Hyperbaric Oxygen Treatment Ameliorates Hearing Loss and Auditory Cortex Injury in Noise Exposed Mice by Repressing Local Ceramide Accumulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
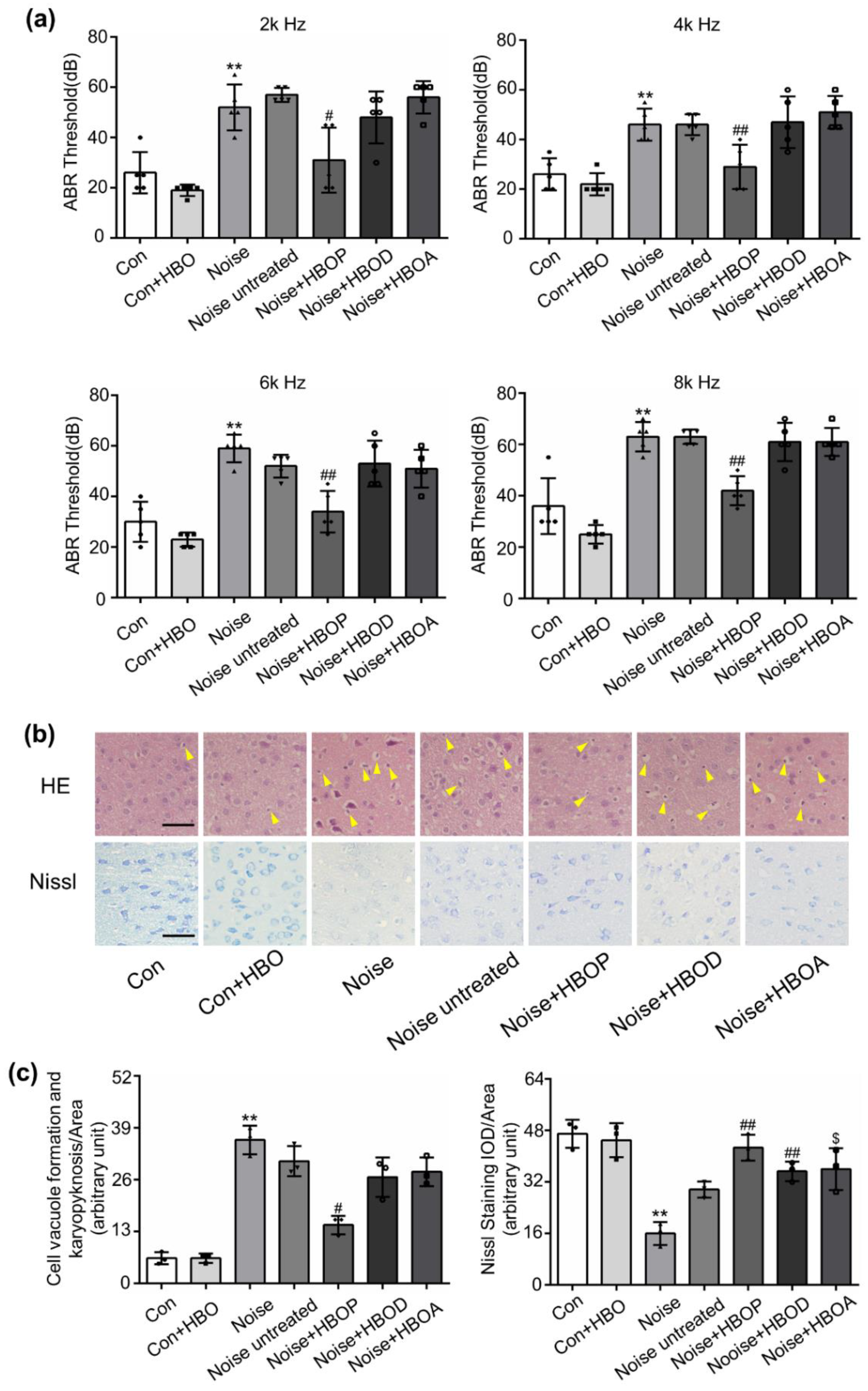
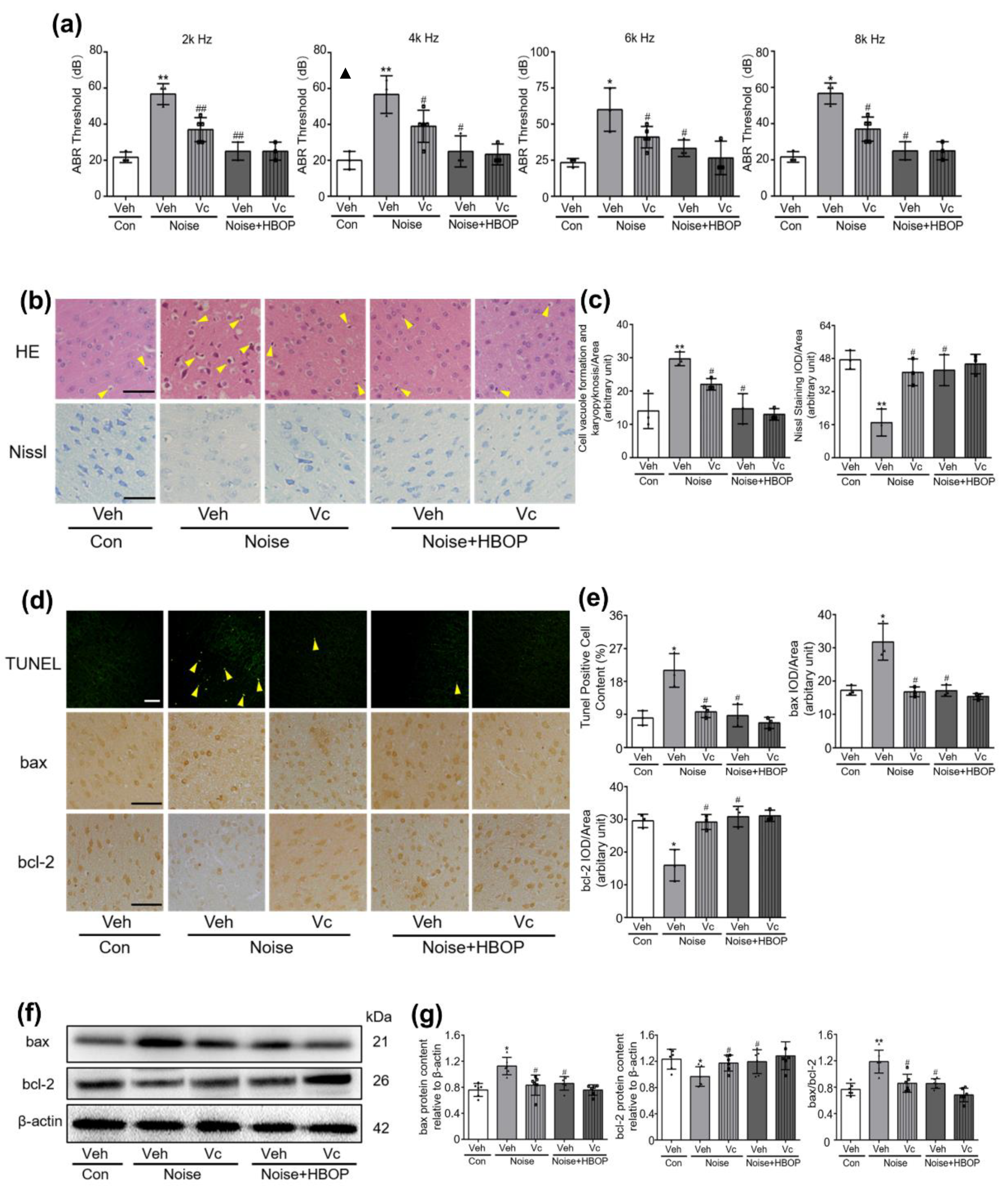
2.1. HBOT Corrected Noise-Induced Hearing Loss and Morphologic Disorder in AC of Mice
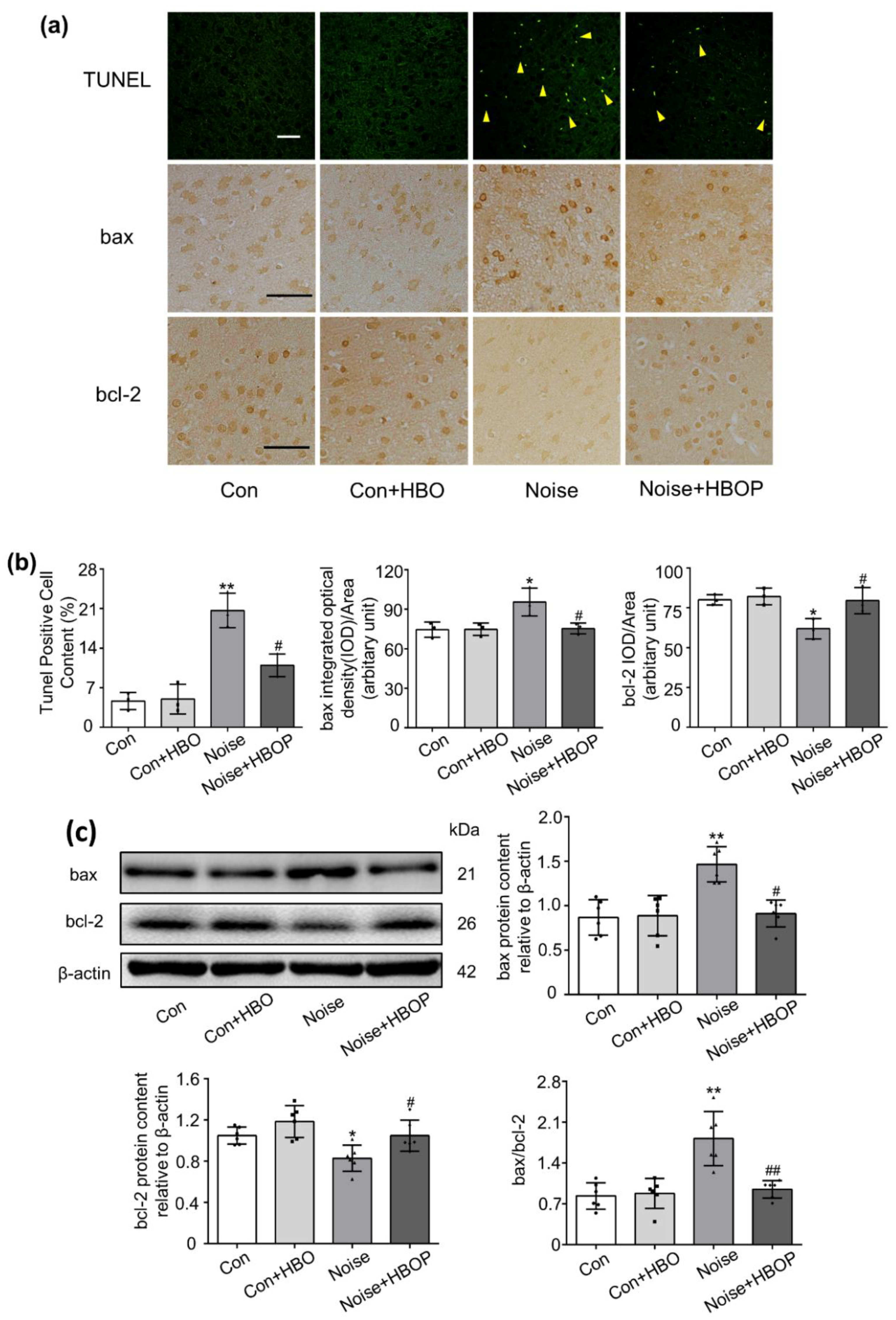
2.2. HBOP Suppressed Neuronal Apoptosis in AC of Noise Exposed Mice
2.3. HBOP Normalized the Cer Generation in AC of Noise Exposed Mice
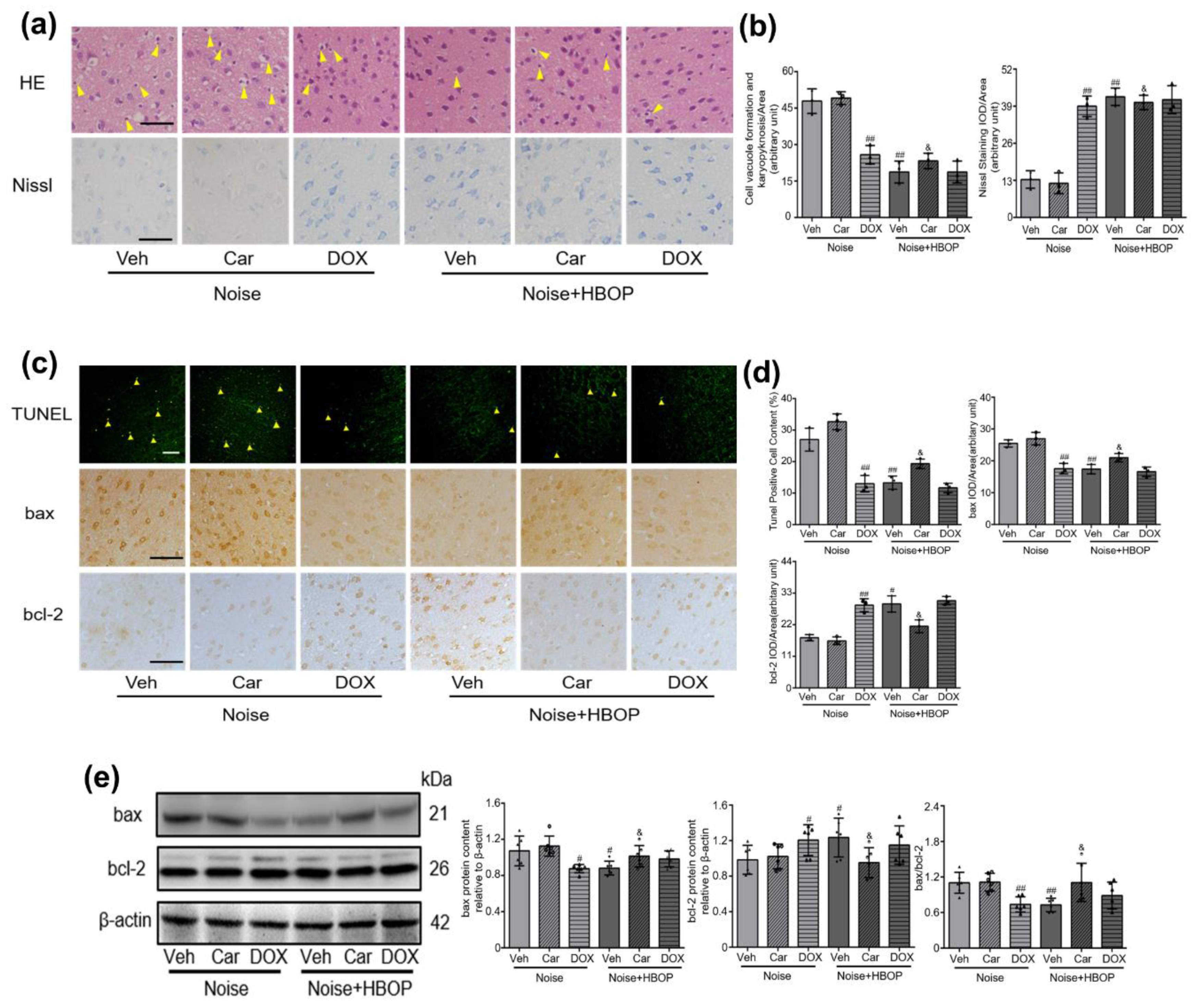
2.4. Cer Mediated the Curative Effect of HBOP on Noise-Induced Neuronal Damage and Apoptosis
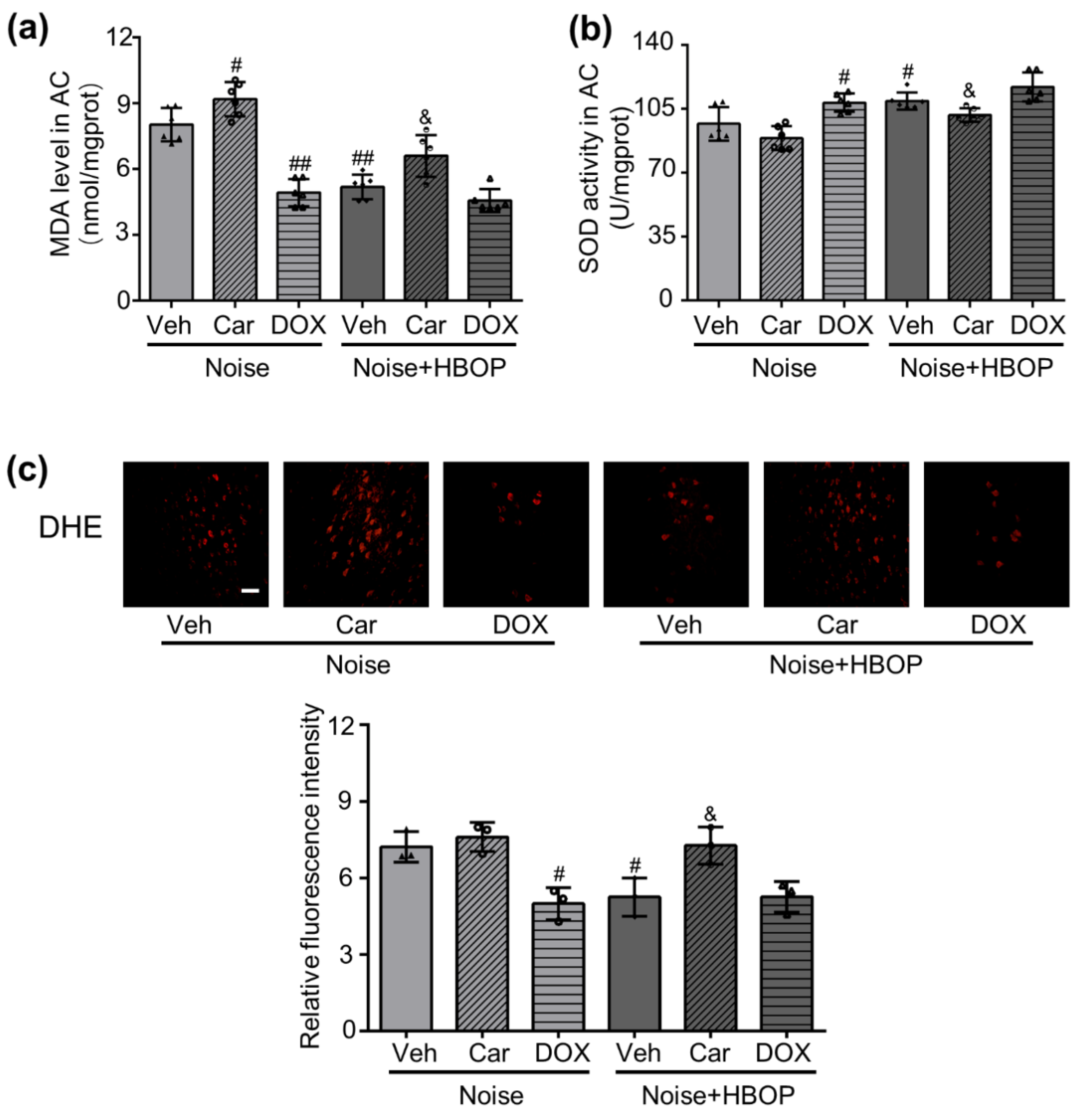
2.5. HBOP Restrained the Oxidative Stress in AC of Noise Exposed Mice by Regulating Cer
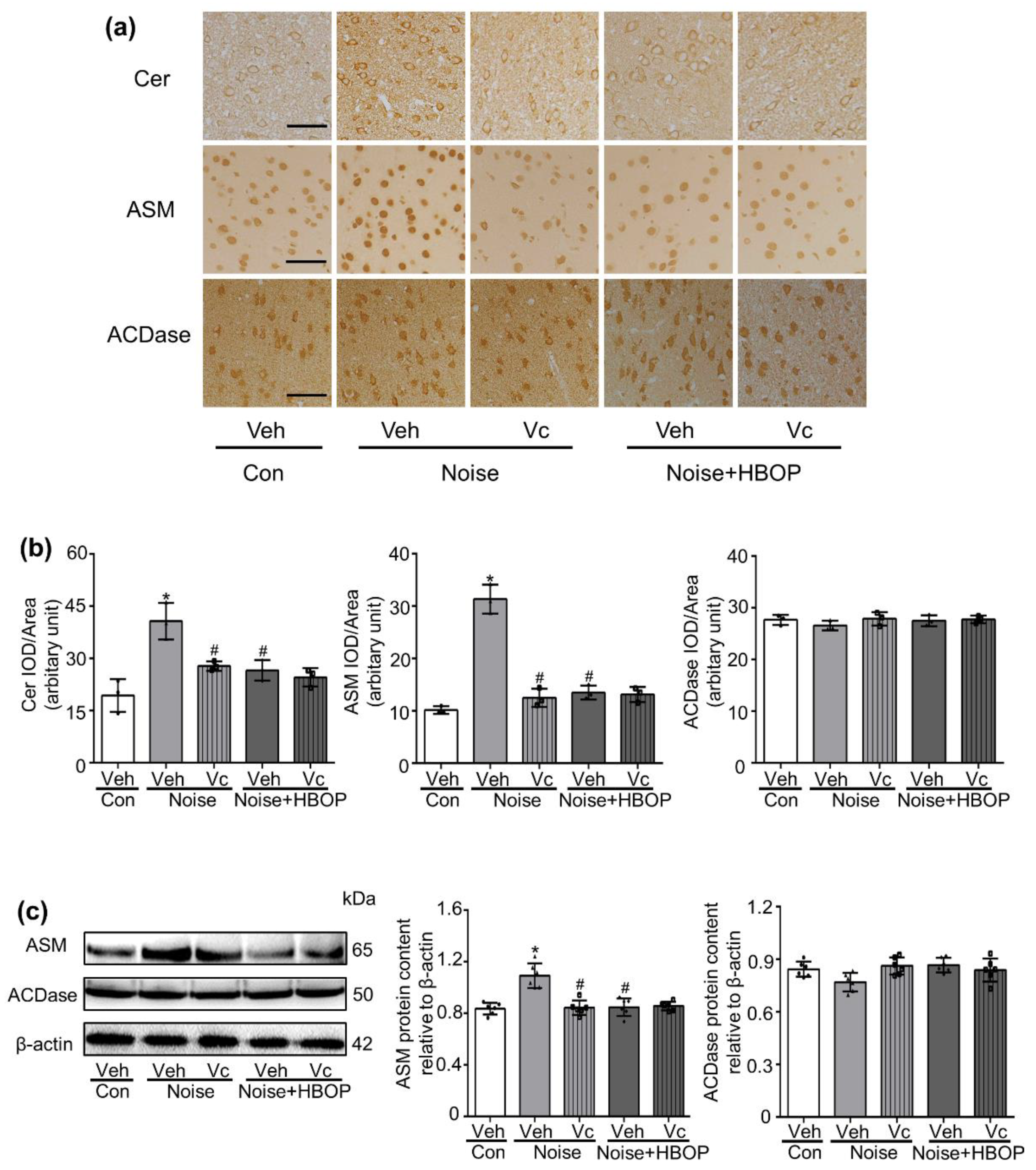
2.6. Vc Neutralized Oxidative Stress, Restoring the Hearing Loss and Neuronal Damage in AC of Noise Exposed Mice
2.7. Vc Normalized the Cer Generation in AC of Noise Exposed Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Animal Experimental Groups and Experimental Design
4.4. Noise Exposure
4.5. Hyperbaric Oxygen Therapy
4.6. Auditory Brainstem Response (ABR) Assay
4.7. Tissue Preparation
4.8. Hematoxylin Eosin (HE) Staining
4.9. Nissl Staining
4.10. Transferase-Mediated dUTP Nick end Labeling (TUNEL) Staining
4.11. Immunohistochemistry (IHC) Assay
4.12. Western Blot Analysis
4.13. Detection of Lipid Peroxidation, Superoxide Dismutase (SOD) Activity
4.14. Detection of Superoxide Production
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A1 | Primary auditory cortex |
| ABR | Auditory brainstem response |
| AC | auditory cortex |
| ACDase | acid ceramidase |
| ASM | acid sphingomyelinase |
| bcl-2 | B cell lymphoma 2 |
| bax | bcl-2 associated X protein |
