Gene-Wide Analysis of Aquaporin Gene Family in Malus domestica and Heterologous Expression of the Gene MpPIP2;1 Confers Drought and Salinity Tolerance in Arabidposis thaliana

Abstract
1. Introduction
2. Results
2.1. The Family of AQPs Encoded by the Apple Genome
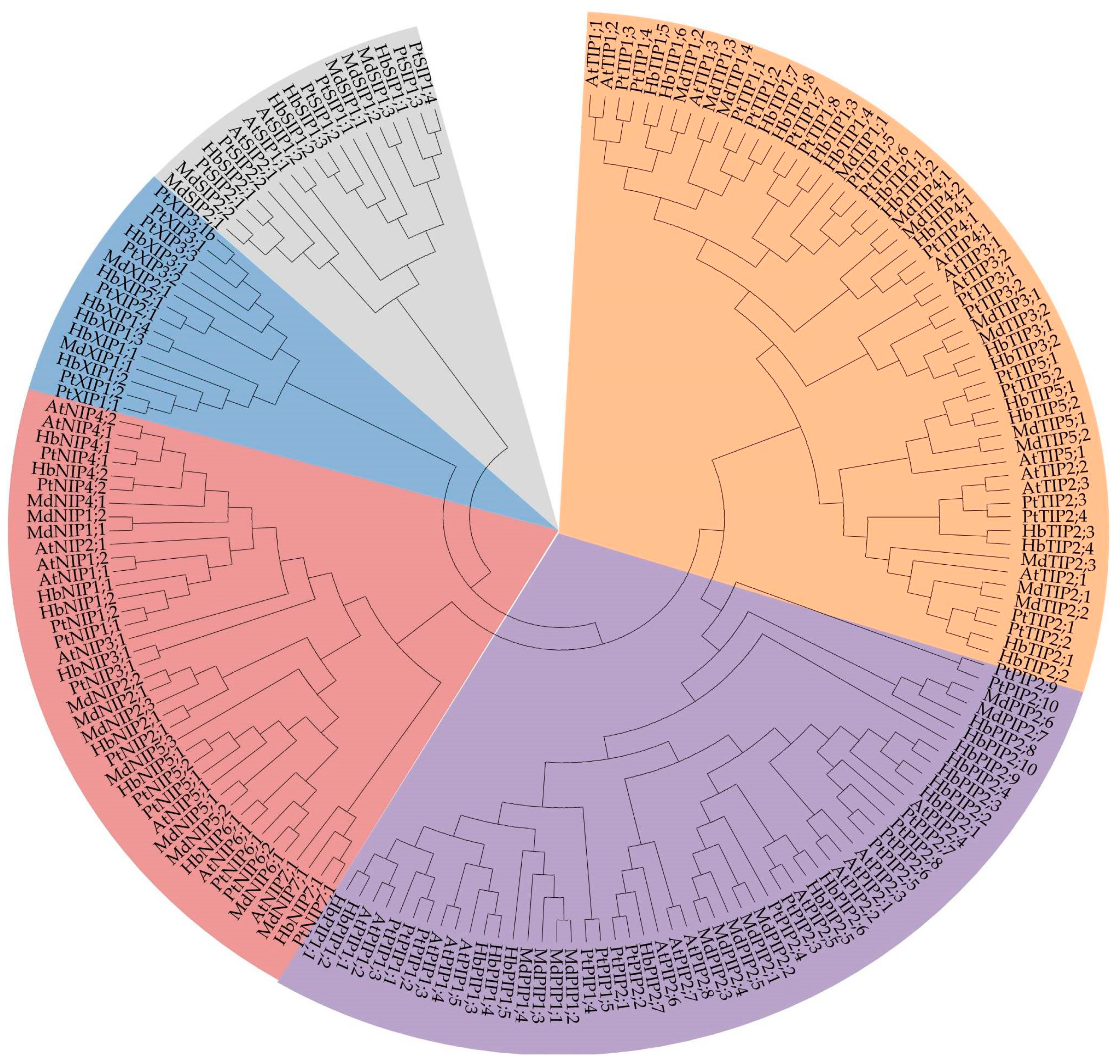
2.2. Phylogenetic Analysis
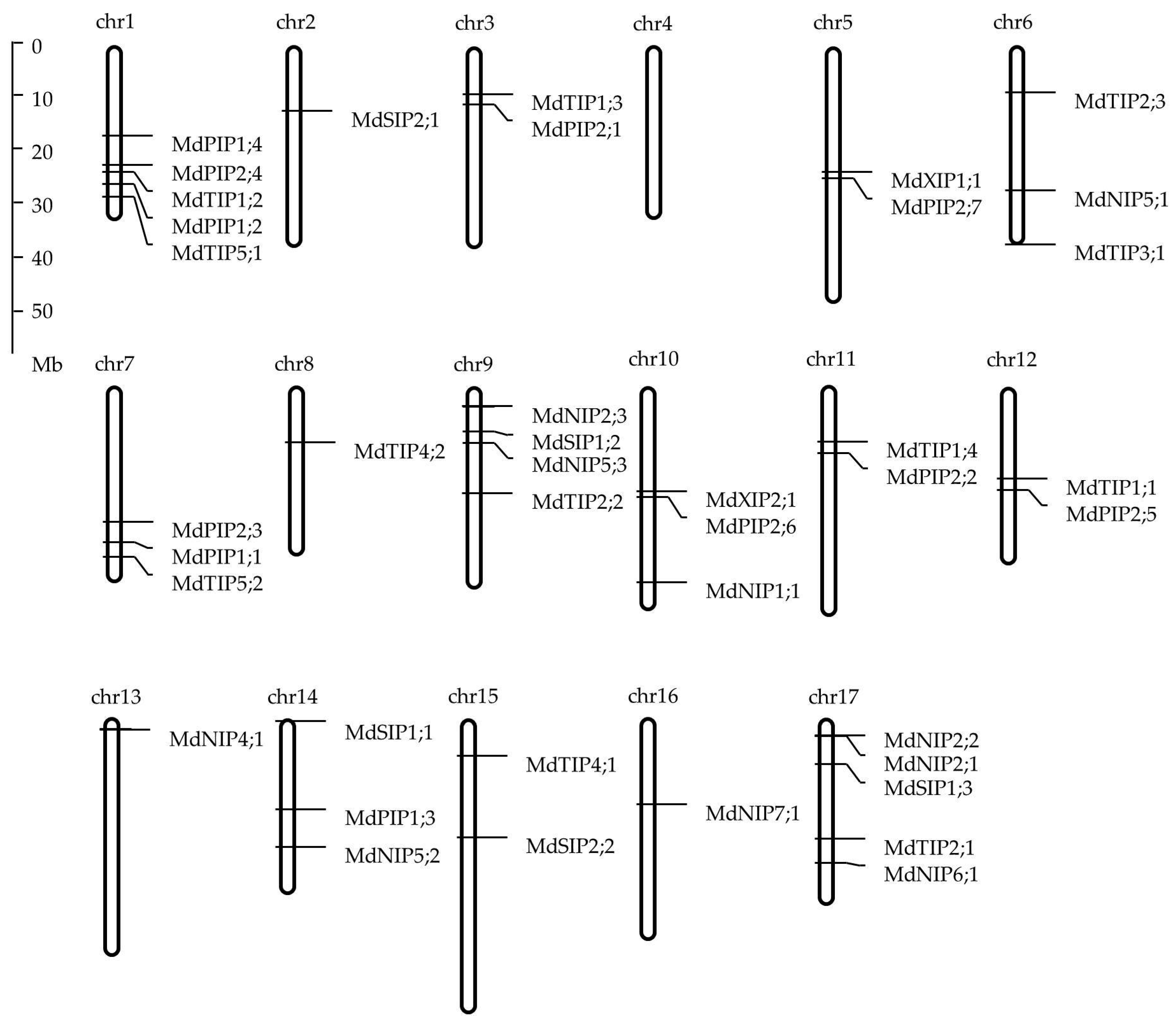
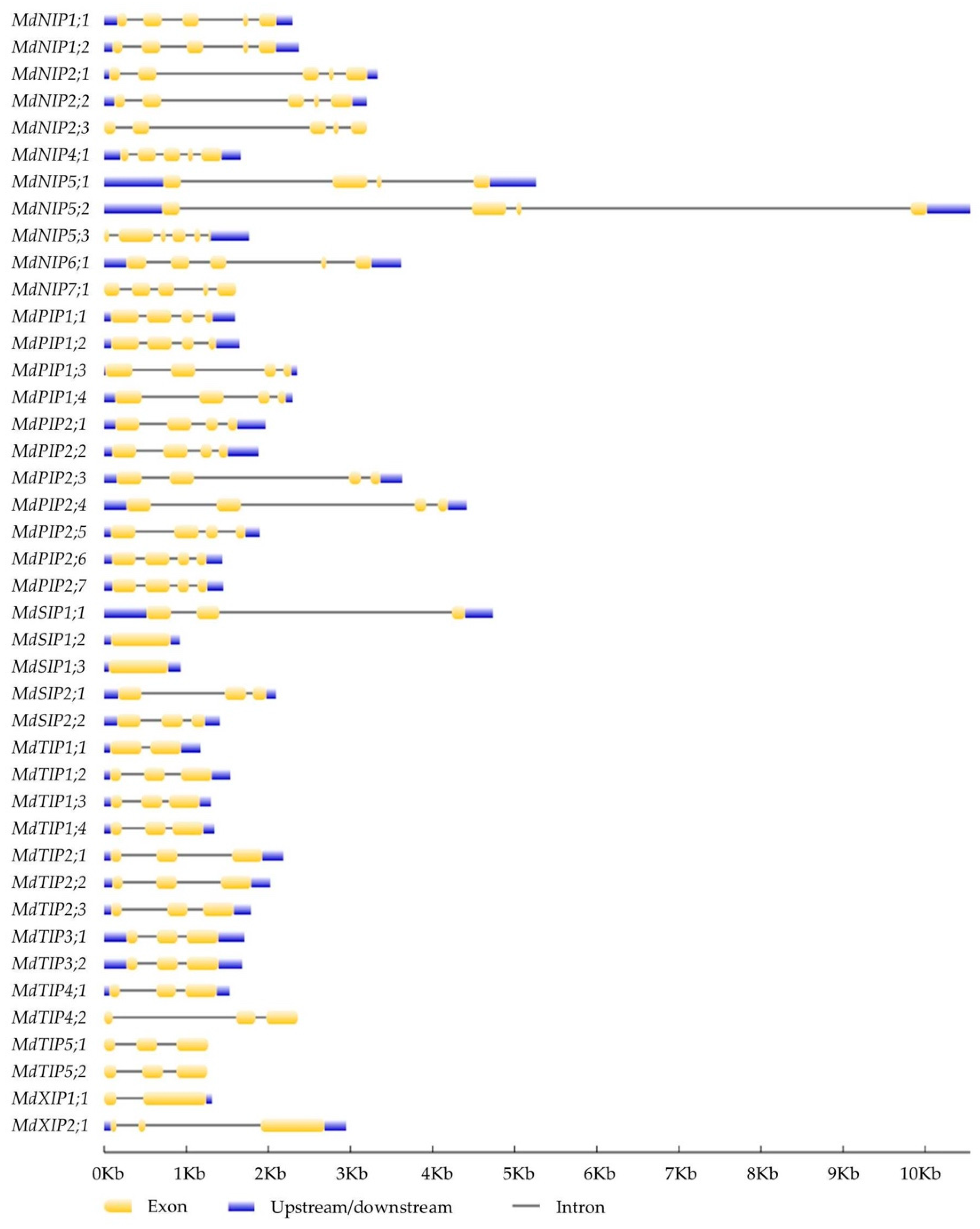
2.3. Chromosomal Location and Gene Structure
2.4. Conserved Residues in the Apple AQPs
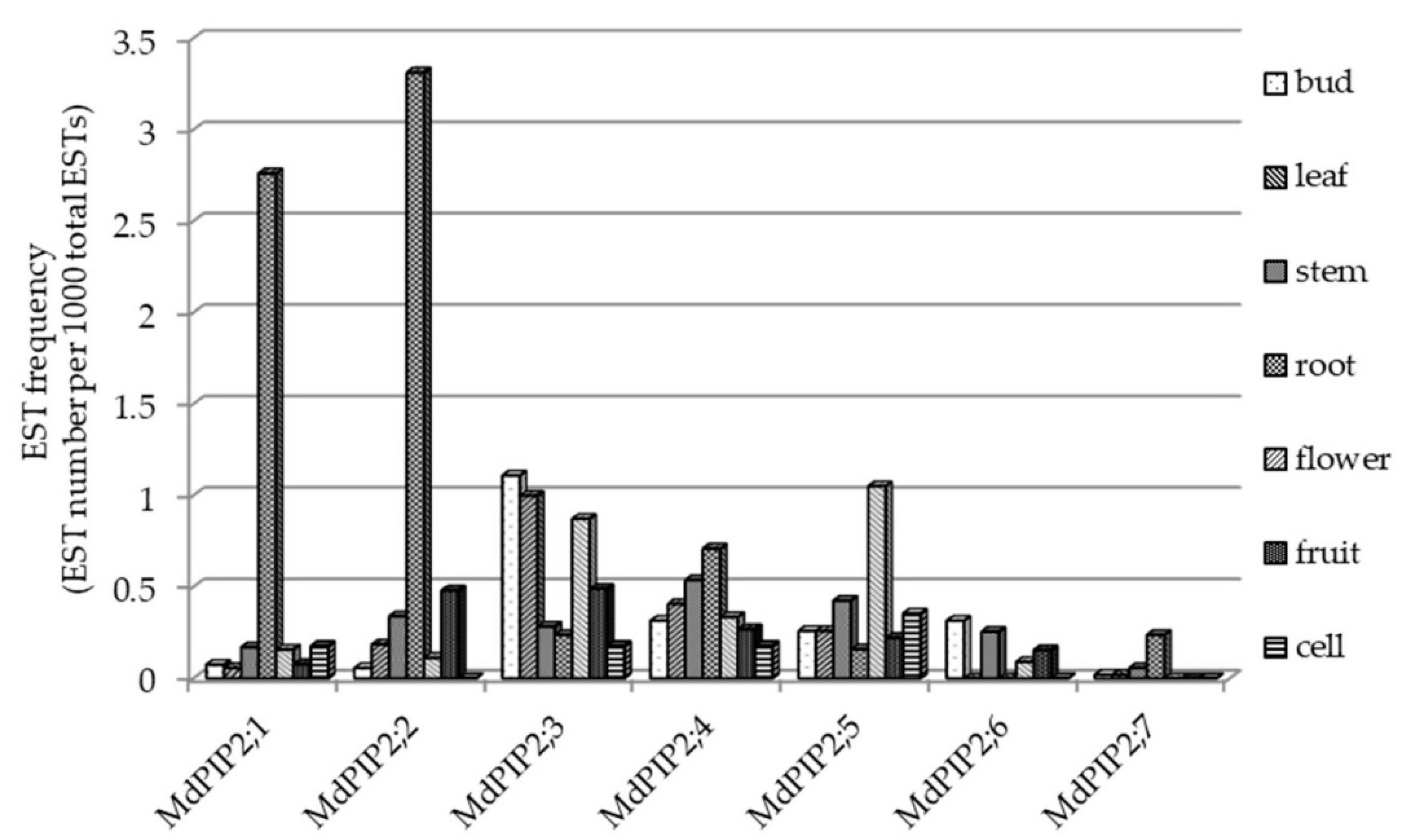
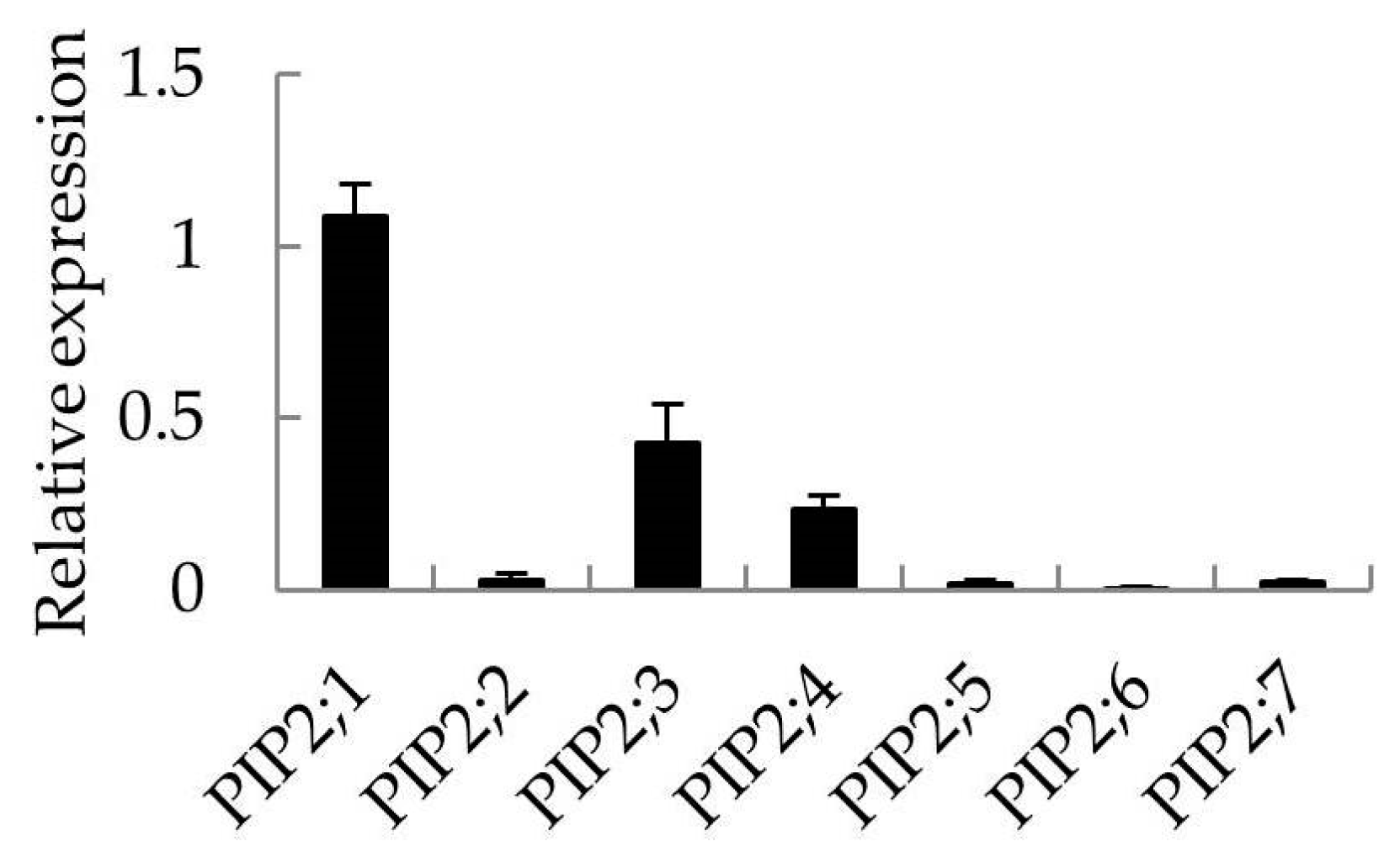
2.5. The Site of Apple PIP2 Expression
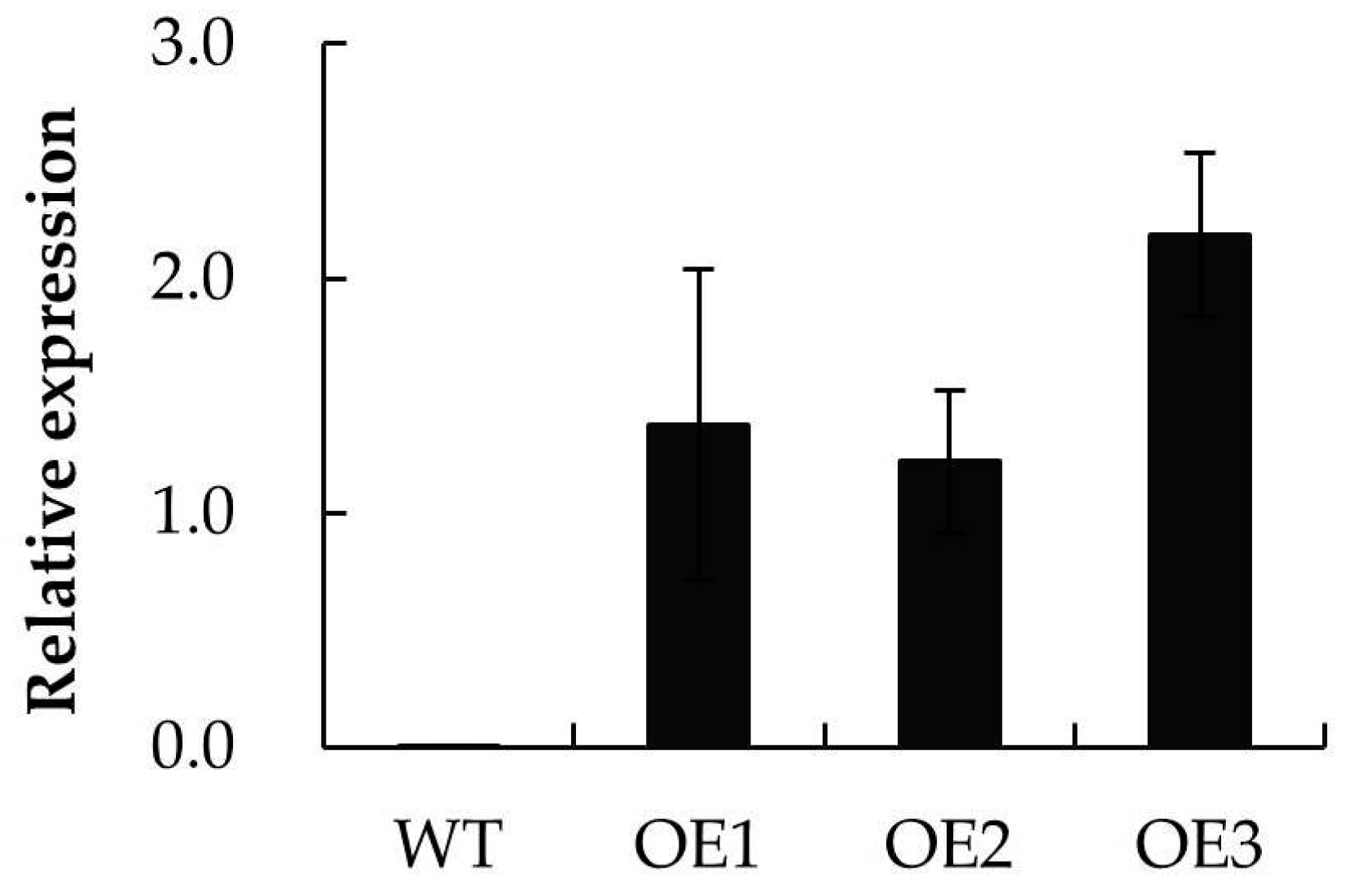
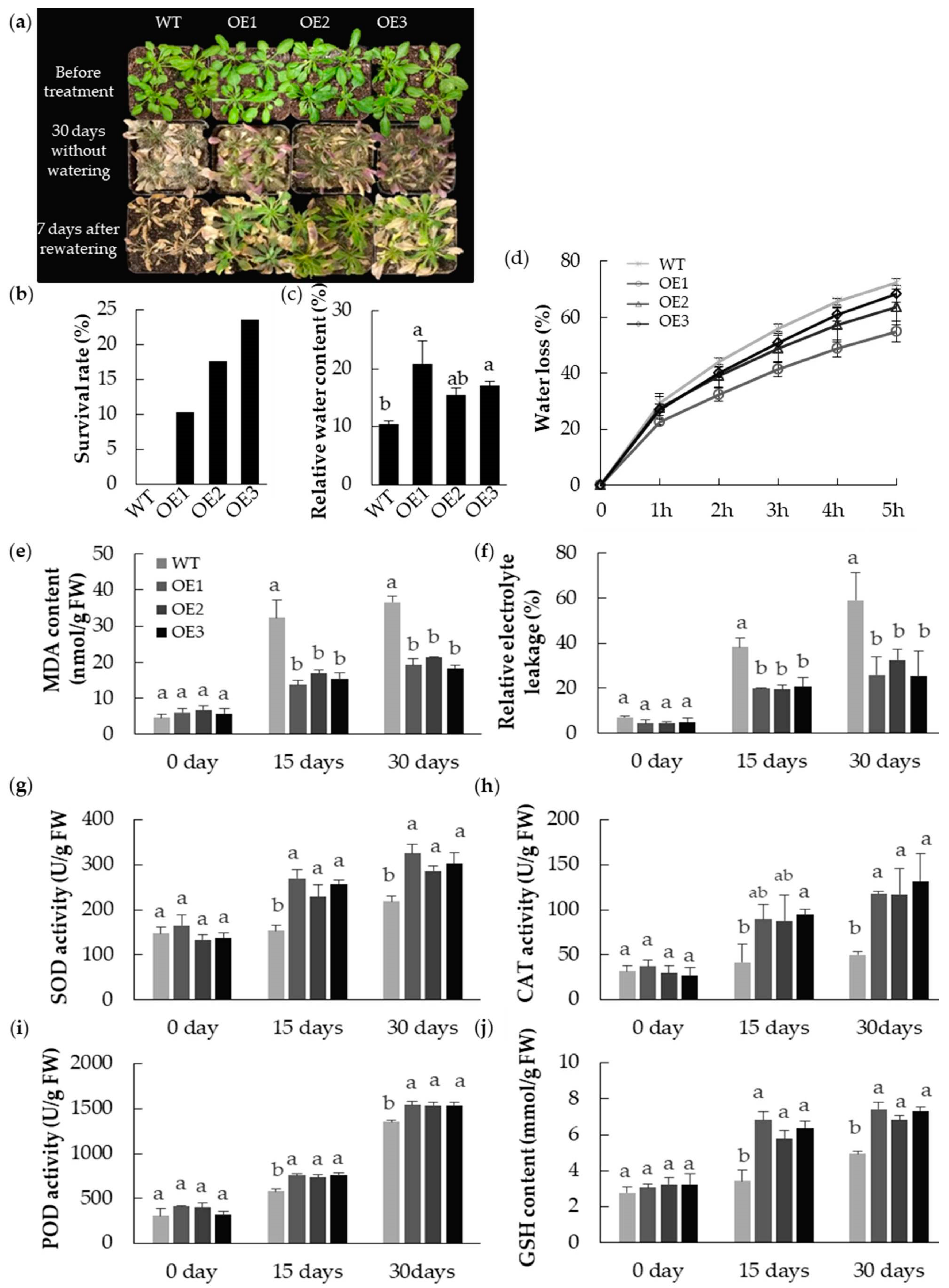
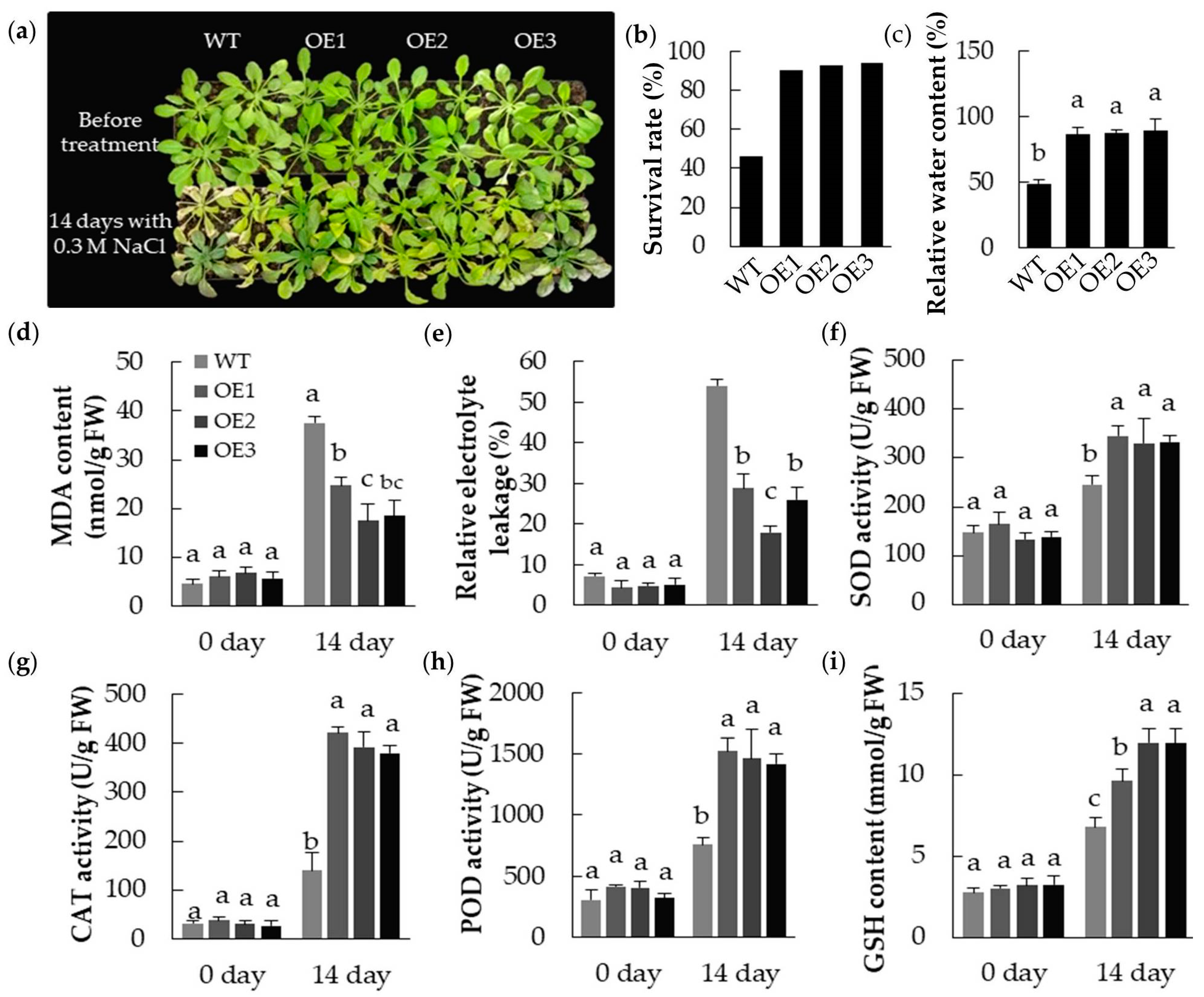
2.6. The Abiotic Stress Tolerance of A. thaliana Plants Heterologously Expressing MpPIP2;1
2.7. Germination and Root Elongation of MpPIP2;1 Transgenics Exposed to Either Salinity or Osmotic Stress
3. Discussion
4. Materials and Methods
4.1. Identification of the Set of AQP Genes in the Apple Genome
4.2. Chromosomal Location, Gene Structure, and Protein Properties of Apple AQPs
4.3. The Sites of Apple PIP2 Expression
4.4. Heterologous Expression of MpPIP2;1 in A. thaliana
4.5. Stress Tolerance Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AQP | aquaporins |
| NPA | asparagine-proline-alanine |
| ar/R | aromatic/arginine |
| NIP | Nod26-like intrinsic proteins |
| PIP | plasma membrane intrinsic proteins |
| SIP | small and basic intrinsic proteins |
| TIP | tonoplast intrinsic proteins |
| XIP | uncharacterized intrinsic proteins |
| MDA | malondialdehyde |
| SOD | superoxide dismutase |
| CAT | catalase |
| POD | peroxidase |
| GSH | glutathione |
References
- Hove, R.M.; Bhave, M. Plant aquaporins with non-aqua functions: Deciphering the signature sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. Biochim. Biophys. Acta. 2014, 1840, 1468–1481. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.X.; Han, B.G.; Lee, J.K.; Walian, P.; Jap, B.K. Structural basis of water-specific transport through the AQP1 water channel. Nature 2001, 414, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Heymann, J.B.; Engel, A. Structural clues in the sequences of the aquaporins. J. Mol. Biol. 2000, 295, 1039–1053. [Google Scholar] [CrossRef] [PubMed]
