Gonadotropin Releasing Hormone Agonists Have an Anti-apoptotic Effect on Cumulus Cells


 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. GnRH Receptors
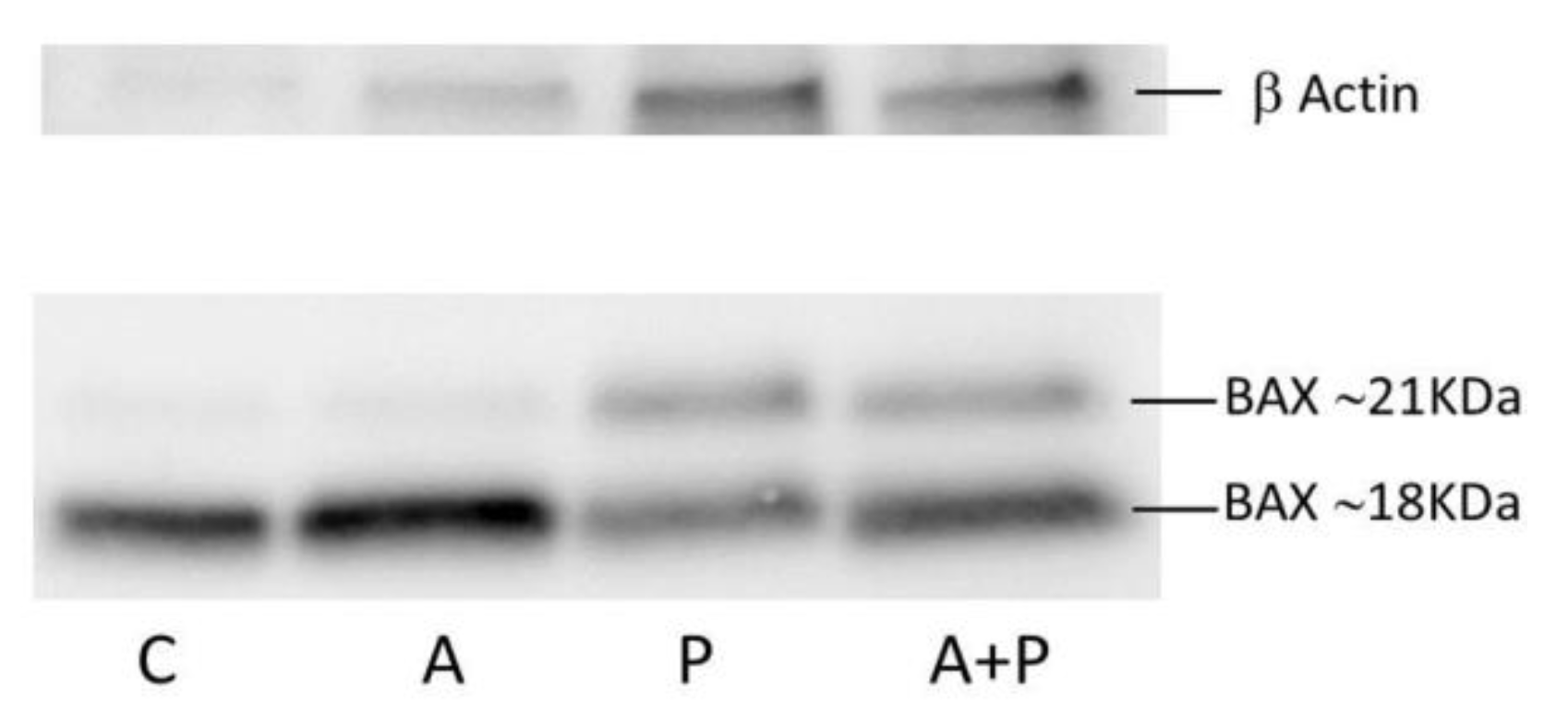
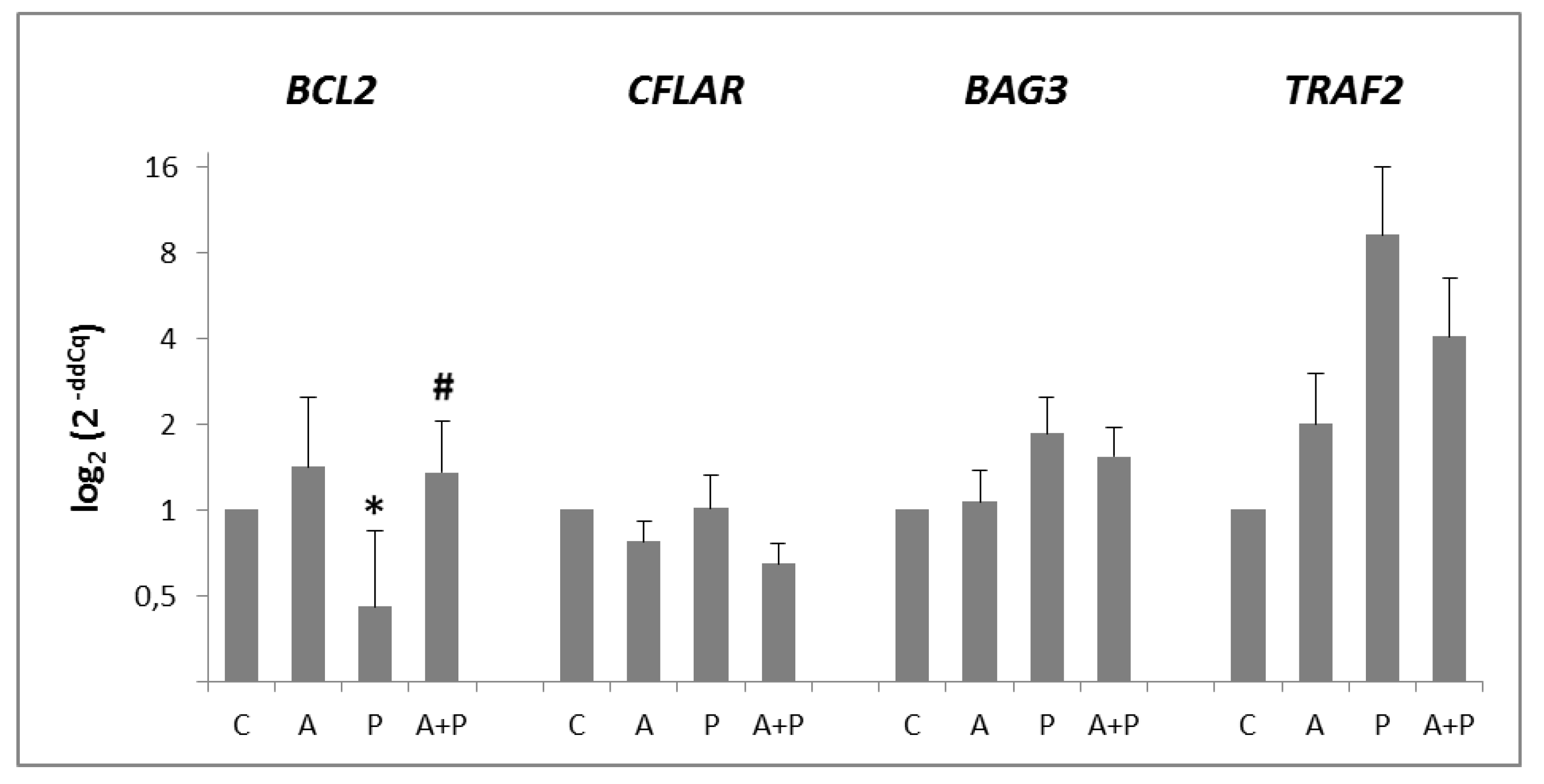
2.2. Apoptosis Pathway
2.3. Sphingomyelin Pathway
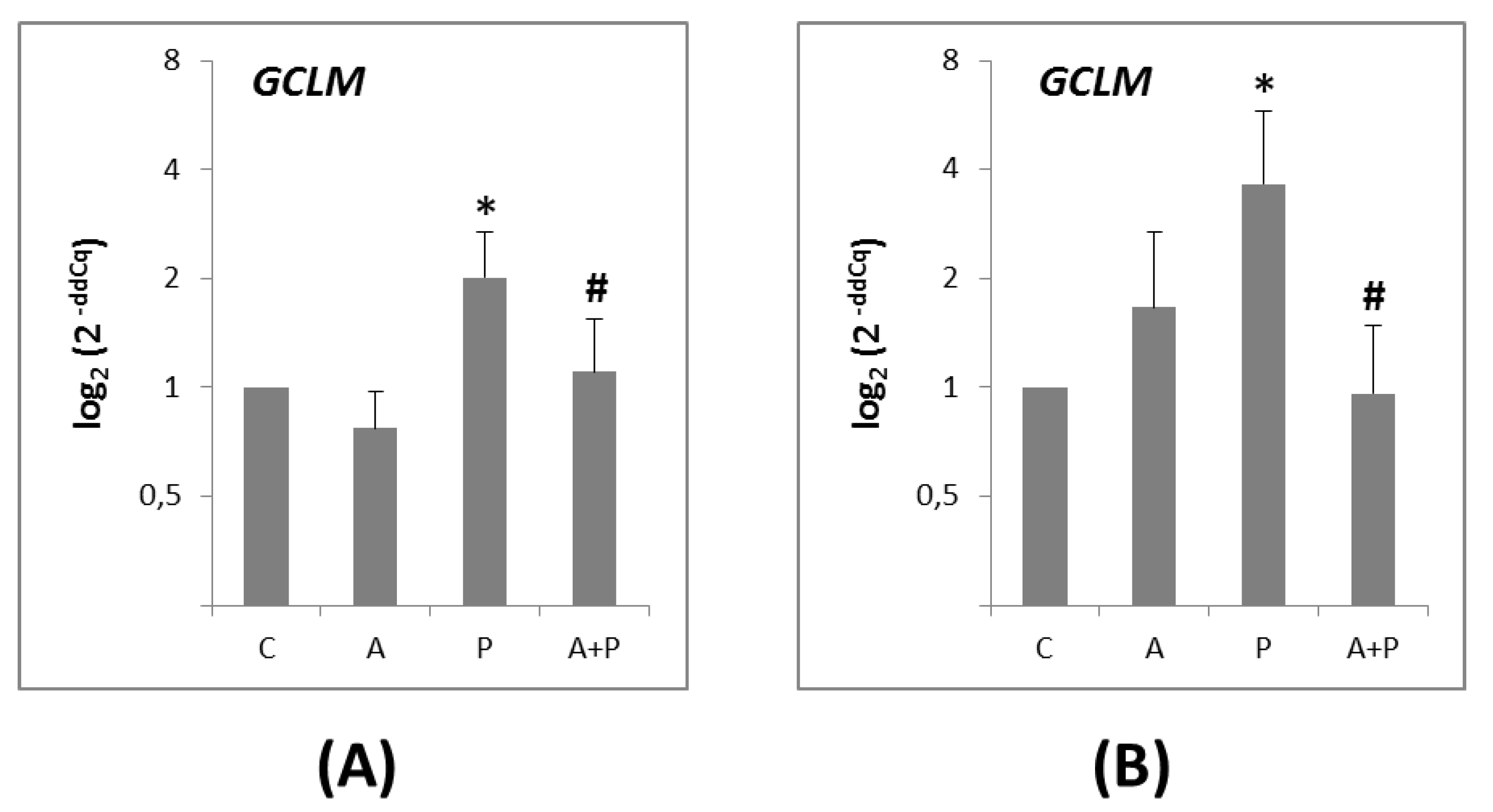
2.4. Glutathione-Mediated Pathway
3. Discussion
4. Materials and Methods
4.1. COC Collection
4.2. Ethical Approval
4.3. Study Design
4.4. In Vitro Treatments
4.5. RNA Isolation, Quantitation and Retro-Transcription
4.6. Quantitative PCR (qPCR)
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GnRHa | Gonadotropin Releasing Hormone agonists |
| COC | Cumulus cell-Oocyte Complexes |
| CC | Cumulus Cells |
| GC | Granulosa Cells |
| IVF | In Vitro Fertilization |
| HSA | Human Serum Albumin |
References
- Wallace, W.H.; Kelsey, T.W. Human ovarian reserve from conception to the menopause. PLoS ONE 2010, 5, e8772. [Google Scholar] [CrossRef] [PubMed]
- Meirow, D.; Nugent, D. The effects of radiotherapy and chemotherapy on female reproduction. Hum. Reprod. Update 2001, 7, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Oktem, O.; Oktay, K. A novel ovarian xenografting model to characterize the impact of chemotherapy agents on human primordial follicle reserve. Cancer Res. 2007, 67, 10159–10162. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.; Anderson, R.A.; Gourley, C.; Wallace, W.H.; Spears, N. How do chemotherapeutic agents damage the ovary? Hum. Reprod. Update 2012, 18, 525–535. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.M. Fertility Preservation in Women. N. Engl. J. Med. 2017, 377, 1657–1665. [Google Scholar] [CrossRef]
- Lambertini, M.; Del Mastro, L.; Pescio, M.C.; Andersen, C.Y.; Azim, H.A.; Peccatori, F.A.; Costa, M.; Revelli, A.; Salvagno, F.; Gennari, A.; et al. Cancer and fertility preservation: International recommendations from an expert meeting. BMC Med. 2016, 14, 1. [Google Scholar] [CrossRef]