| Car | Carmofur |
| Cer | Ceramide |
| DOX | doxepin hydrochloride |
| FDA | U.S. Food and Drug Administration |
| HBOT | hyperbaric oxygen therapy |
| HBOA | hyperbaric oxygen therapy after noise exposure |
| HBOD | hyperbaric oxygen therapy during noise exposure |
| HBOP | hyperbaric oxygen therapy before noise exposure |
| HE | hematoxylin-eosin |
| IHC | immunohistochemistry |
| JNK | c-Jun N-terminal kinase |
| MDA | malondialdehyde |
| MGB | medial geniculate body |
| NMDA | N-methyl-D-aspartic acid receptor |
| NOS | nitric oxide synthase |
| SLs | sphingolipids |
| SOD | superoxide dismutase |
| TUNEL | transferase-mediated dUTP nick end labeling |
| Vc | Vitamin C |
| Veh | Vehicle |
References
- Basta, D.; Tzschentke, B.; Ernst, A. Noise-induced cell death in the mouse medial geniculate body and primary auditory cortex. Neurosci. Lett. 2005, 381, 199–204. [Google Scholar] [CrossRef]
- Coordes, A.; Groschel, M.; Ernst, A.; Basta, D. Apoptotic cascades in the central auditory pathway after noise exposure. J. Neurotrauma 2012, 29, 1249–1254. [Google Scholar] [CrossRef]
- Su, Y.T.; Meng, X.X.; Zhang, X.; Guo, Y.B.; Zhang, H.J.; Cheng, Y.P.; Xie, X.P.; Chang, Y.M.; Bao, J.X. Doxepin Mitigates Noise-induced Neuronal Damage in Primary Auditory Cortex of Mice via Suppression of Acid Sphingomyelinase/Ceramide Pathway. Anat. Rec. 2017, 300, 2220–2232. [Google Scholar] [CrossRef]
- Frohlich, F.; Ernst, A.; Strubing, I.; Basta, D.; Groschel, M. Apoptotic mechanisms after repeated noise trauma in the mouse medial geniculate body and primary auditory cortex. Exp. Brain Res. 2017, 235, 3673–3682. [Google Scholar] [CrossRef]
- Thom, S.R. Hyperbaric oxygen: Its mechanisms and efficacy. Plast. Reconstr. Surg. 2011, 127 (Suppl. 1), 131S–141S. [Google Scholar] [CrossRef]
