Silencing of Transcription Factor Sp1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes


Abstract
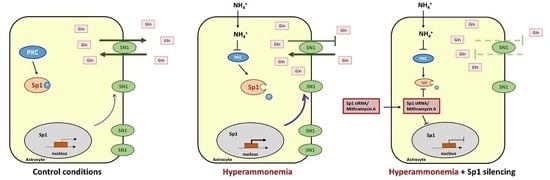
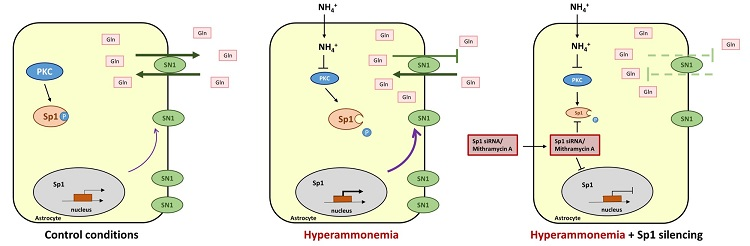
1. Introduction
2. Results
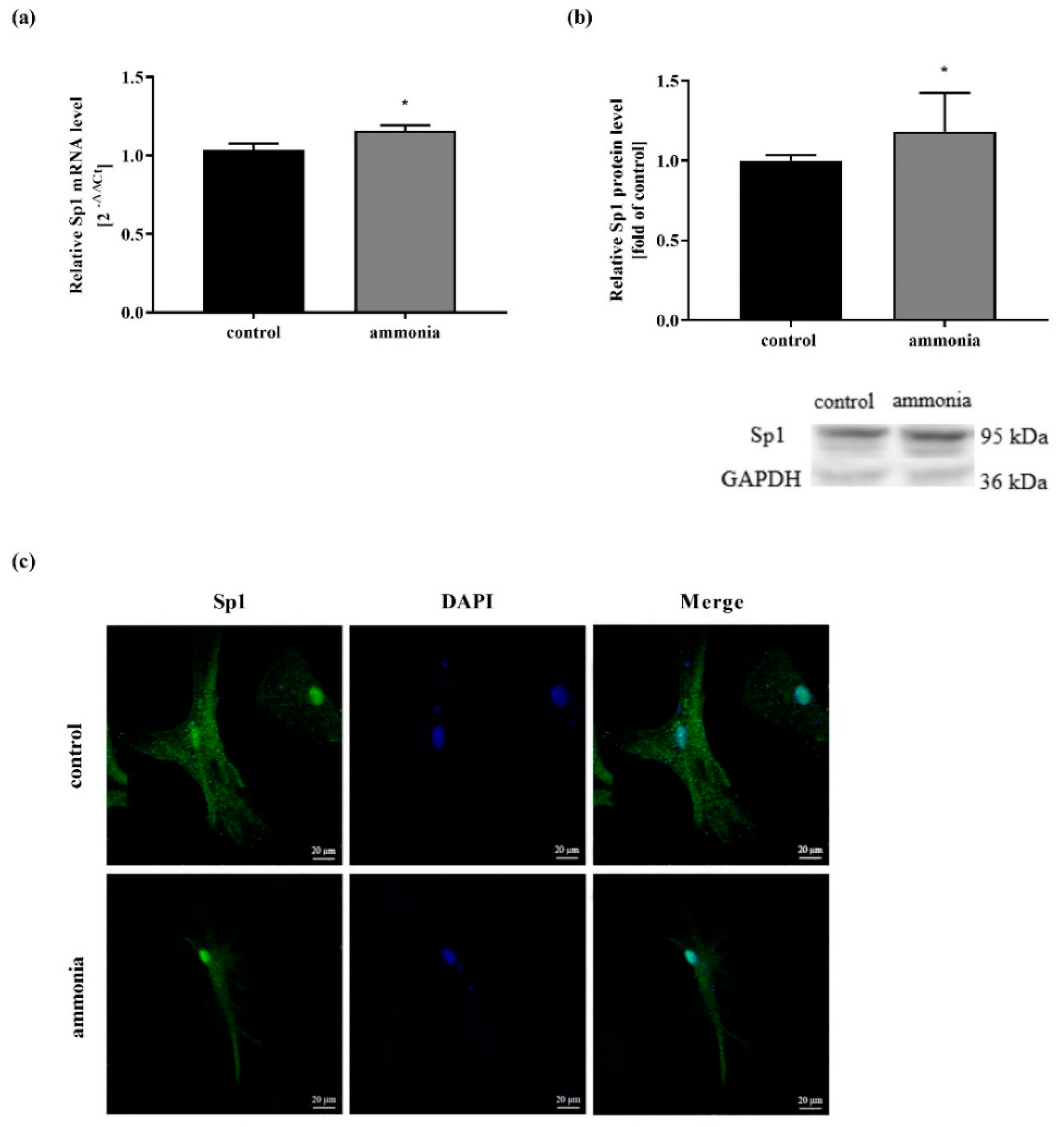
2.1. Ammonia-Induced Sp1 Expression Increase and Sp1 Translocation to the Nucleus
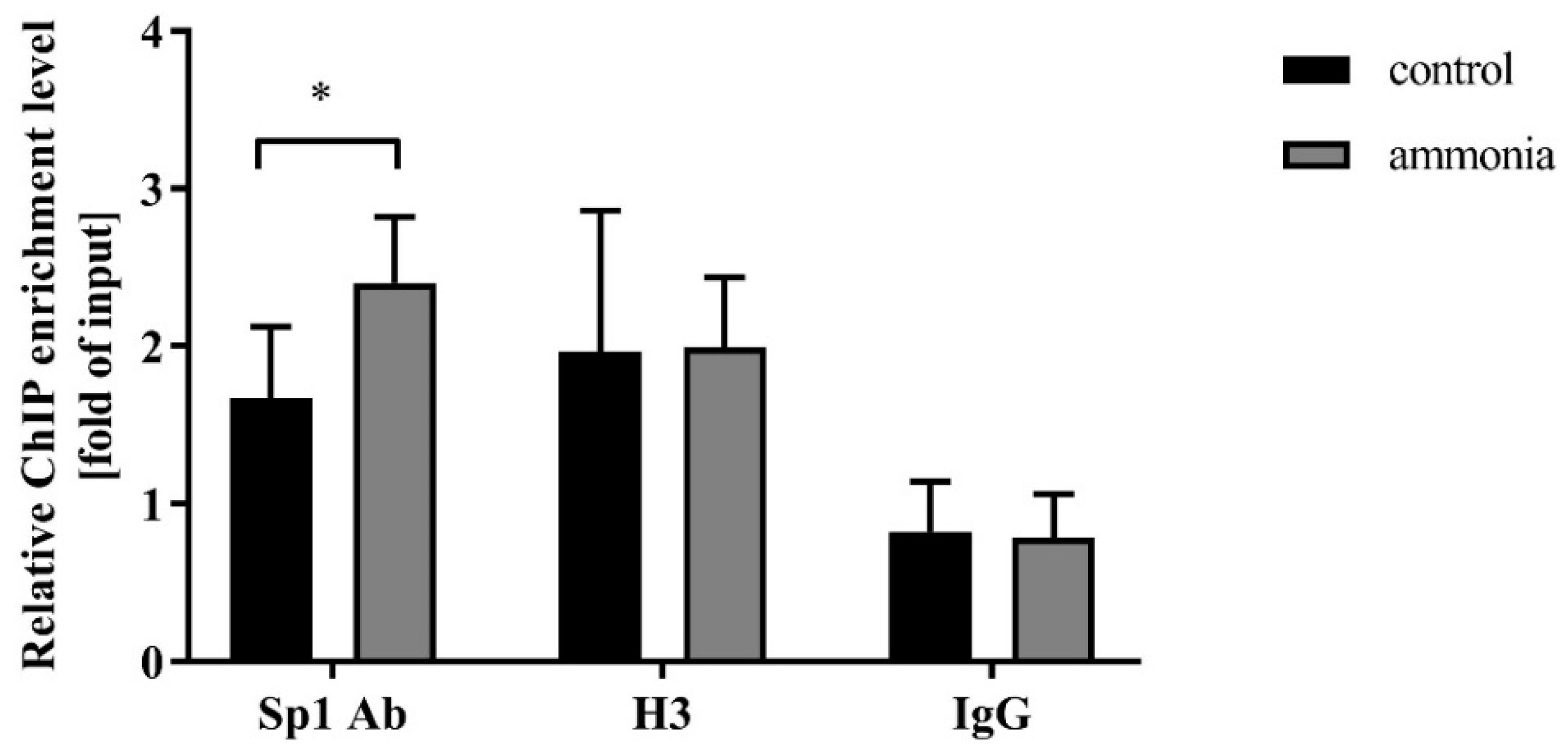
2.2. Ammonia Enhances Sp1 Binding to the Snat3 Promoter Region
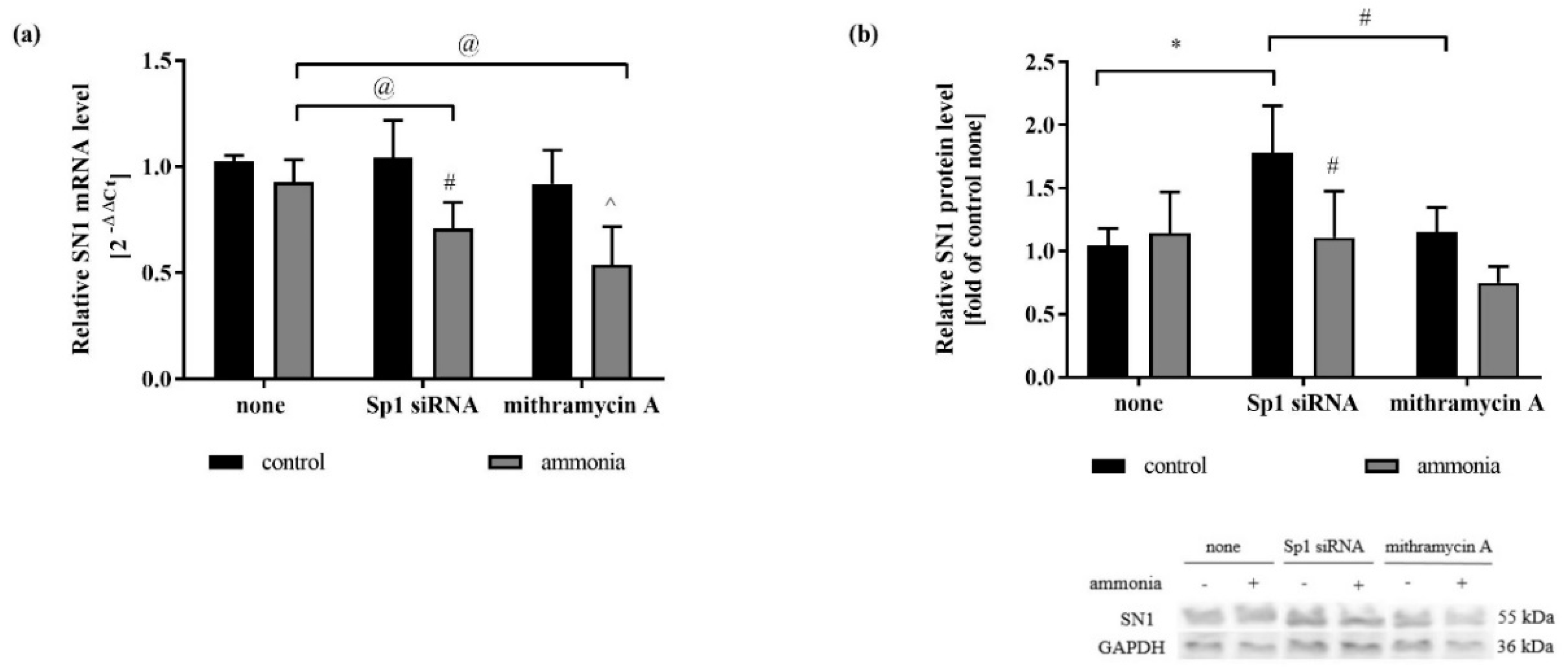
2.3. Silencing of Sp1 Transcription Factor Affects SN1 Expression in the Presence of Ammonia
2.4. The Effect of Sp1 Transcription Factor Silencing on System N Activity in the Presence or Absence of Ammonia
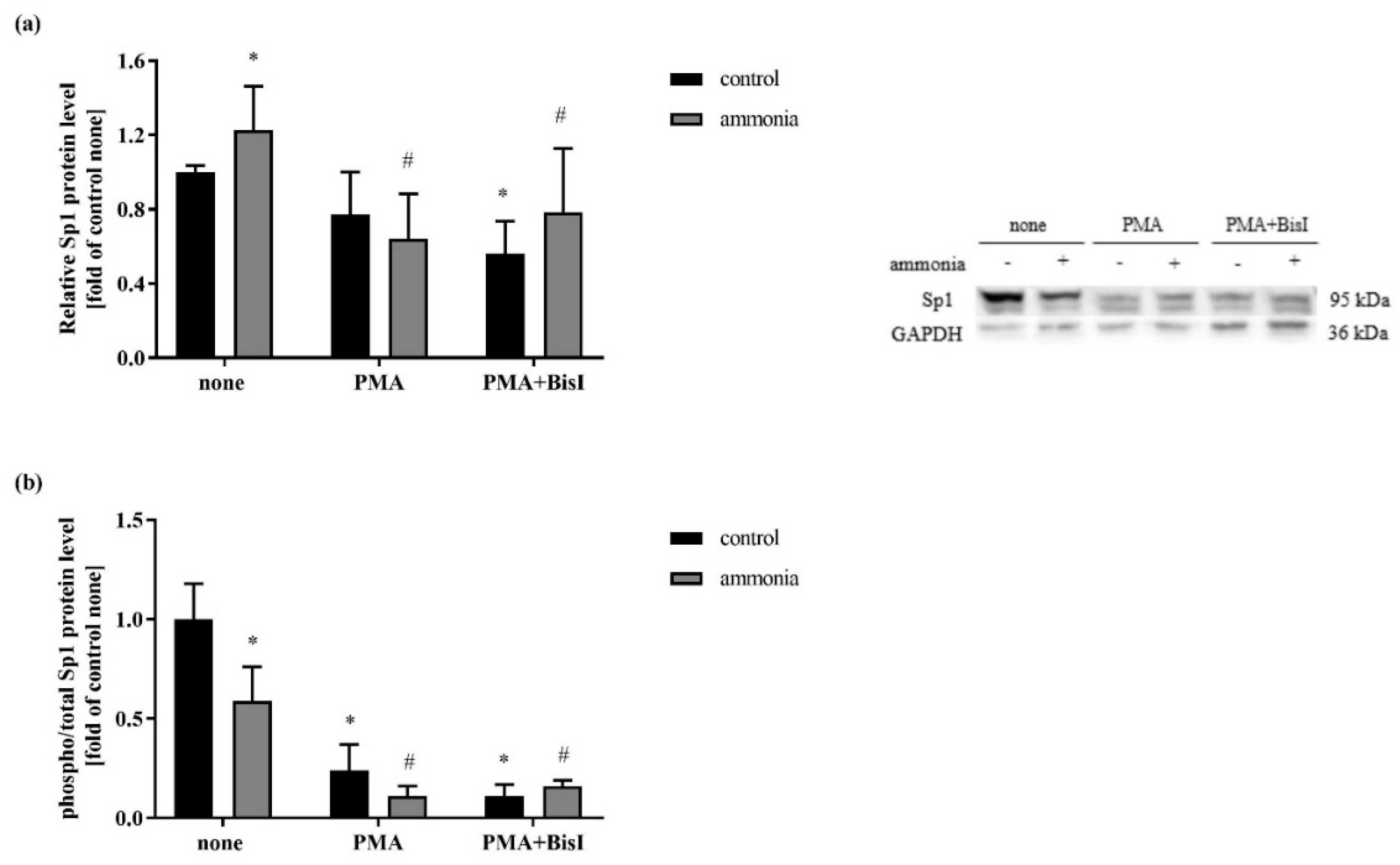
2.5. Sp1 Phosphorylation is Decreased in Astrocytes Treated with Ammonia
2.6. The Effect of PKC Activation on SN1 mRNA Expression in the Presence or Absence of Ammonia
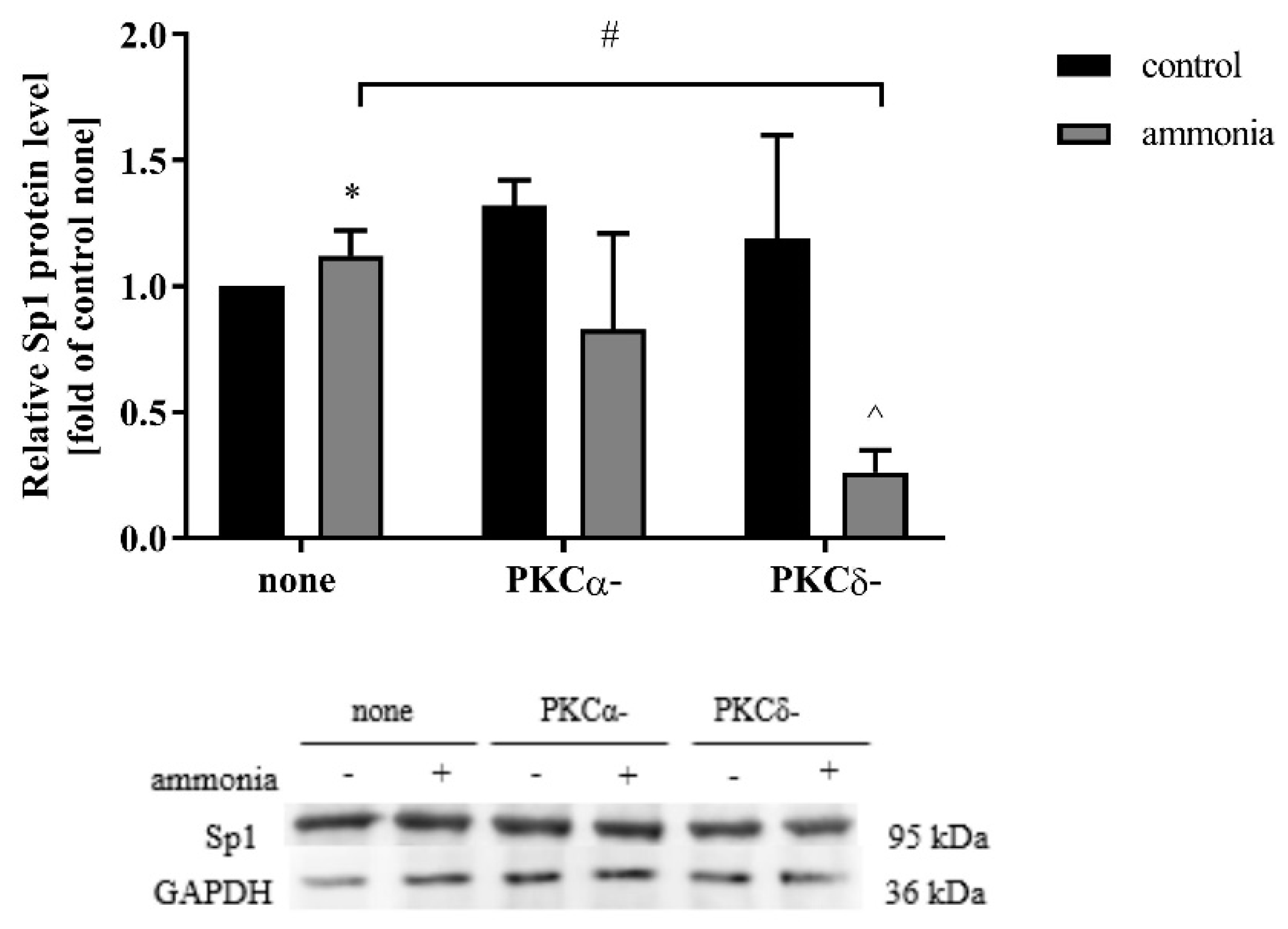
2.7. PKCδ Isoform Depletion Abolishes Ammonia-Induced Increase of Sp1
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Astrocyte Cultures and Treatment
4.3. Sp1 Transcription Factor and PKC Isoforms Silencing
4.4. Real-Time qPCR Analysis
4.5. Protein Isolation and Western Blot
4.6. Gln Uptake
4.7. Chromatin Immunoprecipitation
4.8. Immunocytochemistry
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BisI | bisindolylmaleimide I |
| CDK | cyclin-dependent kinase |
| ChIP | chromatin immunoprecipitation |
| CNS | central nervous system |
| dBcAMP | dibutyryl cyclic adenosine monophosphate |
| ENU | N-ethyl-N-nitrosourea |
| ERK | extracellular signal-regulated kinases |
| GABA | γ-aminobutyric acid |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| Gln | l-glutamine |
| Glu | l-glutamate |
| HE | hepatic encephalopathy |
| MAPK | mitogen-activated protein kinases |
| O-GlcNAc | O-linked N-acetylglucosamine |
| PKA | protein kinase A |
| PKC | protein kinase C |
| PKCα- | silenced expression of protein kinase C isoform α |
| PKCδ- | silenced expression of protein kinase C isoform δ |
| PMA | phorbol 12-myristate 13-acetate |
| SN1 | SNAT3, solute carrier family 38 member 3 |
| SN2 | SNAT5, solute carrier family 38 member 5 |
| Sp1 | specificity protein 1 |
References
