Inhibition of p300/CBP-Associated Factor Attenuates Renal Tubulointerstitial Fibrosis through Modulation of NF-kB and Nrf2

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
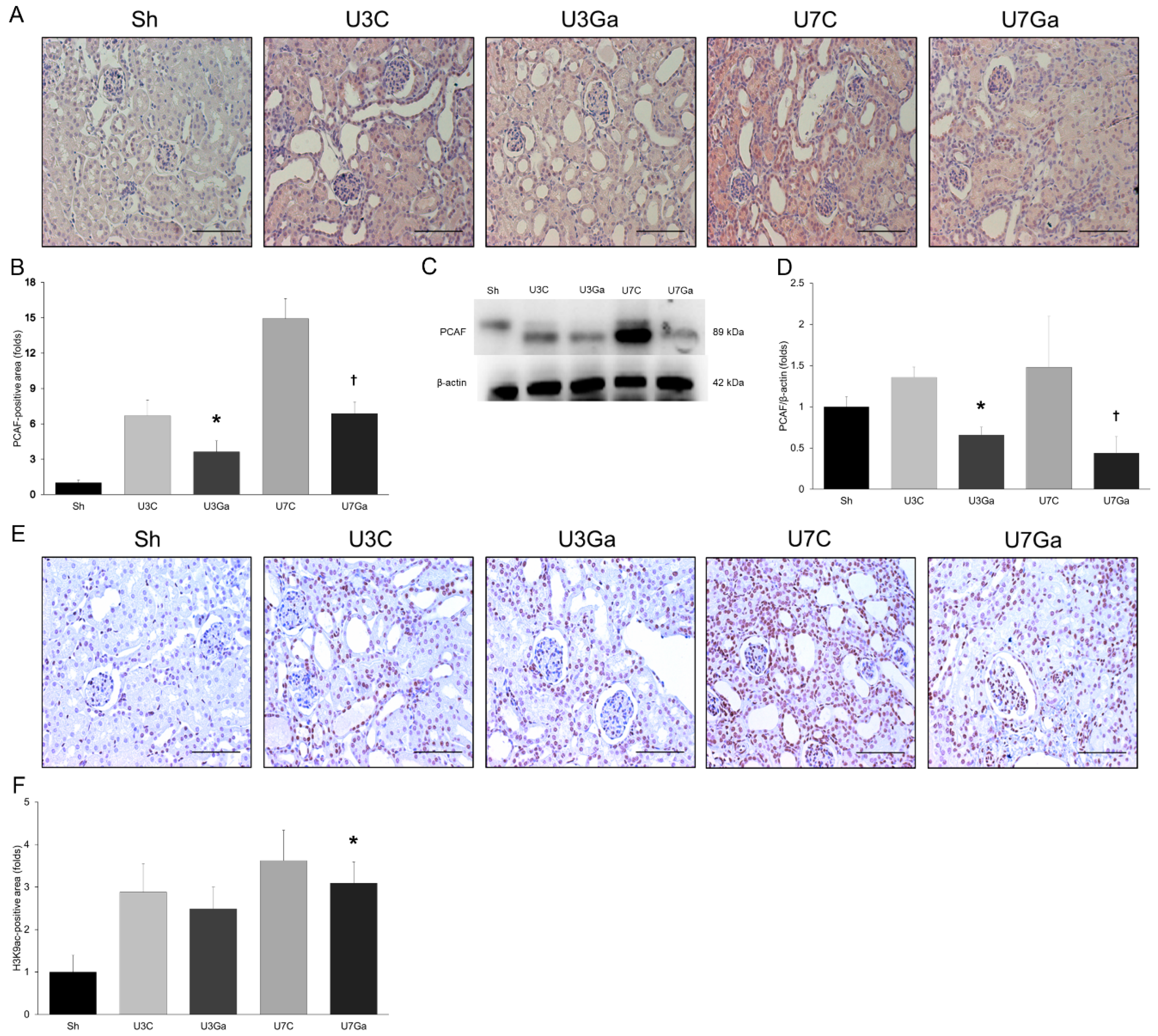
2.1. Expression Levels of PCAF and Histone Acetylation in Kidneys Are Increased after UUO
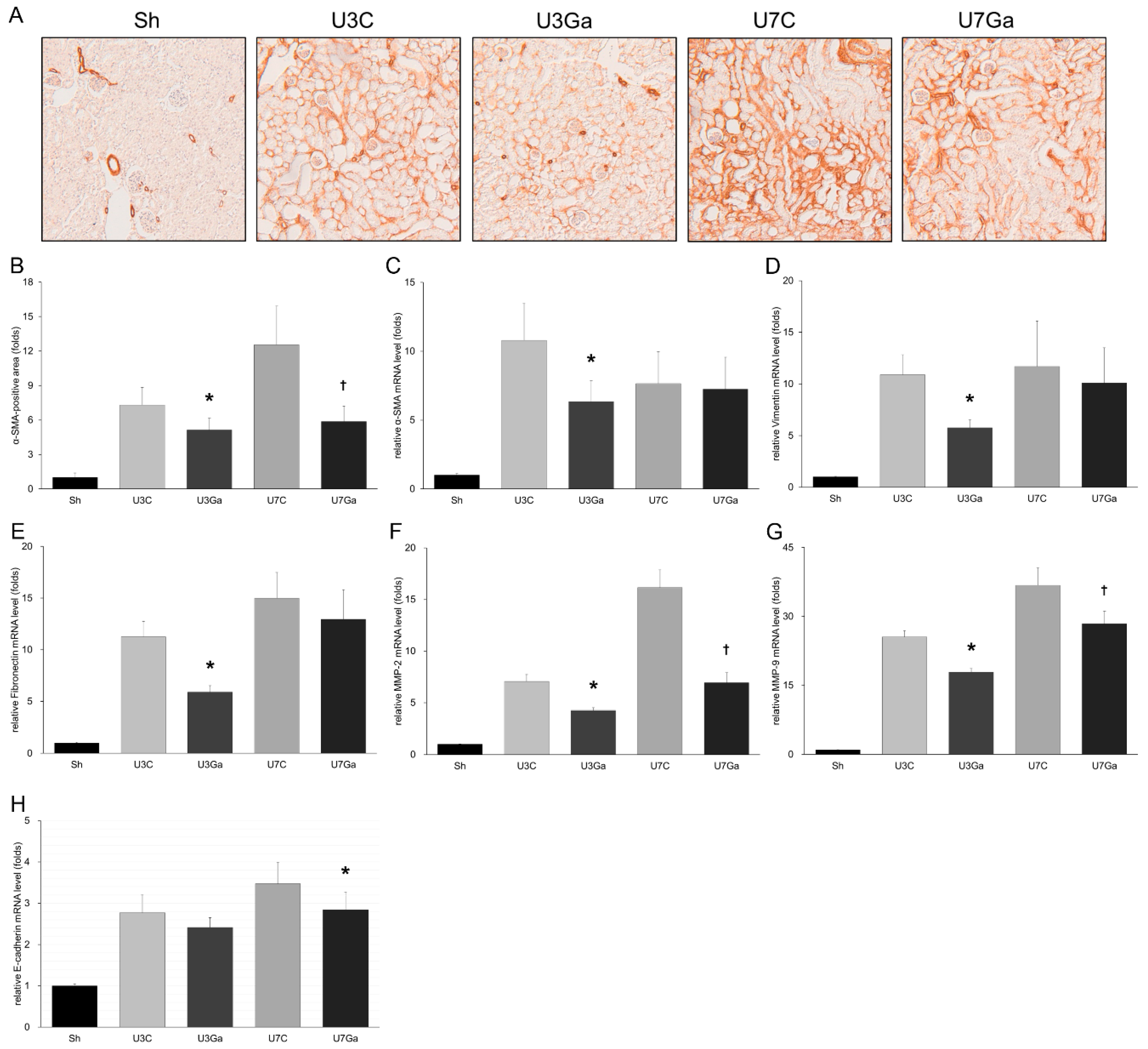
2.2. Inhibition of PCAF Leads to Attenuation of Fibrotic Changes in Obstructed Kidneys
2.3. Inhibition of PCAF Debilitates Epithelial-Mesenchymal Transition in Obstructed Kidneys
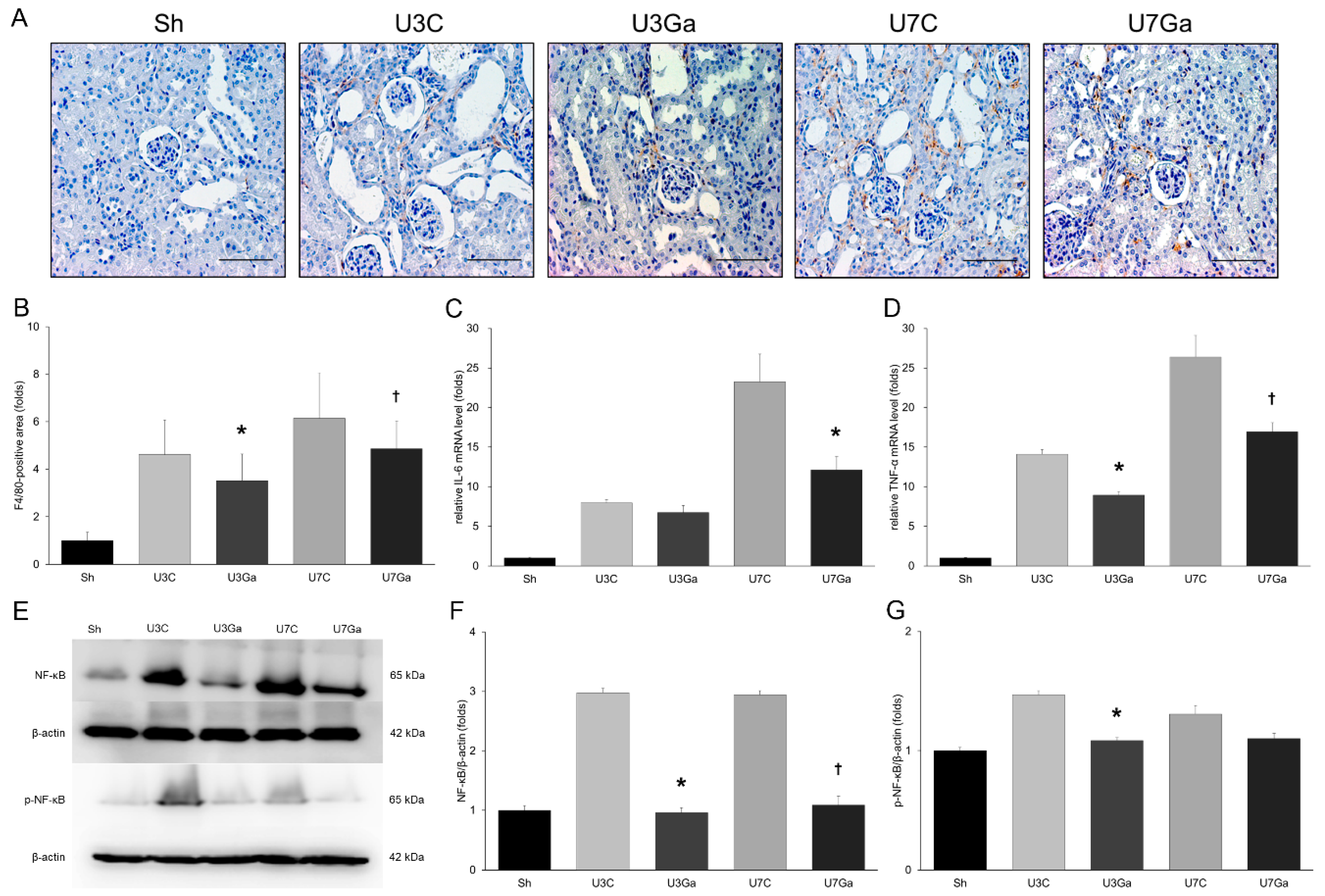
2.4. Inhibition of PCAF Reduces Inflammation in Obstructed Kidneys
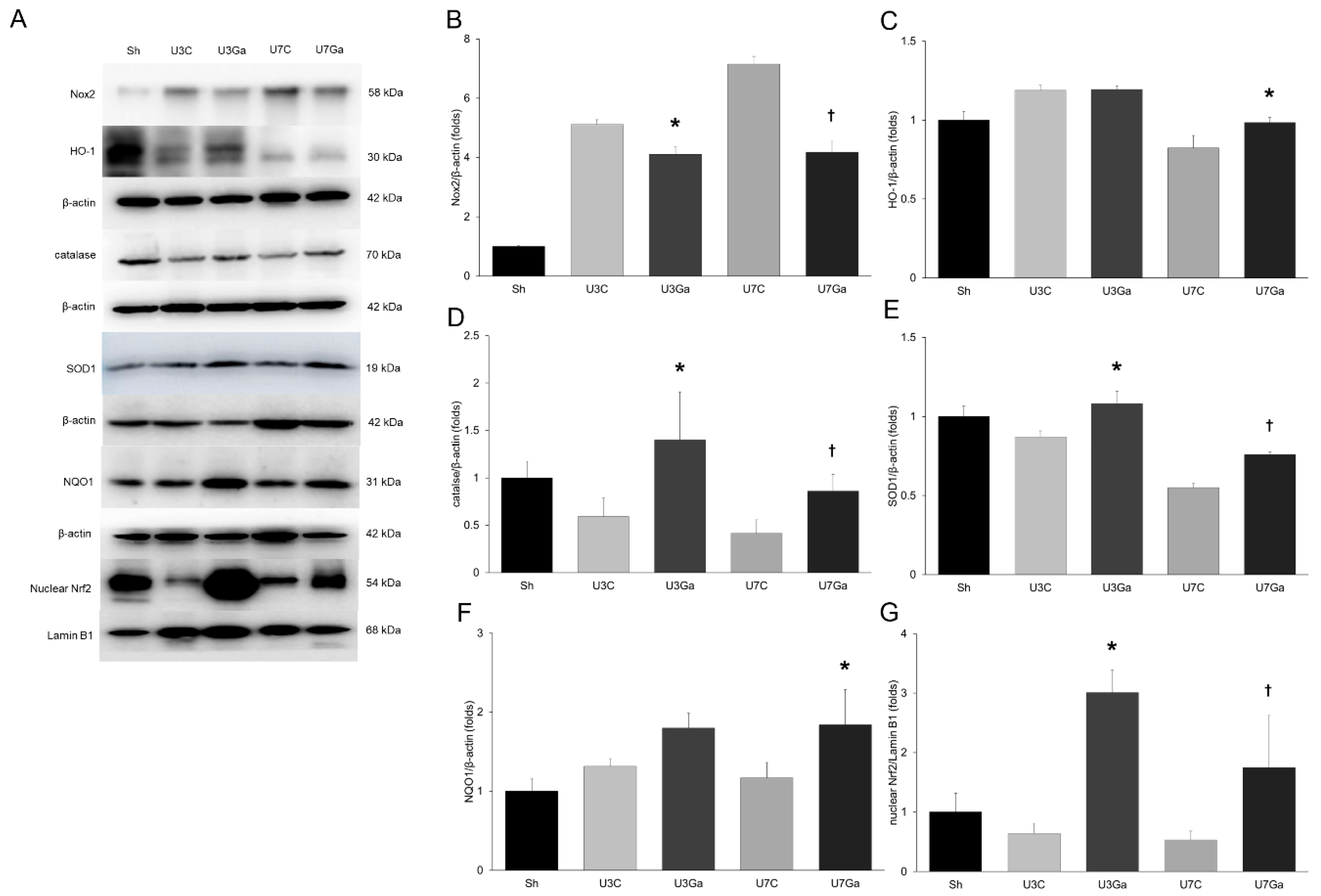
2.5. Inhibition of PCAF Decreases Oxidative Stress and Increases Antioxidant Enzymes in Obstructed Kidneys
2.6. Inhibition of PCAF Decreases Oxidative Stress and Increases Antioxidant Enzymes in Obstructed Kidneys
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histology and Immunohistochemistry
4.3. Renal Collagen Content Assay
4.4. Western Blot Analysis
4.5. Quantitative Real-Time Polymerase Chain Reaction
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ARE | antioxidant response element |
| CBP | CREB binding protein |
| CKD | chronic kidney disease |
| Col IV | collagen IV |
| EMT | epithelial-mesenchymal transition |
| FSP-1 | fibroblast-specific protein-1 |
| GNAT | Gcn5-related N-acetyltransferase |
| H | histone |
| HAT | histone acetyltransferase |
| HDAC | histone deacetylase |
| HO-1 | heme oxygenase-1 |
| IL-1β | interleukin-1β |
| IL-6 | Interleukin-6 |
| IκBα | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, α |
| K | Lysine |
| Keap1 | Kelch-like ECH-associated protein 1 |
| LPS | lipopolysaccharide |
| MMP-2 | matrix metalloproteinase-2 |
| MMP-9 | matrix metalloproteinase-9 |