- Glik, J.; Cholewka, A.; Stanek, A.; Englisz, B.; Sieron, K.; Mikus-Zagorska, K.; Knefel, G.; Nowak, M.; Kawecki, M. Thermal imaging and planimetry evaluation of the results of chronic wounds treatment with hyperbaric oxygen therapy. Adv. Clin. Exp. Med. 2019, 28, 229–236. [Google Scholar] [CrossRef]
- Cholewka, A.; Knefel, G.; Stanek, A.; Kawecki, M.; Nowak, M.; Aleksander, S.; Zofia, D. Thermal imaging and TC oximetry measurements of hyperbaric oxygen therapy (HBO) effects on trophic ulceration of the crura. J. Therm. Anal. Calorim. 2012, 108, 25–31. [Google Scholar]
- Gebala-Prajsnar, K.; Stanek, A.; Pasek, J. Selected physical medicine interventions in the treatment of diabetic foot syndrome. Acta Angiol. 2015, 21, 140–145. [Google Scholar] [CrossRef]
- Lin, P.Y.; Sung, P.H.; Chung, S.Y.; Hsu, S.L.; Chung, W.J.; Sheu, J.J.; Hsueh, S.K.; Chen, K.H.; Wu, R.W.; Yip, H.K. Hyperbaric Oxygen Therapy Enhanced Circulating Levels of Endothelial Progenitor Cells and Angiogenesis Biomarkers, Blood Flow, in Ischemic Areas in Patients with Peripheral Arterial Occlusive Disease. J. Clin. Med. 2018, 7, 548. [Google Scholar] [CrossRef]
- Ajduk, J.; Ries, M.; Trotic, R.; Marinac, I.; Vlatka, K.; Bedekovic, V. Hyperbaric Oxygen Therapy as Salvage Therapy for Sudden Sensorineural Hearing Loss. J. Int. Adv. Otol. 2017, 13, 61–64. [Google Scholar] [CrossRef]
- Bao, J.X.; Su, Y.T.; Cheng, Y.P.; Zhang, H.J.; Xie, X.P.; Chang, Y.M. Vascular sphingolipids in physiological and pathological adaptation. Front. Biosci. 2016, 21, 1168–1186. [Google Scholar]
- Ong, W.Y.; Herr, D.R.; Farooqui, T.; Ling, E.A.; Farooqui, A.A. Role of sphingomyelinases in neurological disorders. Expert Opin. Ther. Targets 2015, 19, 1725–1742. [Google Scholar] [CrossRef]
