Mechanisms of FSH- and Amphiregulin-Induced MAP Kinase 3/1 Activation in Pig Cumulus-Oocyte Complexes During Maturation In Vitro


Abstract
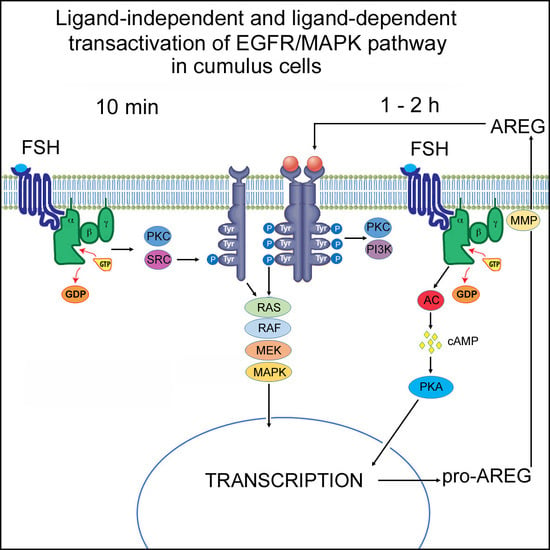
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
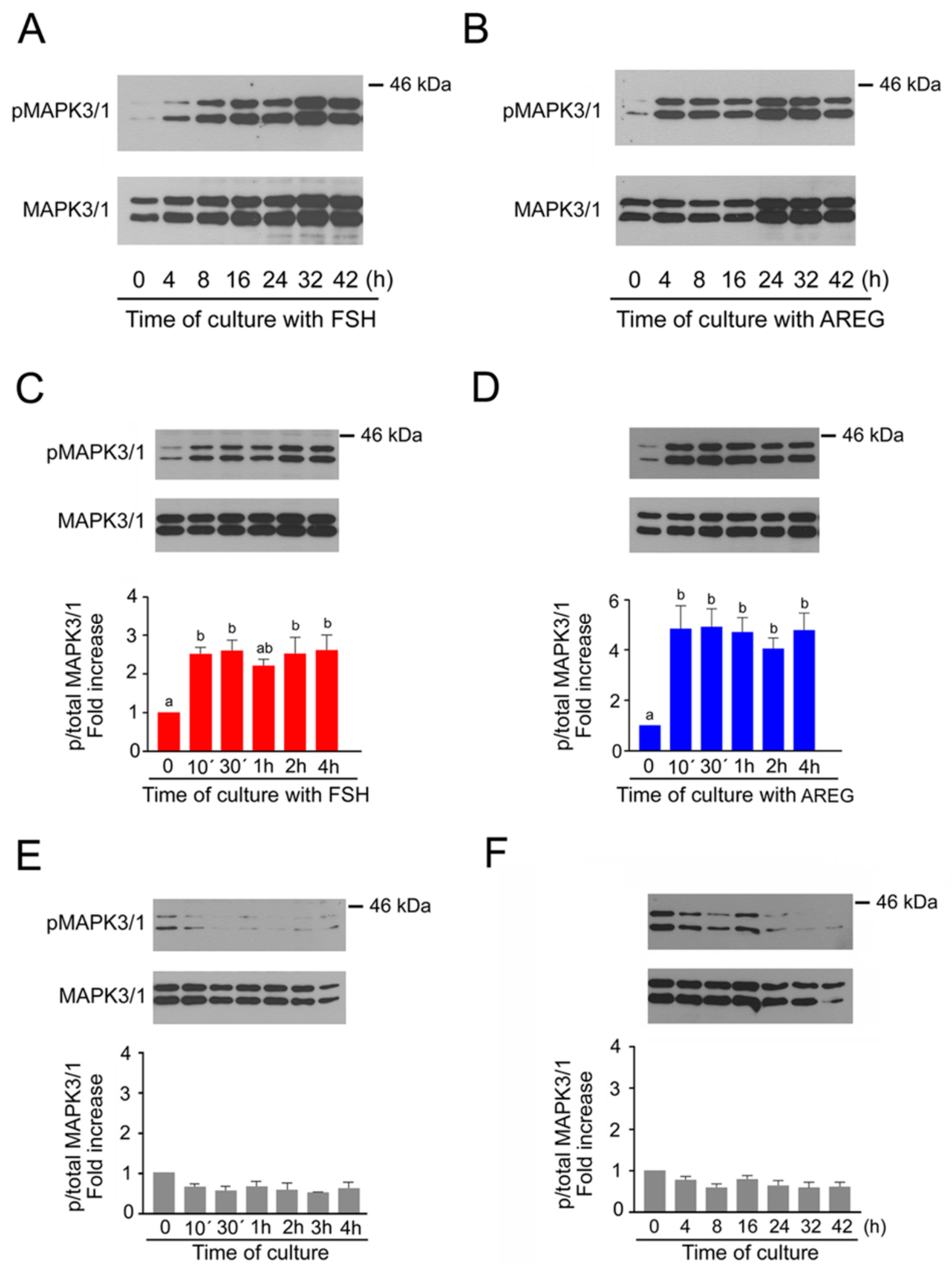
2.1. Time Course of MAPK3/1 Activation in Pig Cumulus-Enclosed Oocytes (COCs) Cultured in Medium Supplemented with Follicle-Stimulating Hormone (FSH) or Amphiregulin (AREG)
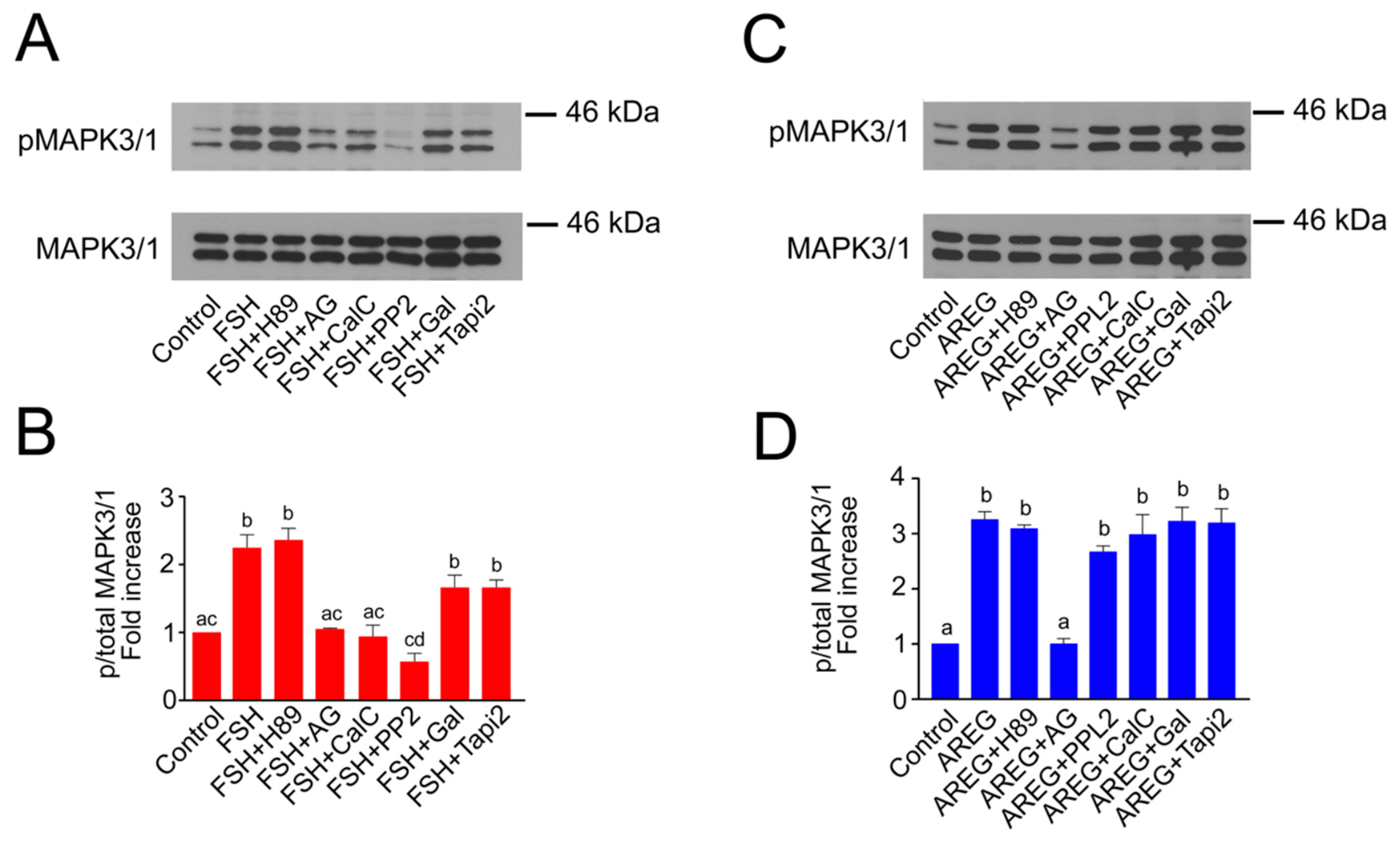
2.2. Molecular Mechanisms of FSH- and AREG-Induced Rapid Activation of MAPK3/1 in Pig COCs
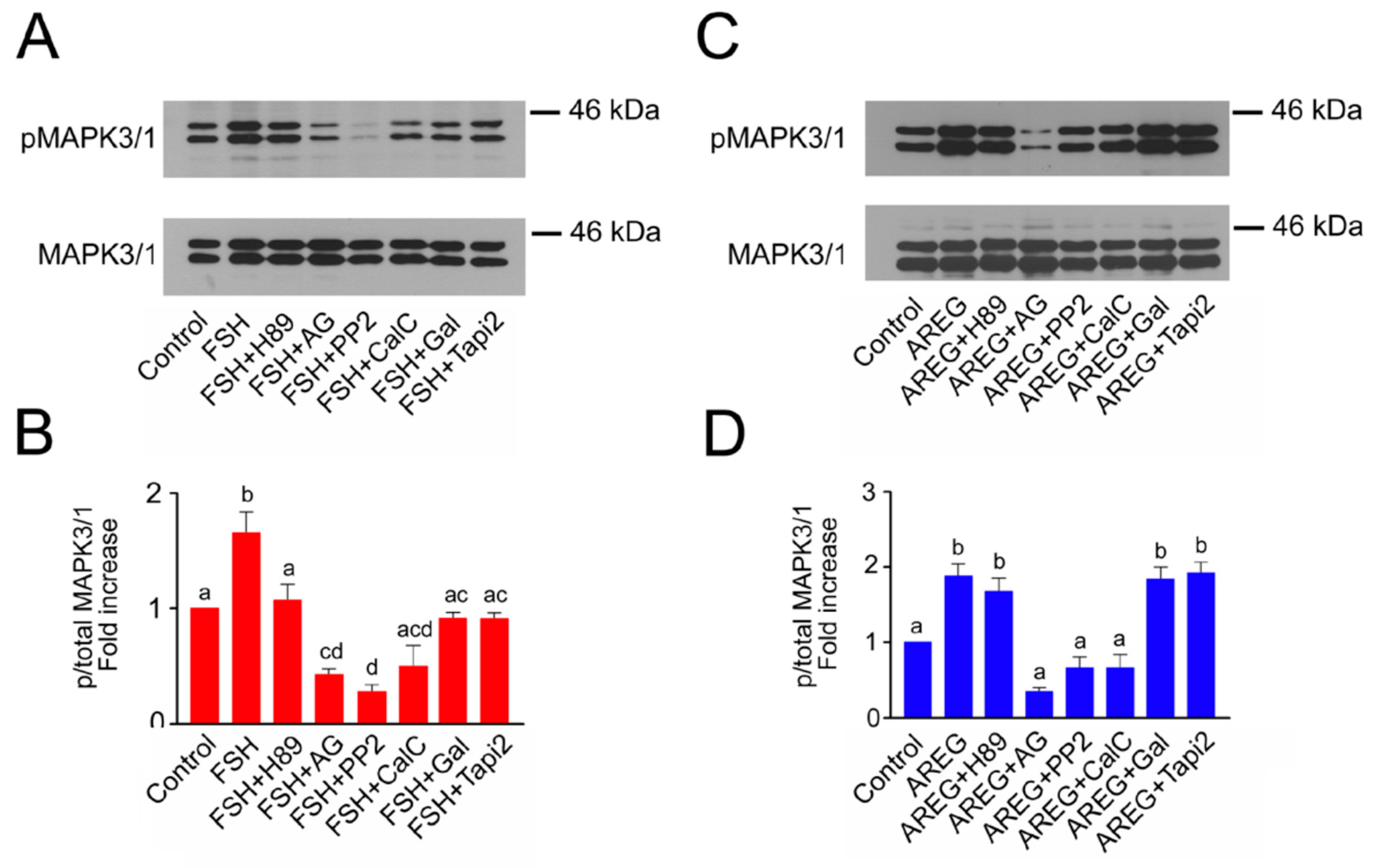
2.3. Molecular Mechanisms Involved in Maintenance of FSH- and AREG-Induced Activation of MAPK3/1
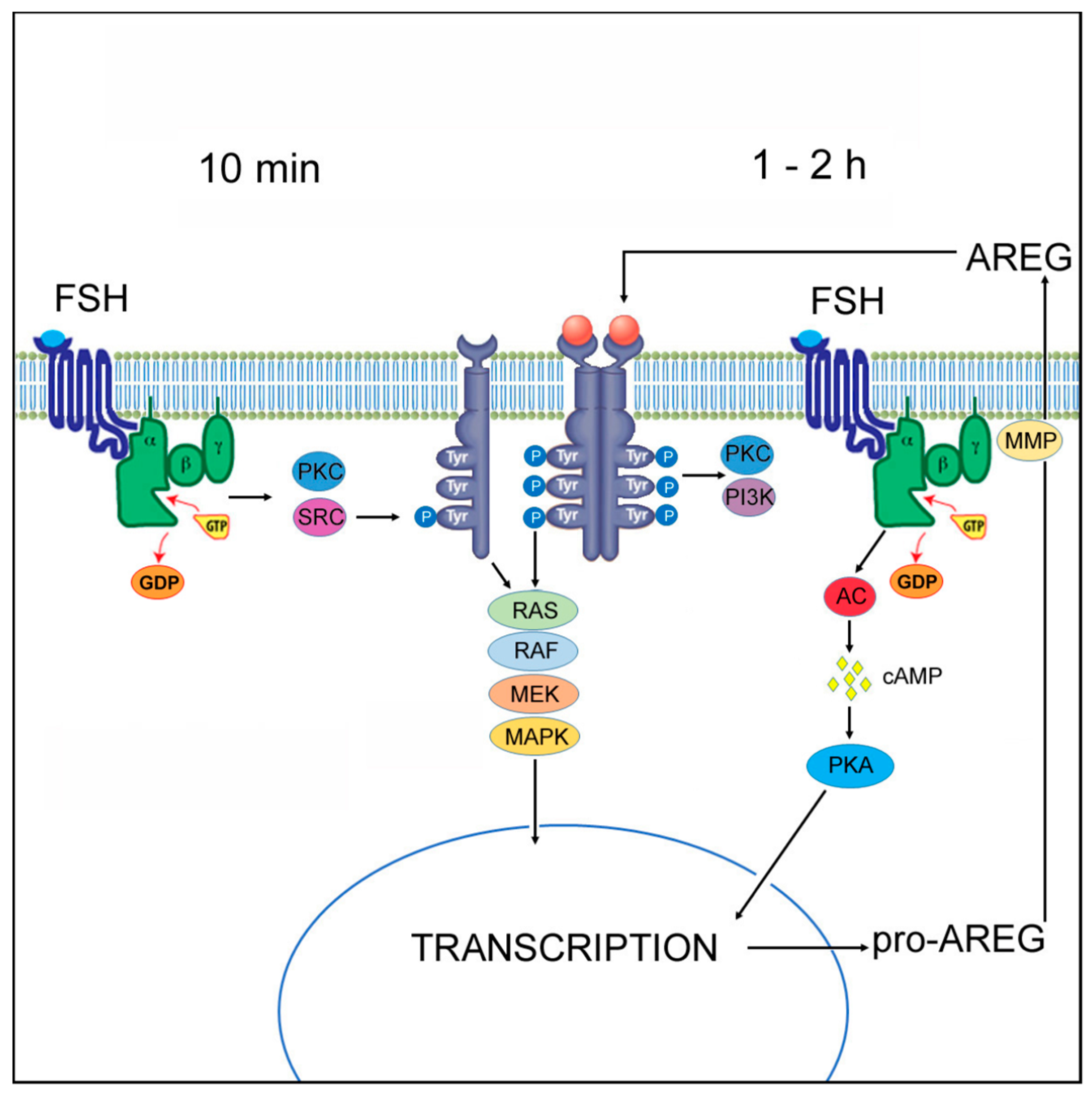
3. Discussion
4. Materials and Methods
4.1. Culture Media and Reagents
4.2. Collection of Cumulus-Oocyte Complexes
4.3. Culture of Cumulus-Oocyte Complexes In Vitro
4.4. Immunoblotting
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Posada, J.; Yew, N.; Ahn, N.G.; Vande Woude, G.F.; Cooper, J.A. Mos stimulates MAP kinase in Xenopus oocytes and activates a MAP kinase kinase in vitro. Mol. Cell. Biol. 1993, 13, 2546–2553. [Google Scholar] [CrossRef] [PubMed]
- Verlhac, M.H.; Lefebvre, C.; Kubiak, J.Z.; Umbhauer, M.; Rassinier, P.; Colledge, W.; Maro, B. Mos activates MAP kinase in mouse oocytes through two opposite pathways. EMBO J. 2000, 19, 6065–6074. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wigglesworth, K.; Pendola, F.L.; O’Brien, M.J.; Eppig, J.J. Mitogen-activated protein kinase activity in cumulus cells is essential for gonadotropin-induced oocyte meiotic resumption and cumulus expansion in the mouse. Endocrinology 2002, 143, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.Q.; Denegre, J.M.; Wigglesworth, K.; Pendola, F.L.; O’Brien, M.J.; Eppig, J.J. Oocyte-dependent activation of mitogen-activated protein kinase (ERK1/2) in cumulus cells is required for the maturation of the mouse oocyte-cumulus cell complex. Dev. Biol. 2003, 263, 126–138. [Google Scholar] [CrossRef]
- Siddappa, D.; Beaulieu, É.; Gévry, N.; Roux, P.P.; Bordignon, V.; Duggavathi, R. Effect of the transient pharmacological inhibition of Mapk3/1 pathway on ovulation in mice. PLoS ONE 2015, 10, e0119387. [Google Scholar] [CrossRef]
- Meinecke, B.; Krischek, C. MAPK/ERK kinase (MEK) signalling is required for resumption of meiosis in cultured cumulus-enclosed pig oocytes. Zygote 2003, 11, 7–16. [Google Scholar] [CrossRef]
- Prochazka, R.; Blaha, M.; Nemcova, L. Signaling pathways regulating FSH- and amphiregulin-induced meiotic resumption and cumulus cell expansion in the pig. Reproduction 2012, 144, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Liu, Z.; Shimada, M.; Sterneck, E.; Johnson, P.F.; Hedrick, S.M.; Richards, J.S. MAPK3/1 (ERK1/2) in ovarian granulosa cells are essential for female fertility. Science 2009, 324, 938–941. [Google Scholar] [CrossRef]
- Mehlmann, L.M. Stops and starts in mammalian oocytes: Recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 2005, 130, 791–799. [Google Scholar] [CrossRef]
