The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Kinases Involved in the Establishment of Chromosome Bipolar Attachment
2.1. Aurora B/Ipl1 Kinase
2.2. Cyclin-Dependent Kinase (CDK)
2.3. Mps1 Kinase
2.4. Cdc5 Kinase
3. Protein Phosphatases Involved in Chromosome Bipolar Attachment
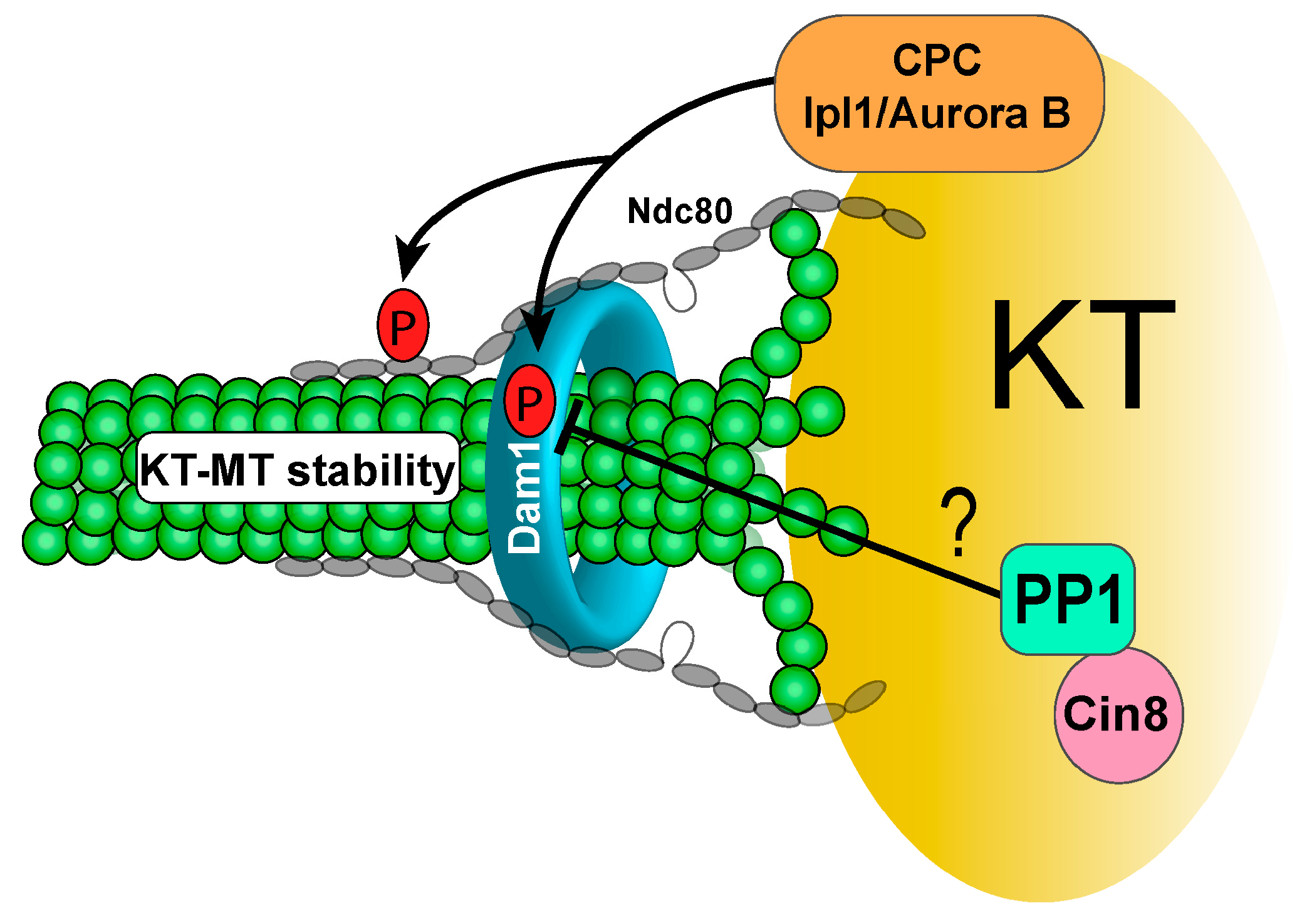
3.1. Protein Phosphatase 1 (PP1) and KT-MT Attachment
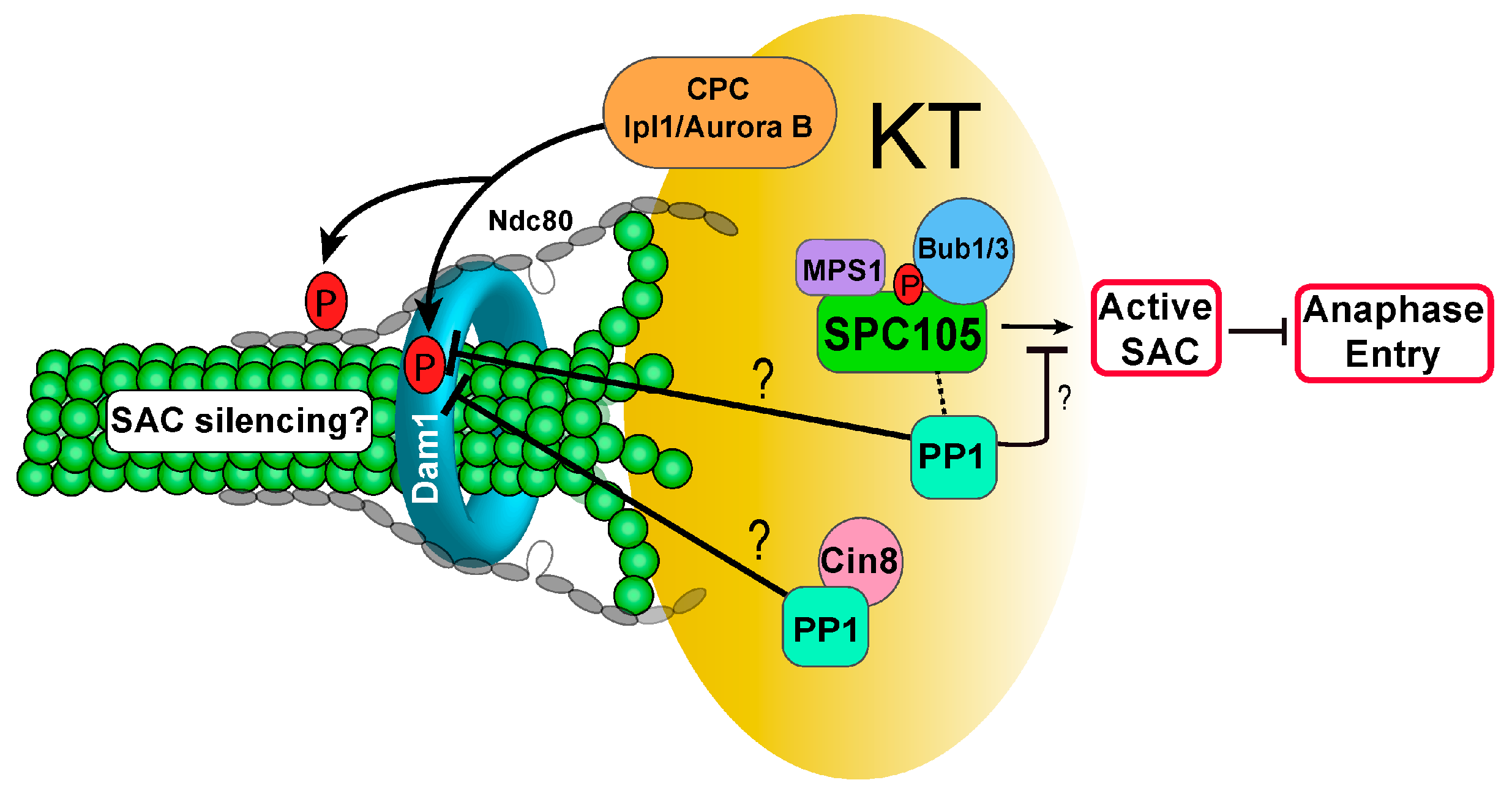
3.2. PP1 and Checkpoint Regulation
3.3. Kinetochore Recruiters for PP1
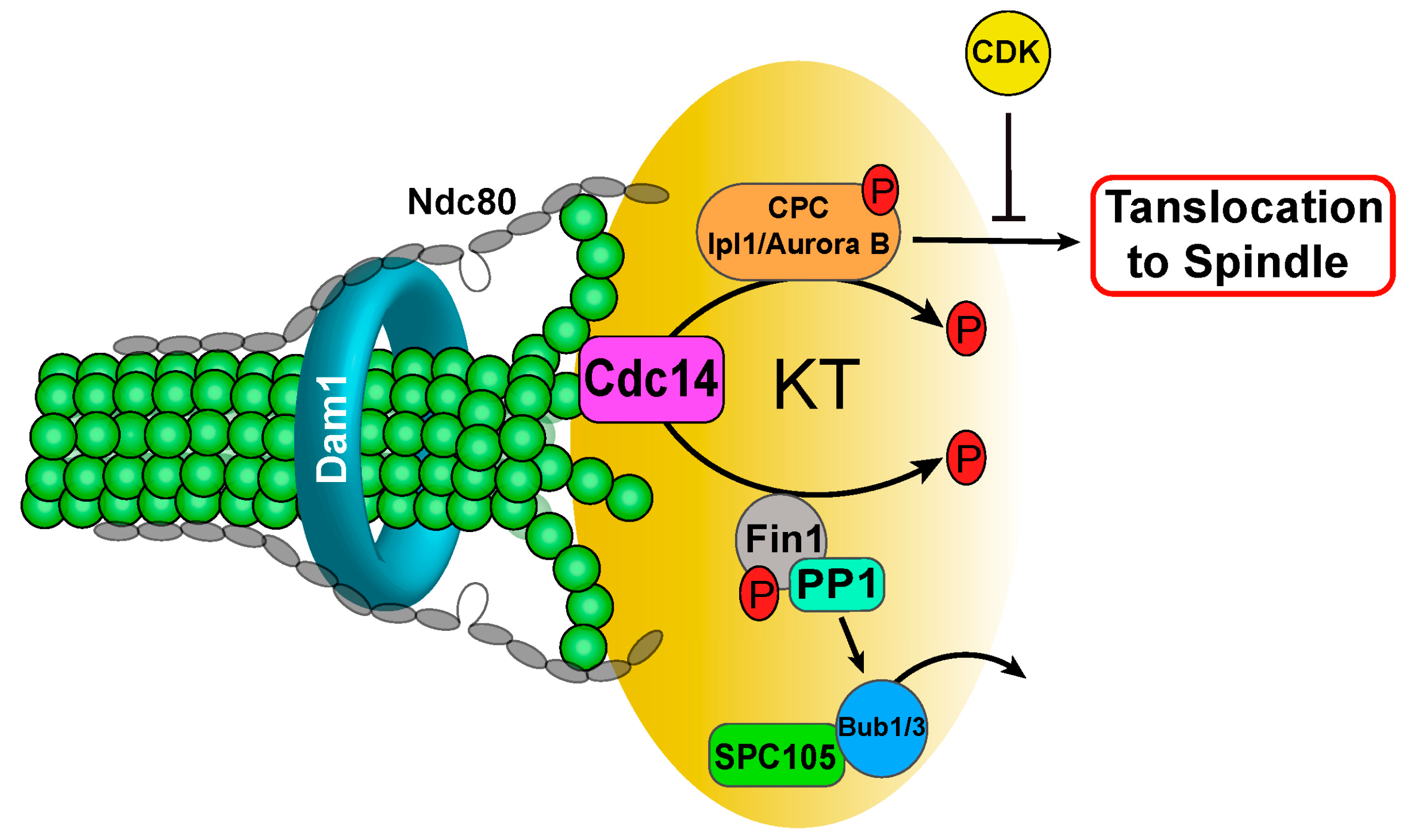
3.4. Phosphatase Cdc14
3.5. Protein Phosphatase 2A (PP2A)
4. The Function of Phosphatases in Meiosis
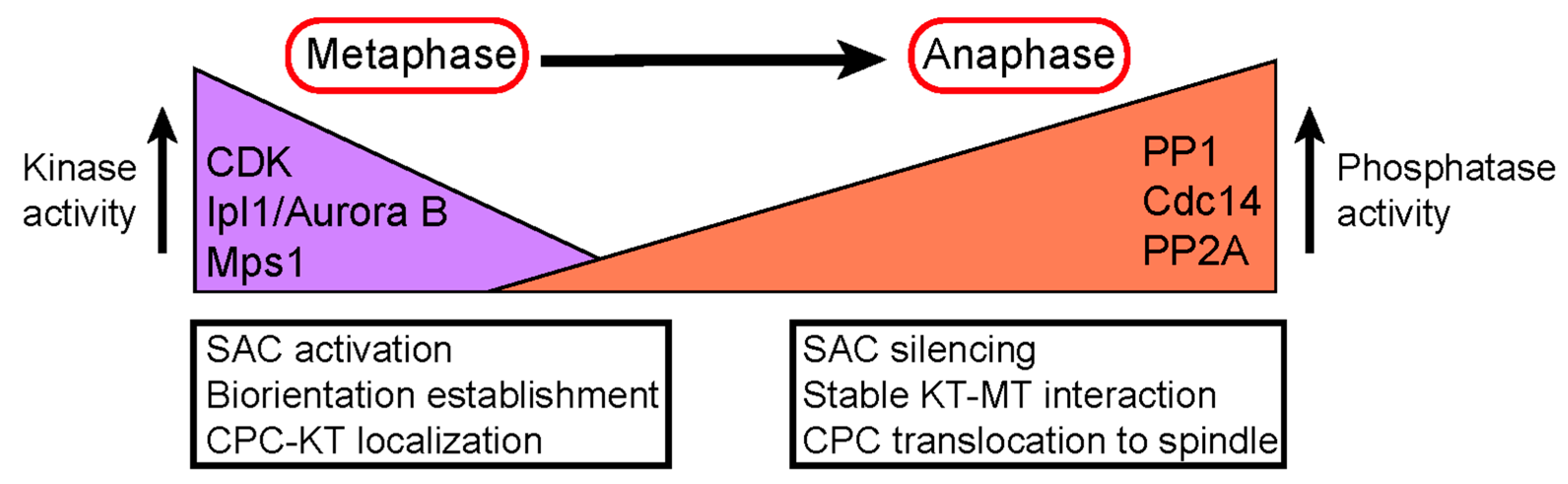
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Biggins, S. The composition, functions, and regulation of the budding yeast kinetochore. Genetics 2013, 194, 817–846. [Google Scholar] [CrossRef] [PubMed]
- Winey, M.; Mamay, C.L.; O’Toole, E.T.; Mastronarde, D.N.; Giddings, T.H., Jr.; McDonald, K.L.; McIntosh, J.R. Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle. J. Cell Biol. 1995, 129, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Lara-Gonzalez, P.; Westhorpe, F.G.; Taylor, S.S. The spindle assembly checkpoint. Curr. Biol. 2012, 22, R966–R980. [Google Scholar] [CrossRef] [PubMed]
- Lampson, M.A.; Cheeseman, I.M. Sensing centromere tension: Aurora B and the regulation of kinetochore function. Trends Cell Biol. 2011, 21, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, F.; Higgins, R.; McKnight, K. The current view for the silencing of the spindle assembly checkpoint. Cell Cycle 2014, 13, 1694–1701. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vader, G.; Medema, R.H.; Lens, S.M. The chromosomal passenger complex: Guiding Aurora-B through mitosis. J. Cell Biol. 2006, 173, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Tyers, R.G.; Wong, C.C.; Yates, J.R., 3rd; Drubin, D.G.; Barnes, G. Nbl1p: A Borealin/Dasra/CSC-1-like protein essential for Aurora/Ipl1 complex function and integrity in Saccharomyces cerevisiae. Mol. Biol. Cell 2009, 20, 1772–1784. [Google Scholar] [CrossRef]
- Buvelot, S.; Tatsutani, S.Y.; Vermaak, D.; Biggins, S. The budding yeast Ipl1/Aurora protein kinase regulates mitotic spindle disassembly. J. Cell Biol. 2003, 160, 329–339. [Google Scholar] [CrossRef]
- Cormier, A.; Drubin, D.G.; Barnes, G. Phosphorylation regulates kinase and microtubule binding activities of the budding yeast chromosomal passenger complex in vitro. J. Biol. Chem. 2013, 288, 23203–23211. [Google Scholar] [CrossRef]
- Tanaka, T.U.; Rachidi, N.; Janke, C.; Pereira, G.; Galova, M.; Schiebel, E.; Stark, M.J.; Nasmyth, K. Evidence that the Ipl1-Sli15 (Aurora kinase-INCENP) complex promotes chromosome bi-orientation by altering kinetochore-spindle pole connections. Cell 2002, 108, 317–329. [Google Scholar] [CrossRef]
- Li, Y.; Bachant, J.; Alcasabas, A.A.; Wang, Y.; Qin, J.; Elledge, S.J. The mitotic spindle is required for loading of the DASH complex onto the kinetochore. Genes Dev. 2002, 16, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, I.M.; Anderson, S.; Jwa, M.; Green, E.M.; Kang, J.; Yates, J.R., 3rd; Chan, C.S.; Drubin, D.G.; Barnes, G. Phospho-regulation of kinetochore-microtubule attachments by the Aurora kinase Ipl1p. Cell 2002, 111, 163–172. [Google Scholar] [CrossRef]