- Bao, J.X.; Jin, S.; Zhang, F.; Wang, Z.C.; Li, N.; Li, P.L. Activation of membrane NADPH oxidase associated with lysosome-targeted acid sphingomyelinase in coronary endothelial cells. Antioxid. Redox Signal. 2010, 12, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Fong, Y.; Tsai, E.M.; Chang, Y.G.; Chou, H.L.; Wu, C.Y.; Teng, Y.N.; Liu, T.C.; Yuan, S.S.; Chiu, C.C. Exogenous C(8)-Ceramide Induces Apoptosis by Overproduction of ROS and the Switch of Superoxide Dismutases SOD1 to SOD2 in Human Lung Cancer Cells. Int. J. Mol. Sci. 2018, 19, 3010. [Google Scholar] [CrossRef] [PubMed]
- Cogolludo, A.; Villamor, E.; Perez-Vizcaino, F.; Moreno, L. Ceramide and Regulation of Vascular Tone. Int. J. Mol. Sci. 2019, 20, 411. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Pallichankandy, S.; Thayyullathil, F.; Galadari, S. Critical role of H2O2 in mediating sanguinarine-induced apoptosis in prostate cancer cells via facilitating ceramide generation, ERK1/2 phosphorylation, and Par-4 cleavage. Free Radic. Biol. Med. 2019, 134, 527–544. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shirachi, D.Y.; Quock, R.M. The acute antinociceptive effect of hyperbaric oxygen is not accompanied by an increase in markers of oxidative stress. Life Sci. 2014, 98, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Jou, I.; Park, S.M. Attenuation of noise-induced hearing loss using methylene blue. Cell Death Dis. 2014, 5, e1200. [Google Scholar] [CrossRef] [PubMed]
- Groschel, M.; Gotze, R.; Ernst, A.; Basta, D. Differential impact of temporary and permanent noise-induced hearing loss on neuronal cell density in the mouse central auditory pathway. J. Neurotrauma 2010, 27, 1499–1507. [Google Scholar] [CrossRef]
- Lynch, E.D.; Kil, J. Compounds for the prevention and treatment of noise-induced hearing loss. Drug Discov. Today 2005, 10, 1291–1298. [Google Scholar] [CrossRef]
