Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis



Abstract
1. Introduction
- (i)
- to investigate the more detailed effects of GnRH and its derivatives in Tetrahymena;
- (ii)
- to find some correlation between the different cell physiological reactions of Tetrahymena induced by various GnRH derivatives;
- (iii)
- to evaluate the selectivity of chemosensory behaviors of Tetrahymena and, consequently, the selectivity of its receptors;
- (iv)
- to interpret the roles of GnRH peptides at the unicellular level of the phylogeny.
2. Results
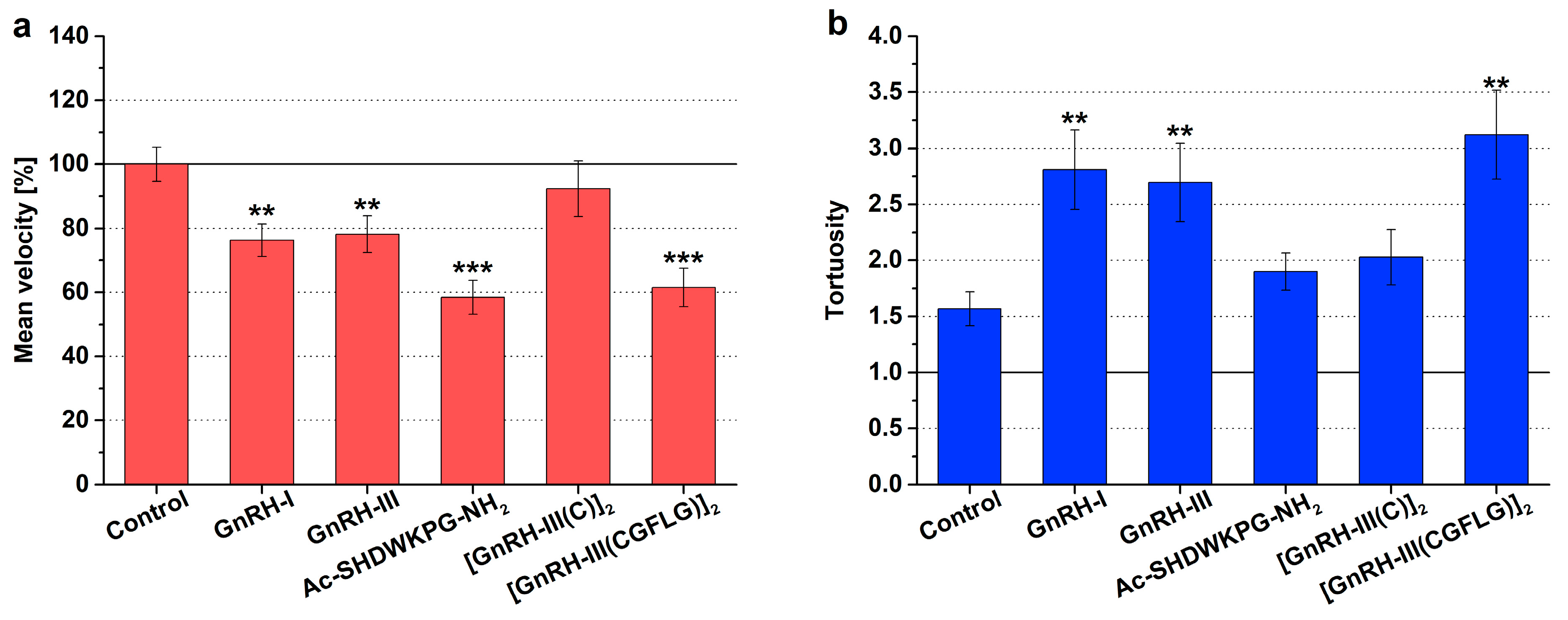
2.1. Chemokinesis Swimming Behavior
2.2. Hormone Content
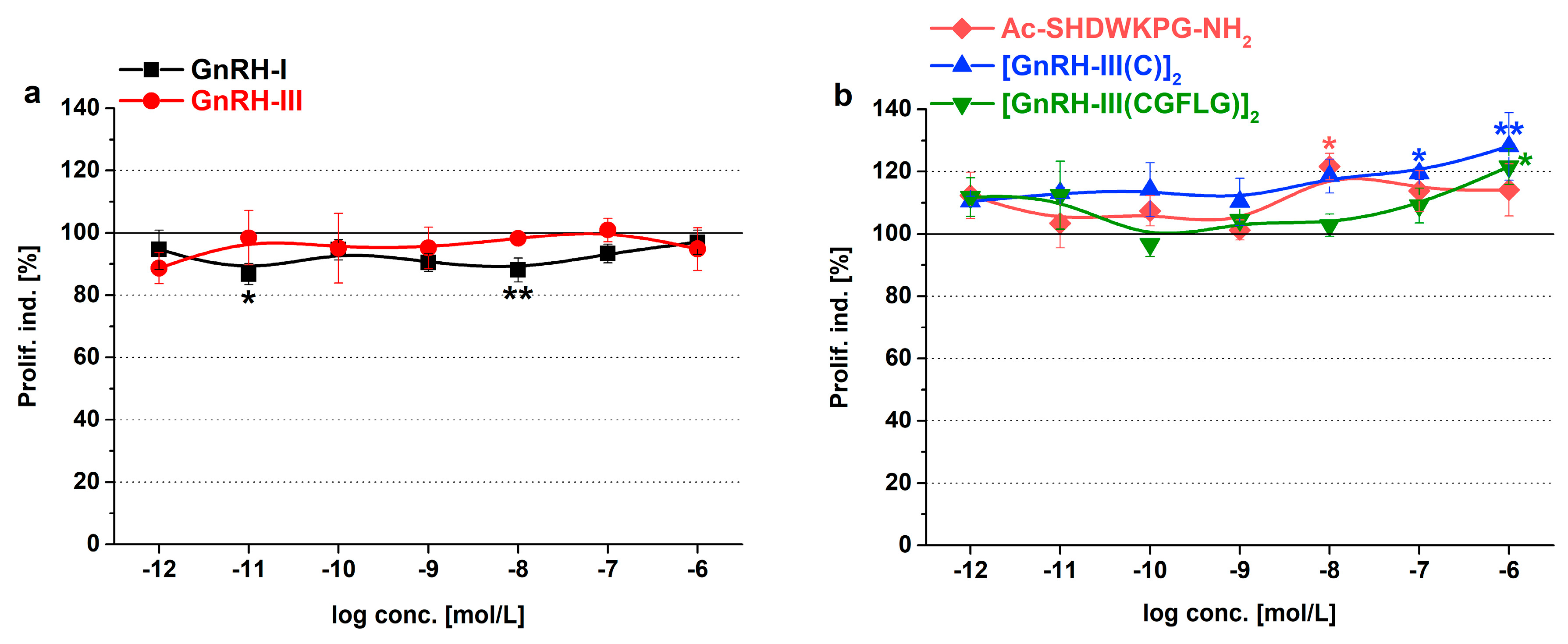
2.3. Cell Proliferation
3. Discussion
3.1. Migratory Responses of Tetrahymena
3.2. Effects of GnRH Derivatives on the Hormone Contents of Tetrahymena
3.3. Effect of GnRH Peptides on the Proliferation of Tetrahymena
3.4. Evolutionary Aspects of Tetrahymena’s Reactions to GnRH Peptides
4. Conclusions
5. Materials and Methods
5.1. Tested Peptides
5.2. Model Cell
5.3. Study of Swimming Behavior
5.4. Analysis of Intracellular Hormone Content
5.5. Study of Cell Growth
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| ESI-MS | Electrospray ionization mass spectrometry |
| FITC | Fluorescein isothiocyanate |
| GeoMean | Geometric mean channel |
| GnRH | Gonadotropin-releasing hormone |
| hCG | Human chorionic-gonadotropic hormone |
| Prolif. Ind. | Proliferation index |
| SPPS | Solid-phase peptide synthesis |
| T3 | Triiodothyronine |
| TSH | Thyroid-stimulating hormone |
| Tort | Tortuosity |
References
- Christopher, G.K.; Sundermann, C.A. Isolation and partial characterization of the insulin binding sites of Tetrahymena pyriformis. Biochem. Biophys. Res. Commun. 1995, 212, 515–523. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.B.; Pert, C.B.; Ruff, M.R.; Smith, C.C.; Higgins, W.J.; Zipser, B. Identification and characterization of the opiate receptor in the ciliated protozoan, Tetrahymena. Brain Res. 1988, 450, 303–315. [Google Scholar] [CrossRef]
- Csaba, G. The hormonal system of the unicellular Tetrahymena: A review with evolutionary aspects. Acta Microbiol. Immunol. Hung. 2012, 59, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Essman, E.J. The serotonergic system in Tetrahymena pyriformis. Ric. Clin. Lab. 1987, 17, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Le Roith, D.; Shiloach, J.; Roth, J.; Lesniak, M.A. Evolutionary origins of vertebrate hormones: Substances similar to mammalian insulins are native to unicellular eukaryotes. Proc. Natl. Acad. Sci. USA 1980, 77, 6184–6188. [Google Scholar] [CrossRef]
- Hegyesi, H.; Kovács, P.; Falus, A.; Csaba, G. Presence and localization of histidine decarboxylase enzyme (HDC) and histamine in Tetrahymena pyriformis. Cell Biol. Int. 1998, 22, 493–497. [Google Scholar] [CrossRef]