- Kreida, S.; Tornroth-Horsefield, S. Structural insights into aquaporin selectivity and regulation. Curr. Opin. Struct. Biol. 2015, 33, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Kshetrinayum, E.; Sahoo, D.P.; Mitra, J.; Panda, S.K. Regulation of seed germination and the role of aquaporins under abiotic stress. Int. J. Agric. Biol. 2017, 2, 607–615. [Google Scholar] [CrossRef]
- Eisenbarth, D.A.; Weig, A.R. Dynamics of aquaporins and water relations during hypocotyl elongation in Ricinus communis L. seedlings. J. Exp. Bot. 2005, 56, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Ribas-Carbo, M.; Hanson, D.T.; Bota, J.; Otto, B.; Cifre, J.; McDowell, N.; Medrano, H.; Kaldenhoff, R. Tobacco aquaporin NtAQP1 is involved in mesophyll conductance to CO2 in vivo. Plant J. 2006, 48, 427–439. [Google Scholar] [CrossRef]
- Grondin, A.; Rodrigues, O.; Verdoucq, L.; Merlot, S.; Leonhardt, N.; Maurel, C. Aquaporins contribute to ABA-triggered stomatal closure through OST1-mediated phosphorylation. Plant Cell. 2015, 27, 1945–1954. [Google Scholar] [CrossRef]
- Mut, P.; Bustamane, C.; Martinez, G.; Alleva, K.; Sutka, M.; Civello, M.; Amodeo, G. A fruit-specific plasma membrane aquaporin subtype PIP1;1 is regulated druing strawberry (fragaria × ananassa) fruit ripening. Physiol. Plant. 2008, 132, 538–551. [Google Scholar] [CrossRef]
- Molina-Hidalgo, J.F.; Medina-Puche, L.; Gelis, S.; Ramos, J. Functional characterization of FaNIP1;1 gene, a ripening-related and receptacle-specific aquaporin in strawberry fruit. Plant Sci 2015, 238, 198–211. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, S.H.; Xiang, K.; Wang, C.T.; Xiang, N.; Yang, Y.Q.; Yang, Y.P. Molecular cloning of a plasma membrane aquaporin in Stipa purpurea, and exploration of its role in drought stress tolerance. Gene 2018, 665, 41–48. [Google Scholar] [CrossRef]
- Li, W.; Qiang, X.J.; Han, R.X.; Jiang, L.L.; Zhang, S.H.; Han, J.; He, R.; Cheng, X.G. Ectopic Expression of a Thellungiella salsuginea aquaporin gene, TsPIP1;1, increased the salt tolerance of rice. Int. J. Mol. Sci. 2018, 19, 2229. [Google Scholar] [CrossRef]
