Dobashi, Y. Gene amplification of ESR1 in breast cancers-fact or fiction? A fluorescence in situ hybridization and multiplex ligation-dependent probe amplification study. J. Pathol. 2012, 227, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Moelans, C.B.; Monsuur, H.N.; de Pinth, J.H.; Radersma, R.D.; de Weger, R.A.; van Diest, P.J. ESR1 amplification is rare in breast cancer and is associated with high grade and high proliferation: A multiplex ligation-dependent probe amplification study. Cell Oncol. (Dordr) 2011, 34, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Holst, F. Estrogen receptor alpha gene amplification in breast cancer: 25 years of debate. World J. Clin. Oncol. 2016, 7, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Zaczek, A.J.; Markiewicz, A.; Seroczynska, B.; Skokowski, J.; Jaskiewicz, J.; Pienkowski, T.; Olszewski, W.P.; Szade, J.; Rhone, P.; Welnicka-Jaskiewicz, M.; et al. Prognostic Significance of TOP2A Gene Dosage in HER-2-Negative Breast Cancer. Oncologist 2012, 17, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Shen, P.; Loggie, B.W.; Chang, Y.; Deuel, T.F. A variant of estrogen receptor-{alpha}, hER-{alpha}36: Transduction of estrogen- and antiestrogen-dependent membrane-initiated mitogenic signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 9063–9068. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Shen, P.; Loggie, B.W.; Chang, Y.; Deuel, T.F. Identification, cloning, and expression of human estrogen receptor-alpha36, a novel variant of human estrogen receptor-alpha66. Biochem Biophys Res. Commun. 2005, 336, 1023–1027. [Google Scholar] [CrossRef]
- Sołtysik, K.; Czekaj, P. ERα36--Another piece of the estrogen puzzle. Eur. J. Cell. Biol. 2015, 94, 611–625. [Google Scholar] [CrossRef]
- Chaudhri, R.A.; Schwartz, N.; Elbaradie, K.; Schwartz, Z.; Boyan, B.D. Role of ERα36 in membrane-associated signaling by estrogen. Steroids 2014, 81, 74–80. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, J.; Ying, G.; Xie, X.Q.; Zhang, X.; Xu, W.; Song, E.; Bu, H.; Ping, Y.F.; Yao, X.H.; et al. Tamoxifen enhances stemness and promotes metastasis of ERα36 + breast cancer by upregulating ALDH1A1 in cancer cells. Cell. Res. 2018, 28, 336–358. [Google Scholar] [CrossRef] [PubMed]
- Chaudhri, R.A.; Olivares-Navarrete, R.; Cuenca, N.; Hadadi, A.; Boyan, B.D.; Schwartz, Z. Membrane estrogen signaling enhances tumorigenesis and metastatic potential of breast cancer cells via estrogen receptor-α36 (ERα36). J. Biol. Chem. 2012, 287, 7169–7181. [Google Scholar] [CrossRef]
- Zhang, X.T.; Kang, L.G.; Ding, L.; Vranic, S.; Gatalica, Z.; Wang, Z.Y. A positive feedback loop of ER-α36/EGFR promotes malignant growth of ER-negative breast cancer cells. Oncogene 2011, 30, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Wallacides, A.; Chesnel, A.; Ajj, H.; Chillet, M.; Flament, S.; Dumond, H. Estrogens promote proliferation of the seminoma-like TCam-2 cell line through a GPER-dependent ERα36 induction. Mol. Cell. Endocrinol. 2012, 350, 61–71. [Google Scholar] [CrossRef]