- Cameron, M.R.; Foster, J.S.; Bukovsky, A.; Wimalasena, J. Activation of mitogen-activated protein kinases by gonadotropins and cyclic adenosine 5′-monophosphates in porcine granulosa cells. Biol. Reprod. 1996, 55, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Su, Y.Q.; Ariga, M.; Law, E.; Jin, S.L.; Conti, M. EGF-like growth factors as mediators of LH action in the ovulatory follicle. Science 2004, 303, 682–684. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, H.; Cao, X.; Motola, S.; Popliker, M.; Conti, M.; Tsafriri, A. Epidermal growth factor family members: Endogenous mediators of the ovulatory response. Endocrinology 2005, 46, 77–84. [Google Scholar] [CrossRef]
- Panigone, S.; Hsieh, M.; Fu, M.; Persani, L.; Conti, M. Luteinizing hormone signaling in preovulatory follicles involves early activation of the epidermal growth factor receptor pathway. Mol. Endocrinol. 2008, 22, 924–936. [Google Scholar] [CrossRef]
- Wayne, C.M.; Fan, H.Y.; Cheng, X.; Richards, J.S. Follicle-stimulating hormone induces multiple signaling cascades: Evidence that activation of Rous sarcoma oncogene, RAS, and the epidermal growth factor receptor are critical for granulosa cell differentiation. Mol. Endocrinol. 2007, 21, 1940–1957. [Google Scholar] [CrossRef] [PubMed]
- Sela-Abramovich, S.; Chorev, E.; Galiani, D.; Dekel, N. Mitogen-activated protein kinase mediates luteinizing hormone-induced breakdown of communication and oocyte maturation in rat ovarian follicles. Endocrinology 2005, 146, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, S.; Naito, K.; Sugiura, K.; Iwamori, N.; Goto, S.; Naruoka, H.; Tojo, H. Analyses of mitogen-activated protein kinase function in the maturation of porcine oocytes. Biol. Reprod. 2003, 68, 604–609. [Google Scholar] [CrossRef]