- Matsumoto, T.; Lian, H.L.; Su, W.A.; Tanaka, D.; Liu, C.W.; Iwasaki, I.; Kitagawa, Y. Role of the aquaporin PIP1 subfamily in the chilling tolerance of rice. Plant Cell Physiol. 2009, 50, 216–229. [Google Scholar] [CrossRef]
- Shekoofa, A.; Sinclair, T.R. Aquaporin activity to improve crop drought tolerance. Cells 2018, 7, 123. [Google Scholar] [CrossRef]
- Zhang, S.; Feng, M.; Chen, W.; Zhou, X.; Lu, J.; Wang, Y.; Li, Y.; Jiang, C.Z.; Gan, S.S.; Ma, N.; et al. In rose, transcription factor PTM balances growth and drought survival via PIP2;1 aquaporin. Nat. Plants 2019, 5, 290–299. [Google Scholar] [CrossRef]
- Johanson, U.; Karlsson, M.; Johansson, I.; Gustavsson, S.; Sjövall, S.; Fraysse, L.; Laure Fraysse, L.; Weig, A.R.; Kjellbom, P. The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol. 2001, 126, 1358–1369. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guerin, V.; Sonah, H.; Labbe, C.; Belzile, F.; Belanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Gupta, A.B.; Sankararamakrishnan, R. Genome-wide analysis of major intrinsic proteins in the tree plant Populus trichocarpa: Characterization of XIP subfamily of aquaporins from evolutionary perspective. BMC Plant Biol. 2009, 9, 134. [Google Scholar] [CrossRef]
- Danielson, J.A.; Johanson, U. Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens. BMC Plant Biol. 2008, 8, 45. [Google Scholar] [CrossRef]
- Fox, A.R.; Maistriaux, L.C.; Chaumont, F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant Sci 2017, 264, 179–187. [Google Scholar] [CrossRef]
- Wang, S.C.; Liang, D.; Li, C.; Hao, Y.L.; Ma, F.W.; Shu, H.R. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks. Plant Physiol. Biochem. 2012, 51, 81–97. [Google Scholar] [CrossRef]
- Li, J.; Ban, L.; Wen, H.Y.; Wang, Z.; Dzyubenko, N.; Chapurin, V.; Gao, H.W.; Wang, X.M. An aquaporin protein is associated with drought stress tolerance. Biochem. Biophys. Res. Commun. 2015, 459, 208–213. [Google Scholar] [CrossRef]
- Hu, C.G.; Hao, Y.J.; Honda, C.; Kita, M.; Moriguchi, T. Putative PIP1 genes isolated from apple: Expression analyses during fruit development and under osmotic stress. J. Exp. Bot. 2003, 54, 2193–2194. [Google Scholar] [CrossRef]
