Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism

 , , and
, , and 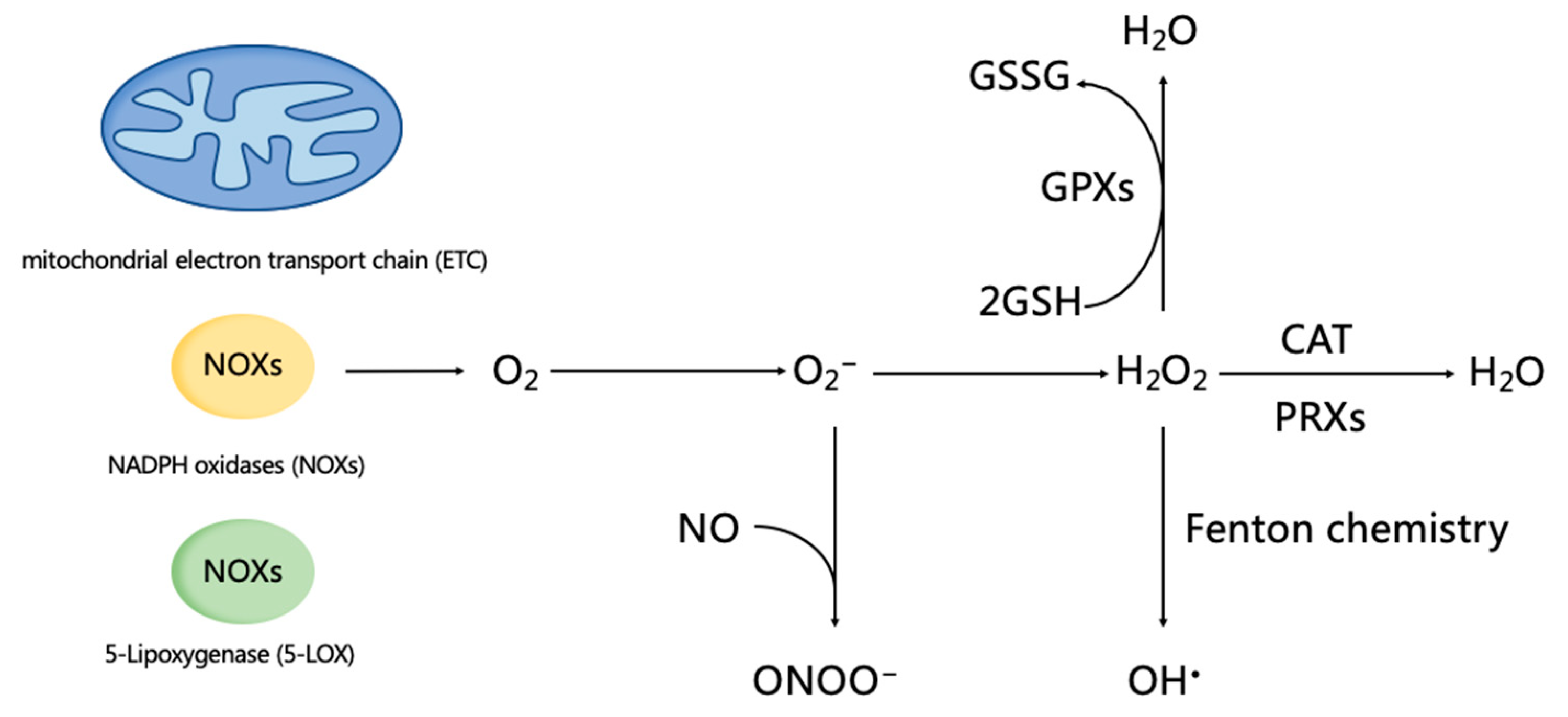{kind=link}
{kind=link}
Abstract
1. Introduction
2. Cancer
3. ROS and Cancer
3.1. ROS Homeostasis and Regulation
3.2. ROS and Carcinogenesis
3.3. ROS as a Cancer Therapy Agent
4. Tea Resists Carcinogenesis
4.1. Tea Polyphenols
4.2. Others
4.3. Tea Types and Anti-Cancer
5. Anti-Cancer Mechanisms of Tea through Regulating ROS Homeostasis
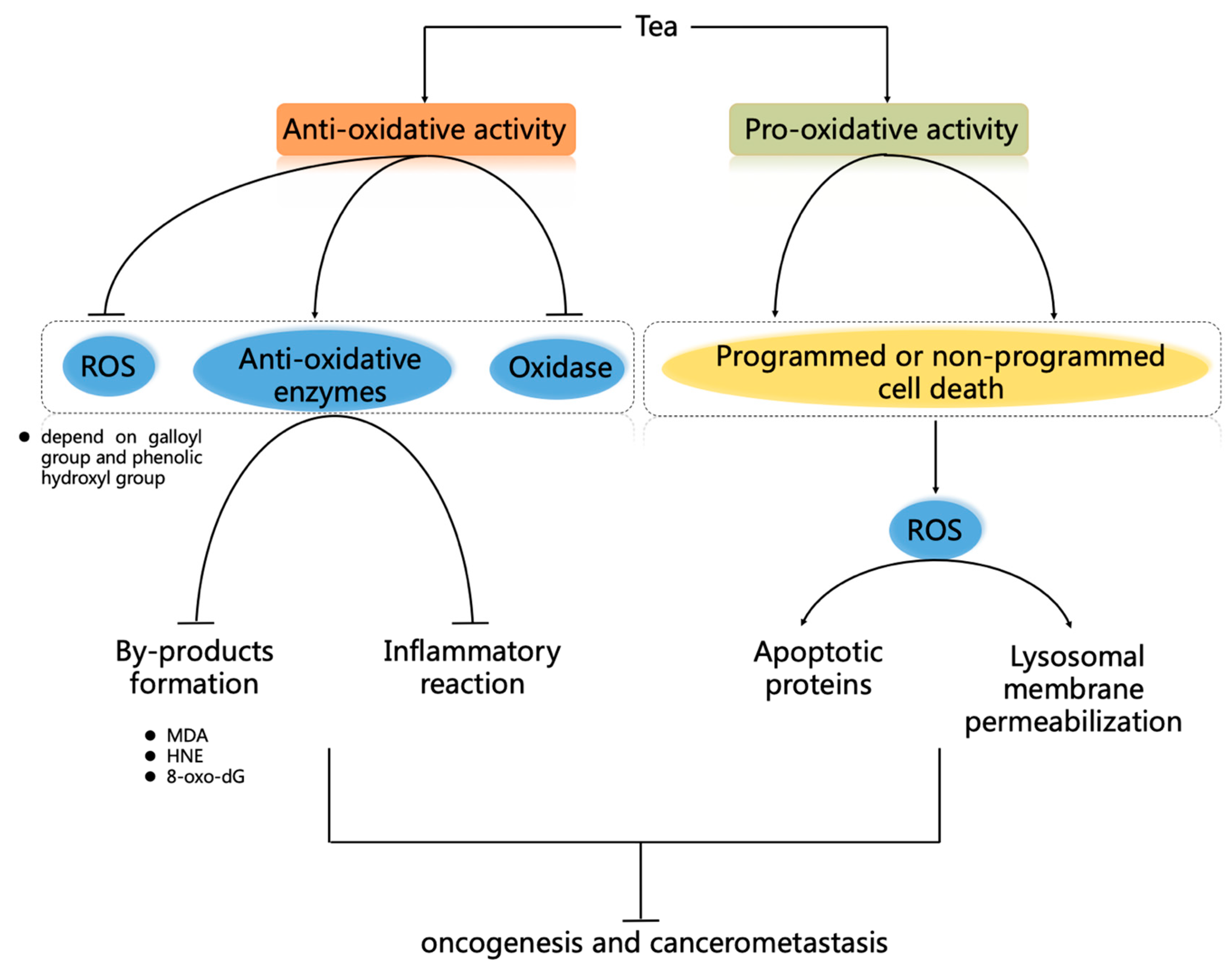
5.1. Anti-Oxidant Capacity of Tea
5.1.1. Tea as a Direct ROS Scavenger
5.1.2. Tea as an Indirect ROS Scavenger
5.2. Pro-Oxidative Activity of Tea
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, C.S.; Wang, Z.Y. Tea and cancer. J. Natl. Cancer Inst. 1993, 85, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Kuo, K.L.; Weng, M.S.; Chiang, C.T.; Tsai, Y.J.; Lin-Shiau, S.Y.; Lin, J.K. Comparative studies on the hypolipidemic and growth suppressive effects of oolong, black, pu-erh, and green tea leaves in rats. J. Agric. Food Chem. 2005, 53, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, Y.; Zhang, Y.; Wan, X.; Li, J.; Liu, K.; Wang, F.; Liu, K.; Liu, Q.; Yang, C.; et al. Anti-cancer activities of tea epigallocatechin-3-gallate in breast cancer patients under radiotherapy. Curr. Mol. Med. 2012, 12, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Wang, J.; Pan, S.; Lu, C. Tea consumption and the risk of ovarian cancer: A meta-analysis of epidemiological studies. Oncotarget 2017, 8, 37796–37806. [Google Scholar] [CrossRef]
- Fabiola, G.O.; Stephens, B.R.; Neilson, A.P.; Rodney, G.; Ferruzzi, M.G.; Bomser, J.A. Green and black tea inhibit cytokine-induced IL-8 production and secretion in AGS gastric cancer cells via inhibition of NF-κB activity. Planta Med. 2010, 76, 1659–1665. [Google Scholar]
- Tomtitchong, P.; Robinson, P.A.; Crabtree, J.E. The green tea catechin epigallocatechin-3-gallate inhibits H. pylori-Induced and TNF-α-induced IL-8 transcription and IL-8 secretion in gastric epithelial cells. Gastroenterology 2009, 136. [Google Scholar] [CrossRef]
- Fei, T.; Fei, J.; Huang, F.; Xie, T.; Yang, P. The anti-aging and anti-oxidation effects of tea water extract in Caenorhabditis elegans. Exp. Gerontol. 2017, 97, 89–96. [Google Scholar] [CrossRef]
- Borra, S.K.; Mahendra, J.; Gurumurthy, P.; Jayamathi; Iqbal, S.S.; Mahendra, L. Effect of curcumin against oxidation of biomolecules by hydroxyl radicals. J. Clin. Diagn. Res. 2014, 8, CC01–CC05. [Google Scholar]
- Zhou, Y.; Shu, F.; Liang, X.; Chang, H.; Shi, L.; Peng, X.; Zhu, J.; Mi, M. Ampelopsin induces cell growth inhibition and apoptosis in breast cancer cells through ROS generation and endoplasmic reticulum stress pathway. PLoS ONE 2014, 9, e89021. [Google Scholar]
- Hoekstra, H.J.; Wobbes, T.; Heineman, E.; Haryono, S.; Aryandono, T.; Balch, C.M. Fighting global disparities in cancer care: A surgical oncology view. Ann. Surg. Oncol. 2016, 23, 2131–2136. [Google Scholar] [CrossRef]
- Yang, C.S.; Lambert, J.D.; Ju, J.; Lu, G.; Sang, S. Tea and cancer prevention: Molecular mechanisms and human relevance. Toxicol. Appl. Pharmacol. 2007, 224, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Chen, J.F.; Canfell, K.; Feng, X.X.; Ma, J.F.; Zhang, Y.Z.; Zhao, F.H.; Li, R.; Ma, L.; Li, Z.F.; et al. Estimation of the costs of cervical cancer screening, diagnosis and treatment in rural Shanxi Province, China: A micro-costing study. BMC Health Serv. Res. 2012, 12, 123. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.L.; Currow, D.C. Cancer as a chronic disease. Collegian 2010, 17, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Duesberg, P.; Rasnick, D. Aneuploidy, the somatic mutation that makes cancer a species of its own. Cell Motil. Cytoskelet. 2000, 32, 81–107. [Google Scholar] [CrossRef]
- Martincorena, I.; Raine, K.M.; Gerstung, M.; Dawson, K.J.; Haase, K.; Van, L.P.; Davies, H.; Stratton, M.R.; Campbell, P.J. Universal Patterns of Selection in Cancer and Somatic Tissues. Cell 2017, 171, 1029–1041. [Google Scholar] [CrossRef]
- Borrello, M.G.; Degl’Innocenti, D.; Pierotti, M.A. Inflammation and cancer: The oncogene-driven connection. Cancer Lett. 2008, 267, 262–270. [Google Scholar] [CrossRef]
- Uchino, S.; Noguchi, M.; Ochiai, A.; Saito, T.; Kobayashi, M.; Hirohashi, S. p53 Mutation in gastric cancer: A genetic model for carcinogenesis is common to gastric and colorectal cancer. Int. J. Cancer 2010, 54, 759–764. [Google Scholar] [CrossRef]