| MYST | MOZ, Ybf2 (Sas3), Sas2, and Tip60 |
| NF-κB | nuclear factor-κB |
| NQO1 | NAD(P)H:quinone oxidoreductase 1 |
| NRC | nuclear coactivators |
| Nrf2 | nuclear factor erythroid-derived 2-like factor 2 |
| PCAF | p300/CBP-associated factor |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| ROS | reactive oxygen species |
| SOD1 | superoxide dismutase 1 |
| TGF-β | transforming growth factor-β |
| TNF-α | tumor necrosis factor-α |
| TUNEL | terminal deoxynucleotidyl transferase dUTP nick end labeling |
| UUO | unilateral ureteral obstruction |
| VE-cadherin | vascular endothelial cadherin |
| α-SMA | α-smooth muscle actin |
References
- Gao, Y.; Tollefsbol, T.O. Impact of Epigenetic Dietary Components on Cancer through Histone Modifications. Curr. Med. Chem. 2015, 22, 2051–2064. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Gaikwad, A.B. Compound 21 and Telmisartan combination mitigates type 2 diabetic nephropathy through amelioration of caspase mediated apoptosis. Biochem. Biophys. Res. Commun. 2017, 487, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Thorsheim, K.; Persson, A.; Siegbahn, A.; Tykesson, E.; Westergren-Thorsson, G.; Mani, K.; Ellervik, U. Disubstituted naphthyl β-d-xylopyranosides: Synthesis, GAG priming, and histone acetyltransferase (HAT) inhibition. Glycoconj. J. 2016, 33, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.P.; Bachli, E.B.; Schaer, D.J.; Schoedon, G. Transcriptome analysis revealed unique genes as targets for the anti-inflammatory action of activated protein C in human macrophages. PLoS ONE 2010, 5, e15352. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Ai, Q.; Lin, L.; Dai, J.; Jia, M.; Zhou, D.; Che, Q.; Wan, J.; Jiang, R.; Zhang, L. Protective effects of garcinol in mice with lipopolysaccharide/D-galactosamine-induced apoptotic liver injury. Int. Immunopharmacol. 2014, 19, 373–380. [Google Scholar] [CrossRef]