- Li, Q.T.; Zhao, Y.; Zhou, Q.Y.; Huang, L.F.; Jiang, Y.Z.; Chen, H.; Kong, J. Cloning and expression analysis of MzPIP1;1 gene from Malus zumi Mats. J.China Agric. Univ. 2012, 17, 63–68. [Google Scholar]
- Wang, L.; Li, Q.T.; Lei, Q.; Feng, C.; Gao, Y.N.; Zheng, X.D.; Zhao, Y.; Wang, Z.; Kong, J. MzPIP2;1: An aquaporin involved in Radial water movement in both water uptake and transportation, altered the drought and salt tolerance of transgenic Arabidopsis. PLoS ONE 2015, 10, e0142446. [Google Scholar] [CrossRef]
- Basssett, C.L.; Baldo, A.M.; Moore, J.T.; Jenkins, R.M.; Soffe, D.S.; Wisniewski, M.E.; Norelli, J.L.; Farrell, R.E., Jr. Genes responding to water deficit in apple (Malus X domestica Borkh.) roots. BMC Plant Bio. 2014, 14, 182–192. [Google Scholar] [CrossRef]
- Wang, L.; Li, Q.T.; Lei, Q.; Feng, C.; Zheng, X.D.; Zhou, F.F.; Li, L.Z.; Liu, X.; Wang, Z.; Kong, J. Ectopically expressing MdPIP1;3, an aquaporin gene, increased fruit size and enhanced drought tolerance of transgenic tomatoes. BMC Plant Biol. 2017, 17, 246–258. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends Plant Sci 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Wang, L.Q.; Zhang, C.R.; Wang, Y.M.; Wang, Y.C.; Yang, C.P.; Lu, M.Z.; Wang, C. Tamarix hispida aquaporin ThPIP2;5 confers salt and osmotic stress tolerance to transgenic Tamarix and Arabidopsis. Environ. Exp. Bot. 2018, 152, 158–166. [Google Scholar] [CrossRef]
- Zou, Z.; Gong, J.; An, F.; Xie, G.S.; Wang, J.K.; Mo, Y.Y.; Yang, L.F. Genome-wide identification of rubber tree (Hevea brasiliensis Muell. Arg.) aquaporin genes and their response to ethephon stimulation in the laticifer, a rubber producing tissue. BMC Genomics 2015, 16, 1001. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choic. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.W.; Liang, D.; Liu, C.H.; Yin, L.H.; Jia, D.F.; Fu, M.Y.; Ma, F.W. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Yao, P.F.; Li, C.L.; Zhao, X.R.; Li, M.F.; Zhao, H.X.; Guo, J.Y.; Cai, Y.; Chen, H.; Wu, W. Overexpression of a tartary buchwheat gene, FtbHLH3, enhances drought/oxidative stress tolerance in transgenic Arabidopsis. Front Plant Sci. 2017, 8, 625–641. [Google Scholar] [CrossRef]
