Serum Proteome Alterations in Human Cystathionine β-Synthase Deficiency and Ischemic Stroke Subtypes


 and
and 
Abstract
1. Introduction
2. Results
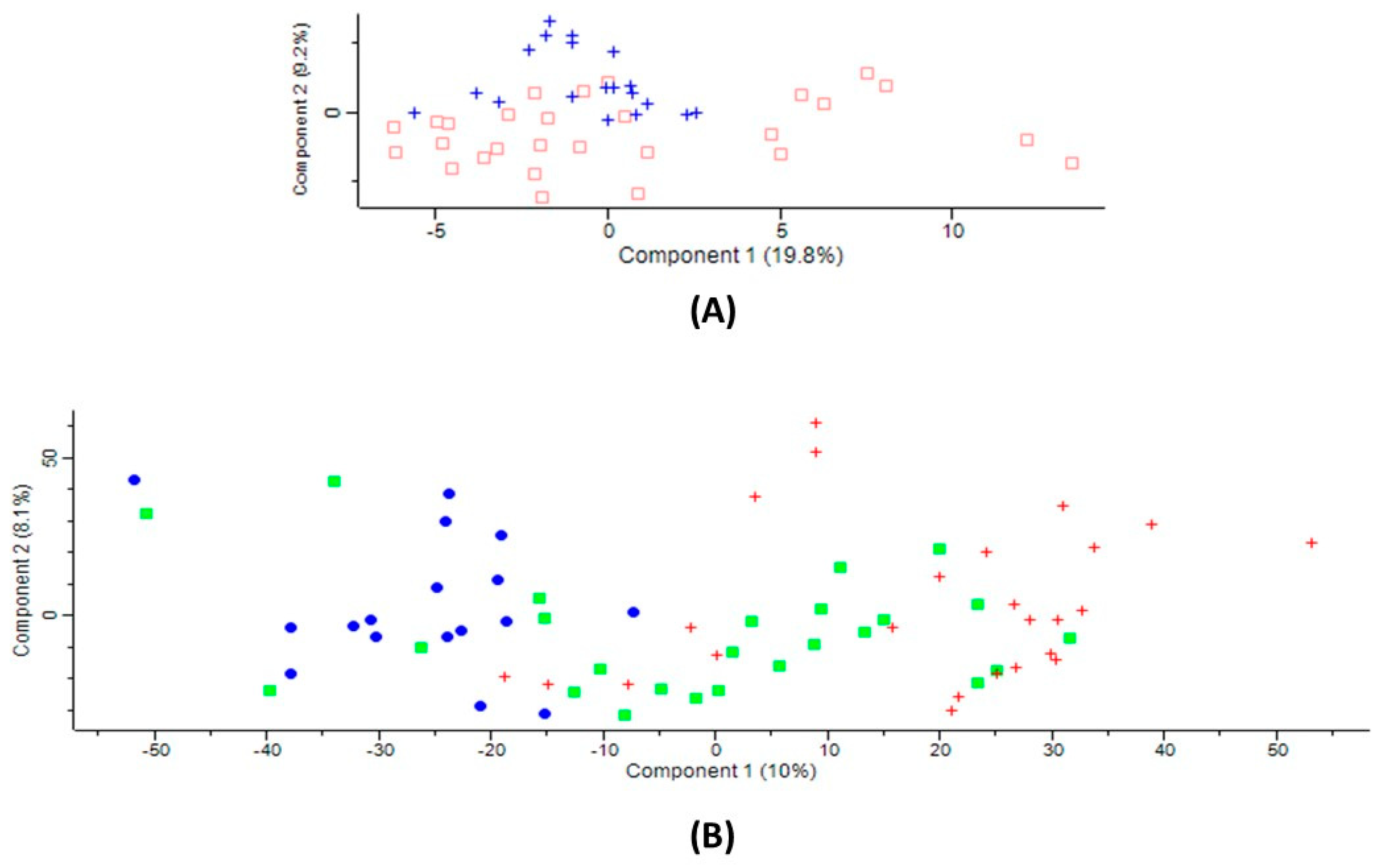
2.1. Serum Proteins Affected by CBS Deficiency
2.2. Serum Proteins Affected by Ischemic Stroke Subtype
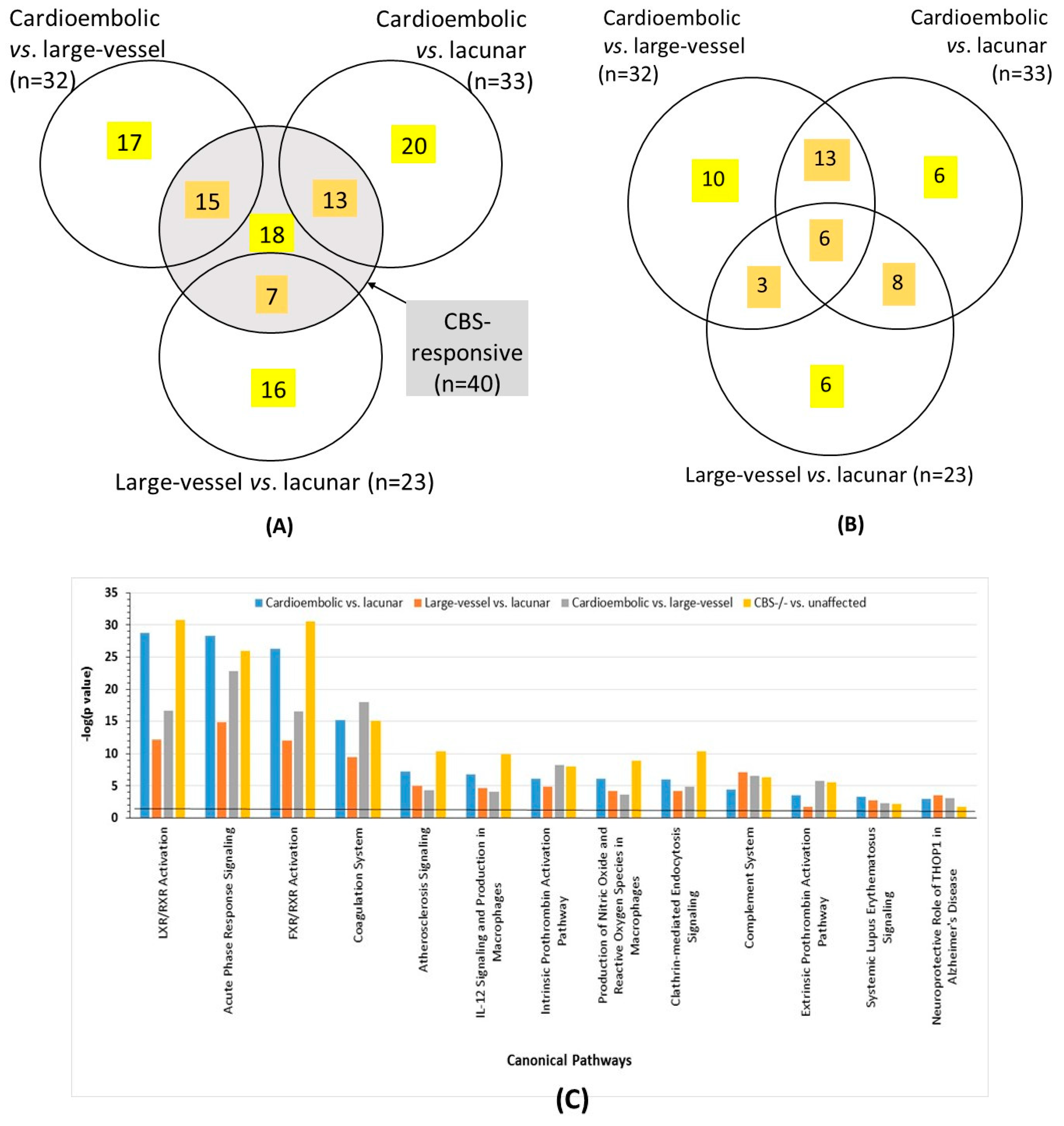
2.3. Ischemic Stroke Subtype-Specific Proteins
2.4. Overlap between Proteins Affected by CBS Deficiency and Ischemic Stroke Subtype
2.5. Validation of Label-Free Mass Spectrometry Analyses by ELISA
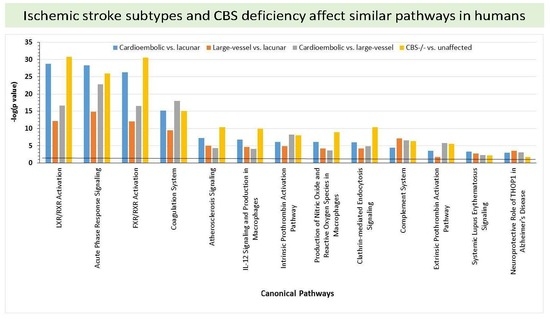
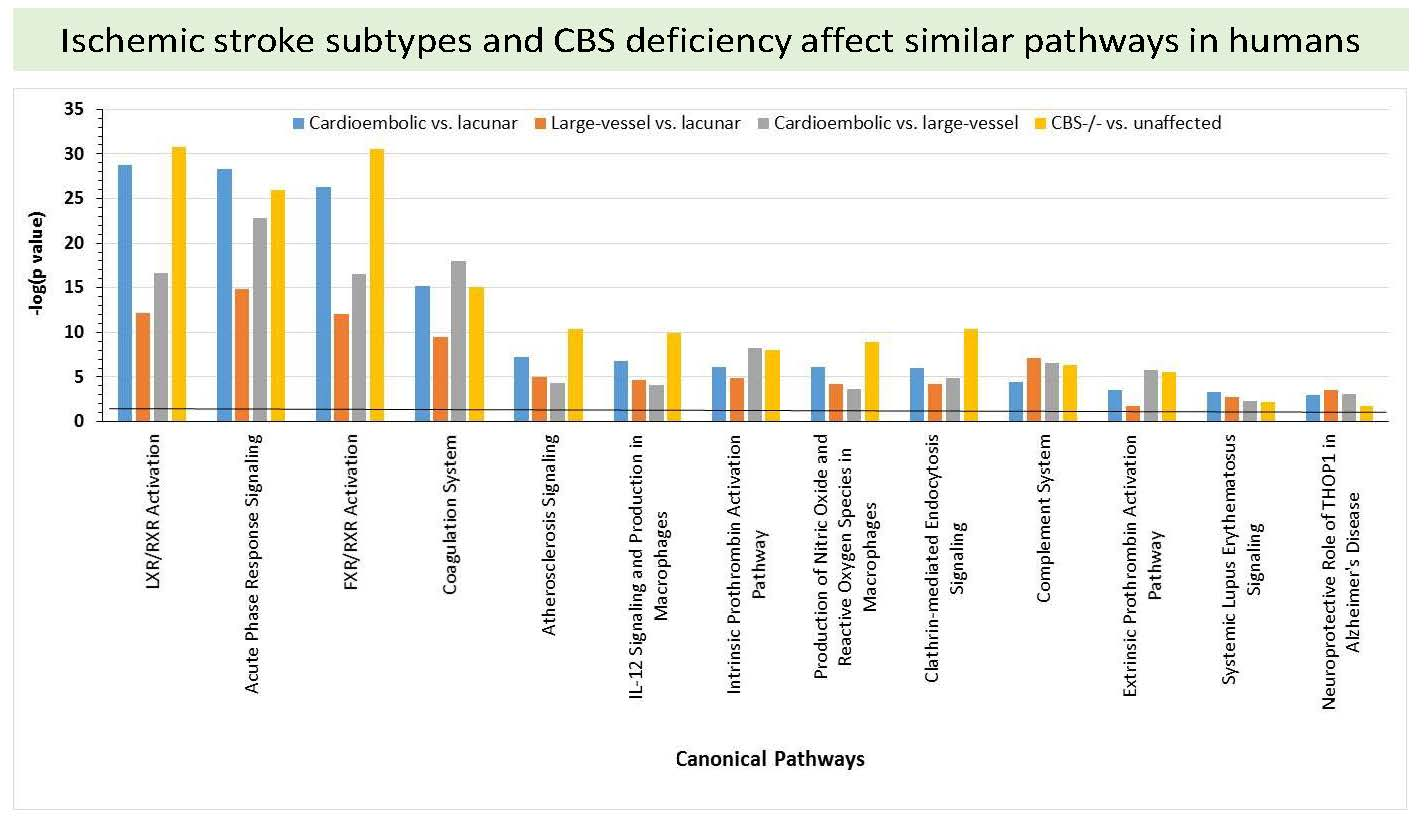
2.6. Bioinformatic Analyses
2.7. Involvement of Homocysteine and Anti-N-Hcy-Protein Autoantibodies
3. Discussion
4. Materials and Methods
4.1. CBS-Deficient Patients
4.2. Ischemic Stroke Patients
4.3. Serum Samples
4.4. Digestion with Trypsin
4.5. Label-Free Mass Spectrometry
4.6. Data Analysis
4.7. Statistical Analyses
4.8. Pathway and Network Analyses
4.9. ELISA Assays
4.10. Anti-N-Hcy-Protein Antibody Assays
4.11. Homocysteine Assays
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart Disease and Stroke Statistics-2019 Update: A Report from the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef] [PubMed]
- Feigin, V.L.; Norrving, B.; Mensah, G.A. Global Burden of Stroke. Circ. Res. 2017, 120, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Spence, J.D. Cardioembolic stroke: everything has changed. Stroke Vasc. Neurol. 2018, 3, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.F.; Sarracino, D.A.; Prakash, A.; Athanas, M.; Krastins, B.; Rezai, T.; Sutton, J.N.; Peterman, S.; Gvozdyak, O.; Chou, S.; et al. Discrimination of ischemic and hemorrhagic strokes using a multiplexed, mass spectrometry-based assay for serum apolipoproteins coupled to multi-marker ROC algorithm. Proteom. Clin. Appl. 2012, 6, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Levy, H.L.; Kraus, J.P. Disorders of transsulfuration. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Childs, B., Kinzler, K.W., Vogelstein, B., Eds.; Mc Graw-Hill: New York, NY, USA, 2001; Volume 2, pp. 2007–2056. [Google Scholar]
- Kelly, P.J.; Furie, K.L.; Kistler, J.P.; Barron, M.; Picard, E.H.; Mandell, R.; Shih, V.E. Stroke in young patients with hyperhomocysteinemia due to cystathionine beta-synthase deficiency. Neurology 2003, 60, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Chwatko, G.; Boers, G.H.; Strauss, K.A.; Shih, D.M.; Jakubowski, H. Mutations in methylenetetrahydrofolate reductase or cystathionine beta-synthase gene, or a high-methionine diet, increase homocysteine thiolactone levels in humans and mice. FASEB J. 2007, 21, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H.; Boers, G.H.; Strauss, K.A. Mutations in cystathionine beta-synthase or methylenetetrahydrofolate reductase gene increase N-homocysteinylated protein levels in humans. FASEB J. 2008, 22, 4071–4076. [Google Scholar] [CrossRef] [PubMed]
