The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics



Abstract
1. Introduction
2. Extracellular Vesicles in Cutaneous Regenerative Medicine
2.1. Use of Extracellular Vesicles to Boost Skin Regeneration
2.2. Regulation of the Pigmentation Process by Exosomes
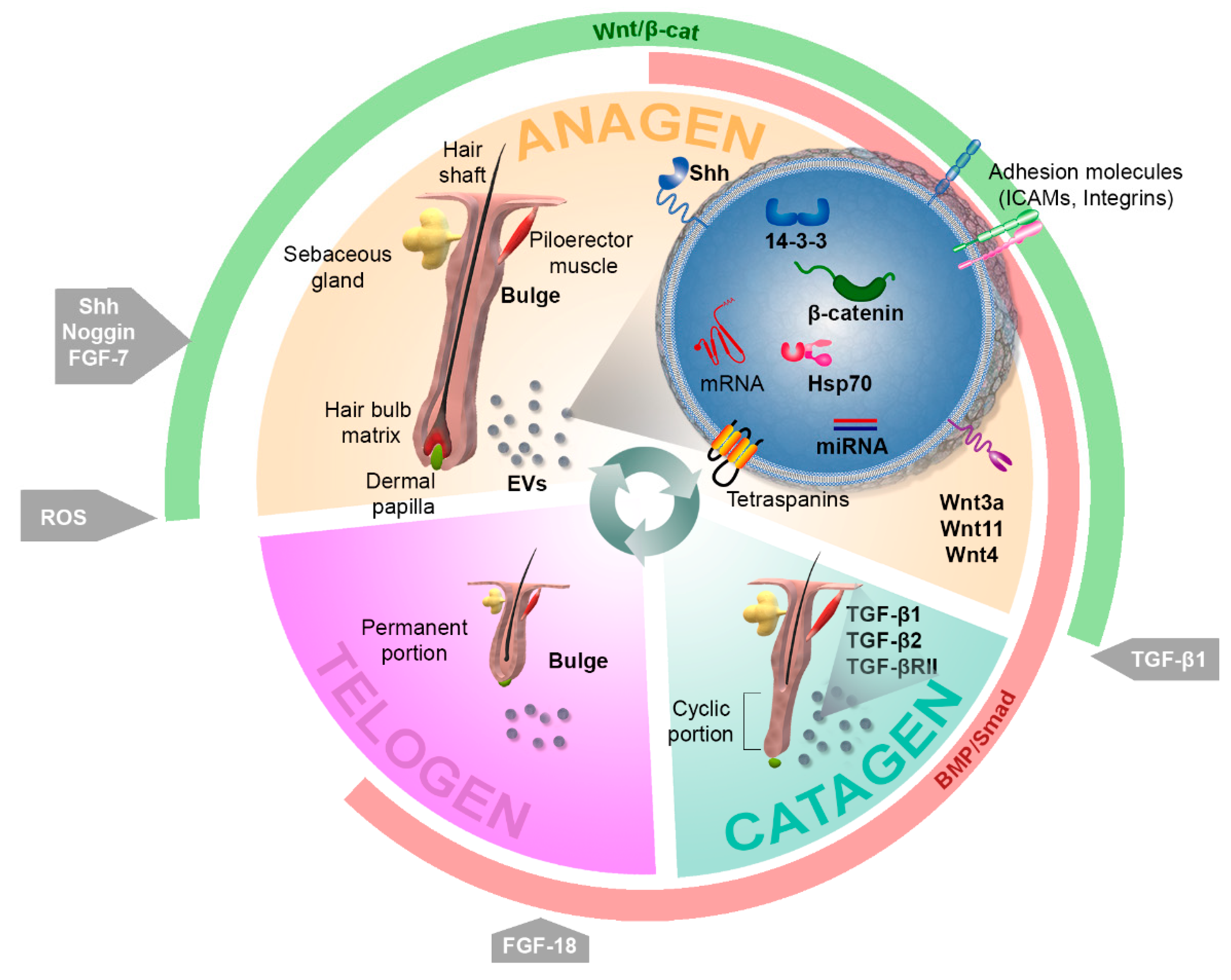
3. Role of Extracellular Vesicles in Hair Follicle Function
3.1. Effects of Extracellular Vesicles on Hair Follicle Dynamics
3.1.1. Exosomes as Signaling Mediators with the Potential to Modulate Hair Cycling
3.1.2. Use of Extracellular Vesicles to Stimulate Hair Growth: Evidence and Clues
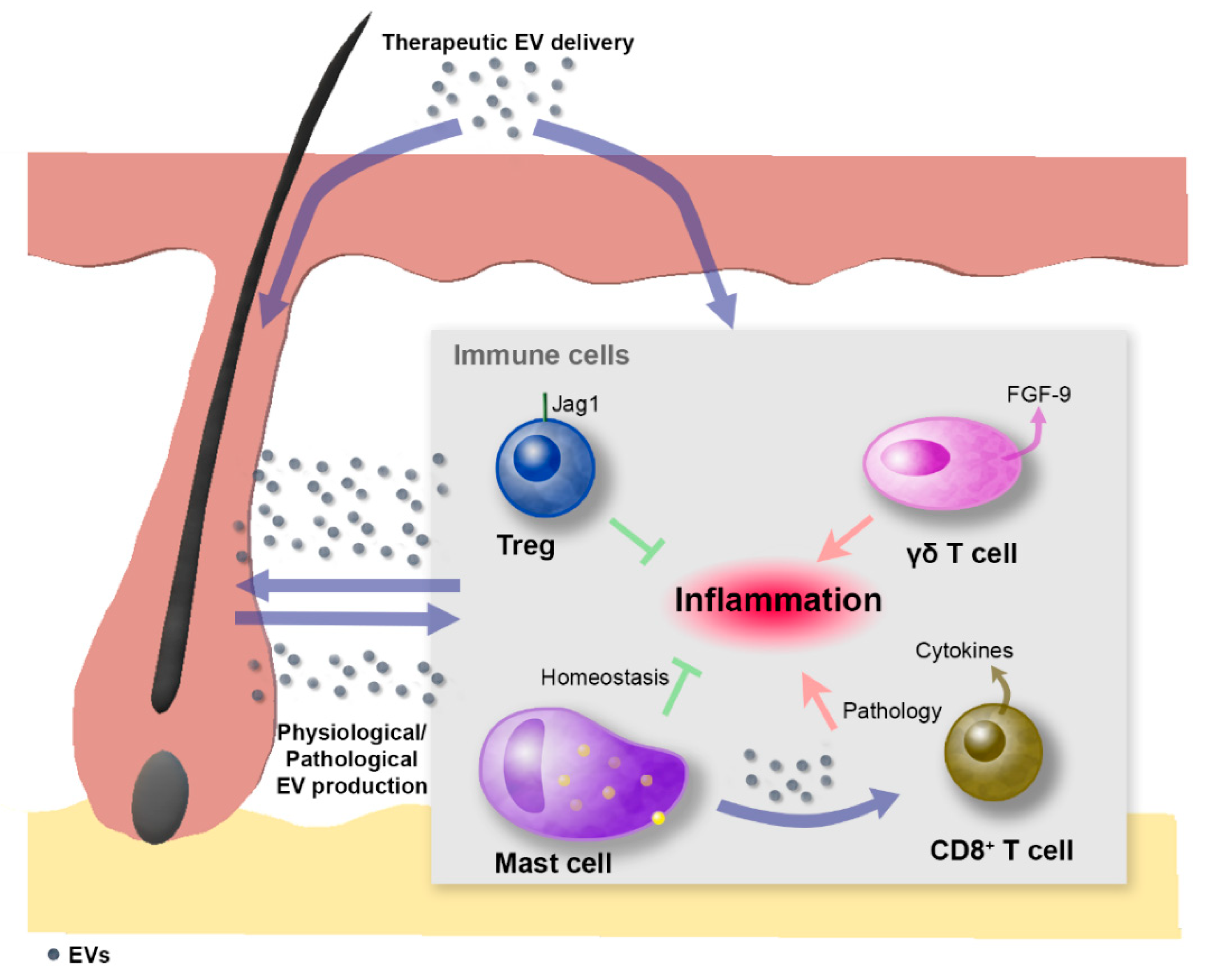
3.2. Immune System Cells and Hair Follicles
4. Concluding Remarks and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP | Bone Morphogenetic Protein |
| DAMPS | Danger-Associated Molecular Patterns |
| DP | Dermal Papilla |
| EGF | Epidermal Growth Factor |
| ESCRT | Endosomal Sorting Complexes Required for Transport |
| EV | Extracellular Vesicles |
| HF | Hair Follicle |
| HSP70 | Heat shock 70 kDa protein |
| miRNA | MicroRNA |
| MITF | Microphthalmia-associated transcription factor |
| MSC | Mesenchymal Stem Cell |
| MVB | Multivesicular Body |
| PGD2 | Prostaglandin D2 |
| SC | Stem Cell |
| Shh | Sonic Hedgehog |
| TERRA | Telomeric repeats-containing RNA |
| TGF | Transforming Growth Factor |
| UVB | Ultraviolet B |
References
- Clark, R.A.F.; Ghosh, K.; Tonnesen, M.G. Tissue Engineering for Cutaneous Wounds. J. Invest. Dermatol. 2007, 127, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H. The Secret Life of the Hair Follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef]
- Solanas, G.; Benitah, S.A. Regenerating the Skin: A Task for the Heterogeneous Stem Cell Pool and Surrounding Niche. Nat. Rev. Mol. Cell Biol. 2013, 14, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane Vesicles as Conveyors of Immune Responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, L.; Yu, H.; McKenzie, A.J.; Franklin, J.L.; Patton, J.G.; Liu, Q.; Weaver, A.M. Quantitative