- Ebeling, S.; Schuon, C.; Meinecke, B. Mitogen-activated protein kinase phosphorylation patterns in pig oocytes and cumulus cells during gonadotrophin-induced resumption of meiosis in vitro. Zygote 2007, 15, 139–147. [Google Scholar] [CrossRef]
- Nemcova, L.; Nagyova, E.; Petlach, M.; Tomanek, M.; Prochazka, R. Molecular mechanisms of insulin-like growth factor 1 promoted synthesis and retention of hyaluronic acid in porcine oocyte-cumulus complexes. Biol. Reprod. 2007, 76, 1016–1024. [Google Scholar] [CrossRef]
- Yamashita, Y.; Okamoto, M.; Kawashima, I.; Okazaki, T.; Nishimura, R.; Gunji, Y.; Hishinuma, M.; Shimada, M. Positive feedback loop between prostaglandin E2 and EGF-like factors is essential for sustainable activation of MAPK3/1 in cumulus cells during in vitro maturation of porcine cumulus oocyte complexes. Biol. Reprod. 2011, 85, 1073–1082. [Google Scholar] [CrossRef]
- Prochazka, R.; Blaha, M. Regulation of mitogen-activated protein kinase 3/1 activity during meiosis resumption in mammals. J. Reprod. Dev. 2015, 61, 495–502. [Google Scholar] [CrossRef]
- Salhab, M.; Dhorne-Pollet, S.; Auclair, S.; Guyader-Joly, C.; Brisard, D.; Dalbies-Tran, R.; Dupont, J.; Ponsart, C.; Mermillod, P.; Uzbekova, S. In vitro maturation of oocytes alters gene expression and signaling pathways in bovine cumulus cells. Mol. Reprod. Dev. 2013, 80, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Reddy, R.J.; Gajadhar, A.S.; Swenson, E.J.; Rothenberg, D.A.; Curran, T.G.; White, F.M. Early signaling dynamics of the epidermal growth factor receptor. Proc. Natl. Acad. Sci. USA 2016, 113, 3114–3119. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Torcia, S.; Xie, F.; Lin, C.J.; Cakmak, H.; Franciosi, F.; Horner, K.; Onodera, C.; Song, J.S.; Cedars, M.I.; et al. Somatic cells regulate maternal mRNA translation and developmental competence of mouse oocytes. Nat. Cell Biol. 2013, 15, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, F.; Manandhar, S.; Conti, M. FSH regulates mRNA translation in mouse oocytes and promotes developmental competence. Endocrinology 2016, 157, 872–882. [Google Scholar] [CrossRef]
- Khan, D.R.; Guillemette, C.; Sirard, M.A.; Richard, F.J. Characterization of FSH signalling networks in bovine cumulus cells: A perspective on oocyte competence acquisition. Mol. Hum. Reprod. 2015, 21, 688–701. [Google Scholar] [CrossRef]
- Wang, Z. Transactivation of Epidermal Growth Factor Receptor by G Protein-Coupled Receptors: Recent Progress, Challenges and Future Research. Int. J. Mol. Sci. 2016, 17, E95. [Google Scholar] [CrossRef]
- Tatosyan, A.G.; Mizenina, O.A. Kinases of the Src family: Structure and functions. Biochemistry 2000, 65, 49–58. [Google Scholar]
- Liebmann, C. EGF receptor activation by GPCRs: An universal pathway reveals different versions. Mol. Cell. Endocrinol. 2011, 331, 222–231. [Google Scholar] [CrossRef]
- George, A.J.; Hannan, R.D.; Thomas, W.G. Unravelling the molecular complexity of GPCR-mediated EGFR transactivation using functional genomics approaches. FEBS J. 2013, 280, 5258–5268. [Google Scholar] [CrossRef]
- Cattaneo, F.; Guerra, G.; Parisi, M.; De Marinis, M.; Tafuri, D.; Cinelli, M.; Ammendola, R. Cell-surface receptors transactivation mediated by G protein-coupled receptors. Int. J. Mol. Sci. 2014, 15, 19700–19728. [Google Scholar] [CrossRef]