- Ghizzoni, M.; Boltjes, A.; Graaf, C.D.; Haisma, H.J.; Dekker, F.J. Improved inhibition of the histone acetyltransferase PCAF by an anacardic acid derivative. Bioorg. Med. Chem. 2010, 18, 5826–5834. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, Y.H.; Seong, A.R.; Lee, J.; Jun, W.; Yoon, H.G. Selective inhibition of PCAF suppresses microglial-mediated β-amyloid neurotoxicity. Int. J. Mol. Med. 2013, 32, 469–475. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Jin, C.; Zhang, M.; Lv, L.; Zhang, X.; Liu, H.; Zhou, Y. Histone H3K9 Acetyltransferase PCAF Is Essential for Osteogenic Differentiation Through Bone Morphogenetic Protein Signaling and May Be Involved in Osteoporosis. Stem Cells 2016, 34, 2332–2341. [Google Scholar] [CrossRef]
- Zhao, J.; Gong, A.Y.; Zhou, R.; Liu, J.; Eischeid, A.N.; Chen, X.M. Downregulation of PCAF by miR-181a/b provides feedback regulation to TNF-α-induced transcription of proinflammatory genes in liver epithelial cells. J. Immunol. 2012, 188, 1266–1274. [Google Scholar] [CrossRef]
- Lee, S.B.; Kalluri, R. Mechanistic connection between inflammation and fibrosis. Kidney Int. Suppl. 2010, S22–S26. [Google Scholar] [CrossRef]
- Huang, J.; Wan, D.; Li, J.; Chen, H.; Huang, K.; Zheng, L. Histone acetyltransferase PCAF regulates inflammatory molecules in the development of renal injury. Epigenetics 2015, 10, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Kawada, N.; Nagatoya, K.; Takeji, M.; Horio, M.; Ando, A.; Imai, E.; Hori, M. Fluvastatin suppresses oxidative stress and fibrosis in the interstitium of mouse kidneys with unilateral ureteral obstruction. Kidney Int. 2001, 59, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Dendooven, A.; Ishola, D.A., Jr.; Nguyen, T.Q.; Van der Giezen, D.M.; Kok, R.J.; Goldschmeding, R.; Joles, J.A. Oxidative stress in obstructive nephropathy. Int. J. Exp. Pathol. 