- Westermann, S.; Avila-Sakar, A.; Wang, H.W.; Niederstrasser, H.; Wong, J.; Drubin, D.G.; Nogales, E.; Barnes, G. Formation of a dynamic kinetochore- microtubule interface through assembly of the Dam1 ring complex. Mol. Cell 2005, 17, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.J.; De Wulf, P.; Sorger, P.K.; Harrison, S.C. The yeast DASH complex forms closed rings on microtubules. Nat. Struct. Mol. Biol. 2005, 12, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Cheeseman, I.M.; Kallstrom, G.; Velmurugan, S.; Barnes, G.; Chan, C.S. Functional cooperation of Dam1, Ipl1, and the inner centromere protein (INCENP)-related protein Sli15 during chromosome segregation. J. Cell Biol. 2001, 155, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Nogales, E.; Ramey, V.H. Structure-function insights into the yeast Dam1 kinetochore complex. J. Cell Sci. 2009, 122, 3831–3836. [Google Scholar] [CrossRef]
- Tien, J.F.; Umbreit, N.T.; Gestaut, D.R.; Franck, A.D.; Cooper, J.; Wordeman, L.; Gonen, T.; Asbury, C.L.; Davis, T.N. Cooperation of the Dam1 and Ndc80 kinetochore complexes enhances microtubule coupling and is regulated by aurora B. J. Cell Biol. 2010, 189, 713–723. [Google Scholar] [CrossRef]
- Gestaut, D.R.; Graczyk, B.; Cooper, J.; Widlund, P.O.; Zelter, A.; Wordeman, L.; Asbury, C.L.; Davis, T.N. Phosphoregulation and depolymerization-driven movement of the Dam1 complex do not require ring formation. Nat. Cell Biol. 2008, 10, 407–414. [Google Scholar] [CrossRef]
- Chan, Y.W.; Jeyaprakash, A.A.; Nigg, E.A.; Santamaria, A. Aurora B controls kinetochore-microtubule attachments by inhibiting Ska complex-KMN network interaction. J. Cell Biol. 2012, 196, 563–571. [Google Scholar] [CrossRef]
- Boeckmann, L.; Takahashi, Y.; Au, W.C.; Mishra, P.K.; Choy, J.S.; Dawson, A.R.; Szeto, M.Y.; Waybright, T.J.; Heger, C.; McAndrew, C.; et al. Phosphorylation of centromeric histone H3 variant regulates chromosome segregation in Saccharomyces cerevisiae. Mol. Biol. Cell 2013, 24, 2034–2044. [Google Scholar] [CrossRef]
- Akiyoshi, B.; Nelson, C.R.; Ranish, J.A.; Biggins, S. Analysis of Ipl1-mediated phosphorylation of the Ndc80 kinetochore protein in Saccharomyces cerevisiae. Genetics 2009, 183, 1591–1595. [Google Scholar] [CrossRef] [PubMed]
- Deluca, J.G.; Gall, W.E.; Ciferri, C.; Cimini, D.; Musacchio, A.; Salmon, E.D. Kinetochore microtubule dynamics and attachment stability are regulated by hec1. Cell 2006, 127, 969–982. [Google Scholar] [CrossRef]
- Biggins, S.; Murray, A.W. The budding yeast protein kinase Ipl1/Aurora allows the absence of tension to activate the spindle checkpoint. Genes Dev. 2001, 15, 3118–3129. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, B.A.; Kung, C.; Shokat, K.M.; Biggins, S. The Ipl1-Aurora protein kinase activates the spindle checkpoint by creating unattached kinetochores. Nat. Cell Biol. 2006, 8, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Liu, H.; Li, P.; Yu, H.G.; Wang, Y. Loss of function of the cik1/kar3 motor complex results in chromosomes with syntelic attachment that are sensed by the tension checkpoint. PLoS Genet. 2012, 8, e1002492. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.; Schiebel, E. Separase regulates INCENP-Aurora B anaphase spindle function through Cdc14. Science 2003, 302, 2120–2124. [Google Scholar] [CrossRef]
- Tsukahara, T.; Tanno, Y.; Watanabe, Y. Phosphorylation of the CPC by Cdk1 promotes chromosome bi-orientation. Nature 2010, 467, 719. [Google Scholar] [CrossRef]
- Campbell, C.S.; Desai, A. Tension sensing by Aurora B kinase is independent of survivin-based centromere localization. Nature 2013, 497, 118–121. [Google Scholar] [CrossRef]
- Garcia-Rodriguez, L.J.; Kasciukovic, T.; Denninger, V.; Tanaka, T.U. Aurora B-INCENP Localization at Centromeres/Inner Kinetochores Is Required for Chromosome Bi-orientation in Budding Yeast. Curr. Biol. 2019, 29, 1536–1544 e1534. [Google Scholar] [CrossRef]