- Gurda, D.; Handschuh, L.; Kotkowiak, W.; Jakubowski, H. Homocysteine thiolactone and N-homocysteinylated protein induce pro-atherogenic changes in gene expression in human vascular endothelial cells. Amino Acids 2015, 47, 1319–1339. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Homocysteine Modification in Protein Structure/Function and Human Disease. Physiol. Rev. 2019, 99, 555–604. [Google Scholar] [CrossRef]
- Undas, A.; Perla, J.; Lacinski, M.; Trzeciak, W.; Kazmierski, R.; Jakubowski, H. Autoantibodies against N-homocysteinylated proteins in humans: implications for atherosclerosis. Stroke 2004, 35, 1299–1304. [Google Scholar] [CrossRef]
- Sikora, M.; Marczak, L.; Twardowski, T.; Stobiecki, M.; Jakubowski, H. Direct monitoring of albumin lysine-525 N-homocysteinylation in human serum by liquid chromatography/mass spectrometry. Anal. Biochem. 2010, 405, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Eikelboom, J.W.; Hankey, G.J.; Anand, S.S.; Lofthouse, E.; Staples, N.; Baker, R.I. Association between high homocyst(e)ine and ischemic stroke due to large- and small-artery disease but not other etiologic subtypes of ischemic stroke. Stroke 2000, 31, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, A.; Brattstrom, L.; Norrving, B.; Hultberg, B.; Andersson, A.; Johansson, B.B. Plasma homocysteine in the acute and convalescent phases after stroke. Stroke 1995, 26, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Coull, B.M.; Malinow, M.R.; Beamer, N.; Sexton, G.; Nordt, F.; de Garmo, P. Elevated plasma homocyst(e)ine concentration as a possible independent risk factor for stroke. Stroke 1990, 21, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Bots, M.L.; Launer, L.J.; Lindemans, J.; Hoes, A.W.; Hofman, A.; Witteman, J.C.; Koudstaal, P.J.; Grobbee, D.E. Homocysteine and short-term risk of myocardial infarction and stroke in the elderly: the Rotterdam Study. Arch. Intern. Med. 1999, 159, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.Y.; Ampil, E.R.; Chen, C.P.; Auchus, A.P. The relationship between homocysteine, cognition and stroke subtypes in acute stroke. J. Neurol. Sci. 2006, 250, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Weng, Y.; Zheng, L.; Li, H.; Gong, Q.; Fu, Y.; Zhao, J. Polymorphism of the complement 5 gene is associated with large artery atherosclerosis stroke in Chinese patients. Arq. Neuropsiquiatr. 2016, 74, 881–886. [Google Scholar] [CrossRef]
- Jickling, G.C.; Sharp, F.R. Biomarker panels in ischemic stroke. Stroke 2015, 46, 915–920. [Google Scholar] [CrossRef]
- Alvarez-Perez, F.J.; Castelo-Branco, M.; Alvarez-Sabin, J. Usefulness of measurement of fibrinogen, D-dimer, D-dimer/fibrinogen ratio, C reactive protein and erythrocyte sedimentation rate to assess the pathophysiology and mechanism of ischaemic stroke. J. Neurol. Neurosurg. Psychiatry 2011, 82, 986–992. [Google Scholar] [CrossRef]
- Turaj, W.; Slowik, A.; Pulyk, R.; Adamski, M.; Szczudlik, A. Comparison of plasma concentrations of fibrinogen in patients with ischemic stroke due to large vessel disease and small vessel disease. Neurol. Neurochir. Pol. 2006, 40, 297–301. [Google Scholar]