2011, 92, 202–210. [Google Scholar] [CrossRef]
- Hung, T.W.; Tsai, J.P.; Lin, S.H.; Lee, C.H.; Hsieh, Y.H.; Chang, H.R. Pentraxin 3 Activates JNK Signaling and Regulates the Epithelial-To-Mesenchymal Transition in Renal Fibrosis. Cell. Physiol. Biochem. 2016, 40, 1029–1038. [Google Scholar] [CrossRef]
- Babelova, A.; Avaniadi, D.; Jung, O.; Fork, C.; Beckmann, J.; Kosowski, J.; Weissmann, N.; Anilkumar, N.; Shah, A.M.; Schaefer, L.; et al. Role of Nox4 in murine models of kidney disease. Free Radic. Biol. Med. 2012, 53, 842–853. [Google Scholar] [CrossRef]
- Chung, S.; Kim, S.; Kim, M.; Koh, E.S.; Yoon, H.E.; Kim, H.S.; Park, C.W.; Chang, Y.S.; Shin, S.J. T-type calcium channel blocker attenuates unilateral ureteral obstruction-induced renal interstitial fibrosis by activating the Nrf2 antioxidant pathway. Am. J. Transl. Res. 2016, 8, 4574–4585. [Google Scholar] [PubMed]
- Steger, D.J.; Workman, J.L. Remodeling chromatin structures for transcription: What happens to the histones? Bioessays 1996, 18, 875–884. [Google Scholar] [CrossRef]
- Vanden Berghe, W.; De Bosscher, K.; Boone, E.; Plaisance, S.; Haegeman, G. The nuclear factor-kappaB engages CBP/p300 and histone acetyltransferase activity for transcriptional activation of the interleukin-6 gene promoter. J. Biol. Chem. 1999, 274, 32091–32098. [Google Scholar] [CrossRef]
- Sheppard, K.A.; Rose, D.W.; Haque, Z.K.; Kurokawa, R.; McInerney, E.; Westin, S.; Thanos, D.; Rosenfeld, M.G.; Glass, C.K.; Collins, T. Transcriptional activation by NF-kappaB requires multiple coactivators. Mol. Cell. Biol. 1999, 19, 6367–6378. [Google Scholar] [CrossRef]