- Albrecht, J.; Sonnewald, U.; Waagepetersen, H.S.; Schousboe, A. Glutamine in the central nervous system: Function and dysfunction. Front. Biosci. 2007, 12, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Cynober, L. Plasma amino acid levels with a note on membrane transport: Characteristics, regulation, and metabolic significance. Nutrition 2002, 18, 761–766. [Google Scholar] [CrossRef]
- Pithon-Curi, T.C.; Trezena, A.G.; Tavares-Lima, W.; Curi, R. Evidence that glutamine is involved in neutrophil function. Cell Biochem. Funct. 2002, 20, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, J.; Sidoryk-Węgrzynowicz, M.; Zielińska, M.; Aschner, M. Roles of glutamine in neurotransmission. Neuron Glia Biol. 2010, 6, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Laake, J.H.; Slyngstad, T.A.; Haug, F.M.; Ottersen, O.P. Glutamine from glial cells is essential for the maintenance of the nerve terminal pool of glutamate: Immunogold evidence from hippocampal slice cultures. Neuroscience 1999, 88, 1137–1151. [Google Scholar] [CrossRef]
- Suárez, I.; Bodega, G.; Fernández, B. Glutamine synthetase in brain: Effect of ammonia. Neurochem. Int. 2002, 41, 123–142. [Google Scholar] [CrossRef]
- Waniewski, R.A.; Martin, D.L. Exogenous glutamate is metabolized to glutamine and exported by rat primary astrocyte cultures. J. Neurochem. 1986, 47, 304–313. [Google Scholar] [CrossRef]
- Lavoie, J.; Giguere, J.F.; Layrargues, G.P.; Butterworth, R.F. Amino acid changes in autopsied brain tissue from cirrhotic patients with hepatic encephalopathy. J. Neurochem. 1987, 49, 692–697. [Google Scholar] [CrossRef]
- Ratnakumari, L.; Qureshi, I.A.; Butterworth, R.F. Regional amino acid neurotransmitter changes in brains of Spf/Y mice with congenital ornithine transcarbamylase deficiency. Metab. Brain Dis. 1994, 9, 43–51. [Google Scholar] [CrossRef]
- Butterworth, R.F. Pathophysiology of brain dysfunction in hyperammonemic syndromes: The many faces of glutamine. Mol. Genet. Metab. 2014, 113, 113–117. [Google Scholar] [CrossRef]