- Hardwick, K.G.; Weiss, E.; Luca, F.C.; Winey, M.; Murray, A.W. Activation of the budding yeast spindle assembly checkpoint without mitotic spindle disruption. Science 1996, 273, 953–956. [Google Scholar] [CrossRef]
- Aravamudhan, P.; Goldfarb, A.A.; Joglekar, A.P. The kinetochore encodes a mechanical switch to disrupt spindle assembly checkpoint signalling. Nat. Cell Biol. 2015, 17, 868–879. [Google Scholar] [CrossRef] [PubMed]
- London, N.; Ceto, S.; Ranish, J.A.; Biggins, S. Phosphoregulation of Spc105 by Mps1 and PP1 Regulates Bub1 Localization to Kinetochores. Curr. Biol. 2012, 22, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Primorac, I.; Weir, J.R.; Chiroli, E.; Gross, F.; Hoffmann, I.; van Gerwen, S.; Ciliberto, A.; Musacchio, A. Bub3 reads phosphorylated MELT repeats to promote spindle assembly checkpoint signaling. eLife 2013, 2, e01030. [Google Scholar] [CrossRef] [PubMed]
- London, N.; Biggins, S. Mad1 kinetochore recruitment by Mps1-mediated phosphorylation of Bub1 signals the spindle checkpoint. Genes Dev. 2014, 28, 140–152. [Google Scholar] [CrossRef]
- Shepperd, L.A.; Meadows, J.C.; Sochaj, A.M.; Lancaster, T.C.; Zou, J.; Buttrick, G.J.; Rappsilber, J.; Hardwick, K.G.; Millar, J.B. Phosphodependent Recruitment of Bub1 and Bub3 to Spc7/KNL1 by Mph1 Kinase Maintains the Spindle Checkpoint. Curr. Biol. 2012, 22, 891–899. [Google Scholar] [CrossRef]
- Vleugel, M.; Omerzu, M.; Groenewold, V.; Hadders, M.A.; Lens, S.M.A.; Kops, G. Sequential multisite phospho-regulation of KNL1-BUB3 interfaces at mitotic kinetochores. Mol. Cell 2015, 57, 824–835. [Google Scholar] [CrossRef]
- Yamagishi, Y.; Yang, C.H.; Tanno, Y.; Watanabe, Y. MPS1/Mph1 phosphorylates the kinetochore protein KNL1/Spc7 to recruit SAC components. Nat. Cell Biol. 2012, 14, 746–752. [Google Scholar] [CrossRef]
- Ji, Z.; Gao, H.; Jia, L.; Li, B.; Yu, H. A sequential multi-target Mps1 phosphorylation cascade promotes spindle checkpoint signaling. eLife 2017, 6, e22513. [Google Scholar] [CrossRef]
- Maure, J.F.; Kitamura, E.; Tanaka, T.U. Mps1 kinase promotes sister-kinetochore bi-orientation by a tension-dependent mechanism. Curr. Biol. 2007, 17, 2175–2182. [Google Scholar] [CrossRef]
- Shimogawa, M.M.; Graczyk, B.; Gardner, M.K.; Francis, S.E.; White, E.A.; Ess, M.; Molk, J.N.; Ruse, C.; Niessen, S.; Yates, J.R., 3rd; et al. Mps1 phosphorylation of Dam1 couples kinetochores to microtubule plus ends at metaphase. Curr. Biol. 2006, 16, 1489–1501. [Google Scholar] [CrossRef]
- Liang, F.; Jin, F.; Liu, H.; Wang, Y. The molecular function of the yeast polo-like kinase Cdc5 in Cdc14 release during early anaphase. Mol. Biol. Cell 2009, 20, 3671–3679. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wang, Y.; Liu, D.; Li, Y.; Qin, J.; Elledge, S.J. Regulation of the Bub2/Bfa1 GAP complex by Cdc5 and cell cycle checkpoints. Cell 2001, 107, 655–665. [Google Scholar] [CrossRef]
- Stegmeier, F.; Visintin, R.; Amon, A. Separase, polo kinase, the kinetochore protein Slk19, and Spo12 function in a network that controls Cdc14 localization during early anaphase. Cell 2002, 108, 207–220. [Google Scholar] [CrossRef]