- Malik, R.; Dau, T.; Gonik, M.; Sivakumar, A.; Deredge, D.J.; Edeleva, E.V.; Götzfried, J.; van der Laan, S.W.; Pasterkamp, G.; Beaufort, N.; et al. Common coding variant in SERPINA1 increases the risk for large artery stroke. Proc. Natl. Acad. Sci. USA 2017, 114, 3613–3618. [Google Scholar] [CrossRef] [PubMed]
- Meschia, J.F. Alpha-1 antitrypsin dysfunction and large artery stroke. Proc. Natl. Acad. Sci. USA 2017, 114, 3555–3557. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Chen, C.P.; Sze, S.K. Discovery of prognostic biomarker candidates of lacunar infarction by quantitative proteomics of microvesicles enriched plasma. PLoS ONE 2014, 9, e94663. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhou, L.; Guo, T.; Wang, N.; Hao, H.; Zhou, Y.; Yu, D. Plasma proteomics reveals coagulation, inflammation, and metabolic shifts in H-type hypertension patients with and without acute ischemic stroke. Oncotarget 2017, 8, 100384–100395. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, H.J.; deVeber, G.A.; Hills, N.K.; Dowling, M.M.; Fox, C.K.; Mackay, M.T.; Kirton, A.; Yager, J.Y.; Bernard, T.J.; Hod, E.A.; et al. Inflammatory Biomarkers in Childhood Arterial Ischemic Stroke: Correlates of Stroke Cause and Recurrence. Stroke 2016, 47, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Zhan, X.; Su, X.; Guo, L.; Lv, L.; Su, B. Proteomic analysis of serum proteins in acute ischemic stroke patients treated with acupuncture. Exp. Biol. Med. (Maywood) 2011, 236, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Ambrosius, W.; Michalak, S.; Kazmierski, R.; Andrzejewska, N.; Kozubski, W. Predictive value of serum transthyretin for outcome in acute ischemic stroke. PLoS ONE 2017, 12, e0179806. [Google Scholar] [CrossRef] [PubMed]
- Allard, L.; Lescuyer, P.; Burgess, J.; Leung, K.Y.; Ward, M.; Walter, N.; Burkhard, P.R.; Corthals, G.; Hochstrasser, D.F.; Sanchez, J.C. ApoC-I and ApoC-III as potential plasmatic markers to distinguish between ischemic and hemorrhagic stroke. Proteomics 2004, 4, 2242–2251. [Google Scholar] [CrossRef]
- Maclean, K.N.; Gaustadnes, M.; Oliveriusova, J.; Janosik, M.; Kraus, E.; Kozich, V.; Kery, V.; Skovby, F.; Rudiger, N.; Ingerslev, J.; et al. High homocysteine and thrombosis without connective tissue disorders are associated with a novel class of cystathionine beta-synthase (CBS) mutations. Hum. Mutat. 2002, 19, 641–655. [Google Scholar] [CrossRef]
- Orendae, M.; Pronicka, E.; Kubalska, J.; Janosik, M.; Sokolova, J.; Linnebank, M.; Koch, H.G.; Kozich, V. Identification and functional analysis of two novel mutations in the CBS gene in Polish patients with homocystinuria. Hum. Mutat. 2004, 23, 631. [Google Scholar] [CrossRef]
- Sikora, M.; Marczak, L.; Kubalska, J.; Graban, A.; Jakubowski, H. Identification of N-homocysteinylation sites in plasma proteins. Amino Acids 2014, 46, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H.; Perla-Kajan, J.; Finnell, R.H.; Cabrera, R.M.; Wang, H.; Gupta, S.; Kruger, W.D.; Kraus, J.P.; Shih, D.M. Genetic or nutritional disorders in homocysteine or folate metabolism increase protein N-homocysteinylation in mice. FASEB J. 2009, 23, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Xu, Y.K.; Chan, P.; Pattengale, P.K. Simple, Fast, and Simultaneous Detection of Plasma Total Homocysteine, Methylmalonic Acid, Methionine, and 2-Methylcitric Acid Using Liquid Chromatography and Mass Spectrometry (LC/MS/MS). JIMD Rep. 2013, 10, 69–78. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | CBS-Deficient Patients (n = 10) | Healthy Controls (n = 14) | p-Value |