- Spitschak, A.; Meier, C.; Kowtharapu, B.; Engelmann, D.; Pützer, B.M. MiR-182 promotes cancer invasion by linking RET oncogene activated NF-κB to loss of the HES1/Notch1 regulatory circuit. Mol. Cancer 2017, 16, 24. [Google Scholar] [CrossRef]
- Rahman, S.; Magnussen, M.; León, T.E.; Farah, N.; Mansour, M.R. Activation of the LMO2 oncogene through a somatically acquired neomorphic promoter in T-cell acute lymphoblastic leukemia. Blood 2017, 129, 3221–3226. [Google Scholar] [CrossRef]
- Jamaspishvili, T.; Berman, D.M.; Ross, A.E.; Scher, H.I.; Marzo, A.M.D.; Squire, J.A.; Lotan, T.L. Clinical implications of PTEN loss in prostate cancer. Nat. Rev. Urol. 2018, 15, 222–234. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Novo, E.; Parola, M. Redox mechanisms in hepatic chronic wound healing and fibrogenesis. Fibrogenesis Tissue Repair 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Reczek, C.R.; Chandel, N.S. The two faces of reactive oxygen species in cancer. Ann. Rev. Cancer Biol. 2017, 1, 79–98. [Google Scholar] [CrossRef]
- Peng, T.; Wong, N.K.; Chen, X.; Chan, Y.K.; Sun, Z.; Hu, J.J.; Shen, J.; Ei-Nezami, H.; Yang, D. Molecular imaging of peroxynitrite with HKGreen-4 in live cells and tissues. J. Am. Chem. Soc. 2014, 136, 11728–11734. [Google Scholar] [CrossRef]
- Ahmad, P.; Islam, B.U.; Allarakha, S.; Rabbani, G.; Dixit, K.; Moinuddin; Khan, R.H.; Siddiqui, S.A.; Ali, A. Preferential recognition of peroxynitrite-modified human serum albumin by circulating autoantibodies in cancer. Int. J. Biol. Macromol. 2015, 72, 875–882. [Google Scholar] [CrossRef]
- Mailloux, R.J.; Mcbride, S.L.; Harper, M.E. Unearthing the secrets of mitochondrial ROS and glutathione in bioenergetics. Trends Biochem. Sci. 2013, 38, 592–602. [Google Scholar] [CrossRef]
- Wu, J.; Sun, B.; Luo, X.; Zhao, M.; Huang, M. Cytoprotective effects of a tripeptide from Chinese Baijiu against AAPH-induced oxidative stress in HepG2 cells: Via Nrf2 signaling. RSC Adv. 2018, 8, 10898–10906. [Google Scholar] [CrossRef]
- Vurusaner, B.; Poli, G.; Basaga, H. Tumor suppressor genes and ROS: Complex networks of interactions. Free Radic. Biol. Med. 2012, 52, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Lara, G.; Marcello, P.; Milena, N.; Sara, D.B.; Erika, R.; Linda, B.; Andrea, C. Interfering with ROS metabolism in cancer cells: The potential role of quercetin. Cancers 2010, 2, 1288–1311. [Google Scholar]
- Mao, X.; Gu, C.; Chen, D.; Yu, B.; He, J. Oxidative stress-induced diseases and tea polyphenols. Oncotarget 2017, 8, 81649–81661. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.Y.; Hsu, T.; Santella, R.M. Immunohistochemical detection of malondialdehyde–DNA adducts in human oral mucosa cells. Carcinogenesis 2002, 23, 207–211. [Google Scholar] [CrossRef]
- Zhong, H.; Yin, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biol. 2015, 4, 193–199. [Google Scholar] [CrossRef]
- Bolin, C.; Stedeford, T.; Cardozopelaez, F. Single extraction protocol for the analysis of 8-hydroxy-2’-deoxyguanosine (oxo8dG) and the associated activity of 8-oxoguanine DNA glycosylase. J. Neurosci. Methods 2004, 136, 69–76. [Google Scholar] [CrossRef]
- Storz, P. Reactive oxygen species in tumor progression. Front. Biosci. 2005, 10, 1881–1896. [Google Scholar] [CrossRef]
- Riemann, A.; Schneider, B.; Ihling, A.; Nowak, M.; Sauvant, C.; Thews, O.; Gekle, M. Acidic environment leads to ROS-induced MAPK signaling in cancer cells. PLoS ONE 2011, 6, e22445. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Chetram, M.A.; Don-Salu-Hewage, A.S.; Hinton, C.V. ROS enhances CXCR4-mediated functions through inactivation of PTEN in prostate cancer cells. Biochem. Biophys. Res. Commun. 2011, 410, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Athanasios, V.; Thomais, V.; Konstantinos, F.; Spyridon, L. Pulmonary oxidative stress, inflammation and cancer: Respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms. Int. J. Environ. Res. Public Health 2013, 10, 3886–3907. [Google Scholar]
- Basu, S.; Rossary, A.; Vasson, M.P. Role of inflammation and eicosanoids in breast cancer. Lipid Technol. 2016, 28, 60–64. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Shivappa, N.; Steck, S.E.; Canzian, F.; Landi, S.; Alonso, M.H.; Hébert, J.R.; Moreno, V. Dietary inflammatory index and inflammatory gene interactions in relation to colorectal cancer risk in the Bellvitge colorectal cancer case–control study. Genes Nutr. 2015, 10, 447. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A. Molecular pathways linking inflammation and cancer. Curr. Mol. Med. 2010, 10, 369–373. [Google Scholar] [CrossRef]