- Rossio, V.; Galati, E.; Ferrari, M.; Pellicioli, A.; Sutani, T.; Shirahige, K.; Lucchini, G.; Piatti, S. The RSC chromatin-remodeling complex influences mitotic exit and adaptation to the spindle assembly checkpoint by controlling the Cdc14 phosphatase. J. Cell Biol. 2010, 191, 981–997. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Ciftci-Yilmaz, S.; Reynolds, D.; Au, W.C.; Boeckmann, L.; Dittman, L.E.; Jowhar, Z.; Pachpor, T.; Yeh, E.; Baker, R.E.; et al. Polo kinase Cdc5 associates with centromeres to facilitate the removal of centromeric cohesin during mitosis. Mol. Biol. Cell 2016, 27, 2286–2300. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, G.; Uhlmann, F.; Mechtler, K.; Poupart, M.A.; Nasmyth, K. Phosphorylation of the cohesin subunit Scc1 by Polo/Cdc5 kinase regulates sister chromatid separation in yeast. Cell 2001, 105, 459–472. [Google Scholar] [CrossRef]
- Uhlmann, F.; Wernic, D.; Poupart, M.A.; Koonin, E.V.; Nasmyth, K. Cleavage of cohesin by the CD clan protease separin triggers anaphase in yeast. Cell 2000, 103, 375–386. [Google Scholar] [CrossRef]
- Baldwin, M.L.; Julius, J.A.; Tang, X.; Wang, Y.; Bachant, J. The yeast SUMO isopeptidase Smt4/Ulp2 and the polo kinase Cdc5 act in an opposing fashion to regulate sumoylation in mitosis and cohesion at centromeres. Cell Cycle 2009, 8, 3406–3419. [Google Scholar] [CrossRef][Green Version]
- Mishra, P.K.; Olafsson, G.; Boeckmann, L.; Westlake, T.J.; Jowhar, Z.M.; Dittman, L.E.; Baker, R.E.; D’Amours, D.; Thorpe, P.H.; Basrai, M.A. Cell cycle-dependent association of polo kinase Cdc5 with CENP-A contributes to faithful chromosome segregation in budding yeast. Mol. Biol. Cell 2019, 30, 1020–1036. [Google Scholar] [CrossRef]
- Mishra, P.K.; Basrai, M.A. Protein kinases in mitotic phosphorylation of budding yeast CENP-A. Curr. Genet. 2019, 65, 1325–1332. [Google Scholar] [CrossRef]
- De Wulf, P.; Montani, F.; Visintin, R. Protein phosphatases take the mitotic stage. Curr. Opin. Cell Biol. 2009, 21, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Gelens, L.; Qian, J.; Bollen, M.; Saurin, A.T. The Importance of Kinase-Phosphatase Integration: Lessons from Mitosis. Trends Cell Biol. 2018, 28, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Bokros, M.; Wang, Y. The phosphorylation of a kinetochore protein Dam1 by Aurora B/Ipl1 kinase promotes chromosome bipolar attachment in yeast. Sci. Rep. 2017, 7, 11880. [Google Scholar] [CrossRef] [PubMed]
- Keating, P.; Rachidi, N.; Tanaka, T.U.; Stark, M.J. Ipl1-dependent phosphorylation of Dam1 is reduced by tension applied on kinetochores. J. Cell Sci. 2009, 122, 4375–4382. [Google Scholar] [CrossRef]
- Emanuele, M.J.; Lan, W.; Jwa, M.; Miller, S.A.; Chan, C.S.; Stukenberg, P.T. Aurora B kinase and protein phosphatase 1 have opposing roles in modulating kinetochore assembly. J. Cell Biol. 2008, 181, 241–254. [Google Scholar] [CrossRef]
- Stark, M.J. Yeast protein serine/threonine phosphatases: Multiple roles and diverse regulation. Yeast 1996, 12, 1647–1675. [Google Scholar] [CrossRef]
- Barford, D.; Das, A.K.; Egloff, M.P. The structure and mechanism of protein phosphatases: Insights into catalysis and regulation. Annu. Rev. Biophys. Biomol. Struct. 1998, 27, 133–164. [Google Scholar] [CrossRef]
- Pinsky, B.A.; Kotwaliwale, C.V.; Tatsutani, S.Y.; Breed, C.A.; Biggins, S. Glc7/protein phosphatase 1 regulatory subunits can oppose the Ipl1/aurora protein kinase by redistributing Glc7. Mol. Cell Biol. 2006, 26, 2648–2660. [Google Scholar] [CrossRef]
- Francisco, L.; Wang, W.; Chan, C.S. Type 1 protein phosphatase acts in opposition to IpL1 protein kinase in regulating yeast chromosome segregation. Mol. Cell Biol. 1994, 14, 4731–4740. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Sun, Z.W.; Li, X.; Reuben, M.; Tatchell, K.; Bishop, D.K.; Grushcow, J.M.; Brame, C.J.; Caldwell, J.A.; Hunt, D.F.; et al. Mitotic phosphorylation of histone H3 is governed by Ipl1/aurora kinase and Glc7/PP1 phosphatase in budding yeast and nematodes. Cell 2000, 102, 279–291. [Google Scholar] [CrossRef]