Proteomic Analysis of Small and Large Extracellular Vesicles (EVs) Reveals Enrichment of Adhesion Proteins in Small EVs. J. Proteome Res. 2019, 18, 947–959. [Google Scholar] [CrossRef]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Sánchez-Madrid, F. Intercellular Communication: Diverse Structures for Exchange of Genetic Information. Nat. Rev. Mol. Cell Biol. 2012, 13, 328–335. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Baixauli, F.; Gutiérrez-Vázquez, C.; Sánchez-Madrid, F.; Mittelbrunn, M. Sorting It out: Regulation of Exosome Loading. Semin. Cancer Biol. 2014, 28, 3–13. [Google Scholar] [CrossRef]
- Harding, C.; Heuser, J.; Stahl, P. Receptor-Mediated Endocytosis of Transferrin and of the Transferrin Receptor in Rat Reticulocytes Recycling. J. Cell. Biol. 1983, 97, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron Microscopic Evidence for Xternalization of the Transferrin Receptor in Vesicular Form in Sheep Reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.H.W.; Nijman, W.; Stoorvogel, R.; Liejendekker, C.V.; Harding, C.J.; Melief, J.G.B. Lymphocytes Secrete Antigen-PresentingVesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Zitvogel, L.; Regnault, A.; et al. Eradication of Established Murine Tumors Using a Novel Cell-Free Vaccine: Dendritic Cell-Derived Exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Cabral, J.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Extracellular Vesicles as Modulators of Wound Healing. Adv. Drug Deliv. Rev. 2018, 129, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Shaw, T.J.; Martin, P. Wound Repair at a Glance. J. Cell Sci. 2009, 122, 3209–3213. [Google Scholar] [CrossRef] [PubMed]
- Riazifar, M.; Pone, E.J.; Lötvall, J.; Zhao, W. Stem Cell Extracellular Vesicles: Extended Messages of Regeneration. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 125–154. [Google Scholar] [CrossRef] [PubMed]