- Lambertini, M.; Cinquini, M.; Moschetti, I.; Peccatori, F.A.; Anserini, P.; Valenzano Menada, M.; Tomirotti, M.; Del Mastro, L. Temporary ovarian suppression during chemotherapy to preserve ovarian function and fertility in breast cancer patients: A GRADE approach for evidence evaluation and recommendations by the Italian Association of Medical Oncology. Eur. J. Cancer 2017, 71, 25–33. [Google Scholar] [CrossRef]
- Oktay, K.; Harvey, B.E.; Partridge, A.H.; Quinn, G.P.; Reinecke, J.; Taylor, H.S.; Wallace, W.H.; Wang, E.T.; Loren, A.W. Fertility preservation in patients with cancer: ASCO clinical practice guideline update. J. Clin. Oncol. 2018, 36, 1994–2001. [Google Scholar] [CrossRef]
- Paluch-Shimon, S.; Pagani, O.; Partridge, A.H.; Abulkhair, O.; Cardoso, M.J.; Dent, R.A.; Gelmon, K.; Gentilini, O.; Harbeck, N.; Margulies, A.; et al. ESO-ESMO 3rd international consensus guidelines for breast cancer in young women (BCY3). Breast 2017, 35, 203–217. [Google Scholar] [CrossRef]
- Lambertini, M.; Horicks, F.; Del Mastro, L.; Partridge, A.H.; Demeestere, I. Ovarian protection with gonadotropin-releasing hormone agonists during chemotherapy in cancer patients: From biological evidence to clinical application. Cancer Treat. Rev. 2019, 72, 65–77. [Google Scholar] [CrossRef]
- Lambertini, M.; Moore, H.C.F.; Leonard, R.C.F.; Loibl, S.; Munster, P.; Bruzzone, M.; Boni, L.; Unger, J.M.; Anderson, R.A.; Mehta, K.; et al. Gonadotropin-releasing hormone agonists during chemotherapy for preservation of ovarian function and fertility in premenopausal patients with early breast cancer: A systematic review and meta-analysis of individual patient-level data. J. Clin. Oncol. 2018, 36, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Demeestere, I.; Brice, P.; Peccatori, F.A.; Kentos, A.; Dupuis, J.; Zachee, P.; Casasnovas, O.; Van Den Neste, E.; Dechene, J.; De Maertelaer, V.; et al. No Evidence for the Benefit of Gonadotropin-Releasing Hormone Agonist in Preserving Ovarian Function and Fertility in Lymphoma Survivors Treated With Chemotherapy: Final Long-Term Report of a Prospective Randomized Trial. J. Clin. Oncol. 2016, 34, 2568–2574. [Google Scholar] [CrossRef] [PubMed]