- Lie, A.; Skogstad, M.; Johannessen, H.A.; Tynes, T.; Mehlum, I.S.; Nordby, K.C.; Engdahl, B.; Tambs, K. Occupational noise exposure and hearing: A systematic review. Int. Arch. Occup. Environ. Health 2016, 89, 351–372. [Google Scholar] [CrossRef]
- Liu, Y.D.; Wang, Z.B.; Han, G.; Zhao, P. Hyperbaric oxygen treatment attenuates neuropathic pain by elevating autophagy flux via inhibiting mTOR pathway. Am. J. Transl. Res. 2017, 9, 2629–2638. [Google Scholar]
- Katznelson, R.; Segal, S.C.; Clarke, H. Successful Treatment of Lower Limb Complex Regional Pain Syndrome following Three Weeks of Hyperbaric Oxygen Therapy. Pain Res. Manag. 2016, 2016, 3458371. [Google Scholar] [CrossRef]
- Efrati, S.; Golan, H.; Bechor, Y.; Faran, Y.; Daphna-Tekoah, S.; Sekler, G.; Fishlev, G.; Ablin, J.N.; Bergan, J.; Volkov, O.; et al. Hyperbaric oxygen therapy can diminish fibromyalgia syndrome-prospective clinical trial. PLoS ONE 2015, 10, e0127012. [Google Scholar] [CrossRef]
- Weaver, L. Hyperbaric Oxygen Therapy Indications, 13th ed.; Best Publishing Company: North Palm Beach, FL, USA, 2014. [Google Scholar]
- Appaix, A.; Demard, F. Hyperbaric oxygenotherapy and sudden perceptive deafness. Rev. Laryngol. Otol. Rhinol. 1970, 91, 951–972. [Google Scholar]
- Nagahara, K.; Fisch, U.; Yagi, N. Perilymph oxygenation in sudden and progressive sensorineural hearing loss. Acta Otolaryngol. 1983, 96, 57–68. [Google Scholar] [CrossRef]
- Melotte, G.; Parmentier, E.; Michel, C.; Herrel, A.; Boyle, K. Hearing capacities and morphology of the auditory system in Serrasalmidae (Teleostei: Otophysi). Sci. Rep. 2018, 8, 1281. [Google Scholar] [CrossRef]
- Hu, Q.; Manaenko, A.; Matei, N.; Guo, Z.; Xu, T.; Tang, J.; Zhang, J.H. Hyperbaric oxygen preconditioning: A reliable option for neuroprotection. Med. Gas. Res. 2016, 6, 20–32. [Google Scholar]