- Liu, C.H.; Li, C.; Liang, D.; Ma, F.W.; Wang, S.C.; Wang, P.; Wang, R.W. Aquaporin expression in response to water-deficit stress in two Malus species: Relationship with physiological status and drought tolerance. Plant Growth Regul. 2013, 70, 187–197. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dong, Q.L.; Duan, D.Y.; Zhao, S.; Xu, B.Y.; Luo, J.W.; Wang, Q.; Liu, C.H.; Li, C.; Gong, X.Q.; Mao, K.; et al. Genome-wide analysis and cloning of the apple stress-associated protein gene family reveals MdSAP15, which confers tolerance to drought and osmotic stresses in transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2478. [Google Scholar] [CrossRef]
- Hou, H.M.; Jia, H.; Yan, Q.; Wang, X.P. Overexpression of a SBP-Box Gene (VpSBP16) from Chinese Wild Vitis Species in Arabidopsis Improves Salinity and Drought Stress Tolerance. Int. J. Mol. Sci. 2018, 19, 940. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | GeneBank accession no. | Gene ID | Size(aa) | MW(Da) | pI | TMD | Loc |
|---|---|---|---|---|---|---|---|
| MdNIP1;1 | XP_008383590.1 | 271 | 28742.49 | 9.03 | 6 | Plas | |
| MdNIP1;2 | XP_028954386.1 | 271 | 28743.38 | 8.99 | 6 | Plas | |
| MdNIP2;1 | XP_008341577.1 | 290 | 30456.02 | 6.70 | 6 | Plas | |
| MdNIP2;2 | XP_008341755.2 | 290 | 30513.07 | 6.70 | 6 | Plas | |
| MdNIP2;3 | MD09G1051500 1 | 260 | 27306.65 | 6.65 | 6 | Plas | |
| MdNIP4;1 | MD13G1025400 1 | 278 | 29945.16 | 8.97 | 7 | Plas | |
| MdNIP5;1 | XP_008356814.1 | 298 | 30960.84 | 8.94 | 5 | Vacu | |
| MdNIP5;2 | XP_008360732.1 | 298 | 31052.15 | 8.85 | 6 | Vacu | |
| MdNIP5;3 | XP_008348330.2 | 266 | 27575.92 | 9.23 | 5 | Plas | |
| MdNIP6;1 | XP_008343681.1 | 306 | 31641.74 | 8.90 | 6 | Vacu | |
| MdNIP7;1 | XP_028952006.1 | 300 | 31457.78 | 8.05 | 6 | Plas | |
| MdPIP1;1 | NP_001280914.1 | 289 | 30875.87 | 9.32 | 5 | Plas | |
| MdPIP1;2 | NP_001280922.1 | 289 | 30849.79 | 9.30 | 6 | Plas | |
| MdPIP1;3 | NP_001315794.1 | 286 | 30759.57 | 9.08 | 6 | Plas | |
| MdPIP1;4 | NP_001280950.1 | 286 | 30660.58 | 9.15 | 6 | Plas | |
| MdPIP2;1 | XP_008363507.1 | 281 | 30166.00 | 7.65 | 6 | Plas | |
| MdPIP2;2 | XP_008385311.2 | 281 | 30179.06 | 8.25 | 6 | Plas | |
| MdPIP2;3 | XP_008365039.2 | 287 | 30461.39 | 8.95 | 6 | Plas | |
| MdPIP2;4 | XP_008367680.2 | 283 | 30050.96 | 8.68 | 6 | Plas | |
| MdPIP2;5 | XP_008387595.1 | 285 | 30156.86 | 6.89 | 6 | Plas | |