- Senra, J.C.; Roque, M.; Talim, M.C.T.; Reis, F.M.; Tavares, R.L.C. Gonadotropin-releasing hormone agonists for ovarian protection during cancer chemotherapy: Systematic review and meta-analysis. Ultrasound Obstet. Gynecol. 2018, 51, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, M.; Ceppi, M.; Poggio, F.; Peccatori, F.A.; Azim, H.A.; Ugolini, D.; Pronzato, P.; Loibl, S.; Moore, H.C.; Partridge, A.H.; et al. Ovarian suppression using luteinizing hormone-releasing hormone agonists during chemotherapy to preserve ovarian function and fertility of breast cancer patients: A meta-analysis of randomized studies. Ann. Oncol. 2015, 26, 2408–2419. [Google Scholar] [CrossRef] [PubMed]
- Munhoz, R.R.; Pereira, A.A.L.; Sasse, A.D.; Hoff, P.M.; Traina, T.A.; Hudis, C.A.; Marques, R.J. Gonadotropin-releasing hormone agonists for ovarian function preservation in premenopausal women undergoing chemotherapy for early-stage breast cancer: A systematic review and meta-analysis. JAMA Oncol. 2016, 2, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Lu, Y.; Wu, K.; Chen, Q.; Ding, L.; Ge, M.; Weng, Z. Protecting effects of gonadotropin releasing hormone agonist on chemotherapy-induced ovarian damage in premenopausal breast cancer patients: A systematic review and meta-analysis. Breast Care 2017, 12, 48–52. [Google Scholar] [CrossRef]
- Danforth, D.R.; Arbogast, L.K.; Friedman, C.I. Acute depletion of murine primordial follicle reserve by gonadotropin-releasing hormone antagonists. Fertil. Steril. 2005, 83, 1333–1338. [Google Scholar] [CrossRef]
- Kanter, M.; Sapmaz-Metin, M.; Serez, B. Effects of GnRHa on early embryonic development in mice receiving cyclophosphamide. Arch. Gynecol. Obstet. 2016, 293, 203–209. [Google Scholar] [CrossRef]
- Meirow, D.; Assad, G.; Dor, J.; Rabinovici, J. The GnRH antagonist cetrorelix reduces cyclophosphamide-induced ovarian follicular destruction in mice. Hum. Reprod. 2004, 19, 1294–1299. [Google Scholar] [CrossRef]
- Matsuo, G.; Ushijima, K.; Shinagawa, A.; Takahashi, S.; Fujiyoshi, N.; Takemoto, S.; Terada, A.; Fukui, A.; Kamura, T. GnRH agonist acts as ovarian protection in chemotherapy induced gonadotoxicity: An experiment using a rat model. Kurume Med. J. 2007, 54, 25–29. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.X.; Tao, X.; Lu, Z.Y.; Wang, J.J.; Feng, W.W.; Hua, K.Q. Goserelin can inhibit ovarian cancer proliferation and simultaneously protect ovarian function from cisplatin: An in vitro and in vivo study. J. Chemother. 2013, 25, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Ataya, K.M.; McKanna, J.A.; Weintraub, A.M.; Clark, M.R.; LeMaire, W.J. A luteinizing hormone-releasing hormone agonist for the prevention of chemotherapy-induced ovarian follicular loss in rats. Cancer Res. 1985, 45, 3651–3656. [Google Scholar] [PubMed]
- Ataya, K.M.; Palmer, K.C.; Blacker, C.M.; Moghissi, K.S.; Mohammad, S.H. Inhibition of rat ovarian [3H]thymidine uptake by luteinizing hormone-releasing hormone agonists: A possible mechanism for preventing damage by cytotoxic agents. Cancer Res. 1988, 48, 7252–7256. [Google Scholar] [PubMed]
- Bokser, L.; Szende, B.; Schally, A.V. Protective effects of D-Trp6-luteinising hormone-releasing hormone microcapsules against cyclophosphamideinduced gonadotoxicity in female rats. Br. J. Cancer 1990, 61, 861–865. [Google Scholar] [CrossRef]
- Li, X.; Kang, X.; Deng, Q.; Cai, J.; Wang, Z. Combination of a GnRH agonist with an antagonist prevents flare-up effects and protects primordial ovarian follicles in the rat ovary from cisplatin-induced toxicity: A controlled experimental animal study. Reprod. Biol. Endocrinol. 2013, 11, 16. [Google Scholar] [CrossRef]
- Horicks, F.; Van Den Steen, G.; Houben, S.; Englert, Y.; Demeestere, I. Folliculogenesis Is Not Fully Inhibited during GnRH Analogues Treatment in Mice Challenging Their Efficiency to Preserve the Ovarian Reserve during Chemotherapy in This Model. PLoS ONE 2015, 10, e0137164. [Google Scholar] [CrossRef]
- Horicks, F.; Van Den Steen, G.; Gervy, C.; Clarke, H.J.; Demeestere, I. Both in vivo FSH depletion and follicular exposure to Gonadotrophin-releasing hormone analogues in vitro are not effective to prevent follicular depletion during chemotherapy in mice. Mol. Hum. Reprod. 2018, 24, 221–232. [Google Scholar] [CrossRef]
- Detti, L.; Uhlmann, R.A.; Zhang, J.; Diamond, M.P.; Saed, G.M.; Fletcher, N.M.; Lu, M.; Williams, L.J. Goserelin fosters bone elongation but does not prevent ovarian damage in cyclophosphamide-treated prepubertal mice. Fertil. Steril. 2014, 101, 1157:e1–1164:e1. [Google Scholar] [CrossRef]