- Tofteng, F.; Hauerberg, J.; Hansen, B.A.; Pedersen, C.B.; Jørgensen, L.; Larsen, F.S. Persistent arterial hyperammonemia increases the concentration of glutamine and alanine in the brain and correlates with intracranial pressure in patients with fulminant hepatic failure. J. Cereb. Blood Flow Metab. 2006, 26, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, J.; Zielińska, M.; Norenberg, M.D. Glutamine as a mediator of ammonia neurotoxicity: A critical appraisal. Biochem. Pharmacol. 2010, 80, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, M.; Dąbrowska, K.; Hadera, M.G.; Sonnewald, U.; Albrecht, J. System N transporters are critical for glutamine release and modulate metabolic fluxes of glucose and acetate in cultured cortical astrocytes: Changes induced by ammonia. J. Neurochem. 2016, 2, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Pichili, V.B.; Rao, K.V.; Jayakumar, A.R.; Norenberg, M.D. Inhibition of glutamine ransport into mitochondria protects astrocytes from ammonia toxicity. Glia 2007, 55, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Ziemińska, E.; Dolińska, M.; Lazarewicz, J.W.; Albrecht, J. Induction of permeability transition and swelling of rat brain mitochondria by glutamine. Neurotoxicology 2000, 21, 295–300. [Google Scholar] [PubMed]
- Bai, G.; Rama Rao, C.R.K.; Norenberg, M.D. Ammonia induces the mitochondrial permeability transition in primary cultures of astrocytes. J. Neurosci. Res. 2001, 66, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Kruczek, C.; Görg, B.; Keitel, V.; Bidmon, H.J.; Schliess, F.; Häussinger, D. Ammonia increases nitric oxide, free Zn2+, and metallothionein mRNA expression in cultured rat astrocytes. Biol. Chem. 2011, 392, 1155–1165. [Google Scholar] [CrossRef]
- Sinke, A.P.; Jayakumar, A.R.; Panickar, K.S.; Moriyama, M.; Reddy, P.V.; Norenberg, M.D. NFkappaB in the mechanism of ammonia-induced astrocyte swelling in culture. J. Neurochem. 2008, 106, 2302–2311. [Google Scholar] [CrossRef]
- Blei, A.T.; Larsen, F.S. Pathophysiology of cerebral edema in fulminant hepatic failure. J. Hepatol. 1999, 31, 771–776. [Google Scholar] [CrossRef]
- Häussinger, D.; Kircheis, G.; Fischer, R.; Schliess, F.; vom Dahl, S. Hepatic encephalopathy in chronic liver disease: A clinical manifestation of astrocyte swelling and low-grade cerebral edema? J. Hepatol. 2000, 32, 1035–1038. [Google Scholar] [CrossRef]