- Hardbower, D.M.; De, S.T.; Chaturvedi, R.; Wilson, K.T. Chronic inflammation and oxidative stress: The smoking gun for Helicobacter pylori-induced gastric cancer? Gut Microbes 2013, 4, 475–481. [Google Scholar] [CrossRef]
- Rahat, M.A.; Shakya, J. Parallel aspects of the microenvironment in cancer and autoimmune disease. Mediat. Inflamm. 2016, 2016, 1–17. [Google Scholar] [CrossRef]
- Zhang, R.; Yin, X.; Shi, H.; Wu, J.; Shakya, P.; Liu, D.; Zhang, J. Adiponectin modulates DCA-induced inflammation via the ROS/NF-κB signaling pathway in Esophageal Adenocarcinoma Cells. Dig. Dis. Sci. 2014, 59, 89–97. [Google Scholar] [CrossRef]
- Cao, Z.; Shang, B.; Zhang, G.; Miele, L.; Sarkar, F.H.; Wang, Z.; Zhou, Q. Tumor cell-mediated neovascularization and lymphangiogenesis contrive tumor progression and cancer metastasis. Biochim. Biophys. Acta (BBA) Rev. Cancer 2013, 1836, 273–286. [Google Scholar] [CrossRef]
- Bae, J.; Park, D.; Lee, Y.S.; Jeoung, D. Interleukin-2 promotes angiogenesis by activation of Akt and increase of ROS. J. Microbiol. Biotechnol. 2008, 18, 377–382. [Google Scholar]
- Katsabeki-Katsafli, A.; Kerenidi, T.; Kostikas, K.; Dalaveris, E.; Kiropoulos, T.S.; Gogou, E.; Papaioannou, A.I.; Gourgoulianis, K.I. Serum vascular endothelial growth factor is related to systemic oxidative stress in patients with lung cancer. Lung Cancer 2008, 60, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Foley, K.; Bertin, J.; Chan, K.; Hutchings, A.; Inoue, T.; Kirshner, J.; Korbut, T.; Li, L.; Mihalek, R.; Rao, P.; et al. The oxidative stress inducer STA-4783 enhances the in vivo efficacy of multiple anti-cancer therapies in mouse tumor models. Mol. Cancer Ther. 2007, 3430–3431. [Google Scholar]
- Manohar, M.; Fatima, I.; Saxena, R.; Chandra, V.; Sankhwar, P.L.; Dwivedi, A. (−)-Epigallocatechin-3-gallate induces apoptosis in human endometrial adenocarcinoma cells via ROS generation and p38 MAP kinase activation. J. Nutr. Biochem. 2013, 24, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kim, S.B.; Luitel, K.; Shay, J.W. Cholesterol depletion by TASIN-1 induces apoptotic cell death through the ER Stress/ROS/JNK signaling in colon cancer cells. Mol. Cancer Ther. 2018, 17, 945–951. [Google Scholar] [CrossRef]
- Lei, J.; Shestov, A.A.; Swain, P.; Yang, C.; Parker, S.J.; Wang, Q.A.; Terada, L.S.; Adams, N.D.; Mccabe, M.T.; Pietrak, B. Reductive carboxylation supports redox homeostasis during anchorage-independent growth. Nature 2016, 532, 255–258. [Google Scholar]
- Chandel, N.S.; Tuveson, D.A. The promise and perils of antioxidants for cancer patients. N. Engl. J. Med. 2014, 371, 177–178. [Google Scholar] [CrossRef]
- Sang, S. Tea: Chemistry and Processing. Encycl. Food Health 2016. [Google Scholar] [CrossRef]
- Zhu, G.; Hua, J.; Wang, Z.; She, F.; Chen, Y. Tea consumption and risk of gallbladder cancer: A meta-analysis of epidemiological studies. Mol. Clin. Oncol. 2015, 3, 613–618. [Google Scholar] [CrossRef]
- Riza, E.; Linos, A.; Petralias, A.; De Martinis, L.; Duntas, L.; Linos, D. The effect of Greek herbal tea consumption on thyroid cancer: A case-control study. Eur. J. Public Health 2015, 25, 1001–1005. [Google Scholar] [CrossRef]
- Sing, M.F.; Yang, W.; Gao, S.; Gao, J.; Xiang, Y. Epidemiological studies of the association between tea drinking and primary liver cancer: A meta-analysis. Eur. J. Cancer Prev. 2011, 20, 157–165. [Google Scholar] [CrossRef]
- Sang, L.X.; Chang, B.; Li, X.H.; Jiang, M. Green tea consumption and risk of esophageal cancer: A meta-analysis of published epidemiological studies. Nutr. Cancer 2013, 65, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Hughes, M.C.B.; Arovah, N.I.; Pols, J.C.V.D.; Green, A.L.C. Black tea consumption and risk of skin cancer: An 11-year prospective study. Nutr. Cancer 2015, 67, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Braem, M.G.M.; Onland-Moret, N.C.; Schouten, L.J.; Tjønneland, A.; Hansen, L.; Dahm, C.C.; Overvad, K.; Lukanova, A.; Dossus, L.; Floegel, A.; et al. Coffee and tea consumption and the risk of ovarian cancer: A prospective cohort study and updated meta-analysis. Am. J. Clin. Nutr 2012, 95, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y. Protective effects of tea polyphenols for weaned pigs challenged with oxidative stress. Ph.D. Thesis, Sichuan Agricultural University, Ya’an, China, 2011. [Google Scholar]