- Hasky, N.; Uri-Belapolsky, S.; Goldberg, K.; Miller, I.; Grossman, H.; Stemmer, S.M.; Ben-Aharon, I.; Shalgi, R. Gonadotrophin-releasing hormone agonists for fertility preservation: Unraveling the enigma? Hum. Reprod. 2015, 30, 1089–1101. [Google Scholar] [CrossRef]
- Ataya, K.; Rao, L.V.; Lawrence, E.; Kimmel, R. Luteinizing hormone-releasing hormone agonist inhibits cyclophosphamide induced ovarian follicular depletion in rhesus monkeys. Biol. Reprod. 1995, 52, 365–372. [Google Scholar] [CrossRef]
- Imai, A.; Sugiyama, M.; Furui, T.; Tamaya, T.; Ohno, T. Direct protection by a gonadotropin-releasing hormone analog from doxorubicin-induced granulosa cell damage. Gynecol. Obstet. Investig. 2007, 63, 102–106. [Google Scholar] [CrossRef]
- Bildik, G.; Akin, N.; Senbabaoglu, F.; Sahin, G.N.; Karahuseyinoglu, S.; Ince, U.; Taskiran, C.; Selek, U.; Yakin, K.; Guzel, Y.; et al. GnRH agonist leuprolide acetate does not confer any protection against ovarian damage induced by chemotherapy and radiation in vitro. Hum. Reprod. 2015, 30, 2912–2925. [Google Scholar] [CrossRef]
- Torrealday, S.; Lalioti, M.D.; Guzeloglu-Kayisli, O.; Seli, E. Characterization of the gonadotropin releasing hormone receptor (GnRHR) expression and activity in the female mouse ovary. Endocrinology 2013, 154, 3877–3887. [Google Scholar] [CrossRef][Green Version]
- Akgul, C.; Moulding, D.A.; Edwards, S.W. Alternative splicing of Bcl-2-related genes: Functional comnseguences and potential therapeutic applications. Cell Mol. Life Sci. 2004, 61, 2189–2199. [Google Scholar] [CrossRef]
- Blumenfeld, Z.; von Wolff, M. GnRH-analogues and oral contraceptives for fertility preservation in women during chemotherapy. Hum. Reprod. Update 2008, 14, 543–552. [Google Scholar] [CrossRef]
- Blumenfeld, Z. How to preserve fertility in young women exposed to chemotherapy? The role of GnRH agonist cotreatment in addition to cryopreservation of embrya, oocytes, or ovaries. Oncologist 2007, 12, 1044–1054. [Google Scholar] [CrossRef]