- Csaba, G.; Kovács, P. Localization of beta-endorphin in tetrahymena by confocal microscopy. Induction of the prolonged production of the hormone by hormonal imprinting. Cell Biol. Int. 1999, 23, 695–702. [Google Scholar] [CrossRef]
- Csaba, G.; Nagy, U. Effect of vertebrate hormones on the cyclic AMP level in Tetrahymena. Acta Biol. Med. Ger. 1976, 35, 1399–1401. [Google Scholar]
- Jamieson, G.A., Jr.; Vanaman, T.C.; Blum, J.J. Presence of calmodulin in Tetrahymena. Proc. Natl. Acad. Sci. USA 1979, 76, 6471–6475. [Google Scholar] [CrossRef]
- Kőhidai, L.; Bársony, J.; Roth, J.; Marx, S.J. Rapid effects of insulin on cyclic GMP location in an intact protozoan. Experientia 1992, 48, 476–481. [Google Scholar] [CrossRef]
- Le Roith, D.; Shiloach, J.; Berelowitz, M.; Frohman, L.A.; Liotta, A.S.; Krieger, D.T.; Roth, J. Are messenger molecules in microbes the ancestors of the vertebrate hormones and tissue factors? Fed. Proc. 1983, 42, 2602–2607. [Google Scholar] [PubMed]
- Kőhidai, L.; Karsa, J.; Csaba, G. Effects of hormones on the chemotaxis in Tetrahymena—Investigations on receptor memory. Microbios 1994, 77, 75–85. [Google Scholar] [PubMed]
- Kovács, P.; Lovas, G.; Csaba, G. Influence of insulin on the movement of Tetrahymena pyriformis. Hormonal imprinting alters the velocity. Comp. Biochem. Physiol. Comp. Physiol. 1994, 107, 375–379. [Google Scholar] [CrossRef]
- Christensen, S.T.; Quie, H.; Kemp, K.; Rasmussen, L. Insulin produces a biphasic response in Tetrahymena thermophila by stimulating cell survival and activating proliferation in two separate concentration intervals. Cell Biol. Int. 1996, 20, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Hellung-Larsen, P.; Leick, V.; Tommerup, N. Chemoattraction in Tetrahymena: On the role of chemokinesis. Biol. Bull. 1986, 170, 357–367. [Google Scholar] [CrossRef]
- Kőhidai, L.; Csaba, G. Chemotaxis and chemotactic selection induced with cytokines (IL-8, RANTES and TNF-alpha) in the unicellular Tetrahymena pyriformis. Cytokine 1998, 10, 481–486. [Google Scholar] [CrossRef]
- Kőhidai, L.; Kovács, K.; Csaba, G. Direct chemotactic effect of bradykinin and related peptides-significance of amino- and carboxyterminal character of oligopeptides in chemotaxis of tetrahymena pyriformis. Cell Biol. Int. 2002, 26, 55–62. [Google Scholar] [CrossRef]
- Csaba, G.; Kovács, K.; Kőhidai, L. Effect of oxytocin and its analogues on the chemotaxis of Tetrahymena: Evolutionary conclusions. Acta Protozool. 2000, 39, 345–347. [Google Scholar]
- Csaba, G.; Pállinger, É. Is there a hormonal network in Tetrahymena? A systematic investigation of hormonal effects on the hormone content. Cell Biochem. Funct. 2008, 26, 303–308. [Google Scholar] [CrossRef]