- Jayakumar, A.R.; Panickar, K.S.; Murthy, Ch.R.K.; Norenberg, M.D. Oxidative stress and MAPK phosphorylation mediate ammonia-induced cell swelling and glutamate uptake inhibition in cultured astrocytes. J. Neurosci. 2006, 26, 4774–4784. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-activated protein (MAP) kinase pathways: Regulation and physiological functions. Endocrinol. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef]
- Bröer, A.; Deitmer, J.W.; Bröer, S. Astroglial glutamine transport by system N is upregulated by glutamate. GLIA 2004, 48, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K.; Albrecht, J.; Zielińska, M. Protein kinase C-mediated impairment of glutamine outward transport and SN1 transporter distribution by ammonia in mouse cortical astrocytes. Neurochem. Int. 2018, 118, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Busque, S.M.; Sailer, M.; Stoeger, C.; Bröer, S.; Daniel, H.; Rubio-Aliaga, I.; Wagner, C.A. Loss of function mutation of the Slc38a3 glutamine transporter reveals its critical role for amino acid metabolism in the liver, brain, and kidney. Pflugers Arch. 2016, 468, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Boulland, J.L.; Osen, K.K.; Levy, L.M.; Danbolt, N.C.; Edwards, R.H.; Storm-Mathisen, J.; Chaudhry, F.A. Cell-specific expression of the glutamine transporter SN1 suggests differences in dependence on the glutamine cycle. Eur. J. Neurosci. 2002, 15, 1615–1631. [Google Scholar] [CrossRef] [PubMed]
- Balkrishna, S.; Bröer, A.; Welford, S.M.; Hatzoglou, M.; Bröer, S. Expression of glutamine transporter Slc38a3 (SNAT3) during acidosis is mediated by a different mechanism than tissue-specific expression. Cell. Physiol. Biochem. 2014, 33, 1591–1606. [Google Scholar] [CrossRef] [PubMed]
- Solbu, T.T.; Boulland, J.L.; Zahid, W.; Lyamouri Bredahl, M.K.; Amiry-Moghaddam, M.; Storm-Mathisen, J.; Roberg, B.A.; Chaudhry, F.A. Induction and targeting of the glutamine transporter SN1 to the basolateral membranes of cortical kidney tubule cells during chronic metabolic acidosis suggest a role in pH regulation. J. Am. Soc. Nephrol. 2005, 16, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Black, A.R.; Black, J.D.; Azizkhan-Clifford, J. Sp1 and krüppel-like factor family of transcription factors in cell growth regulation and cancer. J. Cell. Physiol. 2001, 188, 143–160. [Google Scholar] [CrossRef]