- Drube, S.; Stirnweiss, J.; Valkova, C.; Liebmann, C. Ligand-independent and EGF receptor-supported transactivation: Lessons from beta2-adrenergic receptor signalling. Cell Signal. 2006, 18, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peng, F.F.; Jin, J.; Chen, H.M.; Yu, H.; Zhang, B.F. Src-mediated ligand release-independent EGFR transactivation involves TGF-β-induced Smad3 activation in mesangial cells. Biochem. Biophys. Res. Commun. 2017, 493, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Downs, S.M.; Cottom, J.; Hunzicker-Dunn, M. Protein kinase C and meiotic regulation in isolated mouse oocytes. Mol. Reprod. Dev. 2001, 58, 101–115. [Google Scholar] [CrossRef]
- Fan, H.Y.; Huo, L.J.; Chen, D.Y.; Schatten, H.; Sun, Q.Y. Protein kinase C and mitogen-activated protein kinase cascade in mouse cumulus cells: Cross talk and effect on meiotic resumption of oocyte. Biol. Reprod. 2004, 70, 1178–1187. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, B.; Yan, J.; Xu, B.; Tai, P.; Li, J.; Peng, S.; Zhang, M.; Xia, G. Epidermal growth factor receptor activation by protein kinase C is necessary for FSH-induced meiotic resumption in porcine cumulus-oocyte complexes. J. Endocrinol. 2008, 197, 409–419. [Google Scholar] [CrossRef]
- Yamashita, Y.; Hishinuma, M.; Shimada, M. Activation of PKA, p38 MAPK and ERK1/2 by gonadotropins in cumulus cells is critical for induction of EGF-like factor and TACE/ADAM17 gene expression during in vitro maturation of porcine COCs. J. Ovarian Res. 