- Sanchez, E.C. Mechanisms of action of hyperbaric oxygenation in stroke: A review. Crit. Care Nurs. Q. 2013, 36, 290–298. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Grodzki, A.C.; Giulivi, C.; Lein, P.J. Oxygen tension modulates differentiation and primary macrophage functions in the human monocytic THP-1 cell line. PLoS ONE 2013, 8, e54926. [Google Scholar] [CrossRef]
- Law, B.A.; Liao, X.; Moore, K.S.; Southard, A.; Roddy, P.; Ji, R.; Szulc, Z.; Bielawska, A.; Schulze, P.C.; Cowart, L.A. Lipotoxic very-long-chain ceramides cause mitochondrial dysfunction, oxidative stress, and cell death in cardiomyocytes. FASEB J. 2018, 32, 1403–1416. [Google Scholar] [CrossRef]
- Matsunami, T.; Sato, Y.; Sato, T.; Ariga, S.; Shimomura, T.; Yukawa, M. Oxidative stress and gene expression of antioxidant enzymes in the streptozotocin-induced diabetic rats under hyperbaric oxygen exposure. Int. J. Clin. Exp. Pathol. 2009, 3, 177–188. [Google Scholar]
- Simsek, K.; Olzer, M.; Ucar, E. Pressure-related effects of hyperbaric oxygen exposure on oxidation products and antioxidant enzymes in the rat lung. J. Exp. Integr. Med. 2011, 1, 37–42. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Rossignol, L.W.; James, S.J.; Melnyk, S.; Mumper, E. The effects of hyperbaric oxygen therapy on oxidative stress, inflammation, and symptoms in children with autism: An open-label pilot study. BMC Pediatr. 2007, 7, 36. [Google Scholar] [CrossRef]
- Ozden, T.A.; Uzun, H.; Bohloli, M.; Toklu, A.S.; Paksoy, M.; Simsek, G.; Durak, H.; Issever, H.; Ipek, T. The effects of hyperbaric oxygen treatment on oxidant and antioxidants levels during liver regeneration in rats. Tohoku J. Exp. Med. 2004, 203, 253–265. [Google Scholar] [CrossRef]