| MdPIP2;6 | XP_008382110.2 | 281 | 29856.60 | 9.08 | 6 | Plas | |
| MdPIP2;7 | XP_008377729.1 | 281 | 29924.69 | 9.22 | 6 | Plas | |
| MdSIP1;1 | XP_008357207.2 | 242 | 25827.57 | 9.55 | 6 | Vacu | |
| MdSIP1;2 | XP_008348137.2 | 240 | 25347.21 | 9.96 | 5 | Vacu/Plas | |
| MdSIP1;3 | XP_008342016.1 | 240 | 25427.44 | 9.97 | 5 | Vacu | |
| MdSIP2;1 | XP_008354672.2 | 236 | 25496.16 | 9.17 | 5 | Vacu | |
| MdSIP2;2 | XP_008338069.3 | 236 | 25364.97 | 9.48 | 6 | Chlo | |
| MdTIP1;1 | XP_008387528.2 | 252 | 26029.00 | 5.18 | 6 | Plas | |
| MdTIP1;2 | XP_008343557.2 | 252 | 26029.25 | 5.62 | 6 | Plas | |
| MdTIP1;3 | XP_008357781.1 | 252 | 25944.08 | 4.96 | 6 | Plas | |
| MdTIP1;4 | XP_008366336.1 | 252 | 25922.12 | 4.96 | 6 | Plas | |
| MdTIP2;1 | XP_008342659.1 | 248 | 25234.34 | 5.76 | 7 | Plas | |
| MdTIP2;2 | XP_008380900.2 | 248 | 25308.41 | 5.62 | 7 | Plas | |
| MdTIP2;3 | XP_008373770.1 | 248 | 25109.15 | 4.86 | 6 | Vacu | |
| MdTIP3;1 | XP_008351935.2 | 256 | 26996.43 | 7.10 | 6 | Plas | |
| MdTIP3;2 | MDP0000868372 2 | 255 | 26710.01 | 7.06 | 5 | Plas | |
| MdTIP4;1 | XP_008393878.2 | 248 | 25864.24 | 5.51 | 7 | Plas | |
| MdTIP4;2 | MD08G1115300 1 | 239 | 25098.22 | 4.98 | 7 | Plas | |
| MdTIP5;1 | XP_008354229.2 | 254 | 26270.51 | 7.74 | 6 | E.R. | |
| MdTIP5;2 | XP_008376403.3 | 256 | 26196.42 | 6.24 | 6 | Chlo/plas | |
| MdXIP1;1 | XP_017182198.2 | 304 | 32165.86 | 5.96 | 6 | Plas | |
| MdXIP2;12 | XP_028965550.1 | 309 | 33249.21 | 8.21 | 7 | Plas |
| Name | NPA motifs | Ar/R filter | Froger’s Positions | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| LB | LE | H2 | H5 | LE1 | LE2 | P1 | P2 | P3 | P4 | P5 | |
| MdNIP1;1 | NPA | NPA | W | V | A | R | F | S | A | Y | I |
| MdNIP1;2 | NPA | NPA | W | V | A | R | F | S | A | Y | I |
| MdNIP2;1 | NPA | NPA | G | S | G | R | L | T | A | Y | V |
| MdNIP2;2 | NPA | NPA | G | S | G | R | L | T | A | Y | V |
| MdNIP2;3 | NPA | NPE | G | S | - | R | L | T | A | Y | V |
| MdNIP4;1 | NPA | NPA | W | V | A | R | L | S | A | Y | F |
| MdNIP5;1 | NPS | NPV | A | I | G | R | F | T | A | Y | L |
| MdNIP5;2 | NPS | NPV | A | I | G | R | L | T | A | Y | L |
| MdNIP5;3 | NPS | NPI | A | I | G | R | F | T | A | Y | L |
| MdNIP6;1 | NPA | NPV | T | I | A | R | F | T | A | Y | L |