- Than, U.T.T.; Guanzon, D.; Leavesley, D.; Parker, T. Association of Extracellular Membrane Vesicles with Cutaneous Wound Healing. Int. J. Mol. Sci. 2017, 18, 956. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal Stem Cell Exosomes Induce Proliferation and Migration of Normal and Chronic Wound Fibroblasts, and Enhance Angiogenesis In Vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guan, J.; Niu, X.; Hu, G.; Guo, S.; Li, Q.; Xie, Z.; Zhang, C.; Wang, Y. Exosomes Released from Human Induced Pluripotent Stem Cells-Derived MSCs Facilitate Cutaneous Wound Healing by Promoting Collagen Synthesis and Angiogenesis. J. Transl. Med. 2015, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-F.; Fan, J.; Fedesco, M.; Guan, S.; Li, Y.; Bandyopadhyay, B.; Bright, A.M.; Yerushalmi, D.; Liang, M.; Chen, M.; et al. Transforming Growth Factor (TGF )-Stimulated Secretion of HSP90: Using the Receptor LRP-1/CD91 To Promote Human Skin Cell Migration against a TGF -Rich Environment during Wound Healing. Mol. Cell. Biol. 2008, 28, 3344–3358. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.C.; Tao, S.C.; Yin, W.J.; Qi, X.; Yuan, T.; Zhang, C.Q. Exosomes Derived from Platelet-Rich Plasma Promote the Re-Epithelization of Chronic Cutaneous Wounds via Activation of YAP in a Diabetic Rat Model. Theranostics 2017, 7, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, M.; Gong, A.; Zhang, X.; Wu, X.; Zhu, Y.; Shi, H.; Wu, L.; Zhu, W.; Qian, H.; et al. HucMSc-Exosome Mediated-Wnt4 Signaling Is Required for Cutaneous Wound Healing. Stem Cells 2015, 33, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, C.; Zhao, J. Human Endothelial Progenitor Cells-Derived Exosomes Accelerate Cutaneous Wound Healing in Diabetic Rats by Promoting Endothelial Function. J. Diabetes Complicat. 2016, 30, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.-C.; Guo, S.-C.; Li, M.; Ke, Q.-F.; Guo, Y.-P.; Zhang, C.