|---|---|---|---|
| Female sex, n (%) | 5 (50) | 7 (50) | NS |
| Mean age, years | 37.3 ± 7.3 | 38.0 ± 12.5 | NS |
| History of stroke, n (%) | 4 (40) | 0 (0) | <0.05 |
| Total cholesterol, mg/dL | 180 ± 48 | 180 ± 37 | NS |
| HDL cholesterol, mg/dL | 58 ± 9 | 67 ± 22 | NS |
| LDL cholesterol, mg/dL | 193 ± 42 | 96 ± 35 | NS |
| Triglyceride, mg/dL | 102 ± 46 | 84 ± 44 | NS |
| Methionine, µM | 567 ± 16 | 14.9 ± 4.5 | <0.001 |
| tHcy, µM | 71.2 ± 55.6 | 9.7 ± 5.9 | <0.001 |
| Anti-N-Hcy-protein antibodies, A492 | 0.33 ± 0.16 | 0.12 ± 0.10 | 0.005 |
| Variable | All Patients (n = 68) | Ischemic Stroke Subtype Patients | ANOVA p-Value | ||
|---|---|---|---|---|---|
| Large-Vessel (n = 28) | Cardioembolic (n = 16) | Lacunar (n = 24) | |||
| Female sex, % | 41.2 | 32.1 | 56.2 | 41.7 | |
| Age, years | 66.4 ± 12.7 | 63.8 ± 11.0 | 78.3 ± 15.0 | 62.0 ± 8.0 | <0.001 |
| Atrial fibrillation, % | 34 | 6 | 91 | 0 | |
| Hypertension, % | 72 | 67 | 67 | 83 | |
| Cholesterol, mg/dL | 207.6 ± 57.4 | 218.0 ± 64.7 | 173.0 ± 64.5 | 211.5 ± 40.5 | 0.005 |
| HDL cholesterol, mg/dL | 52.0 ± 14.0 | 55.9 ± 14.75 | 51.0 ± 15.0 | 49.3 ± 13.0 | NS |
| LDL cholesterol, mg/dL | 131.6 ± 48.7 | 138.0 ± 59.0 | 114.5 ± 53.3 | 136.0 ± 33.3 | NS |
| Triglycerides, mg/dL | 116.0 ± 59.4 | 120.0 ± 51.0 | 77.8 ± 25.8 | 133.0 ± 72.0 | 0.001 |
| Creatinine, µmol/L | 94.0 ± 34.0 | 89.0 ± 24.7 | 100.6 ± 38.4 | 94.3 ± 34.6 | NS |
| Glucose, mmol/L | 5.85 ± 1.37 | 5.70 ± 1.26 | 5.96 ± 1.54 | 5.86 ± 1.36 | NS |
| Alanine aminotransferase, U/L | 24.87 ± 18.7 | 21.7 ± 8.6 | 26.8 ± 27.6 | 26.2 ± 13.2 | NS |
| Aspartate aminotransferase, U/l | 27.4 ± 16.4 | 25.9 ± 10.0 | 30.2 ± 23.3 | 26.2 ± 11.7 | NS |
| Thyroid-stimulating hormone, mU/L | 1.64 ± 1.47 | 1.28 ± 0.86 | 1.68 ± 1.08 | 1.94 ± 2.23 | NS |
| Free triiodothyronine, pmol/L | 5.08 ± 3.43 | 6.94 ± 5.4 | 3.76 ± 1.09 | 4.57 ± 0.72 | 0.007 |
| Leukocytes, × 109/L | 8.5 ± 2.75 | 9.8 ± 3.4 | 7.3 ± 1.9 | 7.9 ± 2.0 | 0.01 |
| tHcy, μM | 3.3 ± 1.6 | 3.3 ± 1.2 | 3.5 ± 1.6 | 3.0 ± 2.0 | NS |
| Anti-N-Hcy-protein antibodies, A492 | 0.13 ± 0.08 | 0.12 ± 0.08 | 0.14 ± 0.11 | 0.10 ± 0.07 | NS |
| Gene Name | Protein Name | Cardioembolic vs. Large-Vessel Stroke | Cardioembolic vs. Lacunar Stroke | Large-Vessel vs. Lacunar Stroke | ANOVA p Value | CBS−/− vs. Control | Molecular Function/Biological Process | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fold Change | p Value | Fold Change | p Value | Fold Change | p Value | Fold Change | T-test p Value | ||||
| AFM | Afamin | 0.84 | 0.011 | 0.85 | 0.005 | 0.0148 | 0.83 | 0.007 | Vitamin transport | ||
| ORM1 | α-1-acid glycoprotein1 | 1.35 | 0.001 | 1.20 | 0.010 | 0.0018 | Acute inflammatory response/acute-phase response | ||||
