Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
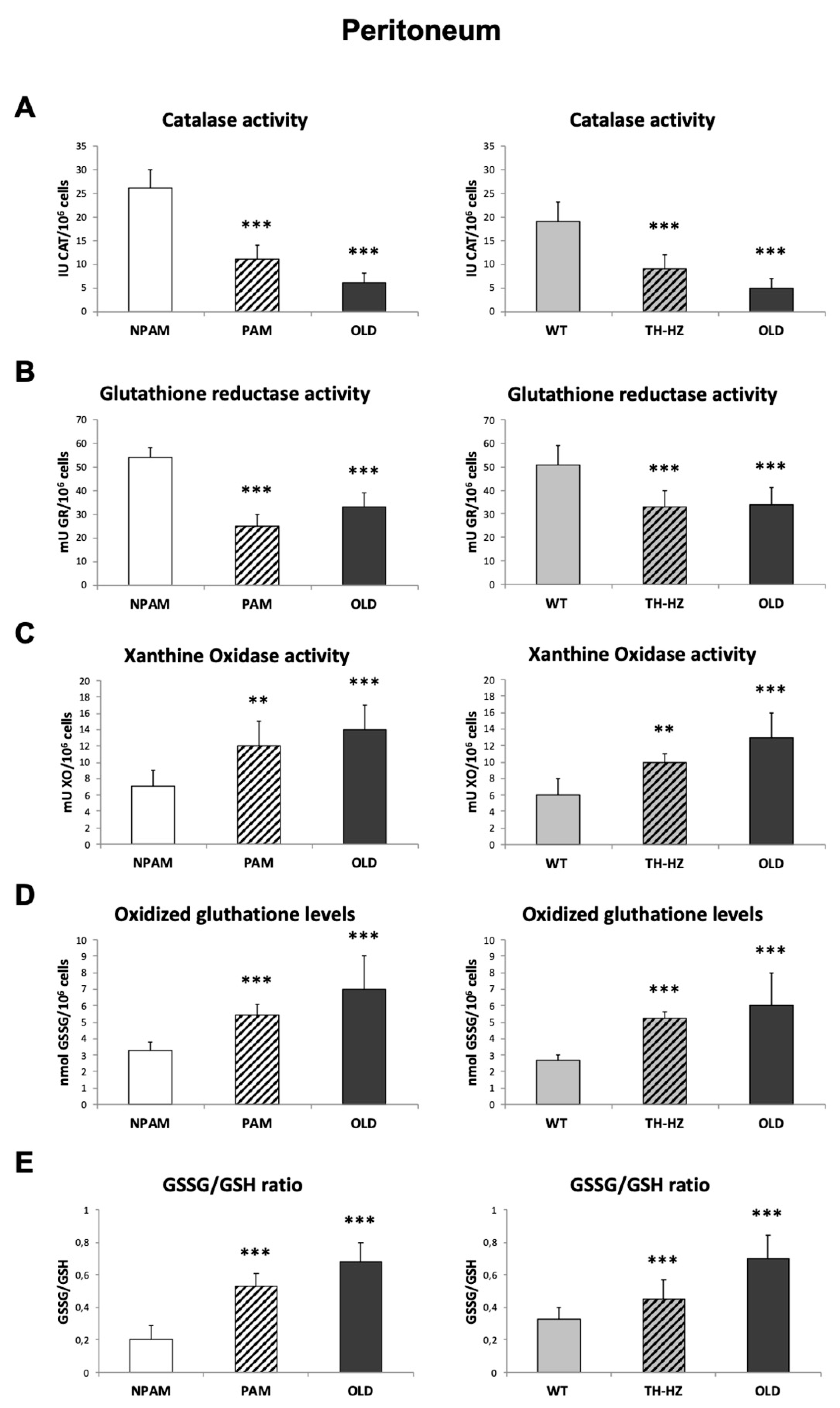
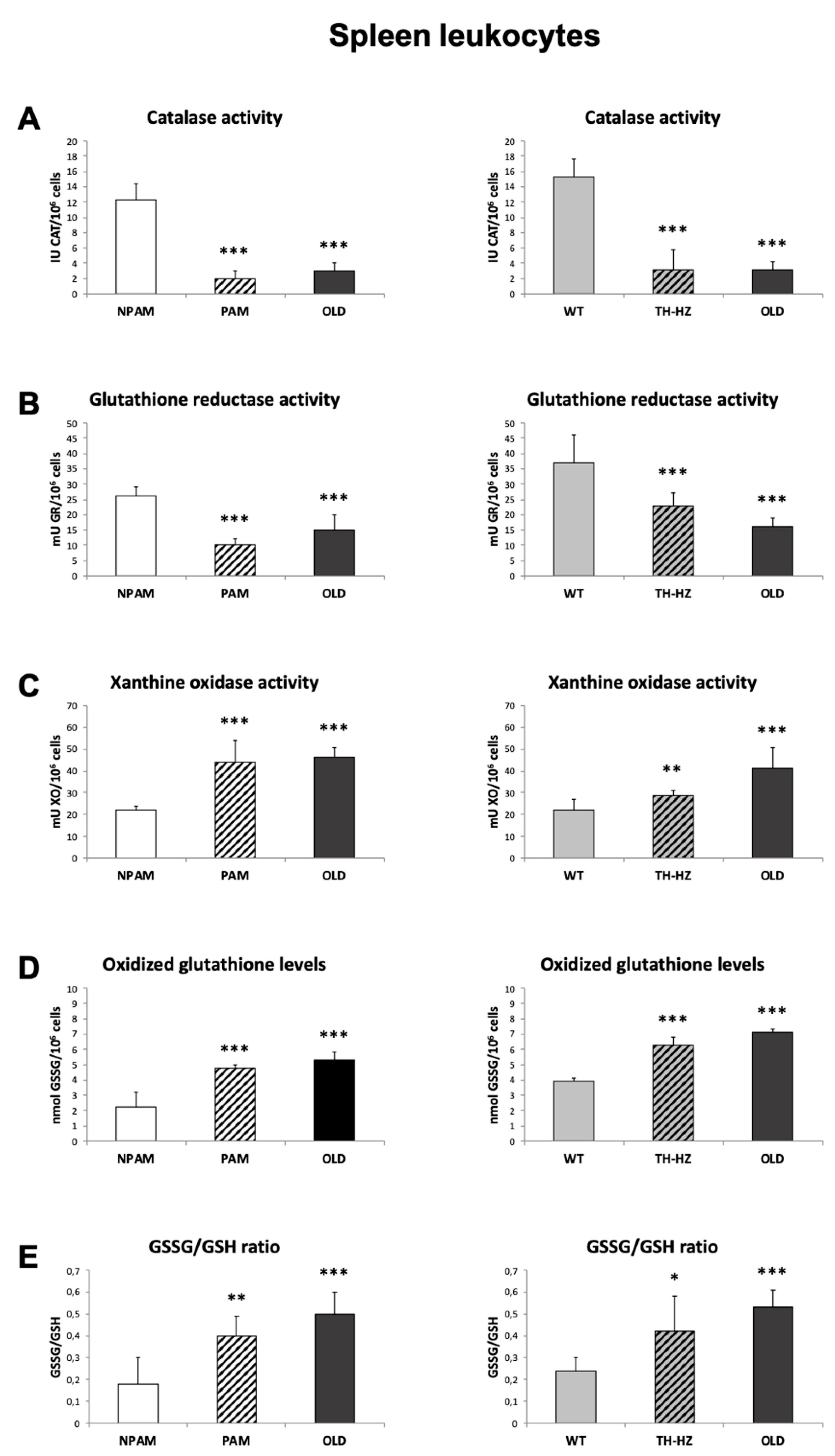
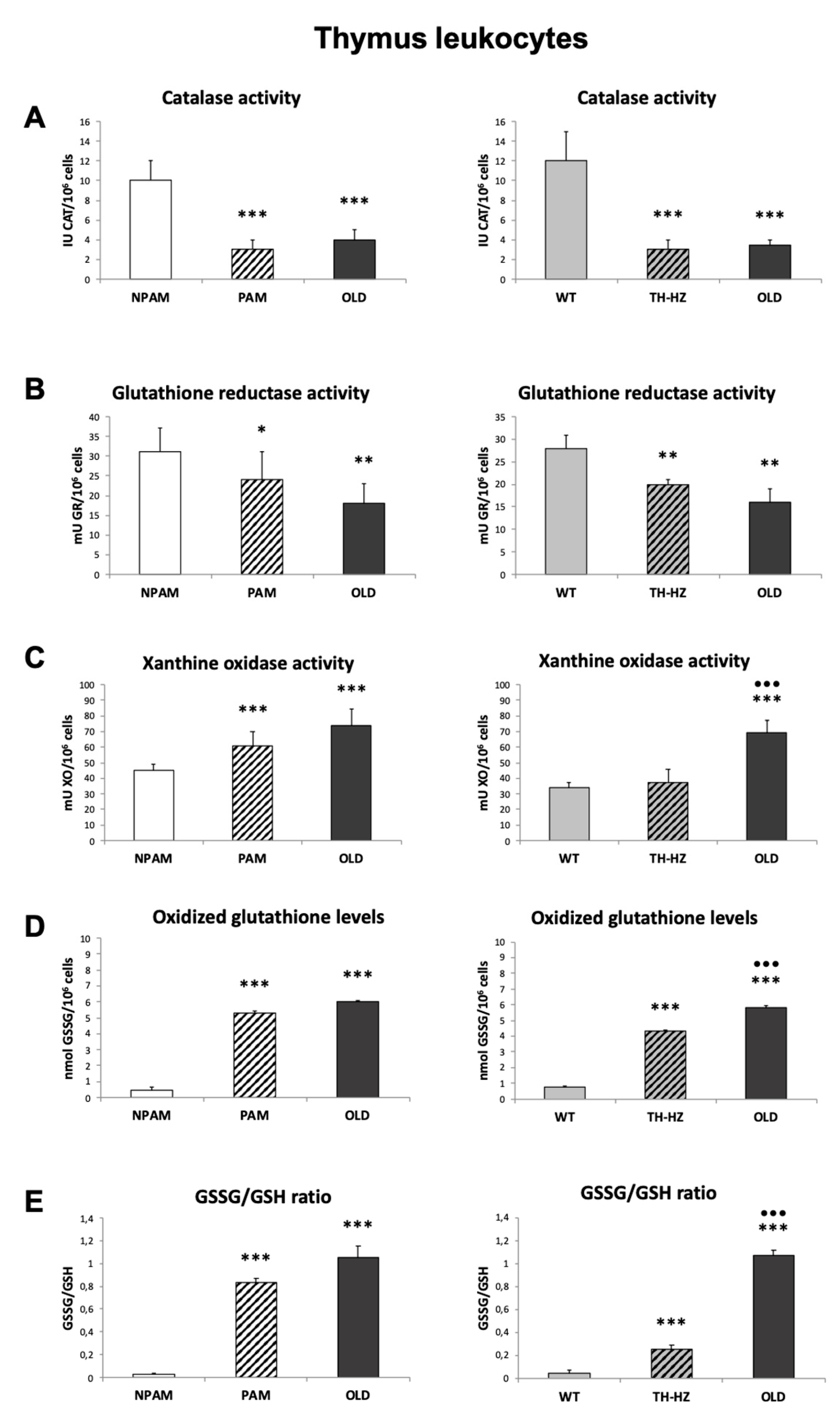
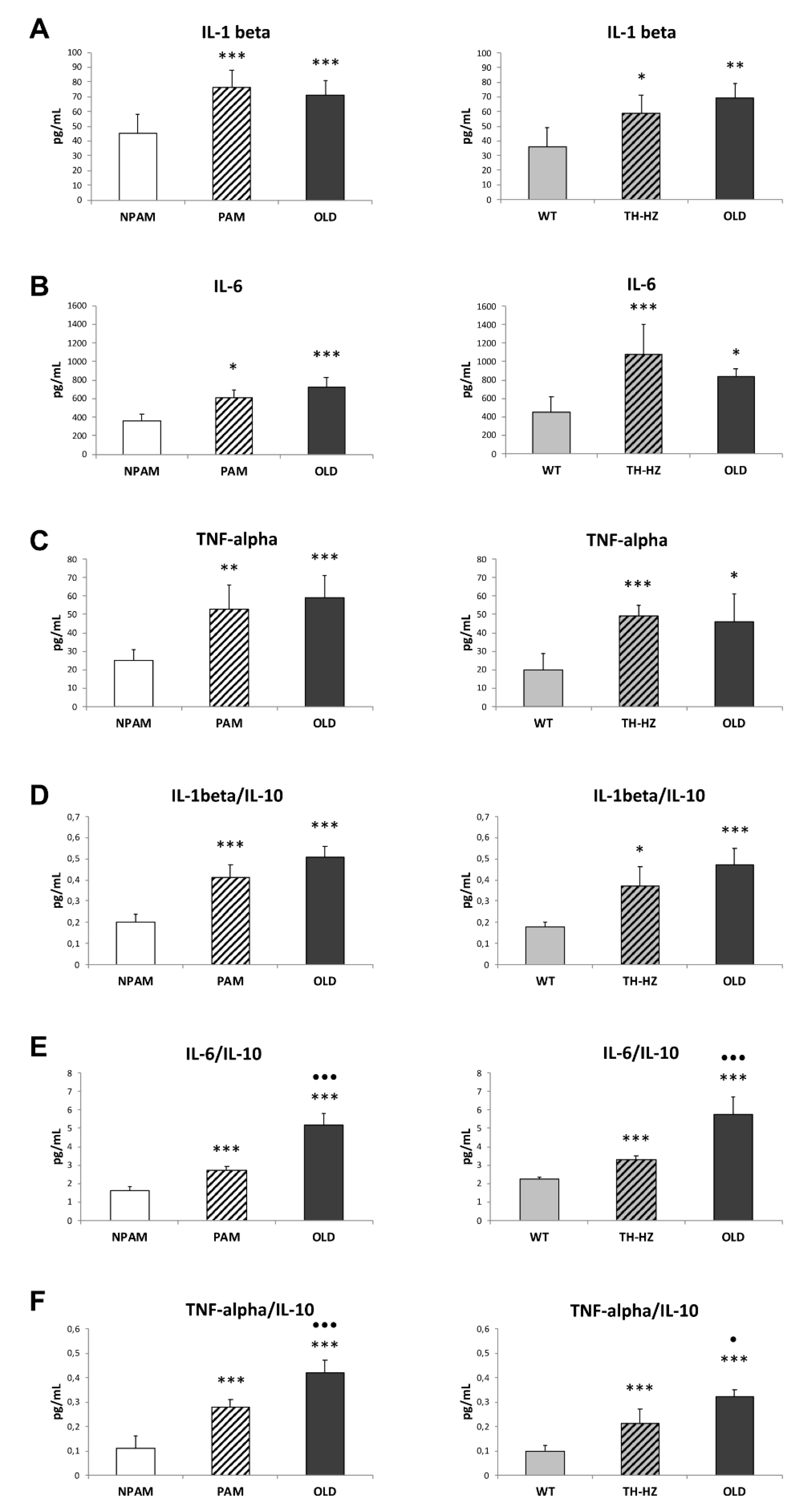
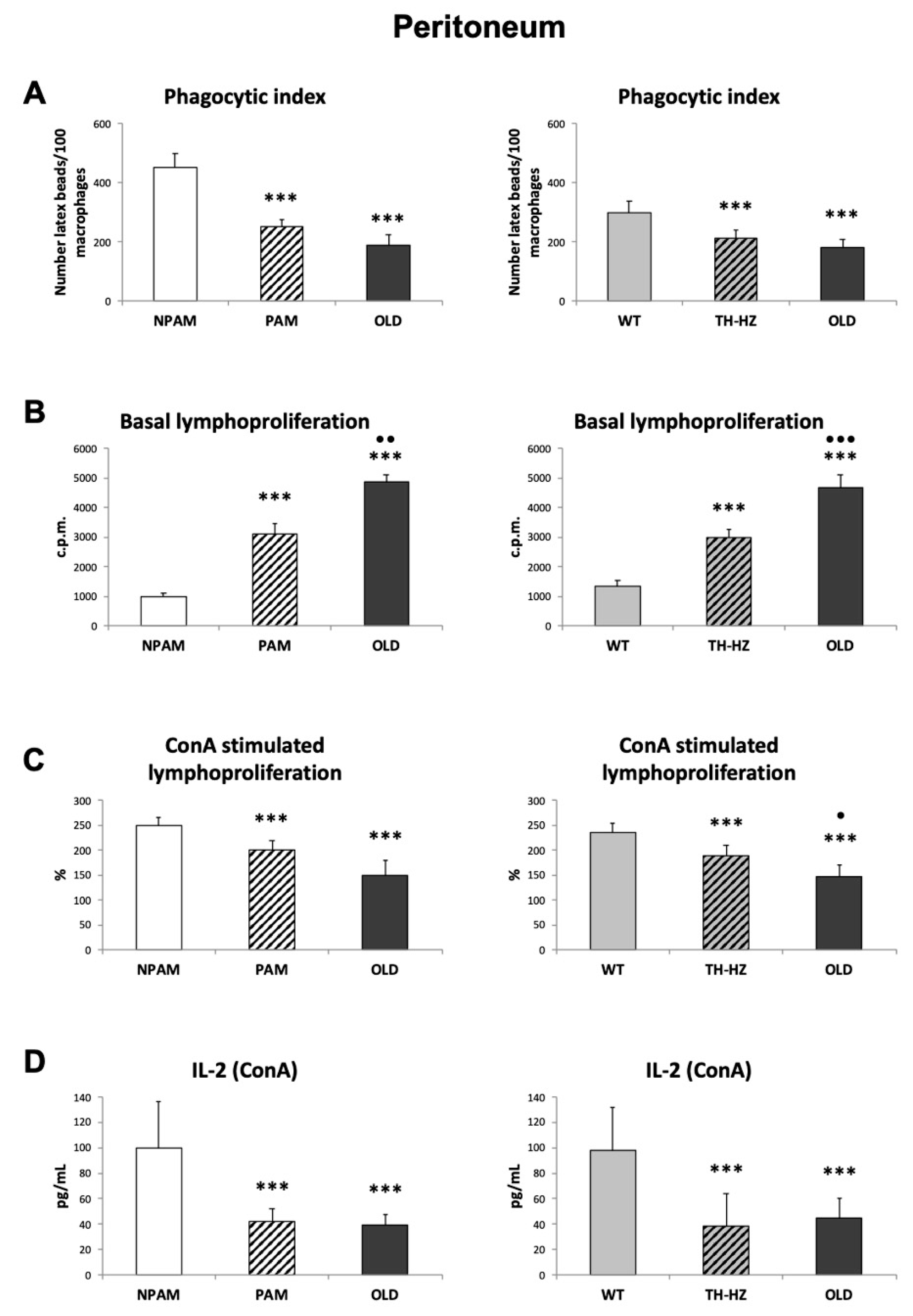
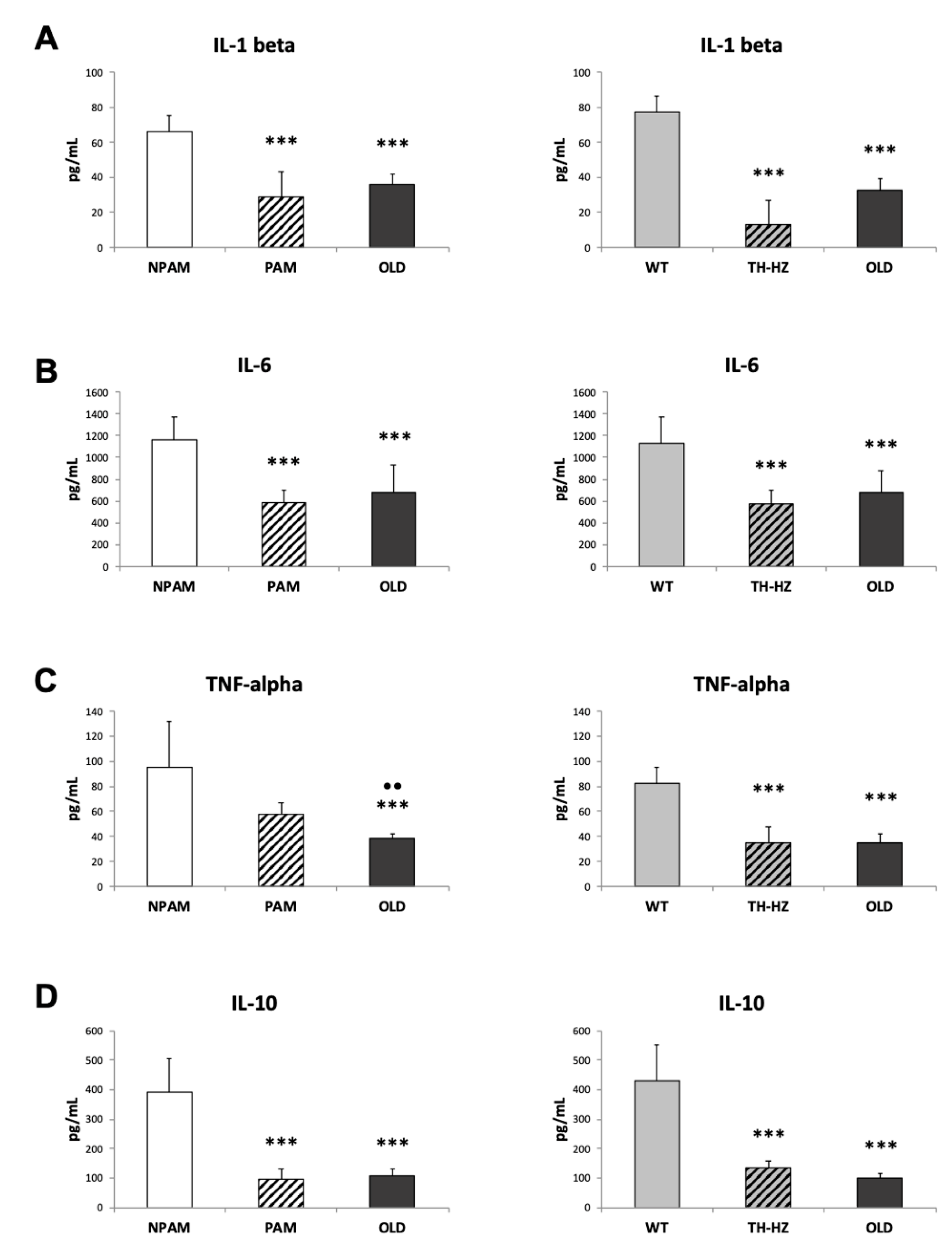
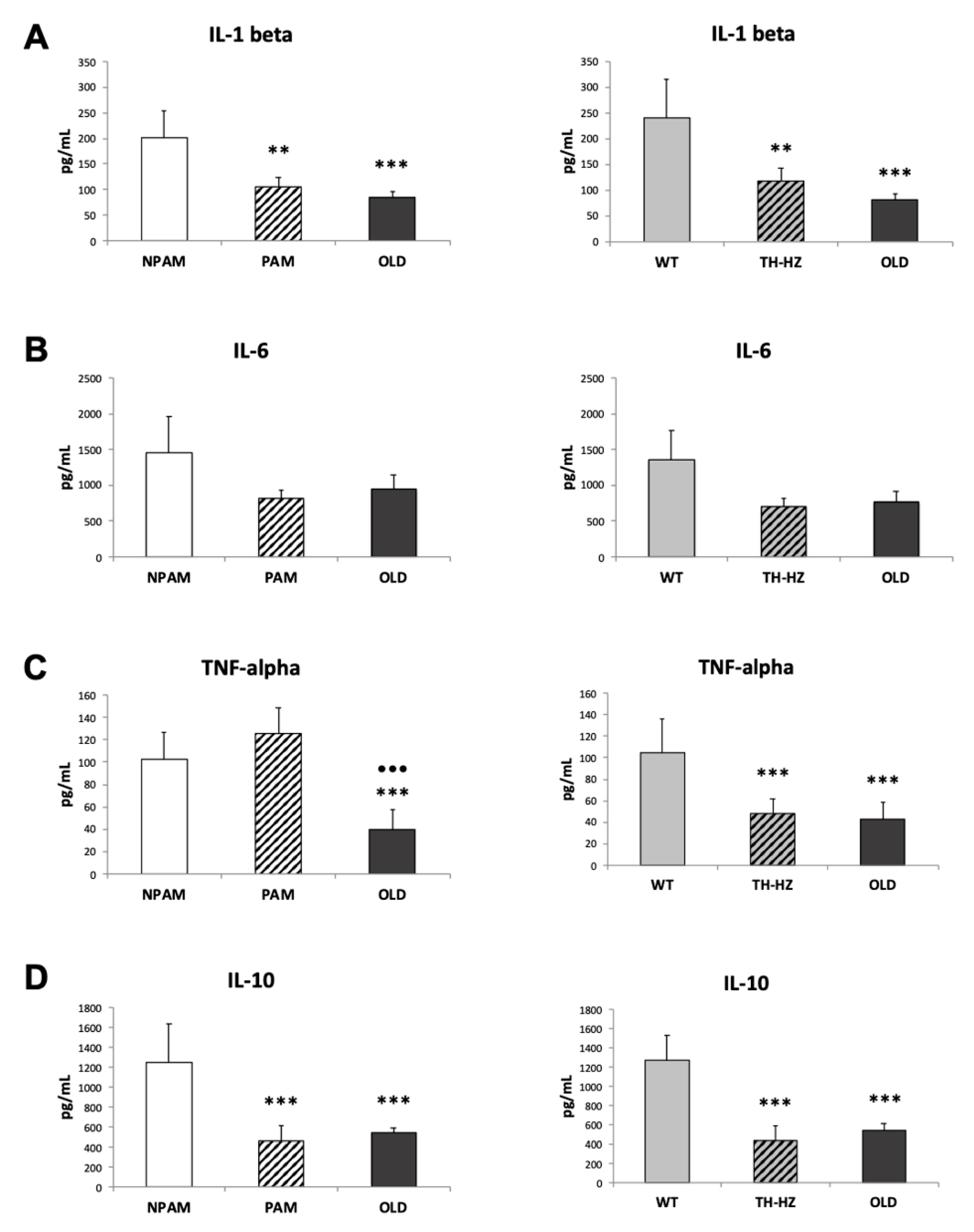
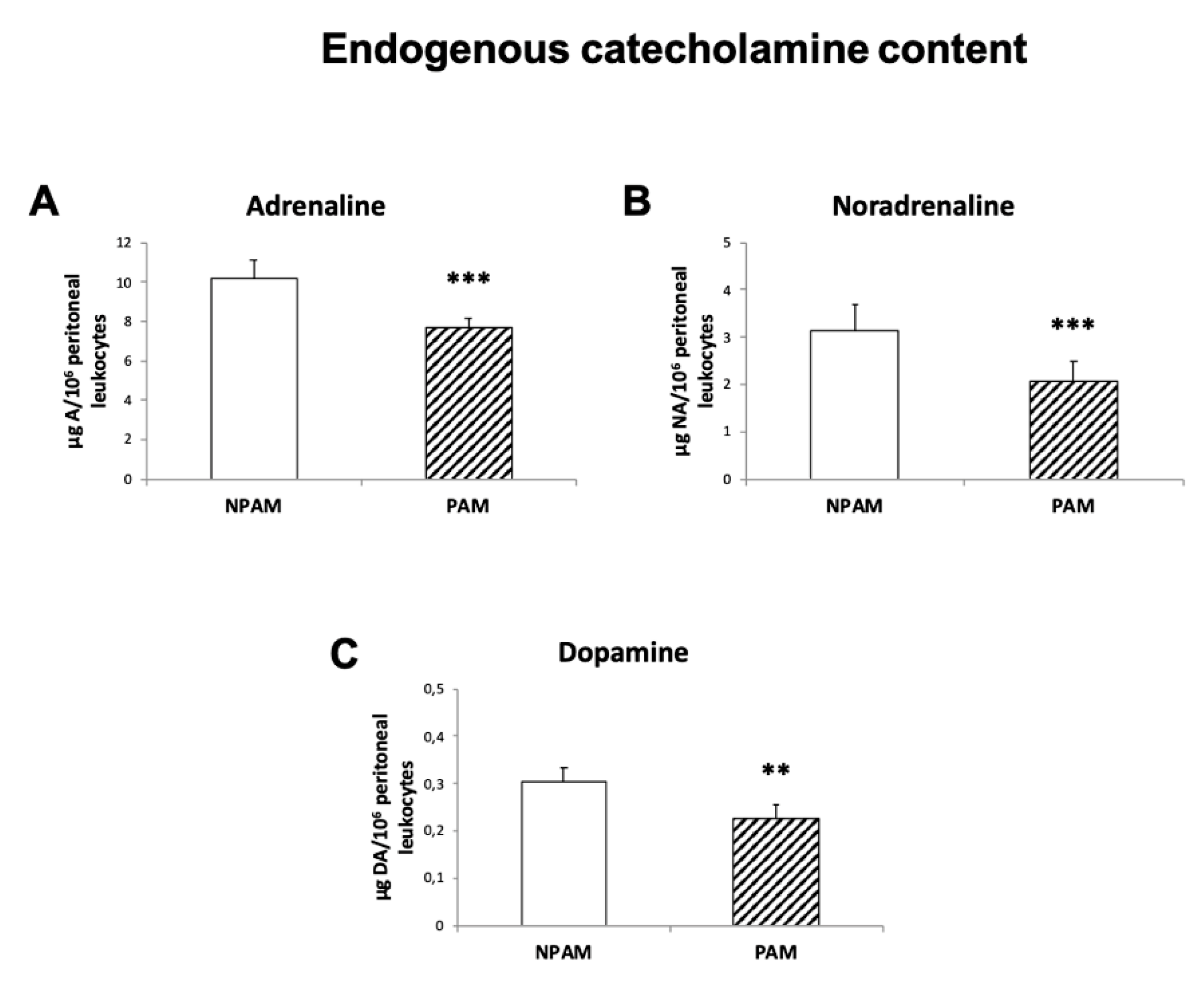
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Collection of Samples
4.2.1. Peritoneal Leukocytes
4.2.2. Spleen and Thymus Leukocytes
4.3. Oxidative Stress Parameters: Antioxidants and Oxidants, in Peritoneal, Spleen and Thymus Leukocytes
4.3.1. Catalase Activity
4.3.2. Glutathione Reductase Activity
4.3.3. Glutathione Content
4.3.4. Xanthine Oxidase Activity
4.4. Inflammatory Stress. Pro-inflammatory and Anti-Inflammatory Cytokines in Peritoneal Leukocyte Cultures
4.5. Functions of Peritoneal Immune Cells
4.5.1. Phagocytic Index
4.5.2. Lymphoproliferation
4.5.3. IL-2 Release Measurement
4.6. Endogenous Catecholamine Content
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PAM | Prematurely aging mice. |
| NPAM | Non-prematurely aging mice |
| TH-HZ | Tyrosine hydroxylase haploinsufficient mice |
| LPS | Lipopolissacharide |
| ConA | Concanavaline A |
| th | Tyrosine hydroxylase gene |
| TH | Tyrosine hydroxylase enzyme |
| A | Adrenaline |
| NA | Noradrenaline |
| DA | Dopamine |
References
- Besedovsky, H.O.; del Rey, A. Physiology of psychoneuroimmunology: A personal view. Brain Behav. Immun. 2007, 21, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G. Age and immunity: What is “immunosenescence”? Exp. Gerontol. 2018, 105, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Dupuis, G.; Witkowski, J.M.; Larbi, A. The role of immunosenescence in the development of age-related diseases. Rev. Investig. Clin. 2016, 68, 84–91. [Google Scholar]
- Fulop, T.; McElhaney, J.; Pawelec, G.; Cohen, A.A.; Morais, J.A.; Dupuis, G.; Baehl, S.; Camous, X.; Witkowski, J.M.; Larbi, A. Frailty, inflammation and immunosenescence. Interdiscip. Top. Gerontol. Geriatr. 2015, 41, 26–40. [Google Scholar] [PubMed]
- Wayne, S.J.; Rhyne, R.L.; Garry, P.J.; Goodwin, J.S. Cell-mediated immunity as a predictor of morbidity and mortality in subjects over 60. J. Gerontol. 1990, 45, 45–48. [Google Scholar] [CrossRef]
- Ferguson, F.G.; Wikby, A.; Maxson, P.; Olsson, J.; Johansson, B. Immune parameters in a longitudinal study of a very old population of Swedish people: A comparison between survivors and non-survivors. J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, B378–B383. [Google Scholar] [CrossRef] [PubMed]