- Poggio, F.; Lambertini, M.; Bighin, C.; Conte, B.; Blondeaux, E.; D’Alonzo, A.; Dellepiane, C.; Buzzati, G.; Molinelli, C.; Boccardo, F.; et al. Potential mechanisms of ovarian protection with gonadotropin-releasing hormone agonist in breast cancer patients: A review. Clin. Med. Insights Reprod. Health 2019, 13, 1179558119864584. [Google Scholar] [CrossRef]
- Ozcelik, B.; Turkyilmaz, C.; Ozgun, M.T.; Serin, I.S.; Batukan, C.; Ozdamar, S.; Ozturk, A. Prevention of paclitaxel and cisplatin induced ovarian damage in rats by a gonadotropin-releasing hormone agonist. Fertil. Steril. 2010, 93, 1609–1614. [Google Scholar] [CrossRef]
- Tan, S.J.; Yeh, Y.C.; Shang, W.J.; Wu, G.J.; Liu, J.Y.; Chen, C.H. Protective effect of a gonadotropin-releasing hormone analogue on chemotherapeutic agent-induced ovarian gonadotoxicity: A mouse model. Eur. J. Obstet. Gynecol. Reprod. Biol. 2010, 149, 182–185. [Google Scholar] [CrossRef]
- Lin, Q.Y.; Wang, Y.F.; Weng, H.N.; Sheng, X.J.; Jiang, Q.P.; Yang, Z.Y. Influence of gonadotropin-releasing hormone agonist on the effect of chemotherapy upon ovarian cancer and the prevention of chemotherapy-induced ovarian damage: An experimental study with nu/nu athymic mice. J. Zhejiang Univ. Sci. B 2012, 13, 894–903. [Google Scholar] [CrossRef]
- Kishk, E.A.; Mohammed Ali, M.H. Effect of a gonadotropin-releasing hormone analogue on cyclophosphamide-induced ovarian toxicity in adult mice. Arch. Gynecol. Obstet. 2013, 287, 1023–1029. [Google Scholar] [CrossRef]
- Park, I.; Lee, S.; Ryu, K.J.; Min, K.J.; Hong, J.H.; Song, J.Y.; Lee, J.K.; Lee, N.W. A gonadotropin-releasing hormone agonist for the prevention of docetaxel-induced gonadal damage. J. Obstet. Gynaecol. 2017, 37, 783–789. [Google Scholar] [CrossRef]
- Grundker, C.; Schulz, K.; Gunthert, A.; Emons, G. Luteinizing hormone releasing hormone induced nuclear factor kappaB activation and inhibits apoptosis in ovarian cancer cells. J. Clin. Endocrinol. Metab. 2000, 85, 3810–3820. [Google Scholar] [CrossRef][Green Version]
- Carabatsos, M.J.; Sellitto, C.; Goodenough, D.A.; Albertini, D.F. Oocyte-granulosa cell heterologous gap junctions are required for the coordination of nuclear and cytoplasmic meiotic competence. Dev. Biol. 2000, 226, 167–179. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Lane, M.; Thompson, J.G. Oocyte-secreted factors: Regulators of cumulus cell function and oocyte quality. Hum. Reprod. Update 2008, 14, 159–177. [Google Scholar] [CrossRef]