- Philipsen, S.; Suske, G. A tale of three fingers: The family of mammalian Sp/XKLF transcription factors. Nucleic Acids Res. 1999, 27, 2991–3000. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium. A user’s guide to the encyclopedia of DNA elements (ENCODE). PLoS Biol. 2011, 9, e1001046. [Google Scholar]
- Bodega, G.; Suarez, I.; Lopez-Fernandez, L.A.; Almonacid, L.; Zaballos, A.; Fernandez, B. Possible implication of ciliary neurotrophic factor (CNTF) and b-synuclein in the ammonia effect on cultured rat astroglial cells: A study using DNA and protein microarrays. Neurochem. Int. 2006, 48, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Eviatar-Ribak, T.; Miskimins, R. Sp1 phosphorylation is involved in myelin basic protein gene transcription. J. Neurosci. Res. 2010, 88, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.Y.; Midgley, V.C.; Kavurma, M.M.; Santiago, F.S.; Luo, X.; Peden, R.; Fahmy, R.G.; Berndt, M.C.; Molloy, M.P.; Khachigian, L.M. Angiotensin II-inducible platelet-derived growth factor-D transcription requires specific Ser/Thr residues in the second zinc finger region of Sp1. Circ. Res. 2008, 102, e38–e51. [Google Scholar] [CrossRef] [PubMed]
- Rohlff, C.; Ahmad, S.; Borellini, F.; Lei, J.; Glazer, R.I. Modulation of transcription factor Sp1 by cAMP-dependent protein kinase. J. Biol. Chem. 1997, 272, 21137–21141. [Google Scholar] [CrossRef] [PubMed]
- Fojas de Borja, P.; Collins, N.K.; Du, P.; Azizkhan-Clifford, J.; Mudryj, M. Cyclin A-CDK phosphorylates Sp1 and enhances Sp1-mediated transcription. EMBO J. 2001, 20, 5737–5747. [Google Scholar] [CrossRef]
- Milanini-Mongiat, J.; Pouysségur, J.; Pagès, G. Identification of two Sp1 phosphorylation sites for p42/p44 mitogen-activated protein kinases: Their implication in vascular endothelial growth factor gene transcription. J. Biol. Chem. 2002, 277, 20631–20639. [Google Scholar] [CrossRef]
- Lichter-Konecki, U.; Mangin, J.M.; Gordish-Dressman, H.; Hoffman, E.P.; Gallo, V. Gene expression profiling of astrocytes from hyperammonemic mice reveals altered pathways for water and potassium homeostasis in vivo. GLIA 2008, 56, 365–377. [Google Scholar] [CrossRef]