- Csaba, G.; Kovács, P.; Kőhidai, L. Tetrahymena cells distinguish insulin preparations according to either their amorphous and crystalline form or their bovine and porcine origin—Aspects of hormone-binding and chemotaxis in relation to imprinting. Microbios 1994, 80, 215–222. [Google Scholar]
- Kőhidai, L.; Csaba, G. Impact of length of exposure of different molecular mass on the establishment of imprinting in Tetrahymena. Acta Protozool. 1990, 29, 315–319. [Google Scholar]
- Csaba, G.; Kovacs, P.; Pallinger, E. How does the unicellular Tetrahymena utilise the hormones that it produces? Paying a visit to the realm of atto-and zeptomolar concentrations. Cell Tissue Res. 2007, 327, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Kovács, P.; Tóthfalusi, L.; Pállinger, E. Effects of extremely low concentrations of hormones on the insulin binding of Tetrahymena. Cell Biol. Int. 2006, 30, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Millar, R.P.; Lu, Z.L.; Pawson, A.J.; Flanagan, C.A.; Morgan, K.; Maudsley, S.R. Gonadotropin-releasing hormone receptors. Endocr. Rev. 2004, 25, 235–275. [Google Scholar] [CrossRef]
- Zandawala, M.; Tian, S.; Elphick, M.R. The evolution and nomenclature of GnRH-type and corazonin-type neuropeptide signaling systems. Gen. Comp. Endocrinol. 2017. [Google Scholar] [CrossRef]
- Vadakkadath Meethal, S.; Gallego, M.J.; Haasl, R.J.; Petras, S.J., 3rd; Sgro, J.Y.; Atwood, C.S. Identification of a gonadotropin-releasing hormone receptor orthologue in Caenorhabditis elegans. BMC Evol. Biol. 2006, 6, 103. [Google Scholar] [CrossRef]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. GnRH receptors and peptides: Skating backward. Gen. Comp. Endocrinol. 2014, 209, 118–134. [Google Scholar] [CrossRef]
- Powell, J.F.; Reska-Skinner, S.M.; Prakash, M.O.; Fischer, W.H.; Park, M.; Rivier, J.E.; Craig, A.G.; Mackie, G.O.; Sherwood, N.M. Two new forms of gonadotropin-releasing hormone in a protochordate and the evolutionary implications. Proc. Natl. Acad. Sci. USA 1996, 93, 10461–10464. [Google Scholar] [CrossRef]
- Csaba, G.; Kovács, P. Human chorionic gonadotropin (HCG)-like hormones (FSH, LH, TSH) in Tetrahymena. A confocal microscopic analysis. Acta. Protozool. 2000, 39, 191–198. [Google Scholar]
- Csaba, G.; Pállinger, E. Thyrotropin (TSH) regulates triiodothyronine (T3) production in the unicellular Tetrahymena. Acta Biol. Hung. 2011, 62, 228–234. [Google Scholar] [CrossRef]
- Lajkó, E.; Szabó, I.; Andódy, K.; Pungor, A.; Mező, G.; Kőhidai, L. Investigation on chemotactic drug targeting (chemotaxis and adhesion) inducer effect of GnRH-III derivatives in Tetrahymena and human leukemia cell line. J. Pept. Sci. 2013, 19, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I. Synthesis of GnRH-III Derivatives to Enhance its Antitumor Activity. Ph.D. Thesis, Eötvös L. University, Budapest, Hungary, 2009. [Google Scholar]