| ORM2 | α-1-acid glycoprotein2 | 0.81 | 0.042 | Acute-phase response | |||||||
| SERPINA3 | α-1-antichymotrypsin | 1.21 | 0.046 | 1.42 | <1 × 10−4 | 1.18 | 0.021 | 0.0002 | Acute inflammatory response/acute-phase response | ||
| SERPINA1 | α-1-antitrypsin | 1.17 | 0.007 | 1.18 | 0.011 | 0.0106 | 1.14 | 0.045 | Acute phase response; Blood coagulation | ||
| SERPINF2 | α-2-antiplasmin | 0.85 | 0.027 | 0.0305 | 0.80 | 1.2 × 10−5 | Complement/coagulation cascades | ||||
| AHSG | α-2-HS-glycoprotein | 0.85 | 0.001 | 0.81 | 0.0001 | 0.0001 | 0.80 | 5.3 × 10−5 | Acute phase response | ||
| SERPINC1 | Antithrombin-III | 0.84 | 4.5 × 10−5 | Complement/coagulation cascades | |||||||
| A2M | α-2-macroglobulin | 1.29 | 0.024 | 1.29 | 0.006 | Blood coagulation | |||||
| APOA1 | Apolipoprotein A-I | 0.87 | 0.038 | Fat digestion/absorption | |||||||
| APOA4 | Apolipoprotein A-IV | 0.71 | 0.006 | Cholesterol transport | |||||||
| APOC1 | Apolipoprotein C-I | 0.67 | 0.004 | 0.52 | <1 × 10−4 | 0.77 | 0.035 | 4.5E-5 | 1.40 | 0.032 | Cholesterol efflux/lipid and lipoprotein metabolic process |
| APOC3 | Apolipoprotein C-III | 0.68 | 0.014 | Cholesterol transport | |||||||
| APOL1 | Apolipoprotein L1 | 1.23 | 0.042 | Cholesterol metabolism | |||||||
| APOM | Apolipoprotein M | 0.81 | 0.012 | 0.0375 | 0.70 | 0.001 | Cholesterol transport, antioxidant activity | ||||
| APOH | Beta-2-glycoprotein 1T | 0.86 | 0.015 | Blood coagulation | |||||||
| C4BPA | C4b-binding protein α-chain | 0.44 | 0.032 | Complement activation/immune response | |||||||
| CPB2 | Carboxypeptidase B2 | 0.74 | 0.018 | 0.0431 | 1.12 | 0.044 | Complement/coagulation cascades; Protein digestion | ||||
| CP | Ceruloplasmin | 1.21 | 0.001 | 1.12 | 0.007 | 0.0020 | Cellular iron ion homeostasis | ||||
| CLU | Clusterin | 0.88 | 0.014 | 0.88 | 0.004 | 0.0072 | 0.93 | 0.019 | Negative regulation of amyloid-beta formation | ||
| F9;factor IX | Coagulation factor IX | 1.45 | 0.004 | 1.29 | 0.018 | 0.0010 | Blood coagulation | ||||
| F13B | Coagulation factor XIII B-chain | 0.74 | 0.010 | 0.71 | 0.005 | 0.0010 | 0.69 | 0.0005 | Blood coagulation | ||
| C1QA | Complement C1q subunit A | 1.66 | 0.042 | Complement and coagulation cascades | |||||||
| C1QB | Complement C1q subunit B | 1.12 | 0.038 | 0.90 | 0.047 | 0.0433 | Complement/coagulation cascades | ||||
| C5 | Complement C5 | 1.14 | 0.003 | 0.0136 | Complement/coagulation cascades | ||||||
| C1R | Complement C1r subcomponent | 0.82 | 5 × 10−5 | Complement/coagulation cascades | |||||||
| C1S | Complement C1s subcomponent | 0.86 | 0.004 | Complement/coagulation cascades | |||||||
| C9 | Complement component C9 | 1.36 | 1 × 10−4 | 1.25 | 0.002 | 0.0002 | 1.36 | 0.004 | Complement/coagulation cascades | ||
| CFI | Complement factor I | 0.85 | 0.002 | Complement/coagulation cascades | |||||||
| CRP | C-reactive protein | 2.44 | 0.046 | 5.89 | 1 × 10−5 | 2.41 | 0.011 | 2.5 × 10−7 | Acute inflammatory response/acute-phase response | ||
| HEL-213 | Epididymis luminal protein 213 | 1.55 | 0.003 | ||||||||