| MdNIP7;1 | NPA | NPA | A | V | G | R | Y | S | A | Y | I |
| MdPIP1;1 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| MdPIP1;2 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| MdPIP1;3 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP1;4 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;1 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;2 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;3 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;4 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;5 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdPIP2;7 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| MdSIP1;1 | NPT | NPA | V | L | P | N | M | A | A | Y | W |
| MdSIP1;2 | NPS | NPA | S | L | P | N | M | A | A | Y | W |
| MdSIP1;3 | NPS | NPA | S | L | P | N | M | A | A | Y | W |
| MdSIP2;1 | NPL | NPA | S | L | G | S | F | V | A | Y | W |
| MdSIP2;2 | NPL | SPA | S | L | G | S | F | V | A | Y | W |
| MdTIP1;1 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| MdTIP1;2 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| MdTIP1;3 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| MdTIP1;4 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| MdTIP2;1 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| MdTIP2;2 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| MdTIP2;3 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| MdTIP3;1 | NPA | NPA | H | I | A | R | T | A | A | Y | W |
| MdTIP3;2 | NPA | NPA | H | I | A | R | T | A | A | Y | W |
| MdTIP4;1 | NPA | NPA | H | I | A | R | S | S | A | Y | W |
| MdTIP4;2 | NPA | NPA | H | I | A | R | S | S | A | Y | W |
| MdTIP5;1 | NPA | NPA | N | V | G | C | T | A | A | Y | W |
| MdTIP5;2 | NPA | NPA | N | V | G | C | I | A | A | Y | W |
| MdXIP1;1 | SPV | NPA | V | V | V | R | M | C | A | F | W |
| MdXIP2;1 | NPV | NPA | I | T | V | R | V | C | A | F | W |
| Gene Name 1 | Sequence (5′-3′) | Use | Reference |
|---|---|---|---|
| MpPIP2;1 | F: ATGGCAAAAGATATTGAGGG | Gene cloning | This study |
| R: TTAAGCATTGCTCCTGAAAG | |||
| F: CTTGGCTCGCAAGGTTTCAC | qRT-PCR for MpPIP2;1 in transgenic Arabidopsis | This study | |
| R: GTAGCCATCAGCCAACTCGT | |||
| AtACTIN | F: CCTTCTACCACCAATACATTC | qRT-PCR for reference gene | [40] |