- Li, Y.Y.; Duan, X.D.; Zhao, J.; Yu, B.; Mao, X.B.; Mao, Q.; Chen, H.; Chen, D.W. Effects of tea polyphenolon growth performance and immune function of weaned piglets chalenged with oxidative stress. Chin. J. Anim. Sci. 2011, 47, 53–57. [Google Scholar]
- Chao, Y.M.; Chen, D.W.; Yu, B.; He, J.; Yu, J.; Mao, X.B.; Luo, Y.H.; Huang, Z.Q.; Luo, J.Q.; Wang, S.H. Effects of tea polyphenol on growth performance, antioxidant capacity, carcass performance and meat quality of finishing pigs. Chin. J. Anim. Nutr. 2016, 28, 3996–4005. [Google Scholar]
- Zhou, F.; Shen, T.; Duan, T.; Xu, Y.Y.; Khor, S.C.; Li, J.; Ge, J.; Zheng, Y.F.; Hsu, S.; De, S.J. Antioxidant effects of lipophilic tea polyphenols on diethylnitrosamine/phenobarbital-induced hepatocarcinogenesis in rats. Vivo 2014, 28, 495–503. [Google Scholar]
- Li, H.; Jiang, N.; Liu, Q.; Gao, A.; Zhou, X.; Liang, B.; Li, R.; Li, Z.; Zhu, H. Topical treatment of green tea polyphenols emulsified in carboxymethyl cellulose protects against acute ultraviolet light B-induced photodamage in hairless mice. Photochem. Photobiol. Sci. 2016, 15, 1264–1271. [Google Scholar] [CrossRef]
- Park, J.S.; Khoi, P.N.; Joo, Y.-E.; Lee, Y.H. EGCG inhibits recepteur d’origine nantais expression by suppressing Egr-1 in gastric cancer cells. Int. J. Oncol. 2013, 42, 1120–1126. [Google Scholar] [CrossRef]
- Morris, J.; Moseley, V.R.; Cabang, A.B.; Coleman, K.; Wei, W.; Garrettmayer, E.; Wargovich, M.J. Reduction in promotor methylation utilizing EGCG (Epigallocatechin-3-gallate) restores RXRα expression in human colon cancer cells. Oncotarget 2016, 7, 35313–35326. [Google Scholar] [CrossRef]
- Jiang, P.; Wu, X.; Wang, X.; Huang, W.; Feng, Q. NEAT1 upregulates EGCG-induced CTR1 to enhance cisplatin sensitivity in lung cancer cells. Oncotarget 2016, 7, 43337–43351. [Google Scholar] [CrossRef]
- Sang, S.M.; Lambert, J.D.; Ho, C.T.; Yang, C.S. The chemistry and biotransformation of tea constituents. Pharmacol. Res. 2011, 64, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.S.; Sang, S.; Cheng, K.H.; Ho, C.T.; Wang, Y.J.; Pan, M.H. Peracetylated (−)-epigallocatechin-3-gallate (AcEGCG) potently prevents skin carcinogenesis by suppressing the PKD1-dependent signaling pathway in CD34+ skin stem cells and skin tumors. Carcinogenesis 2013, 34, 1315–1322. [Google Scholar] [CrossRef]
- Ling, T.; Forester, S.C.; Lambert, J.D. The role of reactive oxygen species in (−)-epigallocatechin-3-gallate (EGCG)-induced cell growth inhibition and apoptosis in oral cancer cells. Cancer Res. 2012, 72, 5436. [Google Scholar]
- Lecumberri, E.; Dupertuis, Y.M.; Miralbell, R.; Pichard, C. Green tea polyphenol epigallocatechin-3-gallate (EGCG) as adjuvant in cancer therapy. Clin. Nutr. 2013, 32, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Park, J.; Oh, B.; Min, H.J.; Jeong, T.S.; Lee, J.H.; Park, S.B.; Lee, D.S. (−)-epigallocatechin-3-gallate (EGCG), green tea component, antagonize the anti-myeloma activity of proteasome inhibitor PS-341 by direct chemical interaction. Blood 2007, 110, 4850. [Google Scholar] [CrossRef]
- Du, L.L.; Fu, Q.Y.; Xiang, L.P.; Zheng, X.Q.; Lu, J.L.; Ye, J.H.; Li, Q.S.; Polito, C.A.; Liang, Y.R. Tea Polysaccharides and Their Bioactivities. Molecules 2016, 21, 1449. [Google Scholar] [CrossRef]
- Yang, J.J.; Chen, B.; Gu, Y. Pharmacological evaluation of tea polysaccharides with antioxidant activity in gastric cancer mice. Carbohydr. Polym. 2012, 90, 943–947. [Google Scholar] [CrossRef]
- He, N.; Shi, X.; Zhao, Y.; Tian, L.; Wang, D.; Yang, X. Inhibitory effects and molecular mechanisms of selenium-containing tea polysaccharides on human breast cancer MCF-7 cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef]
- Wei, N.; Zhu, Q.; Chen, J.; Tong, L.I.; Li, Y.F.; Huang, Y.; Xiao, M.A.; Wang, X.; Sheng, J. Tea polysaccharide increased Doxorubicin inhibition of lung cancer A549 cells. J. Tea Sci. 2016, 36, 477–483. [Google Scholar]
- Li, L.; Wang, X.; Xiong, Y.; Ren, W.; Huang, M.; Zhao, G.; Ding, Y.; Wu, X.; Su, D. L-theanine: A promising substance in tumor research. J. Food Agric. Environ. 2013, 11, 25–27. [Google Scholar]
- Liu, J.; Sun, Y.; Zhang, H.; Ji, D.; Fei, W.; Tian, H.; Liu, K.; Ying, Z.; Wu, B.; Zhang, G. Theanine from tea and its semi-synthetic derivative TBrC suppress human cervical cancer growth and migration by inhibiting EGFR/Met-Akt/NF-κB signaling. Eur. J. Pharmacol. 2016, 791, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, T. Neuroprotective effects of the green tea components theanine and catechins. Biol. Pharm. Bull. 2002, 25, 1513–1518. [Google Scholar] [CrossRef] [PubMed]