- Fischer, B.; Bavister, B.D. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J. Reprod. Fertil. 1993, 99, 673–679. [Google Scholar] [CrossRef]
- Catt, J.W.; Henman, M. Toxic effects of oxygen on human embryo development. Hum. Reprod. 2000, 15, 199–206. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Choi, J.H.; Gilks, C.B.; Auersperg, N.; Leung, P.C. Immunolocalization of gonadotropin-releasing hormone (GnRH)-I, GnRH-II, and type I GnRH receptor during follicular development in the human ovary. J. Clin. Endocrinol. Metab. 2006, 91, 4562–4570. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef]
- Singh, S.; Khan, A.R.; Gupta, A.K. Role of glutathione in cancer pathophysiology and therapeutic interventions. J. Exp. Ther. Oncol. 2012, 9, 303–316. [Google Scholar]
- Gurtoo, H.L.; Hipkens, J.H.; Sharma, S.D. Role of glutathione in the metabolism-dependent toxicity and chemotherapy of cyclophosphamide. Cancer Res. 1981, 41, 3584–3591. [Google Scholar]
- Del Mastro, L.; Boni, L.; Michelotti, A.; Gamucci, T.; Olmeo, N.; Gori, S.; Giordano, M.; Garrone, O.; Pronzato, P.; Bighin, C.; et al. Effect of the gonadotropin-releasing hormone analogue triptorelin on the occurrence of chemotherapy-induced early menopause in premenopausal women with breast cancer: A randomized trial. JAMA 2011, 306, 269–276. [Google Scholar] [CrossRef]
- Liang, L.; Dunn, J.P.; Chen, S.; Tsai, M.S.; Hornburg, D.; Newmann, S.; Chung, P.; Avina, M.; Leng, Y.; Holman, R.; et al. Smart Diaphragm Study: Multi-omics profiling and cervical device measurements during pregnancy. AJOG 2019, 220, S649. [Google Scholar] [CrossRef]
- Minde, D.P.; Dunker, A.K.; Lilley, K.S. Time, space, and disorder in the expanding proteome universe. Proteomics 2017, 17, 7. [Google Scholar] [CrossRef]
- La Marca, A.; Spada, E.; Grisendi, V.; Argento, C.; Papaleo, E.; Milani, S.; Volpe, A. Normal serum anti-Müllerian hormone levels in the general female population and the relationship with reproductive history. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 163, 180–184. [Google Scholar] [CrossRef]