- Mező, G.; Czajlik, A.; Manea, M.; Jakab, A.; Farkas, V.; Majer, Z.; Vass, E.; Bodor, A.; Kapuvári, B.; Boldizsár, M.; et al. Structure, enzymatic stability and antitumor activity of sea lamprey GnRH-III and its dimer derivatives. Peptides 2007, 28, 806–820. [Google Scholar] [CrossRef] [PubMed]
- Cleverly, K.; Wu, T.J. Is the metalloendopeptidase EC 3.4.24.15 (EP24.15), the enzyme that cleaves luteinizing hormone-releasing hormone (LHRH), an activating enzyme? Reproduction (Camb. Engl.) 2010, 139, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Rivier, J.; Amoss, M.; Rivier, C.; Vale, W. Synthetic luteinizing hormone releasing factor. Short chain analogs. J. Med. Chem. 1974, 17, 230–233. [Google Scholar] [CrossRef]
- Rivier, J.; Vale, W.; Burgus, R.; Ling, N.; Amoss, M.; Blackwell, R.; Guillemin, R. Synthetic luteinizing hormone-releasing factor analogs. Series of short-chain amide LRF homologs converging to the amino terminus. J. Med. Chem. 1973, 16, 545–549. [Google Scholar] [CrossRef]
- Haviv, F.; Palabrica, C.A.; Bush, E.N.; Diaz, G.; Johnson, E.S.; Love, S.; Greer, J. Active reduced-size hexapeptide analogues of luteinizing hormone-releasing hormone. J. Med. Chem. 1989, 32, 2340–2344. [Google Scholar] [CrossRef]
- Larco, D.O.; Cho-Clark, M.; Mani, S.K.; Wu, T.J. The metabolite GnRH-(1-5) inhibits the migration of immortalized GnRH neurons. Endocrinology 2013, 154, 783–795. [Google Scholar] [CrossRef]
- Muto, Y.; Kőhidai, L. Image analysis for recording cell locomotion. Bull. Coll. Gifu Univ. 1999, 5, 87–93. [Google Scholar]
- Almagor, M.; Ron, A.; Bar-Tana, J. Chemotaxis in Tetrahymena thermophila. Cell Motil. 1981, 1, 261–268. [Google Scholar] [CrossRef]
- Leick, V.; Hellung-Larsen, P. Chemosensory behaviour of Tetrahymena. BioEssays: News Rev. Mol. Cell. Dev. Biol. 1992, 14, 61–66. [Google Scholar] [CrossRef]
- Kőhidai, L.; Csaba, G. Different and selective chemotactic responses of Tetrahymena pyriformis to two families of signal molecules: Lectins and peptide hormones. Acta Microbiol. Immunol. Hung. 1996, 43, 83–91. [Google Scholar] [PubMed]
- Lajkó, E.; Pállinger, E.; Csaba, G. Investigations on the triiodothyronine (T3)-specificity of thyrotropic (TSH) and gonadotropic (HCG) hormone in the unicellular Tetrahymena. Acta Microbiol. Immunol. Hung. 2011, 58, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.; Christensen, S.T.; Schousboe, P.; Wheatley, D.N. Cell survival and multiplication. The overriding need for signals: From unicellular to multicellular systems. FEMS Microbiol. Lett. 1996, 137, 123–128. [Google Scholar] [CrossRef]