- Nathan, P.J.; Lu, K.; Gray, M.; Oliver, C. The neuropharmacology of L-theanine(N-ethyl-L-glutamine): A possible neuroprotective and cognitive enhancing agent. J. Herb. Pharmacother. 2006, 6, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kang, J.; Yao, X.; Xin, Y.; Wang, Q.; Ye, Y.; Luo, L.; Yin, Z. The component of green tea, L-theanine protects human hepatic L02 cells from hydrogen peroxide-induced apoptosis. Eur. Food Res. Technol. 2011, 233, 427–435. [Google Scholar] [CrossRef]
- Sadzuka, Y.; Sugiyama, T.; Miyagishima, A.; Nozawa, Y.; Hirota, S. The effects of theanine, as a novel biochemical modulator, on the antitumor activity of adriamycin. Cancer Lett. 1996, 105, 203–209. [Google Scholar] [CrossRef]
- Tomita, M.; Irwin, K.I.; Xie, Z.J.; Santoro, T.J. Tea pigments inhibit the production of type 1 (TH1) and type 2 (TH2) helper T cell cytokines in CD4+ T cells. Phytother. Res. 2002, 16, 36–42. [Google Scholar] [CrossRef]
- Han, C.; Gong, Y. Experimental studies on the cancer chemoprevention of tea pigments. J. Hyg. Res. 1999, 28, 343. [Google Scholar]
- Gong, Y.; Han, C.; Chen, J. Effect of tea polyphenols and tea pigments on the inhibition of precancerous liver lesions in rats. Nutr. Cancer 2000, 38, 81–86. [Google Scholar] [CrossRef]
- Jia, X.D.; Han, C. Chemoprevention of tea on colorectal cancer induced by dimethylhydrazine in Wistar rats. World J. Gastroenterol. 2000, 6, 699–703. [Google Scholar] [CrossRef]
- Graham, D.M. Caffeine—Its Identity, Dietary Sources, Intake and Biological Effects. Nutr. Rev. 2010, 36, 97–102. [Google Scholar] [CrossRef]
- Okano, J.; Nagahara, T.; Matsumoto, K.; Murawaki, Y. Caffeine inhibits the proliferation of liver cancer cells and activates the MEK/ERK/EGFR signalling pathway. Basic Clin. Pharmacol. Toxicol. 2010, 102, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Petrek, J.A.; Sandberg, W.A.; Cole, M.N.; Silberman, M.S.; Collins, D.C. The inhibitory effect of caffeine on hormone-induced rat breast cancer. Cancer 1985, 56, 1977–1981. [Google Scholar] [CrossRef]
- Minton, J.P.; Abou-Issa, H.; Foecking, M.K.; Sriram, M.G. Caffeine and unsaturated fat diet significantly promotes DMBA-induced breast cancer in rats. Cancer 1983, 51, 1249–1253. [Google Scholar] [CrossRef]
- Lee, A.H.; Su, D.; Pasalich, M.; Binns, C.W. Tea consumption reduces ovarian cancer risk. Cancer Epidemiol. 2013, 37, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Wu, Y.; Zhou, B.; Wang, B.; Yu, R. Green tea, black tea consumption and risk of lung cancer: A meta-analysis. Lung Cancer 2009, 65, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, B.; Huang, T.; Yu, Y.; Yang, J.; Li, D. Green Tea and Black Tea Consumption and Prostate Cancer Risk: An Exploratory Meta-Analysis of Observational Studies. Nutr. Cancer 2011, 63, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.H.; Jin, J.; Liang, Y.R. Antioxidant Properties of Green Tea, Oolong Tea, Black Tea and Pu-erh Tea. J. Korea Tea 2015, 1, 221–228. [Google Scholar]
- Record, I.R.; Dreosti, I.E. Protection by black tea and green tea against UVB and UVA + B induced skin cancer in hairless mice. Mutat. Res. 1998, 422, 191–199. [Google Scholar] [CrossRef]
- Zhang, G.; Miura, Y.; Yagasaki, K. Effects of green, oolong and black teas and related components on the proliferation and invasion of hepatoma cells in culture. Cytotechnology 1999, 31, 37–44. [Google Scholar] [CrossRef]
- Dai, D.Z.; Chen, S.H. Tea Polyphenols and Quercetin Preventing the Heart, Brain and Liver from the Injury by Free Radicals in Comparison with Ascorbic Acid. Chin. J. Nat. Med. 2004, 2, 223–231. [Google Scholar]
- Ma, L.; Liu, Z.; Zhou, B.; Yang, L.; Liu, Z. Inhibition of free radical induced oxidative hemolysis of red blood cells by green tea polyphenols. Chin. Sci. Bull. 2000, 45, 2052–2056. [Google Scholar] [CrossRef]
- Hussain, S. Comparative efficacy of epigallocatechin-3-gallate against H2O2-induced ROS in cervical cancer biopsies and HeLa cell lines. Contemp. Oncol. 2017, 21, 209–212. [Google Scholar]
- Henning, S.M.; Wang, P.; Said, J.; Magyar, C.; Castor, B.; Doan, N.; Tosity, C.; Moro, A.; Gao, K.; Li, L.; et al. Polyphenols in brewed green tea inhibit prostate tumor xenograft growth by localizing to the tumor and decreasing oxidative stress and angiogenesis. J. Nutr. Biochem. 2012, 23, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Chen, S.J.; Zhang, C. Capacity of hesperidin, limonene and tea polyphenols on sequestering free radical. Oral Sci. Res. 2015, 31, 11–14. [Google Scholar]
