Spatiotemporal Expression of Anticoagulation Factor Antistasin in Freshwater Leeches

Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Phylogeny of Antistasin Orthologs in Serine Protease Inhibitor Groups
2.2. Temporal Expression of Hau-Antistasin Genes during Embryogenesis
2.3. Spatial Expression of Hau-Antistasin during Organogenesis
2.4. Co-Linearity and Diversity of Antistasin Orthologs in Glossiphoniidae Leeches
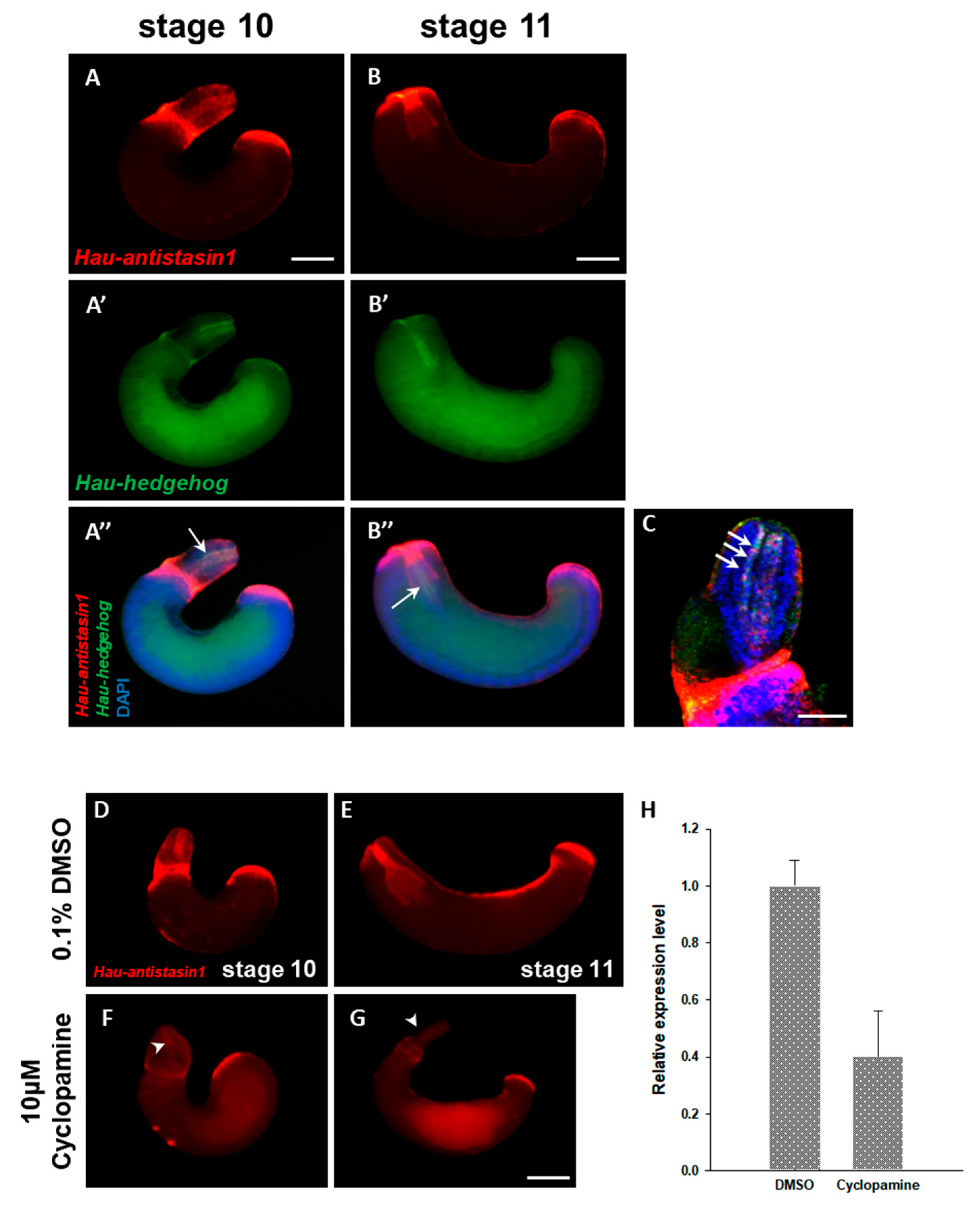
2.5. Hau-Antistasin1 is Regulated by Hedgehog (hh) Signaling
3. Materials and Methods
3.1. Animals
3.2. Phylogenetic Analysis and Domain Prediction
3.3. Gene Identification, Gene Cloning, and Probe Synthesis
3.4. Developmental Semiquantitative RT-PCR
3.5. Whole-mount In Situ Hybridization and Cross Sectioning
3.6. Drug treatments and qPCR
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koh, C.Y.; Kini, R.M. Anticoagulants from hematophagous animals. Expert Rev. Hematol. 2008, 1, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Kvist, S.; Sarkar, I.N.; Siddall, M.E. Genome-wide search for leech antiplatelet proteins in the non-blood-feeding leech H elobdella robusta (R hyncobdellida: G lossiphoniidae) reveals evidence of secreted anticoagulants. Invertebr. Biol. 2011, 130, 344–350. [Google Scholar] [CrossRef]
- Salzet, M. Anticoagulants and inhibitors of platelet aggregation derived from leeches. FEBS Lett. 2001, 492, 187–192. [Google Scholar] [CrossRef]
- Dunwiddie, C.; Thornberry, N.; Bull, H.; Sardana, M.; Friedman, P.; Jacobs, J.; Simpson, E. Antistasin, a leech-derived inhibitor of factor Xa. Kinetic analysis of enzyme inhibition and identification of the reactive site. J. Biol. Chem. 1989, 264, 16694–16699. [Google Scholar] [PubMed]
- Nutt, E.; Gasic, T.; Rodkey, J.; Gasic, G.; Jacobs, J.W.; Friedman, P.A.; Simpson, E. The amino acid sequence of antistasin. A potent inhibitor of factor Xa reveals a repeated internal structure. J. Biol. Chem. 1988, 263, 10162–10167. [Google Scholar] [PubMed]
- Tuszynski, G.; Gasic, T.; Gasic, G. Isolation and characterization of antistasin. An inhibitor of metastasis and coagulation. J. Biol. Chem. 1987, 262, 9718–9723. [Google Scholar] [PubMed]
- Lee, M.S.; Tak, E.S.; Park, S.K.; Cho, S.J.; Hahn, Y.; Joo, S.S.; Lee, D.I.; Ahn, C.H.; Park, S.C. Eisenstasin, new antistasin family inhibitor from the earthworm. Biologia 2010, 65, 284–288. [Google Scholar] [CrossRef]
- Holstein, T.W.; Mala, C.; Kurz, E.; Bauer, K.; Greber, M.; David, C.N. The primitive metazoan Hydra expresses antistasin, a serine protease inhibitor of vertebrate blood coagulation: cDNA cloning, cellular localisation and developmental regulation. FEBS Lett. 1992, 309, 288–292. [Google Scholar] [CrossRef]