- Song, G.; Dhodda, V.K.; Blei, A.T.; Dempsey, R.J.; Rao, V.L.R. GeneChip analysis shows altered mRNA expression of transcripts of neurotransmitter and signal transduction pathways in the cerebral cortex of portacaval shunted rats. J. Neurosci. Res. 2002, 68, 730–737. [Google Scholar] [CrossRef]
- Warskulat, U.; Görg, B.; Bidmon, H.J.; Müller, H.W.; Schliess, F.; Häussinger, D. Ammonia-induced heme oxygenase 1 expression in cultured rat astrocytes and in rat brain in vivo. GLIA 2002, 40, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Warskulat, U.; Kreuels, S.; Müller, H.W.; Häussinger, D. Identification of osmosensitive and ammonia-regulated genes in rat astrocytes by northern blotting and differential display reverse transcriptase-polymerase chain reaction. J. Hepatol. 2001, 35, 358–366. [Google Scholar] [CrossRef]
- Bodega, G.; Segura, B.; Ciordia, S.; Mena Mdel, C.; López-Fernández, L.A.; García, M.I.; Trabado, I.; Suárez, I. Ammonia Affects Astroglial Proliferation in Culture. PLoS ONE 2015, 10, e0139619. [Google Scholar] [CrossRef] [PubMed]
- Sidoryk-Wegrzynowicz, M.; Lee, E.; Mingwei, N.; Aschner, M. Disruption of astrocytic glutamine turnover by manganese is mediated by the protein kinase C pathway. GLIA 2011, 59, 1732–1743. [Google Scholar] [CrossRef] [PubMed]
- Leggett, R.W.; Armstrong, S.A.; Barry, D.; Mueller, C.R. Sp1 is phosphorylated and its DNA binding activity down-regulated upon terminal differentiation of the liver. J. Biol. Chem. 1995, 270, 25879–25884. [Google Scholar] [CrossRef] [PubMed]
- Apt, D.; Watts, R.M.; Suske, G.; Bernard, H.U. High Sp1/Sp3 ratios in epithelial cells during epithelial differentiation and cellular transformation correlate with the activation of the HPV-16 promoter. Virology 1996, 224, 281–291. [Google Scholar] [CrossRef] [PubMed]
- You, H.L.; Eng, H.L.; Hsu, S.F.; Chen, C.M.; Ye, T.C.; Liao, W.T.; Huang, M.Y.; Baer, R.; Cheng, J.T. A PKC-Sp1 signaling pathway induces early differentiation of human keratinocytes through upregulation of TSG101. Cell. Signal. 2007, 19, 1201–1211. [Google Scholar] [CrossRef]
- Gottlieb, T.M.; Jackson, S.P. The DNA-dependent protein kinase: Requirement for DNA ends and association with Ku antigen. Cell 1993, 72, 131–142. [Google Scholar] [CrossRef]