- Kondo, K.; Kurihara, M.; Miyata, N. Scavenging mechanisms of (−)-epigallocatechin gallate and (−)-epicatechin gallate on peroxyl radicals and formation of superoxide during the inhibitory action. Free Radical Biol. Med. 1999, 27, 855–863. [Google Scholar] [CrossRef]
- Kuhn, D.; Lam, W.H.; Kazi, A.; Daniel, K.G.; Song, S.; Chow, L.M.; Chan, T.H.; Dou, Q.P. Synthetic peracetate tea polyphenols as potent proteasome inhibitors and apoptosis inducers in human cancer cells. Front. Biosci. 2005, 10, 1010–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.J.; Gao, Y.X.; Yuan, Y.J.; Yang, H.J.; Zhang, J. Study on extraction technology and antioxidant activity of theabrownine from oolong tea. Chin. J. Exp. Tradit. Med. Formulae 2011, 17, 36–40. [Google Scholar]
- Zhao, Z.Y.; Huangfu, L.T.; Dong, L.L.; Liu, S.L. Functional groups and antioxidant activities of polysaccharides from five categories of tea. Ind. Crops Prod. 2014, 58, 31–35. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Huo, J.; Zhao, T.; Ren, J.; Wei, X. Effect of different drying methods on chemical composition and bioactivity of tea polysaccharides. Int. J. Biol. Macromol. 2013, 62, 714–719. [Google Scholar] [CrossRef]
- Xiao, J.; Huo, J.; Jiang, H.; Yang, F. Chemical compositions and bioactivities of crude polysaccharides from tea leaves beyond their useful date. Int. J. Biol. Macromol. 2011, 49, 1143–1151. [Google Scholar] [CrossRef]
- Nie, S.P.; Xie, M.Y.; Cao, S.W. The antioxidative activity and anti-proliferation activity of purified tea polysaccharide against two colon cancer cell lines. Acta Nutr. Sin. 2007, 29, 46–50. [Google Scholar]
- Liao, Y.; Zou, X.; Wang, C.; Xin, Z. Insect tea extract attenuates CCl4-induced hepatic damage through its antioxidant capacities in ICR mice. Food Sci. Biotechnol. 2016, 25, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Paquay, J.B.; Haenen, G.R.; Stender, G.; Wiseman, S.A.; Tijburg, L.B.; Bast, A. Protection against nitric oxide toxicity by tea. J. Agric. Food Chem. 2000, 48, 5768–5772. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Chiou, Y.S.; Pan, M.H.; Ho, C.T.; Shahidi, F. Protective effects of epigallocatechin gallate (EGCG) derivatives on azoxymethane-induced colonic carcinogenesis in mice. J. Funct. Foods 2012, 4, 323–330. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Bishayee, A. Nrf2-mediated redox signaling in arsenic carcinogenesis: A review. Arch. Toxicol. 2013, 87, 383–396. [Google Scholar] [CrossRef]
- Qi, G.; Mi, Y.; Rong, F.; Li, R.; Wang, Y.; Li, X.; Huang, S.; Liu, X. Tea polyphenols ameliorate hydrogen peroxide- and constant darkness-triggered oxidative stress via modulating the Keap1/Nrf2 transcriptional signaling pathway in HepG2 cells and mice liver. RSC Adv. 2017, 7, 32198–32208. [Google Scholar] [CrossRef]
- Patel, R.; Maru, G. Polymeric black tea polyphenols induce phase II enzymes via Nrf2 in mouse liver and lungs. Free Radic. Biol. Med. 2008, 44, 1897–1911. [Google Scholar] [CrossRef]
- Gao, Z.; Han, Y.; Hu, Y.; Wu, X.; Wang, Y.; Zhang, X.; Fu, J.; Zou, X.; Zhang, J.; Chen, X. Targeting HO-1 by epigallocatechin-3-gallate reduces contrast-induced renal injury via anti-oxidative stress and anti-inflammation pathways. PLoS ONE 2015, 11, e0149032. [Google Scholar] [CrossRef]
- Furfaro, A.L.; Traverso, N.; Domenicotti, C.; Piras, S.; Moretta, L.; Marinari, U.M.; Pronzato, M.A.; Nitti, M. The Nrf2/HO-1 axis in cancer cell growth and chemoresistance. Oxid. Med. Cell. Longev. 2015, 2016, 1958174. [Google Scholar] [CrossRef]
- Li, X.; Zou, Z.; Tang, J.; Zheng, Y.; Liu, Y.; Luo, Y.; Liu, Q.; Wang, Y. NOS1 upregulates ABCG2 expression contributing to DDP chemoresistance in ovarian cancer cells. Biol. Pharm. Bull. 2018, 41, 1237–1242. [Google Scholar] [CrossRef]
- Yu, C.; Jiao, Y.; Xue, J.; Zhang, Q.; Yang, H.; Xing, L.; Chen, G.; Wu, J.; Zhang, S.; Zhu, W.; et al. Metformin sensitizes non-small cell lung cancer cells to an epigallocatechin-3-gallate (EGCG) treatment by suppressing the Nrf2/HO-1 signaling pathway. Int. J. Biol. Sci. 2017, 13, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, S.; Saito, K.; Miyoshi, N.; Ohishi, T.; Oishi, Y.; Miyoshi, M.; Nakamura, Y. Anti-cancer effects of green tea by Either anti- or pro- oxidative mechanisms. Asian Pac. J. Cancer Prev. 2016, 17, 1649–1654. [Google Scholar] [PubMed]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS crosstalk in inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [PubMed]