2009, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Okamoto, M.; Ikeda, M.; Okamoto, A.; Sakai, M.; Gunji, Y.; Nishimura, R.; Hishinuma, M.; Shimada, M. Protein kinase C (PKC) increases TACE/ADAM17 enzyme activity in porcine ovarian somatic cells, which is essential for granulosa cell luteinization and oocyte maturation. Endocrinology 2014, 155, 1080–1090. [Google Scholar] [CrossRef]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G protein regulation of MAPK networks. Oncogene 2007, 26, 3122–3142. [Google Scholar] [CrossRef]
- Shimada, M.; Hernandez-Gonzalez, I.; Gonzalez-Robayna, I.; Richards, J.S. Paracrine and autocrine regulation of epidermal growth factor-like factors in cumulus oocyte complexes and granulosa cells: Key roles for prostaglandin synthase 2 and progesterone receptor. Mol. Endocrinol. 2006, 20, 1352–1365. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Luciano, A.M.; Richani, D.; Zeng, H.T.; Wang, X.; Vos, M.D.; Sugimura, S.; Smitz, J.; Richard, F.J.; Thompson, J.G. Oocyte maturation and quality: Role of cyclic nucleotides. Reproduction 2016, 152, R143–R157. [Google Scholar] [CrossRef]
- Egbert, J.R.; Shuhaibar, L.C.; Edmund, A.B.; Van Helden, D.A.; Robinson, J.W.; Uliasz, T.F.; Baena, V.; Geerts, A.; Wunder, F.; Potter, L.R.; et al. Dephosphorylation and inactivation of NPR2 guanylyl cyclase in granulosa cells contributes to the LH-induced decrease in cGMP that causes resumption of meiosis in rat oocytes. Development 2014, 141, 3594–3604. [Google Scholar] [CrossRef] [PubMed]