- Suh, H.N.; Lee, Y.J.; Kim, M.O.; Ryu, J.M.; Han, H.J. Glucosamine-induced Sp1 O-GlcNAcylation ameliorates hypoxia-induced SGLT dysfunction in primary cultured renal proximal tubule cells. J. Cell. Physiol. 2014, 229, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Sangwan, V.; McGinn, O.; Chugh, R.; Dudeja, V.; Vickers, S.M.; Saluja, A.K. Triptolide-induced cell death in pancreatic cancer is mediated by O-GlcNAc modification of transcription factor Sp1. J. Biol. Chem. 2013, 288, 33927–33938. [Google Scholar] [CrossRef]
- Du, X.L.; Edelstein, D.; Rossetti, L.; Fantus, I.G.; Goldberg, H.; Ziyadeh, F.; Wu, J.; Brownlee, M. Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. USA 2000, 97, 12222–12226. [Google Scholar] [CrossRef] [PubMed]
- Roos, M.D.; Su, K.; Baker, J.R.; Kudlow, J.E. O glycosylation of an Sp1-derived peptide blocks known Sp1 protein interactions. Mol. Cell. Biol. 1997, 17, 6472–6480. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.J.; Wang, Y.T.; Chang, W.C. Sp1 deacetylation induced by phorbol ester recruits p300 to activate 12(S)-lipoxygenase gene transcription. Mol. Cell. Biol. 2006, 26, 1770–1785. [Google Scholar] [CrossRef]
- Waby, J.S.; Chirakkal, H.; Yu, C.; Griffiths, G.J.; Benson, R.S.; Bingle, C.D.; Corfe, B.M. Sp1 acetylation is associated with loss of DNA binding at promoters associated with cell cycle arrest and cell death in a colon cell line. Mol. Cancer 2010, 9, 275. [Google Scholar] [CrossRef] [PubMed]
- Tretiakova, A.; Steplewski, A.; Johnson, E.M.; Khalili, K.; Amini, S. Regulation of myelin basic protein gene transcription by Sp1 and Puralpha: Evidence for association of Sp1 and Puralpha in brain. J. Cell. Physiol. 1999, 181, 160–168. [Google Scholar] [CrossRef]
- Chi, P.L.; Lin, C.C.; Chen, Y.W.; Hsiao, L.D.; Yang, C.M. CO Induces Nrf2-Dependent Heme Oxygenase-1 Transcription by Cooperating with Sp1 and c-Jun in Rat Brain Astrocytes. Mol. Neurobiol. 2015, 52, 277–292. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Luo, Y.; He, J.; Ma, H.; Shen, L.; Tang, J.; Fang, Q.; Wang, H.; Qian, Z.; et al. Sp-1 Negatively Regulates miR-20b Expression in Macrophages. Ann. Clin. Lab. Sci. 2017, 47, 706–712. [Google Scholar] [PubMed]