- Li, J.; Sapper, T.N.; Mah, E.; Moller, M.V.; Kim, J.B.; Chitchumroonchokchai, C.; Mcdonald, J.D.; Bruno, R.S. Green tea extract treatment reduces NFκB activation in mice with diet-induced nonalcoholic steatohepatitis by lowering TNFR1 and TLR4 expression and ligand availability. J. Nutr. Biochem. 2017, 41, 34–41. [Google Scholar]
- Santamarina, A.B.; Carvalho-Silva, M.; Gomes, L.M.; Okuda, M.H.; Santana, A.A.; Streck, E.L.; Seelaender, M.; Nascimento, C.M.O.D.; Ribeiro, E.B.; Lira, F.S. Decaffeinated green tea extract rich in epigallocatechin-3-gallate prevents fatty liver disease by increased activities of mitochondrial respiratory chain complexes in diet-induced obesity mice. J. Nutr. Biochem. 2015, 26, 1348–1356. [Google Scholar]
- Mitchell, T.S.; Moots, R.J.; Wright, H.L. Janus kinase inhibitors prevent migration of rheumatoid arthritis neutrophils towards interleukin-8, but do not inhibit priming of the respiratory burst or reactive oxygen species production. Clin. Exp. Immunol. 2017, 189, 250–258. [Google Scholar] [CrossRef]
- Pal, D.; Sur, S.; Mandal, S.; Das, S.; Pandaa, C.K. Regular black tea habit could reduce tobacco associated ROS generation and DNA damage in oral mucosa of normal population. Food Chem. Toxicol. 2012, 50, 2996–3003. [Google Scholar] [CrossRef]
- Li, J.; Sapper, T.N.; Mah, E.; Rudraiah, S.; Schill, K.E.; Chitchumroonchokchai, C.; Moller, M.V.; Mcdonald, J.D.; Rohrer, P.R.; Manautou, J.E.; et al. Green tea extract provides extensive Nrf2-independent protection against lipid accumulation and NF-κB pro- inflammatory responses during nonalcoholic steatohepatitis in mice fed a high-fat diet. Mol. Nutr. Food Res. 2016, 60, 858–870. [Google Scholar]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NF-κB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar]
- Das, D.N.; Panda, P.K.; Naik, P.P.; Mukhopadhyay, S.; Sinha, N.; Bhutia, S.K. Phytotherapeutic approach: A new hope for polycyclic aromatic hydrocarbons induced cellular disorders, autophagic and apoptotic cell death. Toxicol. Mech. Methods 2017, 27, 1–17. [Google Scholar]
- Wang, Z.Y.; Khan, W.A.; Bickers, D.R.; Mukhtar, H. Protection against polycyclic aromatic hydrocarbon-induced skin tumor initiation in mice by green tea polyphenols. Carcinogenesis 1989, 10, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Wilk, A.; Rski, P.W.; Lassak, A.; Vashistha, H.; Lirette, D.; Tate, D.; Zea, A.H.; Koochekpour, S.; Rodriguez, P.; Meggs, L.G.; et al. Polycyclic aromatic hydrocarbons-induced ROS accumulation enhances mutagenic potential of T-antigen from human polyomavirus JC. J. Cell Physiol. 2013, 228, 2127–2138. [Google Scholar] [CrossRef] [PubMed]
- Libalova, H.; Milcova, A.; Cervena, T.; Vrbova, K.; Rossnerova, A.; Novakova, Z.; Topinka, J.; Rossner, P., Jr. Kinetics of ROS generation induced by polycyclic aromatic hydrocarbons and organic extracts from ambient air particulate matter in model human lung cell lines. Mutat. Res. Toxicol. Environ. Mutagen. 2018, 827, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, I.; Nishiumi, S.; Mukai, R.; Yoshida, K.; Ashida, H. Catechins in tea suppress the activity of cytochrome P450 1A1 through the aryl hydrocarbon receptor activation pathway in rat livers. Int. J. Food Sci. Nutr. 2015, 66, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Lambert, J.D.; Hou, Z.; Ju, J.; Lu, G.; Hao, X. Molecular targets for the cancer preventive activity of tea polyphenols. Mol. Carcinog. 2010, 45, 431–435. [Google Scholar] [CrossRef]
- Hider, R.C.; Liu, Z.D.; Khodr, H.H. Metal chelation of polyphenols. Methods Enzym. 2001, 335, 190–203. [Google Scholar]
- Zhou, Y.; Que, K.-T.; Zhang, Z.; Zhao, P.X.; You, Y.; Gong, J.-P.; Liu, Z.-J. Iron ion polarizes macrophages to M1 subtype through ROS-acetyl-P53. J. Third Mil. Med. Univ. 2018, 7, 4012–4022. [Google Scholar]
- Gumiennakontecka, E.; Pyrkoszbulska, M.; Szebesczyk, A.; Ostrowska, M. Iron chelating strategies in systemic metal overload, neurodegeneration and cancer. Curr. Med. Chem. 2014, 21, 3741–3767. [Google Scholar] [CrossRef]
- Yang, W.-H.; Fong, Y.-C.; Lee, C.-Y.; Jin, T.-R.; Tzen, J.T.; Li, T.-M.; Tang, C.-H. Epigallocatechin-3-gallate induces cell apoptosis of human chondrosarcoma cells through apoptosis signal-regulating kinase 1 pathway. J. Cell. Biochem. 2011, 112, 1601–1611. [Google Scholar] [CrossRef]