- Rockswold, S.B.; Rockswold, G.L.; Zaun, D.A.; Liu, J. A prospective, randomized Phase II clinical trial to evaluate the effect of combined hyperbaric and normobaric hyperoxia on cerebral metabolism, intracranial pressure, oxygen toxicity, and clinical outcome in severe traumatic brain injury. J. Neurosurg. 2013, 118, 1317–1328. [Google Scholar] [CrossRef]
- Andrieu-Abadie, N.; Gouaze, V.; Salvayre, R.; Levade, T. Ceramide in apoptosis signaling: Relationship with oxidative stress. Free Radic. Biol. Med. 2001, 31, 717–728. [Google Scholar] [CrossRef]
- Cutler, R.G.; Kelly, J.; Storie, K.; Pedersen, W.A.; Tammara, A.; Hatanpaa, K.; Troncoso, J.C.; Mattson, M.P. Involvement of oxidative stress-induced abnormalities in ceramide and cholesterol metabolism in brain aging and Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 2070–2075. [Google Scholar] [CrossRef]
- Kang, M.S.; Ahn, K.H.; Kim, S.K.; Jeon, H.J.; Ji, J.E.; Choi, J.M.; Jung, K.M.; Jung, S.Y.; Kim, D.K. Hypoxia-induced neuronal apoptosis is mediated by de novo synthesis of ceramide through activation of serine palmitoyltransferase. Cell. Signal. 2010, 22, 610–618. [Google Scholar] [CrossRef]
- Kocot, J.; Luchowska-Kocot, D.; Kielczykowska, M.; Musik, I.; Kurzepa, J. Does Vitamin C Influence Neurodegenerative Diseases and Psychiatric Disorders? Nutrients 2017, 9, 659. [Google Scholar] [CrossRef]
- Realini, N.; Solorzano, C.; Pagliuca, C.; Pizzirani, D.; Armirotti, A.; Luciani, R.; Costi, M.P.; Bandiera, T.; Piomelli, D. Discovery of highly potent acid ceramidase inhibitors with in vitro tumor chemosensitizing activity. Sci. Rep. 2013, 3, 1035. [Google Scholar] [CrossRef]
- Diehl, K.H.; Hull, R.; Morton, D.; Pfister, R.; Rabemampianina, Y.; Smith, D.; Vidal, J.M.; van de Vorstenbosch, C. A good practice guide to the administration of substances and removal of blood, including routes and volumes. J. Appl. Toxicol. 2001, 21, 15–23. [Google Scholar] [CrossRef]