- Tatchell, K.; Makrantoni, V.; Stark, M.J.; Robinson, L.C. Temperature-sensitive ipl1-2/Aurora B mutation is suppressed by mutations in TOR complex 1 via the Glc7/PP1 phosphatase. Proc. Natl. Acad. Sci. USA 2011, 108, 3994–3999. [Google Scholar] [CrossRef] [PubMed]
- Bloecher, A.; Tatchell, K. Defects in Saccharomyces cerevisiae protein phosphatase type I activate the spindle/kinetochore checkpoint. Genes Dev. 1999, 13, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Sassoon, I.; Severin, F.F.; Andrews, P.D.; Taba, M.R.; Kaplan, K.B.; Ashford, A.J.; Stark, M.J.; Sorger, P.K.; Hyman, A.A. Regulation of Saccharomyces cerevisiae kinetochores by the type 1 phosphatase Glc7p. Genes Dev. 1999, 13, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, B.A.; Nelson, C.R.; Biggins, S. Protein phosphatase 1 regulates exit from the spindle checkpoint in budding yeast. Curr. Biol. 2009, 19, 1182–1187. [Google Scholar] [CrossRef]
- Rosenberg, J.S.; Cross, F.R.; Funabiki, H. KNL1/Spc105 recruits PP1 to silence the spindle assembly checkpoint. Curr. Biol. 2011, 21, 942–947. [Google Scholar] [CrossRef]
- Kemmler, S.; Stach, M.; Knapp, M.; Ortiz, J.; Pfannstiel, J.; Ruppert, T.; Lechner, J. Mimicking Ndc80 phosphorylation triggers spindle assembly checkpoint signalling. EMBO J. 2009, 28, 1099–1110. [Google Scholar] [CrossRef]
- Jin, F.; Wang, Y. The signaling network that silences the spindle assembly checkpoint upon the establishment of chromosome bipolar attachment. Proc. Natl. Acad. Sci. USA 2013, 110, 21036–21041. [Google Scholar] [CrossRef]
- Hoyt, M.A.; He, L.; Totis, L.; Saunders, W.S. Loss of function of Saccharomyces cerevisiae kinesin-related CIN8 and KIP1 is suppressed by KAR3 motor domain mutations. Genetics 1993, 135, 35–44. [Google Scholar]
- Saunders, W.S.; Hoyt, M.A. Kinesin-related proteins required for structural integrity of the mitotic spindle. Cell 1992, 70, 451–458. [Google Scholar] [CrossRef]
- Tytell, J.D.; Sorger, P.K. Analysis of kinesin motor function at budding yeast kinetochores. J. Cell Biol. 2006, 172, 861–874. [Google Scholar] [CrossRef]
- Kotwaliwale, C.V.; Frei, S.B.; Stern, B.M.; Biggins, S. A pathway containing the Ipl1/aurora protein kinase and the spindle midzone protein Ase1 regulates yeast spindle assembly. Dev. Cell 2007, 13, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Gupta, A.; Long, S.K.; Evans, R.; Badger, B.L.; Salmon, E.D.; Biggins, S.; Bloom, K. A Kinesin-5, Cin8, Recruits Protein Phosphatase 1 to Kinetochores and Regulates Chromosome Segregation. Curr. Biol. 2018, 28, 2697–2704 e2693. [Google Scholar] [CrossRef] [PubMed]
- Geiser, J.R.; Schott, E.J.; Kingsbury, T.J.; Cole, N.B.; Totis, L.J.; Bhattacharyya, G.; He, L.; Hoyt, M.A. Saccharomyces cerevisiae genes required in the absence of the CIN8-encoded spindle motor act in functionally diverse mitotic pathways. Mol. Biol. Cell 1997, 8, 1035–1050. [Google Scholar] [CrossRef] [PubMed]
- Woodbury, E.L.; Morgan, D.O. The role of self-association in Fin1 function on the mitotic spindle. J. Biol. Chem. 2007, 282, 32138–32143. [Google Scholar] [CrossRef]
- Woodbury, E.L.; Morgan, D.O. Cdk and APC activities limit the spindle-stabilizing function of Fin1 to anaphase. Nat. Cell Biol. 2007, 9, 106–112. [Google Scholar] [CrossRef]
- Akiyoshi, B.; Nelson, C.R.; Ranish, J.A.; Biggins, S. Quantitative proteomic analysis of purified yeast kinetochores identifies a PP1 regulatory subunit. Genes Dev. 2009, 23, 2887–2899. [Google Scholar] [CrossRef]