- De Martinis, M.; Franceschi, C.; Monti, D.; Ginaldi, L. Inflammation markers predicting frailty and mortality in the elderly. Exp. Mol. Phathol. 2006, 80, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Dewan, S.K.; Zheng, S.B.; Xia, S.J.; Bill, K. Senescent remodeling of the immune system and its contribution to the predisposition of the elderly to infections. Chin. Med. J. 2012, 125, 3325–3331. [Google Scholar]
- Fulop, T.; Larbi, A.; Kotb, R.; de Angelis, F.; Pawelec, G. Aging, immunity and cancer. Discov. Med. 2011, 11, 537–550. [Google Scholar]
- Martinez de Toda, I.; Maté, I.; Vida, C.; Cruces, J.; De la Fuente, M. Immune function parameters as markers of biological age and predictors of longevity. Aging 2016, 8, 3110–3119. [Google Scholar] [CrossRef]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 2, 298–300. [Google Scholar] [CrossRef]
- De la Fuente, M.; Miquel, J. An update of the oxidation inflammation theory of aging: The involvement of the immune system in oxi-inflamm-aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Updating the mitochondrial free radical theory of aging: An integrated view, key aspects, and confounding concepts. Antioxid. Redox Signal. 2013, 19, 1420–1445. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. The mitochondrial free radical theory of aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar] [CrossRef]
- Go, Y.M.; Jones, D.P. Redox theory of aging: Implications for health and disease. Clin. Sci. 2017, 131, 1669–1688. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A. Mitochondrial reactive oxygen species: Do they extend or shorten animal lifespan? Biochim. Biophys. Acta 2016, 1857, 1116–1126. [Google Scholar] [CrossRef]
- Sanz, A.; Stefanatos, R.K. The mitocondrial free radical theory of aging: A critical view. Curr. Aging Sci. 2008, 1, 10–21. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafé, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and inflamm-aging as two sides of the same coin: Friends or foes? Front. Immunol. 2018, 10, 1960. [Google Scholar] [CrossRef]
- Salvioli, S.; Capri, M.; Valensin, S.; Tieri, P.; Monti, D.; Ottaviani, E.; Franceschi, C. Inflamm-aging, cytokines and aging: State of the art, new hypotheses on the role of mitochondria and new perspectives from systems biology. Curr. Pharm. Des. 2006, 12, 3161–3171. [Google Scholar] [CrossRef]