| FGA | Fibrinogen α-chain; | 1.53 | 0.006 | 2.01 | 1 × 10−6 | 1.32 | 0.031 | 1.6 × 10−5 | Blood coagulation | ||
| FBLN1 | Fibulin-1 | 1.99 | 0.0005 | 0.0326 | 2.13 | 0.001 | Blood coagulation/fibrin clot formation | ||||
| FCN3 | Ficolin-3 | 0.72 | 0.007 | 0.0211 | 0.76 | 0.003 | Complement activation | ||||
| GSN | Gelsolin | 0.87 | 0.004 | 0.0160 | 1.11 | 0.015 | Actin filament capping/Amyloid fibril formation | ||||
| GPX3 | Glutathione peroxidase 3 | 1.26 | 0.018 | 1.41 | 0.005 | Cellular response to oxidative stress | |||||
| HPR | Haptoglobin-related | 0.61 | 0.002 | 0.64 | 0.001 | 0.0018 | Receptor-mediated endocytosis | ||||
| HPX | Hemopexin | 0.90 | 0.004 | Cellular iron ion homeostasis | |||||||
| SERPIND1 | Heparin cofactor 2 | 0.80 | 0.027 | 0.71 | 4 × 10−4 | 0.0023 | 0.77 | 0.005 | Complement/coagulation cascades | ||
| H2AFJ | Histone H2A | 0.49 | 0.027 | Chromatin silencing | |||||||
| HIST1H4A | Histone H4 | 0.35 | 0.001 | 0.43 | 0.003 | 1.2 × 10−5 | Telomere organization | ||||
| IGHV3-7 | Ig heavy chain V-III region GAL | 1.64 | 0.019 | Immune response | |||||||
| IGKV1D-12 | IgK chain V-I region Wes | 0.77 | 0.032 | Immune response | |||||||
| IGH@ | IGH@ protein | 0.71 | 0.006 | Immune response | |||||||
| IGK@ | IGK@ protein | 0.88 | 0.019 | 1.31 | 0.002 | Immune response | |||||
| IGHD | Immunoglobulin heavy constant delta | 3.39 | 0.009 | Immune response | |||||||
| IGHV3-72 | Ig heavy variable 3-72 | 1.48 | 0.025 | Immune response | |||||||
| IGJ; JCHAIN | Ig J-chain | 1.51 | 0.006 | Immune response | |||||||
| IGKV2D-24 | IgK variable | 0.70 | 0.024 | Immune response | |||||||
| IGFALS | Insulin-like growth fac-tor-binding complex acid-labile subunit | 0.59 | 0.003 | 0.58 | 0.002 | 0.0005 | Cell adhesion | ||||
| ITIH2 | Inter-α-trypsin inhibi-tor heavy chain H2 | 0.92 | 0.022 | Amine metabolic process | |||||||
| ITIH3 | Inter-α-trypsin inhibi-tor heavy chain H3 | 1.46 | 0.001 | 1.72 | 1 × 10−5 | 1.1 × 10−5 | Amine metabolic process | ||||
| ITIH4 | Inter-α-trypsin inhi-bitor heavy chain H4 | 1.12 | 0.001 | Amine metabolic process | |||||||
| N/A | cDNA FLJ53075, high-ly similar to KNG1 | 1.55 | 0.001 | ||||||||
| SERPINA4 | Kallistatin | 0.70 | 0.012 | 0.59 | <1 × 10−4 | 0.84 | 0.018 | 4.8 × 10−5 | Platelet degranulation | ||
| KNG1 | Kininogen-1 | 0.92 | 0.029 | 0.92 | 0.009 | 0.0002 | 0.86 | 0.000 | Blood coagulation/inflammatory response | ||
| LBP | Lipopolysaccharide-binding protein | 1.90 | 0.002 | Acute phase response | |||||||
| LUM | Lumican | 1.27 | 0.017 | 0.78 | 0.002 | 0.0037 | Collagen binding | ||||
| PGLYRP2 | N-acetylmuramoyl-L-alanine amidase | 0.87 | 0.001 | 0.91 | 0.023 | 0.0055 | Immune resonse/inflammatory response | ||||
| KLKB1 | Plasma kallikrein | 0.81 | 0.016 | 0.91 | 0.043 | Complement/coagulation cascades | |||||
| PLG | Plasminogen | 0.82 | 0.0002 | 0.89 | 0.004 | 1.09 | 0.039 | 1 × 10−4 | Blood coagulation, fibrynolysis | ||
| PF4 | Platelet factor 4 | 0.72 | 0.001 | 0.0264 | Platelet degranulation, inflammatory response | ||||||
| PZP | Pregnancy zone protein | 3.87 | 0.0003 | 0.38 | 0.004 | 0.0001 | Female pregnancy | ||||