- Mittal, R.; Peak-Chew, S.Y.; McMahon, H.T. Acetylation of MEK2 and I kappa B kinase (IKK) activation loop residues by YopJ inhibits signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 18574–18579. [Google Scholar] [CrossRef]
- Jung, K.J.; Min, K.J.; Park, J.W.; Park, K.M.; Kwon, T.K. Carnosic acid attenuates unilateral ureteral obstruction-induced kidney fibrosis via inhibition of Akt-mediated Nox4 expression. Free Radic. Biol. Med. 2016, 97, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Sedeek, M.; Nasrallah, R.; Touyz, R.M.; Hébert, R.L. NADPH oxidases, reactive oxygen species, and the kidney: Friend and foe. J. Am. Soc. Nephrol. 2013, 24, 1512–1518. [Google Scholar] [CrossRef]
- Rhee, E.P. NADPH Oxidase 4 at the Nexus of Diabetes, Reactive Oxygen Species, and Renal Metabolism. J. Am. Soc. Nephrol. 2016, 27, 337–339. [Google Scholar] [CrossRef]
- Kim, J.I.; Noh, M.R.; Kim, K.Y.; Jang, H.S.; Kim, H.Y.; Park, K.M. Methionine sulfoxide reductase A deficiency exacerbates progression of kidney fibrosis induced by unilateral ureteral obstruction. Free Radic. Biol. Med. 2015, 89, 201–208. [Google Scholar] [CrossRef]
- Chung, S.; Yoon, H.E.; Kim, S.J.; Kim, S.J.; Koh, E.S.; Hong, Y.A.; Park, C.W.; Chang, Y.S.; Shin, S.J. Oleanolic acid attenuates renal fibrosis in mice with unilateral ureteral obstruction via facilitating nuclear translocation of Nrf2. Nutr. Metab. 2014, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, B.; Du, F.; Su, X.; Sun, G.; Zhou, G.; Bian, X.; Liu, N. Epigallocatechin-3-Gallate Attenuates Oxidative Stress and Inflammation in Obstructive Nephropathy via NF-κB and Nrf2/HO-1 Signalling Pathway Regulation. Basic Clin. Pharmacol. Toxicol. 2015, 117, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Bolati, D.; Shimizu, H.; Yisireyili, M.; Nishijima, F.; Niwa, T. Indoxyl sulfate, a uremic toxin, downregulates renal expression of Nrf2 through activation of NF-κB. BMC Nephrol. 2015, 14, 56. [Google Scholar] [CrossRef]