- Alshatwi, A.A.; Periasamy, V.S.; Athinarayanan, J.; Elango, R. Synergistic anticancer activity of dietary tea polyphenols and bleomycin hydrochloride in human cervical cancer cell: Caspase-dependent and independent apoptotic pathways. Chem. Biol. Interact. 2016, 247, 1–10. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Chen, C.Y.; Chiou, Y.H.; Shyu, H.W.; Lin, K.H.; Chou, M.C.; Huang, M.H.; Wang, Y.F. Epigallocatechin-3-gallate suppresses human herpesvirus 8 replication and induces ROS leading to apoptosis and autophagy in primary effusion lymphoma cells. Int. J. Mol. Sci. 2018, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Satoh, M.; Takemura, Y.; Hamada, H.; Sekido, Y.; Kubota, S. EGCG induces human mesothelioma cell death by inducing reactive oxygen;species and autophagy. Cancer Cell Int. 2013, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Ling, T.; Forester, S.C.; Lambert, J.D. Pro-oxidant effects of the green tea catechin, (−)-epigallocatechin-3-gallate in oral cancer cells: A role for the mitochondria. Cancer Res. 2013, 73, 3667. [Google Scholar]
- Tao, L.; Park, J.Y.; Lambert, J.D. Differential prooxidative effects of the green tea polyphenol, (−)-epigallocatechin-3-gallate, in normal and oral cancer cells are related to differences in sirtuin 3 signaling. Mol. Nutr. Food Res. 2015, 59, 203–211. [Google Scholar] [CrossRef]
- Xie, L.; Luo, Z.; Zhao, Z.; Chen, T. Anticancer and antiangiogenic Iron(II) complexes that target thioredoxin reductase to trigger cancer cell apoptosis. J. Med. Chem. 2016, 60, 202–214. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, D.; Cui, W.; Ji, M.; Qian, X.; Zhong, L. Molecular bases of thioredoxin and thioredoxin reductase-mediated prooxidant actions of (−)-epigallocatechin-3-gallate. Free Radic. Biol. Med. 2010, 49, 2010–2018. [Google Scholar] [CrossRef]
- Park, B.J.; Kang, S.S.; Hong, S.G.; Lee, J.H.; Kim, H.S.; Chun, Y.K.; Hong, S.R.; Kang, Y.S.; Moon, I.G.; Lee, S.K.; et al. Telomerase Activity in Human Breast Tumors. J. Natl. Cancer Inst. 1998, 1, 203–207. [Google Scholar] [CrossRef]
- Na, Y.M.; Kim, J.H.; Choi, J.H.; Wen, L.; Ko, Y.J.; Rhee, S.; Bang, H.; Ham, S.W.; Park, A.J.; Lee, K.H. Selective death of cancer cells by preferential induction of reactive oxygen species in response to (−)-epigallocatechin-3-gallate. Biochem. Biophys. Res. Commun 2012, 421, 91–97. [Google Scholar]
- Gonzalez-Donquiles, C.; Alonso-Molero, J.; Fernandez-Villa, T.; Vilorio-Marqués, L.; Molina, A.J.; Martín, V. The NRF2 transcription factor plays a dual role in colorectal cancer: A systematic review. PLoS ONE 2017, 12, e0177549. [Google Scholar] [CrossRef]
- Amin, A.R.M.R.; Wang, D.; Zhang, H.; Peng, S.; Shin, H.J.C.; Brandes, J.C.; Tighiouart, M.; Khuri, F.R.; Chen, Z.G.; Shin, D.M. Enhanced anti-tumor activity by the combination of the natural compounds (−)-epigallocatechin-3-gallate and luteolin: Potential role of p53. J. Biol. Chem. 2010, 285, 34557–34565. [Google Scholar] [CrossRef]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. Oral administration of copper to rats leads to increased lymphocyte cellular DNA degradation by dietary polyphenols: Implications for a cancer preventive mechanism. Biometals 2011, 24, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, N.D.; Zhou, F.; Shen, T.; Duan, T.; Zhou, J.; Shi, Y.; Zhu, X.Q.; Shen, H.M. (−)-Epigallocatechin-3-gallate induces non-apoptotic cell death in human cancer cells via ROS-mediated lysosomal membrane permeabilization. PLoS ONE 2012, 7, e46749. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, X.; Xiao, X.; Chen, D.; Yu, B.; He, J. Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism. Int. J. Mol. Sci. 2019, 20, 5249. https://doi.org/10.3390/ijms20215249
Mao X, Xiao X, Chen D, Yu B, He J. Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism. International Journal of Molecular Sciences. 2019; 20(21):5249. https://doi.org/10.3390/ijms20215249
Chicago/Turabian StyleMao, Xiangbing, Xiangjun Xiao, Daiwen Chen, Bing Yu, and Jun He. 2019. "Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism" International Journal of Molecular Sciences 20, no. 21: 5249. https://doi.org/10.3390/ijms20215249
APA StyleMao, X., Xiao, X., Chen, D., Yu, B., & He, J. (2019). Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism. International Journal of Molecular Sciences, 20(21), 5249. https://doi.org/10.3390/ijms20215249