- Nikapitiya, C.; De Zoysa, M.; Oh, C.; Lee, Y.; Ekanayake, P.M.; Whang, I.; Choi, C.Y.; Lee, J.S.; Lee, J. Disk abalone (Haliotis discus discus) expresses a novel antistasin-like serine protease inhibitor: Molecular cloning and immune response against bacterial infection. Fish Shellfish Immunol. 2010, 28, 661–671. [Google Scholar] [CrossRef]
- Kutschera, U.; Langguth, H.; Kuo, D.H.; Weisblat, D.; Shankland, M. Description of a new leech species from North America, Helobdella austinensis n. sp. Hirudinea: Glossiphoniidae), with observations on its feeding behaviour. Zoosystematics Evol. 2013, 89, 239–246. [Google Scholar] [CrossRef]
- Sawyer, R.; Casellas, M.; Munro, R.; Jones, C.P. Secretion of hementin and other antihaemostatic factors in the salivary gland complex of the giant Amazon leech Haementeria ghilianii. Comp. Haematol. Int. 1991, 1, 35–41. [Google Scholar] [CrossRef]
- Faria, F.; Junqueira-de-Azevedo Ide, I.; Ho, P.L.; Sampaio, M.U.; Chudzinski-Tavassi, A.M. Gene expression in the salivary complexes from Haementeria depressa leech through the generation of expressed sequence tags. Gene 2005, 349, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Valles, Y.; Weisblat, D.A. Differential expression of conserved germ line markers and delayed segregation of male and female primordial germ cells in a hermaphrodite, the leech Helobdella. Mol. Biol. Evol. 2013, 31, 341–354. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kuo, D.H.; Hsiao, Y.H. Duplicated FoxA genes in the leech Helobdella: Insights into the evolution of direct development in clitellate annelids. Dev. Dyn. 2018, 247, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Weisblat, D.A.; Huang, F.Z. An overview of glossiphoniid leech development. Can. J. Zool. 2001, 79, 218–232. [Google Scholar] [CrossRef]
- Lu, Z.; Shi, P.; You, H.; Liu, Y.; Chen, S. Transcriptomic analysis of the salivary gland of medicinal leech Hirudo nipponia. Plos ONE 2018, 13, e0205875. [Google Scholar] [CrossRef] [PubMed]
- Min, G.S.; Sarkar, I.N.; Siddall, M.E. Salivary transcriptome of the North American medicinal leech, Macrobdella decora. J. Parasitol. 2010, 96, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Babenko, V.V.; Podgorny, O.V.; Manuvera, V.A.; Kasianov, A.S.; Manolov, A.I.; Grafskaia, E.N.; Shirokov, D.A.; Kurdyumov, A.S.; Vinogradov, D.V.; Nikitina, A.S. Far beyond common leeching: Insights into an ancient medical device through integrated omics data. bioRxiv 2018, 357681. [Google Scholar] [CrossRef]
- Gline, S.E.; Nakamoto, A.; Cho, S.J.; Chi, C.; Weisblat, D.A. Lineage analysis of micromere 4d, a super-phylotypic cell for Lophotrochozoa, in the leech Helobdella and the sludgeworm Tubifex. Dev. Biol. 2011, 353, 120–133. [Google Scholar] [CrossRef]
- Kang, D.; Huang, F.; Li, D.; Shankland, M.; Gaffield, W.; Weisblat, D.A. A hedgehog homolog regulates gut formation in leech (Helobdella). Development 2003, 130, 1645–1657. [Google Scholar] [CrossRef]