- Yu, M.; Li, H.; Liu, Q.; Liu, F.; Tang, L.; Li, C.; Yuan, Y.; Zhan, Y.; Xu, W.; Li, W.; et al. Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell Signal. 2011, 23, 883–892. [Google Scholar] [CrossRef]
- Pedruzzi, L.M.; Stockler-Pinto, M.B.; Leite, M., Jr.; Mafra, D. Nrf2-keap1 system versus NF-κB: The good and the evil in chronic kidney disease? Biochimie 2012, 94, 2461–2466. [Google Scholar] [CrossRef]
- Hu, J.; Zhu, Q.; Li, P.L.; Wang, W.; Yi, F.; Li, N. Stem cell conditioned culture media attenuated albumin-induced epithelial-mesenchymal transition in renal tubular cells. Cell. Physiol. Biochem. 2015, 35, 1719–1728. [Google Scholar] [CrossRef]
- Bai, Y.; Lu, H.; Lin, C.; Xu, Y.; Hu, D.; Liang, Y.; Hong, W.; Chen, B. Sonic hedgehog-mediated epithelial-mesenchymal transition in renal tubulointerstitial fibrosis. Int. J. Mol. Med. 2016, 37, 1317–1427. [Google Scholar] [CrossRef]
- Kaufhold, S.; Bonavida, B. Central role of Snail1 in the regulation of EMT and resistance in cancer: A target for therapeutic intervention. J. Exp. Clin. Cancer Res. 2014, 33, 62. [Google Scholar] [CrossRef] [PubMed]
- Lange-Sperandio, B.; Trautmann, A.; Eickelberg, O.; Jayachandran, A.; Oberle, S.; Schmidutz, F.; Rodenbeck, B.; Hömme, M.; Horuk, R.; Schaefer, F. Leukocytes induce epithelial to mesenchymal transition after unilateral ureteral obstruction in neonatal mice. Am. J. Pathol. 2007, 171, 861–871. [Google Scholar] [CrossRef]