- Shi, L.; Dong, B.; Li, Z.; Lu, Y.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Fan, T.; Lin, B.; et al. Expression of ER-{alpha}36, a novel variant of estrogen receptor {alpha}, and resistance to tamoxifen treatment in breast cancer. J. Clin. Oncol. 2009, 27, 3423–3429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, G.; Li, Z.; Yu, X.; Zheng, Y.; Jin, K.; Wang, H.; Gong, Y.; Sun, X.; Teng, X.; et al. Estrogen-independent effects of ER-α36 in ER-negative breast cancer. Steroids 2012, 77, 666–673. [Google Scholar] [CrossRef]
- Lee, L.M.; Cao, J.; Deng, H.; Chen, P.; Gatalica, Z.; Wang, Z.Y. ER-alpha36, a novel variant of ER-alpha, is expressed in ER-positive and -negative human breast carcinomas. Anticancer Res. 2008, 28, 479–483. [Google Scholar] [PubMed]
- Schwartz, N.; Verma, A.; Bivens, C.B.; Schwartz, Z.; Boyan, B.D. Rapid steroid hormone actions via membrane receptors. Biochim. Biophys Acta. 2016, 1863, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Soysal, S.D.; Kilic, I.B.; Regenbrecht, C.R.; Schneider, S.; Muenst, S.; Kilic, N.; Güth, U.; Dietel, M.; Terracciano, L.M.; Kilic, E. Status of estrogen receptor 1 (ESR1) gene in mastopathy predicts subsequent development of breast cancer. Breast Cancer Res. Treat 2015, 151, 709–715. [Google Scholar] [CrossRef]
- Lebeau, A.; Grob, T.; Holst, F.; Seyedi-Fazlollahi, N.; Moch, H.; Terracciano, L.; Turzynski, A.; Choschzick, M.; Sauter, G.; Simon, R. Oestrogen receptor gene (ESR1) amplification is frequent in endometrial carcinoma and its precursor lesions. J. Pathol. 2008, 216, 151–157. [Google Scholar] [CrossRef]
- Tomita, S.; Zhang, Z.; Nakano, M.; Ibusuki, M.; Kawazoe, T.; Yamamoto, Y.; Iwase, H. Estrogen receptor alpha gene ESR1 amplification may predict endocrine therapy responsiveness in breast cancer patients. Cancer Sci. 2009, 100, 1012–1017. [Google Scholar] [CrossRef]
- Laenkholm, A.V.; Knoop, A.; Ejlertsen, B.; Rudbeck, T.; Jensen, M.B.; Muller, S.; Lykkesfeldt, A.E.; Rasmussen, B.B.; Nielsen, K.V. ESR1 gene status correlates with estrogen receptor protein levels measured by ligand binding assay and immunohistochemistry. Mol. Oncol. 2012, 6, 428–436. [Google Scholar] [CrossRef]
- Lin, C.H.; Liu, J.M.; Lu, Y.S.; Lan, C.; Lee, W.C.; Kuo, K.T.; Wang, C.C.; Chang, D.Y.; Huang, C.S.; Cheng, A.L. Clinical significance of ESR1 gene copy number changes in breast cancer as measured by fluorescence in situ hybridisation. J. Clin. Pathol. 2013, 66, 140–145. [Google Scholar] [CrossRef]
- Tomita, S.; Abdalla, M.O.; Fujiwara, S.; Matsumori, H.; Maehara, K.; Ohkawa, Y.; Iwase, H.; Saitoh, N.; Nakao, M. A cluster of noncoding RNAs activates the ESR1 locus during breast cancer adaptation. Nat. Commun. 2015, 6, 6966. [Google Scholar] [CrossRef]
- Zaczek, A.; Markiewicz, A.; Jaskiewicz, J.; Pienkowski, T.; Rhone, P.; Jassem, J.; Welnicka-Jaskiewicz, M. Clinical evaluation of developed PCR-based method with hydrolysis probes for TOP2A copy number evaluation in breast cancer samples. Clinical. Biochemistry 2010, 43, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Pagani, F.; Baralle, F.E. Genomic variants in exons and introns: Identifying the splicing spoilers. Nat. Rev. Genet. 2004, 5, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yin, L.; Zhang, X.T.; Liu, L.J.; Wang, M.L.; Wang, Z.Y. ER-α variant ER-α36 mediates antiestrogen resistance in ER-positive breast cancer stem/progenitor cells. J. Steroid Biochem. Mol. Biol. 2014, 144 Pt B, 417–426. [Google Scholar] [CrossRef]