- Kim, J.S.; Jiménez, B.I.M.; Kwak, H.J.; Park, S.C.; Xiao, P.; Weisblat, D.A.; Cho, S.J. Spatiotemporal expression of a twist homolog in the leech Helobdella austinensis. Dev. Genes Evol. 2017, 227, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kwak, H.J.; Jiménez, B.I.M.; Park, S.C.; Xiao, P.; Weisblat, D.A.; Cho, S.J. Expression patterns of duplicated snail genes in the leech Helobdella. Dev. Genes Evol. 2017, 227, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Bely, A.E.; Weisblat, D.A. Lessons from leeches: A call for DNA barcoding in the lab. Evol. Dev. 2006, 8, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Kuo, D.H.; Weisblat, D.A. Intermediate filament genes as differentiation markers in the leech Helobdella. Dev. Genes Evol. 2011, 221, 225. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, B.I.M.; Kwak, H.J.; Park, J.S.; Kim, J.W.; Cho, S.J. Developmental biology and potential use of Alboglossiphonia lata (Annelida: Hirudinea) as an “Evo-Devo” model organism. Front. Zool. 2017, 14, 60. [Google Scholar] [CrossRef]
- Kuo, D.H.; Weisblat, D.A. A new molecular logic for BMP-mediated dorsoventral patterning in the leech Helobdella. Curr. Biol. 2011, 21, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.J.; Ryu, K.B.; Medina Jiménez, B.I.; Park, S.C.; Cho, S.J. Temporal and spatial expression of the Fox gene family in the Leech Helobdella austinensis. J. Exp. Zool. Part. B: Mol. Dev. Evol. 2018, 330, 341–350. [Google Scholar] [CrossRef]
- Cho, S.J.; Vallès, Y.; Giani Jr, V.C.; Seaver, E.C.; Weisblat, D.A. Evolutionary dynamics of the wnt gene family: A lophotrochozoan perspective. Mol. Biol. Evol. 2010, 27, 1645–1658. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, H.-J.; Park, J.-S.; Medina Jiménez, B.I.; Park, S.C.; Cho, S.-J. Spatiotemporal Expression of Anticoagulation Factor Antistasin in Freshwater Leeches. Int. J. Mol. Sci. 2019, 20, 3994. https://doi.org/10.3390/ijms20163994
Kwak H-J, Park J-S, Medina Jiménez BI, Park SC, Cho S-J. Spatiotemporal Expression of Anticoagulation Factor Antistasin in Freshwater Leeches. International Journal of Molecular Sciences. 2019; 20(16):3994. https://doi.org/10.3390/ijms20163994
Chicago/Turabian StyleKwak, Hee-Jin, Jeong-Su Park, Brenda Irene Medina Jiménez, Soon Cheol Park, and Sung-Jin Cho. 2019. "Spatiotemporal Expression of Anticoagulation Factor Antistasin in Freshwater Leeches" International Journal of Molecular Sciences 20, no. 16: 3994. https://doi.org/10.3390/ijms20163994
APA StyleKwak, H.-J., Park, J.-S., Medina Jiménez, B. I., Park, S. C., & Cho, S.-J. (2019). Spatiotemporal Expression of Anticoagulation Factor Antistasin in Freshwater Leeches. International Journal of Molecular Sciences, 20(16), 3994. https://doi.org/10.3390/ijms20163994