- Shou, Y.; Baron, S.; Poncz, M. An Sp1-binding Silencer Element Is a Critical Negative Regulator of the Megakaryocyte-specific αIIb Gene. J. Biol. Chem. 1998, 273, 5716–5726. [Google Scholar] [CrossRef] [PubMed]
- Lister, A.; Bourgeois, S.; Imenez Silva, P.H.; Rubio-Aliaga, I.; Marbet, P.; Walsh, J.; Shelton, L.M.; Keller, B.; Verrey, F.; Devuyst, O.; et al. NRF2 regulates the glutamine transporter Slc38a3 (SNAT3) in kidney in response to metabolic acidosis. Sci. Rep. 2018, 8, 5629. [Google Scholar] [CrossRef]
- Butterworth, R.F. Pathophysiology of hepatic encephalopathy: A new look at ammonia. Metab. Brain Dis. 2002, 17, 221–227. [Google Scholar] [CrossRef]
- Norenberg, M.D.; Baker, L.; Norenberg, L.O.; Blicharska, J.; Bruce-Gregorios, J.H.; Neary, J.T. Ammonia-induced astrocyte swelling in primary culture. Neurochem. Res. 1991, 16, 833–836. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | [3H]glutamine Uptake | Ammonia | Value (Fold of Control None) |
|---|---|---|---|
| none | total | − | 0.98 ± 0.09 |
| + | 1.01 ± 0.17 | ||
| system N | − | 0.39 ± 0.11 * | |
| + | 0.36 ± 0.13 & | ||
| Sp1 siRNA | total | − | 1.04 ± 0.19 |
| + | 0.73 ± 0.12 ^,& | ||
| system N | − | 0.42 ± 0.06 ^ | |
| + | 0.29 ± 0.06 +,! | ||
| mithramycin A | total | − | 1.28 ± 0.24 * |
| + | 1.10 ± 0.14 $ | ||
| system N | − | 0.24 ± 0.10 # | |
| + | 0.15 ± 0.04 ? |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowska, K.; Zielińska, M. Silencing of Transcription Factor Sp1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes. Int. J. Mol. Sci. 2019, 20, 234. https://doi.org/10.3390/ijms20020234
Dąbrowska K, Zielińska M. Silencing of Transcription Factor Sp1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes. International Journal of Molecular Sciences. 2019; 20(2):234. https://doi.org/10.3390/ijms20020234
Chicago/Turabian StyleDąbrowska, Katarzyna, and Magdalena Zielińska. 2019. "Silencing of Transcription Factor Sp1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes" International Journal of Molecular Sciences 20, no. 2: 234. https://doi.org/10.3390/ijms20020234
APA StyleDąbrowska, K., & Zielińska, M. (2019). Silencing of Transcription Factor Sp1 Promotes SN1 Transporter Regulation by Ammonia in Mouse Cortical Astrocytes. International Journal of Molecular Sciences, 20(2), 234. https://doi.org/10.3390/ijms20020234