- Vida, C.; Gonzalez, E.M.; De la Fuente, M. Increase of oxidation and inflammation in nervous and immune systems with aging and anxiety. Curr. Pharm. Des. 2014, 20, 4656–4678. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Viadero, C.; Jiménez-Sanz, M.; Fernández-Pérez, A.; Verduga Vélez, R.; Crespo Santiago, D. Inflammation and oxidation: Predictive and/or causative factors. Rev. Esp. Geriatr. Gerontol. 2016, 1, 27–33. [Google Scholar] [CrossRef]
- Bauer, M.E.; De la Fuente, M. The role of oxidative and inflammatory stress and persistent viral infections in immunosenescence. Mech. Ageing Dev. 2016, 156, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E. Stress, glucocorticoids and ageing of the immune system. Stress 2005, 8, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Racchi, M.; Bouso, E.; Ronfani, M.; Serafini, M.M.; Galasso, M.; Lanni, C.; Corsini, E. Role of hormones in the regulation of RACK1 Expression as a signaling checkpoint in immunosenescence. Int. J. Mol. Sci. 2017, 18, 1453. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.E.; Muller, G.C.; Correa, B.L.; Vianna, P.; Turner, J.E.; Bosch, J.A. Psychoneuroendocrine interventions aimed at attenuating immunosenescence: A review. Biogerontology 2013, 14, 8–20. [Google Scholar] [CrossRef]
- Turner, J.E. Is immunosenescence influenced by our lifetime “dose” of exercise? Biogerontology 2016, 17, 581–602. [Google Scholar] [CrossRef]
- Bauer, M.E. Chronic stress and immunosenescence: A review. Neuroimmunomodulation 2008, 15, 241–250. [Google Scholar] [CrossRef]
- Cruces, J.; Venero, C.; Pereda-Pérez, J.; De la Fuente, M. The effect of psychological stress and social isolation on neuroendocrine communication. Curr. Pharm. Des. 2014, 20, 4608–4628. [Google Scholar] [CrossRef]
- De la Fuente, M. Oxidation and Inflammation in the immune and nervous systems, a link between aging and anxiety. In Handbook of Immunosenescence; Fulop, T., Franceschi, C., Hirokawa, K., Pawelec, G., Eds.; Springer: Berlin, Germany, 2018; 28p. [Google Scholar]
- Borkan, G.A.; Norris, A.H. Assessment of biological age using a profile of physical parameters. J. Gerontol. 1980, 35, 177–184. [Google Scholar] [CrossRef]
- Bulpitt, C.J.; Antikainen, R.L.; Markowe, H.L. Mortality according to a prior assessment of biological age. Curr. Aging Sci. 2009, 2, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Guayerbas, N.; De la Fuente, M. An impairment of phagocytic function is linked to a shorter life span in two strains of prematurely-aging mice. Dev. Comp. Immunol. 2003, 27, 339–350. [Google Scholar] [CrossRef]