- Hegyesi, H.; Csaba, G. Time- and concentration-dependence of the growth-promoting activity of insulin and histamine in Tetrahymena. Application of the MTT-method for the determination of cell proliferation in a protozoan model. Cell Biol. Int. 1997, 21, 289–293. [Google Scholar] [CrossRef]
- Karrer, K.M.; Peiffer, S.L.; DiTomas, M.E. Two distinct gene subfamilies within the family of cysteine protease genes. Proc. Natl. Acad. Sci. USA 1993, 90, 3063–3067. [Google Scholar] [CrossRef]
- Suzuki, K.M.; Hayashi, N.; Hosoya, N.; Takahashi, T.; Kosaka, T.; Hosoya, H. Secretion of tetrain, a Tetrahymena cysteine protease, as a mature enzyme and its identification as a member of the cathepsin L subfamily. Eur. J. Biochem. 1998, 254, 6–13. [Google Scholar] [CrossRef]
- Csaba, G.; Kőhidai, L. Interrelationship of hormone concentration, hormonal imprinting and receptor down-regulation in Tetrahymena. Acta Protozool. 1989, 28, 183–186. [Google Scholar]
- Iwata, H.; Kariya, K.; Wada, Y. Adrenergic mechanism in Tetrahymena. II. Effect of adrenaline on cell proliferation. Jpn. J. Pharmacol. 1973, 23, 681–688. [Google Scholar] [CrossRef][Green Version]
- Csaba, G. Biogenic amines at a low level of evolution: Production, functions and regulation in the unicellular Tetrahymena. Acta Microbiol. Immunol. Hung. 2015, 62, 93–108. [Google Scholar] [CrossRef]
- Csaba, G.; Németh, G. Effect of hormones and their precursors on protozoa-the selective responsiveness of Tetrahymena. Comp. Biochem. Physiol. 1980, 65B, 387–390. [Google Scholar] [CrossRef]
- Millar, R.P.; Pawson, A.J.; Morgan, K.; Rissman, E.F.; Lu, Z.L. Diversity of actions of GnRHs mediated by ligand-induced selective signaling. Front. Neuroendocrinol. 2008, 29, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G. Hormonal imprinting: Phylogeny, ontogeny, diseases and possible role in present-day human evolution. Cell Biochem. Funct. 2008, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Kovacs, P.; Pallinger, E. EDAC fixation increases the demonstrability of biogenic amines in the unicellular Tetrahymena: A flow cytometric and confocal microscopic comparative analysis. Cell Biol. Int. 2006, 30, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Pállinger, E. How applicable is the general adaptation syndrome to the unicellular Tetrahymena? Cell Biochem. Funct. 2009, 27, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Kovacs, P.; Pallinger, E. Effects of different fixatives on demonstrating epinephrine and ACTH hormones in Tetrahymena. Biotech. Histochem. Off. Publ. Biol. Stain Comm. 2009, 84, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Csaba, G.; Kovacs, P.; Pallinger, E. Increased hormone levels in Tetrahymena after long-lasting starvation. Cell Biol. Int. 2007, 31, 924–928. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Groups | Peptide Names | Structures |