- Bokros, M.; Gravenmier, C.; Jin, F.; Richmond, D.; Wang, Y. Fin1-PP1 Helps Clear Spindle Assembly Checkpoint Protein Bub1 from Kinetochores in Anaphase. Cell Rep. 2016, 14, 1074–1085. [Google Scholar] [CrossRef]
- Mocciaro, A.; Schiebel, E. Cdc14: A highly conserved family of phosphatases with non-conserved functions? J. Cell Sci. 2010, 123, 2867–2876. [Google Scholar] [CrossRef]
- Visintin, R.; Hwang, E.S.; Amon, A. Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus. Nature 1999, 398, 818–823. [Google Scholar] [CrossRef]
- Shou, W.; Seol, J.H.; Shevchenko, A.; Baskerville, C.; Moazed, D.; Chen, Z.W.; Jang, J.; Shevchenko, A.; Charbonneau, H.; Deshaies, R.J. Exit from mitosis is triggered by Tem1-dependent release of the protein phosphatase Cdc14 from nucleolar RENT complex. Cell 1999, 97, 233–244. [Google Scholar] [CrossRef]
- Wang, Y.; Ng, T.Y. Phosphatase 2A Negatively Regulates Mitotic Exit in Saccharomyces cerevisiae. Mol. Biol. Cell 2006, 17, 80–89. [Google Scholar] [CrossRef]
- Queralt, E.; Lehane, C.; Novak, B.; Uhlmann, F. Downregulation of PP2A(Cdc55) phosphatase by separase initiates mitotic exit in budding yeast. Cell 2006, 125, 719–732. [Google Scholar] [CrossRef]
- Jin, F.; Liu, H.; Liang, F.; Rizkallah, R.; Hurt, M.M.; Wang, Y. Temporal control of the dephosphorylation of Cdk substrates by mitotic exit pathways in budding yeast. Proc. Natl. Acad. Sci. USA 2008, 105, 16177–16182. [Google Scholar] [CrossRef]
- Visintin, R.; Craig, K.; Hwang, E.S.; Prinz, S.; Tyers, M.; Amon, A. The phosphatase Cdc14 triggers mitotic exit by reversal of Cdk-dependent phosphorylation. Mol. Cell 1998, 2, 709–718. [Google Scholar] [CrossRef]
- Nakajima, Y.; Cormier, A.; Tyers, R.G.; Pigula, A.; Peng, Y.; Drubin, D.G.; Barnes, G. Ipl1/Aurora-dependent phosphorylation of Sli15/INCENP regulates CPC-spindle interaction to ensure proper microtubule dynamics. J. Cell Biol. 2011, 194, 137–153. [Google Scholar] [CrossRef]
- Bokros, M.; Wang, Y. Spindle assembly checkpoint silencing and beyond. Cell Cycle 2016, 15, 1661–1662. [Google Scholar] [CrossRef]
- Shi, Y. Serine/threonine phosphatases: Mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef]
- Kitajima, T.S.; Kawashima, S.A.; Watanabe, Y. The conserved kinetochore protein shugoshin protects centromeric cohesion during meiosis. Nature 2004, 427, 510–517. [Google Scholar] [CrossRef]
- Riedel, C.G.; Katis, V.L.; Katou, Y.; Mori, S.; Itoh, T.; Helmhart, W.; Galova, M.; Petronczki, M.; Gregan, J.; Cetin, B.; et al. Protein phosphatase 2A protects centromeric sister chromatid cohesion during meiosis I. Nature 2006, 441, 53–61. [Google Scholar] [CrossRef]
- Fernius, J.; Hardwick, K.G. Bub1 kinase targets Sgo1 to ensure efficient chromosome biorientation in budding yeast mitosis. PLoS Genet. 2007, 3, e213. [Google Scholar] [CrossRef]
- Kawashima, S.A.; Yamagishi, Y.; Honda, T.; Ishiguro, K.; Watanabe, Y. Phosphorylation of H2A by Bub1 prevents chromosomal instability through localizing shugoshin. Science 2010, 327, 172–177. [Google Scholar] [CrossRef]
- Mishra, P.K.; Thapa, K.S.; Chen, P.; Wang, S.; Hazbun, T.R.; Basrai, M.A. Budding yeast CENP-A(Cse4) interacts with the N-terminus of Sgo1 and regulates its association with centromeric chromatin. Cell Cycle 2018, 17, 11–23. [Google Scholar] [CrossRef]
- Buehl, C.J.; Kuo, M.H. Critical roles of Shugoshin and histones as tension sensors during mitosis. Curr. Genet. 2018, 64, 1215–1219. [Google Scholar] [CrossRef]
- Indjeian, V.B.; Murray, A.W. Budding yeast mitotic chromosomes have an intrinsic bias to biorient on the spindle. Curr. Biol. 2007, 17, 1837–1846. [Google Scholar] [CrossRef]