- Stigliani, S.; Moretti, S.; Anserini, P.; Casciano, I.; Venturini, P.L.; Scaruffi, P. Storage time does not modify the gene expression profile of cryopreserved human metaphase II oocytes. Hum. Reprod. 2015, 30, 2519–2526. [Google Scholar] [CrossRef]
- Rienzi, L.; Balaban, B.; Ebner, T.; Mandelbaum, J. The oocyte. Hum. Reprod. 2012, 27, i2–i21. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Santonocito, M.; Guglielmino, M.R.; Vento, M.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Casciano, I.; Scollo, P.; Romani, M.; Tatone, C.; et al. The apoptotic transcriptome of the human MII oocyte: Characterization and age-related changes. Apoptosis 2013, 18, 201–211. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scaruffi, P.; Stigliani, S.; Cardinali, B.; Massarotti, C.; Lambertini, M.; Sozzi, F.; Dellepiane, C.; Merlo, D.F.; Anserini, P.; Del Mastro, L. Gonadotropin Releasing Hormone Agonists Have an Anti-apoptotic Effect on Cumulus Cells. Int. J. Mol. Sci. 2019, 20, 6045. https://doi.org/10.3390/ijms20236045
Scaruffi P, Stigliani S, Cardinali B, Massarotti C, Lambertini M, Sozzi F, Dellepiane C, Merlo DF, Anserini P, Del Mastro L. Gonadotropin Releasing Hormone Agonists Have an Anti-apoptotic Effect on Cumulus Cells. International Journal of Molecular Sciences. 2019; 20(23):6045. https://doi.org/10.3390/ijms20236045
Chicago/Turabian StyleScaruffi, Paola, Sara Stigliani, Barbara Cardinali, Claudia Massarotti, Matteo Lambertini, Fausta Sozzi, Chiara Dellepiane, Domenico Franco Merlo, Paola Anserini, and Lucia Del Mastro. 2019. "Gonadotropin Releasing Hormone Agonists Have an Anti-apoptotic Effect on Cumulus Cells" International Journal of Molecular Sciences 20, no. 23: 6045. https://doi.org/10.3390/ijms20236045
APA StyleScaruffi, P., Stigliani, S., Cardinali, B., Massarotti, C., Lambertini, M., Sozzi, F., Dellepiane, C., Merlo, D. F., Anserini, P., & Del Mastro, L. (2019). Gonadotropin Releasing Hormone Agonists Have an Anti-apoptotic Effect on Cumulus Cells. International Journal of Molecular Sciences, 20(23), 6045. https://doi.org/10.3390/ijms20236045