|---|---|---|
| Natural hormones | GnRH-I | <EHWSYGLRPG-NH2 |
| GnRH-III | <EHWSHDWKPG-NH2 | |
| GnRH-III fragment | Ac-SHDWKPG-NH2 | |
| Symmetric GnRH-III dimers | [GnRH-III(C)]2 |  |
| [GnRH-III(CGFLG)]2 |  |
| GnRH Peptides | Intracellular Hormone Content Mean Fluorescence Intensity (Geometric Mean Channel Value) | ||||
|---|---|---|---|---|---|
| T3 | Histamine | Serotonin | Epinephrine | Endorphin | |
| Control | 59.00 ± 1.28 | 21.02 ± 1.31 | 24.04 ± 0.37 | 100.82 ± 3.56 | 47.89 ± 1.26 |
| GnRH-I | 58.96 ± 2.01 | 24.89 *** ± 0.51 | 24.87 ± 0.59 | 108.83 ± 2.11 | 55.78 ** ± 0.51 |
| GnRH-III | 63.23 * ± 0.83 | 21.68 ± 0.92 | 19.65 ** ± 0.97 | 104.04 ± 1.87 | 61.52 *** ± 1.31 |
| Ac-SHDWKPG-NH2 | 62.31 ± 1.94 | 30.27 *** ± 0.51 | 21.91 * ± 0.77 | 73.03 *** ± 1.34 | 43.85 ± 1.1 |
| [GnRH-III(C)]2 | 65.08 ± 5.02 | 28.59 ** ± 0.93 | 22.01 * ± 0.58 | 78.44 ** ± 2.14 | 54.60 * ± 2.51 |
| [GnRH-III(CGFLG)]2 | 54.56 * ± 0.55 | 27.93 * ± 1.75 | 23.01 ± 0.13 | 71.58 ** ± 2.11 | 47.45 ± 1.61 |
| GnRH Peptides | Chemotactic Effects [31] | Chemokinetic Effects | Changes in Intracellular Hormone Contents | Cell Proliferation Modulator Effects | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean velocity | Tortuosity | T3 rep ***/attr ** [12] | Histamine attr *** [12] | Serotonin rep *** [12] | Epinephrine neut a | Endorphin attr * [2] | 6 h | 24 h | ||
| GnRH-I | attr * | - | ++ | 0 | + | 0 | 0 | + | - | 0 |
| GnRH-III | rep * | - | ++ | + | 0 | - | 0 | ++ | 0 | + |
| Ac-SHDWKPG-NH2 | attr ** | -- | 0 | 0 | ++ | - | -- | - | + | 0 |
| [GnRH-III(C)]2 | attr ** | 0 | 0 | 0 | ++ | - | -- | + | ++ | 0 |
| [GnRH-III(CGFLG)]2 | rep ** | -- | +++ | - | ++ | 0 | -- | 0 | + | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lajkó, E.; Pállinger, É.; Kovács, Z.; Szabó, I.; Kőhidai, L. Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis. Int. J. Mol. Sci. 2019, 20, 5711. https://doi.org/10.3390/ijms20225711
Lajkó E, Pállinger É, Kovács Z, Szabó I, Kőhidai L. Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis. International Journal of Molecular Sciences. 2019; 20(22):5711. https://doi.org/10.3390/ijms20225711
Chicago/Turabian StyleLajkó, Eszter, Éva Pállinger, Zsombor Kovács, Ildikó Szabó, and László Kőhidai. 2019. "Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis" International Journal of Molecular Sciences 20, no. 22: 5711. https://doi.org/10.3390/ijms20225711
APA StyleLajkó, E., Pállinger, É., Kovács, Z., Szabó, I., & Kőhidai, L. (2019). Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis. International Journal of Molecular Sciences, 20(22), 5711. https://doi.org/10.3390/ijms20225711