| AMBP | Protein AMBP; α-1-microglobulin; Trypstatin | 0.90 | 0.005 | 0.0497 | Cell adhesion; Heme metabolic process | ||||||
| F2 | Prothrombin | 0.90 | 0.008 | 0.0142 | 0.79 | 8 × 10−6 | Complement/coagulation cascades; Neuroactive ligand-receptor interaction; Regulation of actin cytoskeleton | ||||
| RBP4 | Retinol-binding protein 4 | 0.75 | 0.010 | 0.79 | 0.012 | 0.0093 | Cardiac muscle tissue development | ||||
| SAA1 | Serum amyloid A-1 | 6.31 | 0.004 | 11.13 | <1 × 10−5 | 0.0052 | 1.97 | 0.020 | Acute-phase response | ||
| APCS | Serum amyloid P | 0.83 | 0.010 | 0.0237 | Immune response | ||||||
| HEL111;TTR | Transthyretin | 0.68 | 0.004 | 0.64 | 0.004 | 0.0060 | 0.57 | 5.4 × 10−9 | Retinol metabolic process, thyroid hormone transport | ||
| HEL-S-51;GC | Vit. D-binding protein | 0.91 | 0.020 | 0.87 | <1 × 10−3 | 0.0007 | 0.85 | 0.0001 | Vitamin D metabolic process | ||
| Cardioembolic vs. Large-Vessel Stroke | Cardioembolic vs. Lacunar Stroke | Large-Vessel vs. Lacunar Stroke | CBS−/− vs. Control |
|---|---|---|---|
| APCS | AMBP | APOL1 | APOA1 |
| APOM | APOA4 | C5 | APOC3 |
| C1QA | FCN3 | GSN | APOH |
| C4BPA | ITIH4 | GPX3 | C1R |
| CPB2 | LBP | H2AFJ | C1S |
| FBLN1 | PF4 | IGK@ | CFI |
| IGKV1D-12 | HEL0213 | ||
| KLKB1 | HPX | ||
| SERPINF2 | IGHV3-7 | ||
| IGHD | |||
| IGHV3-7 | |||
| IGH@ | |||
| IGJ; JCHAIN | |||
| IGKV2D-24 | |||
| ITIH2 | |||
| ORM2 | |||
| SERPINC1 | |||
| cDNA FLJ53075, highly similar to KNG1 |
| Analysis | Molecules in Network | Score | Focus Molecules | Top Diseases and Functions |
|---|---|---|---|---|
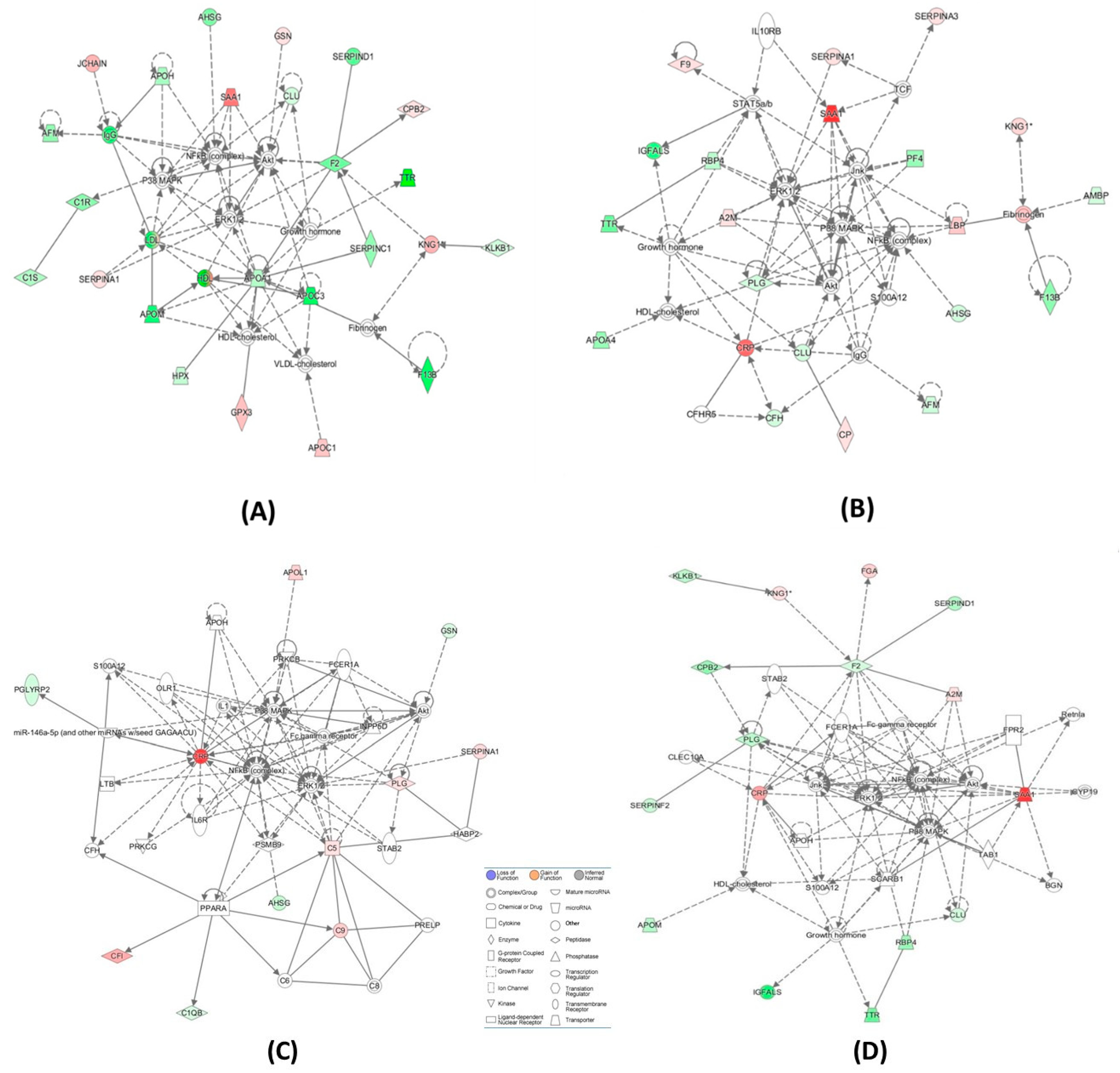
| Cardioembolic vs. large-vessel stroke (Figure 3A) | ↑A2M, Akt, APOH, ↓APOM, BGN, CLEC10A, ↓CLU, ↓CPB2,↑CRP, CYP19, ERK1/2, ↓F2, Fc gamma receptor, FCER1A, ↑FGA, FPR2, Growth hormone, HDL-cholesterol, ↓IGFALS, Jnk, KLKB1,↑KNG1, NFkB (complex), P38 MAPK, ↓PLG, ↓RBP4, Retnla, S100A12, ↑SAA1, SCARB1, ↓SERPIND1, ↓SERPINF2, STAB2, TAB1, ↓TTR | 36 | 16 | Hematological System Development and Function, Cell-To-Cell Signaling and Interaction, Organismal Functions |
| Cardioembolic vs. large-vessel stroke | ↓AFM,ALT,↓APCS, ↓APOC1,APOE,C6, ↑C7,C8,C9,C1q,↑C1QA, ↑C1QB, ↓C4BPA, Ccl2, ↑CRP, CRYAB, CTSB, CXCL2, ↓F13B,↑FBLN1, ↑FGA,↓GC, HNF1A, HNF4A, HRG, IL1R1, INSR, ↑ITIH3, LDL-cholesterol, ↑LUM, PSEN2, SCARB1, ↑SERPINA3, STK40, VLDL-cholesterol | 33 | 15 | Metabolic Disease, Developmental Disorder, Hereditary Disorder |
| Cardioembolic vs. lacunar stroke (Figure 3B) | ↑A2M, ↓AFM, ↓AHSG, Akt, ↓AMBP, ↓APOA4, ↓CFH,CFHR5, ↓CLU, ↑CP, ↑CRP, ERK1/2, ↑F9, ↓F13B, Fibrinogen, Growth hormone, HDL-cholesterol, ↓IGFALS, IgG, IL10RB, Jnk, ↑KNG1, ↑LBP, NFkB (complex), P38 MAPK, ↓PF4, ↓PLG, ↓RBP4,S100A12, ↑SAA1, ↑SERPINA1, ↑SERPINA3,STAT5a/b, TCF, ↓TTR | 50 | 21 | Cell-To-Cell Signaling and Interaction, Hematological System Development and Function, Neurological Disease |
| Cardioembolic vs. lacunar stroke | ↓AHSG, ↓APOC1, ARG1, C5, C6,↑C7, C8,↑C9, CCND1, CPB2, ↑CRP, CTSB, EHF, ↓FCN3, MASP1, MASP2, Fc-γ receptor, FGA, FGB, FGG, ↓GC, ↓HIST1H4H, HNF1A, HNF4A, IL6, ↑ITIH3, ↑ITIH4, LDL-cholesterol, LGALS3, N-cor, NR1H2, NR5A2, ↑ORM1, PGLYRP2, PRELP, ↑SAA1, ↓SERPIND1, TTR | 32 | 15 | Inflammatory Response, Metabolic Disease, Cell-To-Cell Signaling and Interaction |