- Viveros, M.P.; Arranz, L.; Hernanz, A.; Miquel, J.; De la Fuente, M. A model of premature aging in mice based on altered stress-related behavioral response and immunosenescence. Neuroimmunomodulation 2007, 14, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Garrido, A.; Cruces, J.; Ceprián, N.; Hernández-Sánchez, C.; De la Fuente, M. Premature aging in behavior and immune functions in tyrosine hydroxylase haploinsufficient female mice. A longitudinal study. Brain Behav. Immun. 2018, 69, 440–455. [Google Scholar] [CrossRef]
- Osada, K.; Minehira, K.; Inoue, S.; Nakamura, S.; Yamada, K.; Sugano, M. Effect of oxidized colesterol on age-associated changes to immune parameters in spleen lymphocytes and peritoneal exudate cells derived from rats. Biosci. Biotechnol. Biochem. 2000, 64, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Caamano, J.H.; Lord, J.M.; De la Fuente, M. Preserved immune functions and controlled leukocyte oxidative stress in naturally long-lived mice: Possible role of nuclear factor kappa β. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Arranz, L.; Lord, J.M.; De la Fuente, M. Preserved ex vivo inflammatory status and cytokine responses in naturally long-lived mice. Age 2010, 32, 451–466. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 1, S4–S9. [Google Scholar] [CrossRef]
- Frasca, D.; Bloberg, B.B. Inflammaging decreases adaptive and innate immune responses in mice and humans. Biogerontology 2016, 17, 7–19. [Google Scholar] [CrossRef]
- Cannizzo, E.S.; Clement, C.C.; Sahu, R.; Follo, C.; Santambrogio, L. Oxidative stress, inflamm-aging and immunosenescence. J. Proteom. 2011, 74, 2313–2323. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An update on inflamm-aging: Mechanisms, prevention and treatment. J. Immunol. Res. 2016, 8426874. [Google Scholar] [CrossRef] [PubMed]
- Guayerbas, N.; Puerto, M.; Victor, V.M.; Miquel, J.; De la Fuente, M. Leukocyte function and life span in a murine model of premature immunosenescence. Exp. Gerontol. 2002, 37, 249–256. [Google Scholar] [CrossRef]
- Guayerbas, N.; Catalán, M.; Victor, V.M.; Miquel, J.; De la Fuente, M. Relation of behaviour and macrophage function to life span in a murine model of premature immunosenescence. Brain Behav. Res. 2002, 134, 41–48. [Google Scholar] [CrossRef]