- Rhee, H.; Han, M.; Kim, S.S.; Kim, I.Y.; Lee, H.W.; Bae, S.S.; Ha, H.K.; Jung, E.S.; Lee, M.Y.; Seong, E.Y.; et al. The expression of two isoforms of matrix metalloproteinase-2 in aged mouse models of diabetes mellitus and chronic kidney disease. Kidney Res. Clin. Pract. 2018, 37, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.A.; Park, J.T.; Natarajan, R. Epigenetic modifications and diabetic nephropathy. Kidney Res. Clin. Pract. 2012, 31, 139–150. [Google Scholar] [CrossRef]
- Ning, X.H.; Ge, X.F.; Cui, Y.; An, H.X. Ulinastatin inhibits unilateral ureteral obstruction-induced renal interstitial fibrosis in rats via transforming growth factor β (TGF-β)/Smad signalling pathways. Int. Immunopharmacol. 2013, 15, 406–413. [Google Scholar] [CrossRef]
- Wu, W.P.; Chang, C.H.; Chiu, Y.T.; Ku, C.L.; Wen, M.C.; Shu, K.H.; Wu, M.J. A reduction of unilateral ureteral obstruction-induced renal fibrosis by a therapy combining valsartan with aliskiren. Am. J. Physiol. Ren. Physiol. 2010, 299, F929–F941. [Google Scholar] [CrossRef]
- Forbes, M.S.; Thornhill, B.A.; Minor, J.J.; Gordon, K.A.; Galarreta, C.I.; Chevalier, R.L. Fight-or-flight: Murine unilateral ureteral obstruction causes extensive proximal tubular degeneration, collecting duct dilatation, and minimal fibrosis. Am. J. Physiol. Ren. Physiol. 2012, 303, F120–F129. [Google Scholar] [CrossRef] [PubMed]
- Chaabane, W.; Praddaude, F.; Buleon, M.; Jaafar, A.; Vallet, M.; Rischmann, P.; Galarreta, C.I.; Chevalier, R.L.; Tack, I. Renal functional decline and glomerulotubular injury are arrested but not restored by release of unilateral ureteral obstruction (UUO). Am. J. Physiol. Ren. Physiol. 2013, 304, F432–F439. [Google Scholar] [CrossRef]