- Eshleman, H.D.; Morgan, D.O. Sgo1 recruits PP2A to chromosomes to ensure sister chromatid bi-orientation during mitosis. J. Cell Sci. 2014, 127, 4974–4983. [Google Scholar] [CrossRef]
- Peplowska, K.; Wallek, A.U.; Storchova, Z. Sgo1 regulates both condensin and ipl1/aurora B to promote chromosome biorientation. PLoS Genet. 2014, 10, e1004411. [Google Scholar] [CrossRef]
- Verzijlbergen, K.F.; Nerusheva, O.O.; Kelly, D.; Kerr, A.; Clift, D.; de Lima Alves, F.; Rappsilber, J.; Marston, A.L. Shugoshin biases chromosomes for biorientation through condensin recruitment to the pericentromere. eLife 2014, 3, e01374. [Google Scholar] [CrossRef]
- Nerusheva, O.O.; Galander, S.; Fernius, J.; Kelly, D.; Marston, A.L. Tension-dependent removal of pericentromeric shugoshin is an indicator of sister chromosome biorientation. Genes Dev. 2014, 28, 1291–1309. [Google Scholar] [CrossRef]
- Indjeian, V.B.; Stern, B.M.; Murray, A.W. The centromeric protein Sgo1 is required to sense lack of tension on mitotic chromosomes. Science 2005, 307, 130–133. [Google Scholar] [CrossRef]
- Jin, F.; Bokros, M.; Wang, Y. Premature Silencing of the Spindle Assembly Checkpoint Is Prevented by the Bub1-H2A-Sgo1-PP2A Axis in Saccharomyces cerevisiae. Genetics 2017, 205, 1169–1178. [Google Scholar] [CrossRef][Green Version]
- Yellman, C.M.; Burke, D.J. The role of Cdc55 in the spindle checkpoint is through regulation of mitotic exit in Saccharomyces cerevisiae. Mol. Biol. Cell 2006, 17, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Wang, Y. Pds1/Esp1-dependent and -independent sister chromatid separation in mutants defective for protein phosphatase 2A. Proc. Natl. Acad. Sci. USA 2006, 103, 16290–16295. [Google Scholar] [CrossRef] [PubMed]
- Yaakov, G.; Thorn, K.; Morgan, D.O. Separase biosensor reveals that cohesin cleavage timing depends on phosphatase PP2A(Cdc55) regulation. Dev. Cell 2012, 23, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Shu, H.; Qi, W.; Mahmood, N.A.; Mumby, M.C.; Yu, H. PP2A is required for centromeric localization of Sgo1 and proper chromosome segregation. Dev. Cell 2006, 10, 575–585. [Google Scholar] [CrossRef]
- Liu, H.; Rankin, S.; Yu, H. Phosphorylation-enabled binding of SGO1-PP2A to cohesin protects sororin and centromeric cohesion during mitosis. Nat. Cell Biol. 2013, 15, 40–49. [Google Scholar] [CrossRef]
- Meppelink, A.; Kabeche, L.; Vromans, M.J.; Compton, D.A.; Lens, S.M. Shugoshin-1 balances Aurora B kinase activity via PP2A to promote chromosome bi-orientation. Cell Rep. 2015, 11, 508–515. [Google Scholar] [CrossRef]
- Foley, E.A.; Maldonado, M.; Kapoor, T.M. Formation of stable attachments between kinetochores and microtubules depends on the B56-PP2A phosphatase. Nat. Cell Biol. 2011, 13, 1265–1271. [Google Scholar] [CrossRef]
- Hauf, S.; Watanabe, Y. Kinetochore orientation in mitosis and meiosis. Cell 2004, 119, 317–327. [Google Scholar] [CrossRef]
- Toth, A.; Rabitsch, K.P.; Galova, M.; Schleiffer, A.; Buonomo, S.B.; Nasmyth, K. Functional genomics identifies monopolin: A kinetochore protein required for segregation of homologs during meiosis i. Cell 2000, 103, 1155–1168. [Google Scholar] [CrossRef]
- Rabitsch, K.P.; Petronczki, M.; Javerzat, J.P.; Genier, S.; Chwalla, B.; Schleiffer, A.; Tanaka, T.U.; Nasmyth, K. Kinetochore recruitment of two nucleolar proteins is required for homolog segregation in meiosis I. Dev. Cell 2003, 4, 535–548. [Google Scholar] [CrossRef]
- Petronczki, M.; Matos, J.; Mori, S.; Gregan, J.; Bogdanova, A.; Schwickart, M.; Mechtler, K.; Shirahige, K.; Zachariae, W.; Nasmyth, K. Monopolar attachment of sister kinetochores at meiosis I requires casein kinase 1. Cell 2006, 126, 1049–1064. [Google Scholar] [CrossRef]