- Garrido, A.; Cruces, J.; Iriarte, I.; Hernández-Sánchez, C.; de Pablo, F.; De la Fuente, M. Premature immunosenescence in catecholamine synthesis defficient mice. Effect of social environment. Rev. Esp. Geriatr. Gerontol. 2017, 52, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.K.; Smith, C.A.; Sakamoto, K.; Kaminski, N.; Koff, J.L.; Goldstein, D.R. Aging impairs alveolar macrophage phagocytosis and increases influenza-induced mortality in mice. J. Immunol. 2017, 199, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, S.; Carlisi, M.; Santoro, M.; Napolitano, M.; Raso, S.; Siragusa, S. Immunosenescence and lymphomagenesis. Immun. Ageing 2018, 15, 22. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, J.; Tarkowski, A.; Ekman, R.; Erwing, A. Discovery of endogenous catecholamines in lymphocytes and evidence for catecholamine regulation of lymphocyte function via autocrine loop. Proc. Natl. Acad. Sci. USA 1994, 91, 12912–12916. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, M.; Marino, F.; Bombelli, R.; Ferrari, M.; Lecchini, S.; Frigo, G. Endogenous catecholamine synthesis, metabolism, storage and uptake in human neutrophils. Life Sci. 1999, 64, 975–981. [Google Scholar] [CrossRef]
- Qiu, Y.H.; Cheng, C.; Dai, L.; Peng, Y.P. Effect of endogenous catecholamines in lymphocytes on lymphocyte function. J. Neuroimmunol. 2005, 167, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Madden, K.S. Sympathetic neural-immune interactions regulate hematopoiesis, thermoregulation and inflammation in mammals. Dev. Comp. Immun. 2017, 66, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; Martinez de Toda, I.; Cruces, J.; Garrido, A.; González-Sánchez, M.; De la Fuente, M. Role of macrophages in age-related oxidative stress and lipofuscin accumulation in mice. Redox Biol. 2017, 12, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, C.; Álvarez, P.; Jiménez, L.; De la Fuente, M. Oxidative stress in leukocytes from young prematurely aging mice is reversed by supplementation with biscuits rich in antioxidants. Dev. Comp. Immunol. 2006, 30, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.; Kequan, G.; Yasushi, A.; Susumu, I. Immune dysfunction associated with abnormal bone marrow-derived mesenchymal stroma cells in senescence accelerated mice. Int. J. Mol. Sci. 2016, 17, 183. [Google Scholar] [CrossRef]
- Caballero, B.; Vega-Naredo, I.; Sierra, V.; DeGonzalo-Calvo, D.; Medrano-Campillo, P.; Guerrero, J.M.; Tolivia, D.; Rodríguez-Colunga, M.J.; Coto-Montes, A. Autophagy upregulation and loss of NF-KappaB in oxidative stress-related immunodeficient SAMP8 mice. Mech. Ageing Dev. 2009, 130, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, T.; Asada, S.; Nishitani, S.; Hazeki, O. Age-related changes in manganese superoxide dismutase activity in the cerebral cortex of senescence-accelerated prone and resistant mouse. Neurosci. Lett. 2001, 298, 135–138. [Google Scholar] [CrossRef]