- Deng, H.; Zhang, X.T.; Wang, M.L.; Zheng, H.Y.; Liu, L.J.; Wang, Z.Y. ER-α36-mediated rapid estrogen signaling positively regulates ER-positive breast cancer stem/progenitor cells. PLoS ONE 2014, 9, e88034. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Pan, X.; Zhang, X.T.; Guo, Y.M.; Wang, Z.Y.; Gong, Y.; Wang, M. Downregulation of ER-α36 expression sensitizes HER2 overexpressing breast cancer cells to tamoxifen. Am. J. Cancer Res. 2015, 5, 530–544. [Google Scholar]
- Zhao, Y.; Deng, C.; Lu, W.; Xiao, J.; Ma, D.; Guo, M.; Recker, R.R.; Gatalica, Z.; Wang, Z.; Xiao, G.G. let-7 microRNAs induce tamoxifen sensitivity by downregulation of estrogen receptor α signaling in breast cancer. Mol. Med. 2011, 17, 1233–1241. [Google Scholar] [CrossRef]
- Li, L.; Wang, Q.; Lv, X.; Sha, L.; Qin, H.; Wang, L. Expression and localization of estrogen receptor in human breast cancer and its clinical significance. Cell Biochem. Biophys. 2015, 71, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Chamard-Jovenin, C.; Jung, A.C.; Chesnel, A.; Abecassis, J.; Flament, S.; Ledrappier, S.; Macabre, C.; Boukhobza, T.; Dumond, H. From ERα66 to ERα36: A generic method for validating a prognosis marker of breast tumor progression. BMC Syst. Biol. 2015, 9, 28. [Google Scholar] [CrossRef]
- Li, G.; Zhang, J.; Jin, K.; He, K.; Zheng, Y.; Xu, X.; Wang, H.; Li, Z.; Yu, X.; Teng, X.; et al. Estrogen receptor-α36 is involved in development of acquired tamoxifen resistance via regulating the growth status switch in breast cancer cells. Mol. Oncol. 2013, 7, 611–624. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Header | ESR1Gene Dosage Status (qPCR) [Number of Cases (%)] | Total | p | ESR1Gene Copy Number (FISH) [Number of Cases (%)] | Total | p | ||
| Normal | Increased | Normal | Amplified | |||||
| ERα66 Protein Status | ||||||||
| negative | 134 (37%) | 24 (65%) | 158 | 0.001 | 21 (36%) | 0 | 21 | 0.0038 |
| positive | 227 (63%) | 13 (25%) | 240 | 37 (64%) | 15 (100%) | 52 | ||
| Total | 361 (100%) | 37 (100%) | 58 (100%) | 15 (100%) | 73 | |||
| ESR1Gene Dosage Status (qPCR) | Total | p | ESR1Gene Copy Number (FISH) | Total | p | |||
| Normal | Increased | Normal | Amplified | |||||
| ERa66 mRNA Expression, Median (25–75 Percentile) | 10.63 (0.84–33.1) | 0.21 (0–3.22) | 145 | 0.0086 | 10.28 (0.2–24.5) | 64.14 (39–165.3) | 45 | 0.00002 |
| Variable | N | Median ERα36 Expression (25–75th Percentile) | p |
|---|---|---|---|
| T stage | p = 0.41 | ||
| T1–2 | 127 | 13.17 (3.75–42.90) | |
| T3–4 | 10 | 58.02 (7.37–160.88) | |
| N stage | p = 0.69 | ||
| N0 | 78 | 17.20 (3.52–63.44) | |
| N1 | 58 | 9.95 (4.23–40.05) | |
| Grading | p = 0.92 | ||
| 1 | 13 | 18.84 (5.34–27.19) | |
| 2 | 63 | 9.89 (3.75–37.41) | |
| 3 | 42 | 9.97 (2.89–39.08) | |