| Large-vessel vs. lacunar stroke (Figure 3C) | ↓AHSG, Akt, APOH, ↑APOL1, ↑C5, C6, C8, ↑C9, ↓C1QB, CFH, ↑CFI, ↑CRP, ERK1/2, Fc-γ receptor, FCER1A, ↓GSN, HABP2, IL1, IL6R, INPP5D, LTB, miR-146a-5p (and other miRNAs w/seed GAGAACU), NFkB (complex), OLR1, P38 MAPK, ↓PGLYRP2, ↑PLG, PPARA, PRELP, PRKCB, PRKCG, PSMB9, S100A12, ↑SERPINA1, STAB2 | 24 | 11 | Inflammatory Response, Cellular Movement, Immune Cell Trafficking |
| Large-vessel vs. lacunar stroke | AGER, ↓APOC1, CCND1, CDK4, ↑CP, CRYAB, EHF, F2, ↑F9, Fc-γ receptor, FCER1A, Ferritin, ↑FGA, FGB, FGG, ↑GPX3, ↓H2AFJ, ↓HIST1H4H, INPP5D, ↑KNG1, Ldh (complex), LRPAP1, ↓LUM, MAPK1,N-cor, NR5A2, Nuclear factor 1, ↑ORM1,PLCG2, PSMB9, PSME2, ↑SERPINA3, TNF, TNFSF10, TP53 | 24 | 11 | Cell-To-Cell Signaling and Interaction, Hematological System Development and Function, Inflammatory Response |
| CBS−/− vs. control (Figure 3D) | ↓AFM, ↓AHSG, Akt, ↓APOA1,↑APOC1,↓APOC3, ↓APOH, ↓APOM, ↓C1R, ↓C1S, ↓CLU,↑CPB2,ERK1/2,↓F2, ↓F13B, Fibrinogen, ↑GPX3, Growth hormone, ↑GSN, HDL, HDL-cholesterol, ↓HPX, IgG, ↑JCHAIN, ↓KLKB1, ↑KNG1, LDL, NFkB (complex), P38 MAPK, ↑SAA1, ↑SERPINA1, ↓SERPINC1, ↓SERPIND1, ↓TTR, VLDL-cholesterol | 60 | 24 | Metabolic Disease, Hematological System Development and Function, Lipid Metabolism |
| CBS−/− vs. control | ADAMTS1, ↓AHSG,ALT, ↓APOC3, BGN, BIRC5, C6, C7,↑C9, ↓C1S,CCND1, ↓CFI, ↑CPB2, ↓CPN1, F13A1, ↑FBLN1, ↓FCN3, FGA, FPR2, ↓GC, GCNT3, GPR119, HNF1A, ↓IGHG1, IL1B, ↓ITIH2, MASP1, MASP2, ↓ORM2, PPARA, ↑SAA1, ↓SERPIND1, ↓SERPINF2, TG, TNF | 35 | 16 | Humoral Immune Response, Inflammato-ry Response, Develop-mental Disorder |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikora, M.; Lewandowska, I.; Kupc, M.; Kubalska, J.; Graban, A.; Marczak, Ł.; Kaźmierski, R.; Jakubowski, H. Serum Proteome Alterations in Human Cystathionine β-Synthase Deficiency and Ischemic Stroke Subtypes. Int. J. Mol. Sci. 2019, 20, 3096. https://doi.org/10.3390/ijms20123096
Sikora M, Lewandowska I, Kupc M, Kubalska J, Graban A, Marczak Ł, Kaźmierski R, Jakubowski H. Serum Proteome Alterations in Human Cystathionine β-Synthase Deficiency and Ischemic Stroke Subtypes. International Journal of Molecular Sciences. 2019; 20(12):3096. https://doi.org/10.3390/ijms20123096
Chicago/Turabian StyleSikora, Marta, Izabela Lewandowska, Małgorzata Kupc, Jolanta Kubalska, Ałła Graban, Łukasz Marczak, Radosław Kaźmierski, and Hieronim Jakubowski. 2019. "Serum Proteome Alterations in Human Cystathionine β-Synthase Deficiency and Ischemic Stroke Subtypes" International Journal of Molecular Sciences 20, no. 12: 3096. https://doi.org/10.3390/ijms20123096
APA StyleSikora, M., Lewandowska, I., Kupc, M., Kubalska, J., Graban, A., Marczak, Ł., Kaźmierski, R., & Jakubowski, H. (2019). Serum Proteome Alterations in Human Cystathionine β-Synthase Deficiency and Ischemic Stroke Subtypes. International Journal of Molecular Sciences, 20(12), 3096. https://doi.org/10.3390/ijms20123096