- Farr, S.A.; Poon, H.F.; Dogrokol-Ak, D.; Drake, J.; Banks, W.A.; Everman, E.; Butterfield, D.A.; Morley, J.E. The antioxidants a-lipoic and N-acetylcyteine reverse memory impairment and brain oxidative stress in aged SAMP8 mice. J. Neurochem. 2003, 84, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Sureda, F.X.; Gutierrez-Cuesta, J.; Romeu, M.; Mulero, M.; Canudas, A.M.; Camins, A.; Mallol, J.; Pallás, M. Changes in oxidative stress parameters and neurodegeneration markers in the brain of the senescence-accelerated mice SAMP-8. Exp. Gerontol. 2006, 41, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Baeza, I.; De Castro, N.M.; Arranz, L.; Fdez-Tresguerres, J.; De la Fuente, M. Ovariectomy causes immunosenescence and oxi-inflamm-ageing in peritoneal leukocytes of age female mice similar to that in aged males. Biogerontology 2011, 12, 227–238. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Vida, C.; De la Fuente, M. An appropriate modulation of lymphoproliferative response and cytokine release as possible contributors to longevity. Int. J. Mol. Sci. 2017, 18, 1598. [Google Scholar] [CrossRef]
- Vassileva, V.; Piquette-Miller, M. Inflammation: The dynamic force of health and disease. Clin. Pharmacol. Ther. 2014, 96, 401–405. [Google Scholar] [CrossRef]
- Mooradian, A.D.; Reed, R.L.; Osterweil, D.; Scuderi, P. Detectable serum levels of tumor necrosis factor alpha may predict early mortality in elderly institutionalized patients. J. Am. Geriatr. Soc. 1991, 39, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.B.; Ferrucci, L.; Tracy, R.P.; Corti, M.C.; Wacholder, S.; Ettinger, W.H., Jr.; Heimovitz, H.; Cohen, H.J.; Wallace, R. Associations of elevated interleukin-6 and C-reactive protein levels with mortality in the elderly. Am. J. Med. 1999, 106, 506–512. [Google Scholar] [CrossRef]
- Ferrucci, L.; Harris, T.B.; Guralnik, J.M.; Tracy, R.P.; Corti, M.C.; Cohen, H.J.; Penninx, B.; Pahor, M.; Wallace, R.; Havlik, R.J. Serum IL-6 level and the development of disability in older persons. J. Am. Geriatr. Soc. 1999, 47, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Behnia, F.; Sheller, S.; Menon, R. Mechanistic Differences Leading to Infectious and Sterile Inflammation. Am. J. Reprod. Immunol. 2016, 75, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.I.; Escames, G.; López, L.C.; López, A.; García, J.A.; Ortiz, F.; Acuña-Castroviejo, D. Chronic melatonin treatment reduces the age-dependent inflammatory process in senescence-accelerated mice. J. Pineal Res. 2007, 42, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shao, W.F.; Yuan, L.F.; Tu, P.F.; Ma, Z.Z. Decreasing pro-inflamatory cytokine and reversing the immunosenescence with extracts of Pu-erh tea in senescence accelerated mouse (SAM). Food Chem. 2012, 135, 2222–2228. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, O.; Pawelec, G.; Peralbo, E.; Wikby, A.; Mariani, E.; Mocchegiani, E.; Tarazona, R.; Solana, R. Immunological biomarkers of ageing in man: Changes in both innate and adaptive immunity are associated with health and longevity. Biogerontology 2006, 7, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Lectin receptors as lymphocyte surface markers. Adv. Immunol. 1983, 83, 213–298. [Google Scholar]
- Hallgren, H.M.; Bergh, N.; Rodysill, J.K.; O’Leary, J.J. Lymphocyte proliferative response to PHA and anti-CD3/Ti monoclonal antibodies T cell surface marker expression, and serum IL-2 receptor levels as biomarkers of age and health. Mech. Aging Dev. 1998, 43, 175–185. [Google Scholar] [CrossRef]
- Wikby, A.; Ferguson, F.; Forsey, R.; Thompson, J.; Strindhall, J.; Lofgren, S.; Nilsson, B.O.; Ernerudh, J.; Pawelec, G.; Johansson, B. An immune risk phenotype, cognitive impairment, and survival in very late life: Impact of allostatic load in wedish octogenarian and nonagenarian humans. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 556–565. [Google Scholar] [CrossRef]
- Pera, A.; Campos, C.; López, N.; Hassouneh, F.; Alonso, C.; Tarazona, R.; Solana, R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas 2015, 82, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Kobayasi, K.; Morita, S.; Sawada, H.; Mizuguchi, T.; Yamada, K.; Nagatsu, I.; Hata, T.; Watenabe, Y.; Fujita, K.; Nagatsu, T. Targeted disruption of the tyrosine hydroxylase locus results in severe catecholamine depletion and perinatal lethality in mice. J. Biol. Chem. 1995, 270, 27235–27243. [Google Scholar] [CrossRef]
- Webster Marketon, J.I.; Glaser, R. Stress hormones and immune function. Cell. Immunol. 2008, 252, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; De la Fuente, M. Stress-related behavioural responses, immunity and ageing in animal models. Immunosenescence. In Immunosenescence: Psychosocial and Behavioral Determinant; Bosch, J.A., Phillips, C., Lord, J.M., Eds.; Springer: New York, NY, USA, 2013; pp. 125–144. [Google Scholar]