| Histological subtype | p = 0.21 | ||
| Ductal | 54 | 31.28 (4.1–144.7) | |
| Lobular | 7 | 74.39 (1.1–180.7) | |
| Other | 4 | 162.13 (78.1–205.5) | |
| Molecular type | p = 0.84 | ||
| Luminal A | 17 | 16.70 (4.68–150.35) | |
| Luminal B HER2– | 13 | 11.18 (3.98–24.24) | |
| Luminal B HER2+ | 2 | 97.21 (0–194.42) | |
| Non luminal HER2+ | 6 | 83.57 (0–160.88) | |
| Triple negative | 13 | 39.56 (2.88–160.1) | |
| ER status | p = 0.99 | ||
| 0 | 50 | 17.84 (1.91–139.11) | |
| 1 | 87 | 13.71 (4.41–39.08) | |
| PR status | p = 0.08 | ||
| 0 | 52 | 10.46 (1.52–53.17) | |
| 1 | 85 | 15.77 (5.34–54.09) | |
| HER2 status | p = 0.35 | ||
| 0 | 104 | 13.82 (3.94–59.45) | |
| 1 | 24 | 10.41 (2.56–31.21) |
| Variable | Number of Cases (%) | |
|---|---|---|
| Age | ||
| <50 | 120 | (29) |
| >50 | 298 | (71) |
| T Stage | ||
| 1 | 140 | (33) |
| 2 | 194 | (46) |
| 3 | 40 | (10) |
| 4 | 39 | (9) |
| Missing data | 5 | (2) |
| N Stage | ||
| negative | 207 | (49) |
| positive | 206 | (49) |
| Missing data | 5 | (2) |
| Grade | ||
| 1 | 30 | (7) |
| 2 | 171 | (41) |
| 3 | 135 | (32) |
| Missing data | 82 | (20) |
| Histologic Type | ||
| Ductal | 218 | (52) |
| Lobular | 54 | (13) |
| Other | 26 | (6) |
| Missing data | 120 | (29) |
| ER Status | ||
| negative | 164 | (39) |
| positive | 250 | (59) |
| Missing data | 4 | (2) |
| PR Status | ||
| negative | 175 | (42) |
| positive | 239 | (57) |
| Missing data | 4 | (2) |
| HER2 Status | ||
| negative | 274 | (66) |
| positive | 59 | (14) |
| Missing data | 85 | (20) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagel, A.; Szade, J.; Iliszko, M.; Elzanowska, J.; Welnicka-Jaskiewicz, M.; Skokowski, J.; Stasilojc, G.; Bigda, J.; Sadej, R.; Zaczek, A.; et al. Clinical and Biological Significance of ESR1 Gene Alteration and Estrogen Receptors Isoforms Expression in Breast Cancer Patients. Int. J. Mol. Sci. 2019, 20, 1881. https://doi.org/10.3390/ijms20081881
Nagel A, Szade J, Iliszko M, Elzanowska J, Welnicka-Jaskiewicz M, Skokowski J, Stasilojc G, Bigda J, Sadej R, Zaczek A, et al. Clinical and Biological Significance of ESR1 Gene Alteration and Estrogen Receptors Isoforms Expression in Breast Cancer Patients. International Journal of Molecular Sciences. 2019; 20(8):1881. https://doi.org/10.3390/ijms20081881
Chicago/Turabian StyleNagel, Anna, Jolanta Szade, Mariola Iliszko, Julia Elzanowska, Marzena Welnicka-Jaskiewicz, Jaroslaw Skokowski, Grzegorz Stasilojc, Jacek Bigda, Rafal Sadej, Anna Zaczek, and et al. 2019. "Clinical and Biological Significance of ESR1 Gene Alteration and Estrogen Receptors Isoforms Expression in Breast Cancer Patients" International Journal of Molecular Sciences 20, no. 8: 1881. https://doi.org/10.3390/ijms20081881
APA StyleNagel, A., Szade, J., Iliszko, M., Elzanowska, J., Welnicka-Jaskiewicz, M., Skokowski, J., Stasilojc, G., Bigda, J., Sadej, R., Zaczek, A., & Markiewicz, A. (2019). Clinical and Biological Significance of ESR1 Gene Alteration and Estrogen Receptors Isoforms Expression in Breast Cancer Patients. International Journal of Molecular Sciences, 20(8), 1881. https://doi.org/10.3390/ijms20081881