- Sethi, G.; Chatterjee, S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Wong, K.F.; Kumar, A.P.; Senapati, P.; Behera, A.K.; Hui, K.M.; et al. Inhibition of STAT3 dimerization and acetylation by garcinol suppresses the growth of human hepatocellular carcinoma in vitro and in vivo. Mol. Cancer 2014, 13, 66. [Google Scholar] [CrossRef]
- Li, F.; Shanmugam, M.K.; Siveen, K.S.; Wang, F.; Ong, T.H.; Loo, S.Y.; Swamy, M.M.; Mandal, S.; Kumar, A.P.; Goh, B.C.; et al. Garcinol sensitizes human head and neck carcinoma to cisplatin in a xenograft mouse model despite downregulation of proliferative biomarkers. Oncotarget 2015, 6, 5147–5163. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, S.; Kim, S.; Son, M.; Kim, M.; Koh, E.S.; Shin, S.J.; Park, C.W.; Kim, H.-S. Inhibition of p300/CBP-Associated Factor Attenuates Renal Tubulointerstitial Fibrosis through Modulation of NF-kB and Nrf2. Int. J. Mol. Sci. 2019, 20, 1554. https://doi.org/10.3390/ijms20071554
Chung S, Kim S, Son M, Kim M, Koh ES, Shin SJ, Park CW, Kim H-S. Inhibition of p300/CBP-Associated Factor Attenuates Renal Tubulointerstitial Fibrosis through Modulation of NF-kB and Nrf2. International Journal of Molecular Sciences. 2019; 20(7):1554. https://doi.org/10.3390/ijms20071554
Chicago/Turabian StyleChung, Sungjin, Soojeong Kim, Mina Son, Minyoung Kim, Eun Sil Koh, Seok Joon Shin, Cheol Whee Park, and Ho-Shik Kim. 2019. "Inhibition of p300/CBP-Associated Factor Attenuates Renal Tubulointerstitial Fibrosis through Modulation of NF-kB and Nrf2" International Journal of Molecular Sciences 20, no. 7: 1554. https://doi.org/10.3390/ijms20071554
APA StyleChung, S., Kim, S., Son, M., Kim, M., Koh, E. S., Shin, S. J., Park, C. W., & Kim, H.-S. (2019). Inhibition of p300/CBP-Associated Factor Attenuates Renal Tubulointerstitial Fibrosis through Modulation of NF-kB and Nrf2. International Journal of Molecular Sciences, 20(7), 1554. https://doi.org/10.3390/ijms20071554