| R: TGTTCCATTGTCGCATAC | |||
| MhPIP2;1 | F:CCTTCTACCACCAATACATTC | qRT-PCR for MhPIP2;1 | This study |
| R:TGATTATCTACAATTCCATAGCC | |||
| MhPIP2;2 | F:GCGGTGGAACTGTAGATA | qRT-PCR for MhPIP2;2 | This study |
| R:GCTTTCTCTGGCATCAAT | |||
| MhPIP2;3 | F:CAAGAGGAGTGCTAGAGAC | qRT-PCR for MhPIP2;3 | This study |
| R:GCCAAGTGGACAATGAAC | |||
| MhPIP2;4 | F:CTTGGACCTGCTGTTATCT | qRT-PCR for MhPIP2;4 | This study |
| R:AATTGCTGCTCCGATGAA | |||
| MhPIP2;5 | F:TGGATTATTCTGGAAGCAT | qRT-PCR for MhPIP2;5 | This study |
| R:GCAACATTAAGGCACATT | |||
| MhTIP2;6 | F:GCAACCCGACCCACTAAA | qRT-PCR for MhPIP2;6 | This study |
| R:ACAACACTCTCAATACACACTACA | |||
| MhTIP2;7 | F:CAGCAACCCAACCAACTAAA | qRT-PCR for MhPIP2;7 | This study |
| R:ATCATCATCCATCCTCTCTCAAT | |||
| MhACTIN | F:TTCGTTTTCGTTTTCGTTTT | qRT-PCR for reference gene | This study |
| R:TGTTCCATTGTCGCATAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Yang, L.; Xin, M.; Ma, F.; Liu, J. Gene-Wide Analysis of Aquaporin Gene Family in Malus domestica and Heterologous Expression of the Gene MpPIP2;1 Confers Drought and Salinity Tolerance in Arabidposis thaliana. Int. J. Mol. Sci. 2019, 20, 3710. https://doi.org/10.3390/ijms20153710
Liu H, Yang L, Xin M, Ma F, Liu J. Gene-Wide Analysis of Aquaporin Gene Family in Malus domestica and Heterologous Expression of the Gene MpPIP2;1 Confers Drought and Salinity Tolerance in Arabidposis thaliana. International Journal of Molecular Sciences. 2019; 20(15):3710. https://doi.org/10.3390/ijms20153710
Chicago/Turabian StyleLiu, Haili, Leilei Yang, Miaomiao Xin, Fengwang Ma, and Jingying Liu. 2019. "Gene-Wide Analysis of Aquaporin Gene Family in Malus domestica and Heterologous Expression of the Gene MpPIP2;1 Confers Drought and Salinity Tolerance in Arabidposis thaliana" International Journal of Molecular Sciences 20, no. 15: 3710. https://doi.org/10.3390/ijms20153710
APA StyleLiu, H., Yang, L., Xin, M., Ma, F., & Liu, J. (2019). Gene-Wide Analysis of Aquaporin Gene Family in Malus domestica and Heterologous Expression of the Gene MpPIP2;1 Confers Drought and Salinity Tolerance in Arabidposis thaliana. International Journal of Molecular Sciences, 20(15), 3710. https://doi.org/10.3390/ijms20153710