- Sarkar, S.; Shenoy, R.T.; Dalgaard, J.Z.; Newnham, L.; Hoffmann, E.; Millar, J.B.; Arumugam, P. Monopolin subunit Csm1 associates with MIND complex to establish monopolar attachment of sister kinetochores at meiosis I. PLoS Genet. 2013, 9, e1003610. [Google Scholar] [CrossRef]
- Monje-Casas, F.; Prabhu, V.R.; Lee, B.H.; Boselli, M.; Amon, A. Kinetochore orientation during meiosis is controlled by Aurora B and the monopolin complex. Cell 2007, 128, 477–490. [Google Scholar] [CrossRef]
- Meyer, R.E.; Kim, S.; Obeso, D.; Straight, P.D.; Winey, M.; Dawson, D.S. Mps1 and Ipl1/Aurora B act sequentially to correctly orient chromosomes on the meiotic spindle of budding yeast. Science 2013, 339, 1071–1074. [Google Scholar] [CrossRef]
- Buonomo, S.B.; Clyne, R.K.; Fuchs, J.; Loidl, J.; Uhlmann, F.; Nasmyth, K. Disjunction of homologous chromosomes in meiosis I depends on proteolytic cleavage of the meiotic cohesin Rec8 by separin. Cell 2000, 103, 387–398. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nurse, P. Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature 1999, 400, 461–464. [Google Scholar] [CrossRef]
- Katis, V.L.; Lipp, J.J.; Imre, R.; Bogdanova, A.; Okaz, E.; Habermann, B.; Mechtler, K.; Nasmyth, K.; Zachariae, W. Rec8 phosphorylation by casein kinase 1 and Cdc7-Dbf4 kinase regulates cohesin cleavage by separase during meiosis. Dev. Cell 2010, 18, 397–409. [Google Scholar] [CrossRef]
- Ishiguro, T.; Tanaka, K.; Sakuno, T.; Watanabe, Y. Shugoshin-PP2A counteracts casein-kinase-1-dependent cleavage of Rec8 by separase. Nat. Cell Biol. 2010, 12, 500–506. [Google Scholar] [CrossRef]
- Arguello-Miranda, O.; Zagoriy, I.; Mengoli, V.; Rojas, J.; Jonak, K.; Oz, T.; Graf, P.; Zachariae, W. Casein Kinase 1 Coordinates Cohesin Cleavage, Gametogenesis, and Exit from M Phase in Meiosis II. Dev. Cell 2017, 40, 37–52. [Google Scholar] [CrossRef]
- Katis, V.L.; Galova, M.; Rabitsch, K.P.; Gregan, J.; Nasmyth, K. Maintenance of cohesin at centromeres after meiosis I in budding yeast requires a kinetochore-associated protein related to MEI-S332. Curr. Biol. 2004, 14, 560–572. [Google Scholar] [CrossRef]
- Marston, A.L.; Tham, W.H.; Shah, H.; Amon, A. A genome-wide screen identifies genes required for centromeric cohesion. Science 2004, 303, 1367–1370. [Google Scholar] [CrossRef]
- Jonak, K.; Zagoriy, I.; Oz, T.; Graf, P.; Rojas, J.; Mengoli, V.; Zachariae, W. APC/C-Cdc20 mediates deprotection of centromeric cohesin at meiosis II in yeast. Cell Cycle 2017, 16, 1145–1152. [Google Scholar] [CrossRef]
- Yu, H.G.; Koshland, D. The Aurora kinase Ipl1 maintains the centromeric localization of PP2A to protect cohesin during meiosis. J. Cell Biol. 2007, 176, 911–918. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherwin, D.; Wang, Y. The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment. Int. J. Mol. Sci. 2019, 20, 6182. https://doi.org/10.3390/ijms20246182
Sherwin D, Wang Y. The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment. International Journal of Molecular Sciences. 2019; 20(24):6182. https://doi.org/10.3390/ijms20246182
Chicago/Turabian StyleSherwin, Delaney, and Yanchang Wang. 2019. "The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment" International Journal of Molecular Sciences 20, no. 24: 6182. https://doi.org/10.3390/ijms20246182
APA StyleSherwin, D., & Wang, Y. (2019). The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment. International Journal of Molecular Sciences, 20(24), 6182. https://doi.org/10.3390/ijms20246182