- Aguilera, G. HPA axis responsiveness to stress: Implications for healthy aging. Exp. Gerontol. 2011, 46, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Kvetnansky, R.; Sabban, E.L.; Palkovits, M. Catecholaminergic systems in stress: Structural and molecular genetic approaches. Physiol. Rev. 2009, 89, 535–606. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, P.; Alvarado, C.; Puerto, M.; Schlumberger, A.; Jiménez, L.; De la Fuente, M. Improvement of leukocyte functions in prematurely aging mice after five weeks of diet supplementation with polyphenol-rich cereals. Nutrition 2006, 22, 913–921. [Google Scholar] [CrossRef]
- Alvarado, C.; Álvarez, P.; Puerto, M.; Gausserés, N.; Jiménez, L.; De la Fuente, M. Dietary supplementation with antioxidants improves functions and decreases oxidative stress of leukocytes from prematurely aging mice. Nutrition 2006, 22, 767–777. [Google Scholar] [CrossRef]
- Guayerbas, N.; Puerto, M.; Álvarez, P.; De la Fuente, M. Improvement of the macrophage functions in prematurely aging mice by a diet supplemented with thiolic antioxidants. Cell. Mol. Biol. 2004, 50, OL677-81. [Google Scholar]
- Zhou, Q.Y.; Quaife, C.J.; Palmiter, R.D. Targeted disruption of the tyrosine hydroxylase gen reveals that catecholamines are required for mouse fetal development. Nature 1995, 374, 640–643. [Google Scholar] [CrossRef]
- Vázquez, P.; Robles, A.M.; de Pablo, F.; Hernández-Sánchez, C. Non-neural tyrosine hydroxylase, via modulation of endocrine pancreatic precursors, is required for normal development of beta-cells in the mouse pancreas. Diabetologia 2014, 57, 2339–2347. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [PubMed]
- Massey, V.; Williams, C. On the reaction mechanism of yeast glutathione reductase. J. Biol. Chem. 1965, 240, 4470–4481. [Google Scholar] [PubMed]
- Hissin, P.J.; Hilf, R. A fluorimetric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, A.; Cruces, J.; Ceprián, N.; Vara, E.; de la Fuente, M. Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging. Int. J. Mol. Sci. 2019, 20, 769. https://doi.org/10.3390/ijms20030769
Garrido A, Cruces J, Ceprián N, Vara E, de la Fuente M. Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging. International Journal of Molecular Sciences. 2019; 20(3):769. https://doi.org/10.3390/ijms20030769
Chicago/Turabian StyleGarrido, Antonio, Julia Cruces, Noemí Ceprián, Elena Vara, and Mónica de la Fuente. 2019. "Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging" International Journal of Molecular Sciences 20, no. 3: 769. https://doi.org/10.3390/ijms20030769
APA StyleGarrido, A., Cruces, J., Ceprián, N., Vara, E., & de la Fuente, M. (2019). Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging. International Journal of Molecular Sciences, 20(3), 769. https://doi.org/10.3390/ijms20030769