- El-Gendy, K.S.; Aly, N.M.; Mahmoud, F.H.; Kenawy, A.; El-Sebae, A.K.H. The role of vitamin C as antioxidant in protection of oxidative stress induced by imidacloprid. Food Chem. Toxicol. 2010, 48, 215–221. [Google Scholar] [CrossRef]
- Rabelo, L.M.; Estrela, F.N.; Silva, B.C.; Mendes, B.O.; Vaz, B.G.; Rodrigues, A.S.L.; Malafaia, G. Protective effect of vitamin C in female Swiss mice dermally-exposed to the tannery effluent. Chemosphere 2017, 181, 492–499. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.-T.; Guo, Y.-B.; Cheng, Y.-P.; Zhang, X.; Xie, X.-P.; Chang, Y.-M.; Bao, J.-X. Hyperbaric Oxygen Treatment Ameliorates Hearing Loss and Auditory Cortex Injury in Noise Exposed Mice by Repressing Local Ceramide Accumulation. Int. J. Mol. Sci. 2019, 20, 4675. https://doi.org/10.3390/ijms20194675
Su Y-T, Guo Y-B, Cheng Y-P, Zhang X, Xie X-P, Chang Y-M, Bao J-X. Hyperbaric Oxygen Treatment Ameliorates Hearing Loss and Auditory Cortex Injury in Noise Exposed Mice by Repressing Local Ceramide Accumulation. International Journal of Molecular Sciences. 2019; 20(19):4675. https://doi.org/10.3390/ijms20194675
Chicago/Turabian StyleSu, Yu-Ting, Yi-Bin Guo, Yao-Ping Cheng, Xi Zhang, Xiao-Ping Xie, Yao-Ming Chang, and Jun-Xiang Bao. 2019. "Hyperbaric Oxygen Treatment Ameliorates Hearing Loss and Auditory Cortex Injury in Noise Exposed Mice by Repressing Local Ceramide Accumulation" International Journal of Molecular Sciences 20, no. 19: 4675. https://doi.org/10.3390/ijms20194675
APA StyleSu, Y.-T., Guo, Y.-B., Cheng, Y.-P., Zhang, X., Xie, X.-P., Chang, Y.-M., & Bao, J.-X. (2019). Hyperbaric Oxygen Treatment Ameliorates Hearing Loss and Auditory Cortex Injury in Noise Exposed Mice by Repressing Local Ceramide Accumulation. International Journal of Molecular Sciences, 20(19), 4675. https://doi.org/10.3390/ijms20194675