- Egbert, J.R.; Uliasz, T.F.; Shuhaibar, L.C.; Geerts, A.; Wunder, F.; Kleiman, R.J.; Humphrey, J.M.; Lampe, P.D.; Artemyev, N.O.; Rybalkin, S.D.; et al. Luteinizing hormone causes phosphorylation and activation of the cGMP phosphodiesterase PDE5 in rat ovarian follicles, contributing, together with PDE1 activity, to the resumption of meiosis. Biol. Reprod. 2016, 94, 110. [Google Scholar] [CrossRef] [PubMed]
- Shuhaibar, L.C.; Egbert, J.R.; Edmund, A.B.; Uliasz, T.F.; Dickey, D.M.; Yee, S.P.; Potter, L.R.; Jaffe, L.A. Dephosphorylation of juxtamembrane serines and threonines of the NPR2 guanylyl cyclase is required for rapid resumption of oocyte meiosis in response to luteinizing hormone. Dev. Biol. 2016, 409, 194–201. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prochazka, R.; Nemcova, L. Mechanisms of FSH- and Amphiregulin-Induced MAP Kinase 3/1 Activation in Pig Cumulus-Oocyte Complexes During Maturation In Vitro. Int. J. Mol. Sci. 2019, 20, 1179. https://doi.org/10.3390/ijms20051179
Prochazka R, Nemcova L. Mechanisms of FSH- and Amphiregulin-Induced MAP Kinase 3/1 Activation in Pig Cumulus-Oocyte Complexes During Maturation In Vitro. International Journal of Molecular Sciences. 2019; 20(5):1179. https://doi.org/10.3390/ijms20051179
Chicago/Turabian StyleProchazka, Radek, and Lucie Nemcova. 2019. "Mechanisms of FSH- and Amphiregulin-Induced MAP Kinase 3/1 Activation in Pig Cumulus-Oocyte Complexes During Maturation In Vitro" International Journal of Molecular Sciences 20, no. 5: 1179. https://doi.org/10.3390/ijms20051179
APA StyleProchazka, R., & Nemcova, L. (2019). Mechanisms of FSH- and Amphiregulin-Induced MAP Kinase 3/1 Activation in Pig Cumulus-Oocyte Complexes During Maturation In Vitro. International Journal of Molecular Sciences, 20(5), 1179. https://doi.org/10.3390/ijms20051179