-Q. Chitosan Wound Dressings Incorporating Exosomes Derived from MicroRNA-126-Overexpressing Synovium Mesenchymal Stem Cells Provide Sustained Release of Exosomes and Heal Full-Thickness Skin Defects in a Diabetic Rat Model. Stem Cells Transl. Med. 2017, 6, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, C.; Hu, B.; Niu, X.; Liu, X.; Zhang, G.; Zhang, C.; Li, Q.; Wang, Y. Exosomes Derived from Human Endothelial Progenitor Cells Accelerate Cutaneous Wound Healing by Promoting Angiogenesis through Erk1/2 Signaling. Int. J. Biol. Sci. 2016, 12, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Walker, A.; Nissen, E. Human Fibrocyte-Derived Exosomes Accelerate Wound Healing in Genetically Diabetic Mice. Biochem. Biophys. Res. Commun. 2015, 467, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes Derived from Human Adipose Mensenchymal Stem Cells Accelerates Cutaneous Wound Healing via Optimizing the Characteristics of Fibroblasts. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-K.; Wu, S.-Y.; Tang, S.-E.; Li, M.-H.; Lin, S.-S.; Chu, S.-J.; Huang, K.-L. Protective Effects of Neural Crest-Derived Stem Cell-Conditioned Media against Ischemia-Reperfusion-Induced Lung Injury in Rats. Inflammation 2017, 40, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.D.; Rodriguez-Menocal, L.; Badiavas, E.V. Extracellular Vesicles as Biomarkers and Therapeutics in Dermatology: A Focus on Exosomes. J. Investig. Dermatol. 2017, 137, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Fisher, D.E. Melanocyte Biology and Skin Pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Denman, C.J.; McCracken, J.; Hariharan, V.; Klarquist, J.; Oyarbide-Valencia, K.; Guevara-Patĩo, J.A.; Caroline Le Poole, I. HSP70i Accelerates Depigmentation in a Mouse Model of Autoimmune Vitiligo. J. Investig. Dermatol. 2008, 128, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Lo Cicero, A.; Delevoye, C.; Gilles-Marsens, F.; Loew, D.; Dingli, F.; Guéré, C.; André, N.; Vié, K.; Van Niel, G.; Raposo, G. Exosomes Released by Keratinocytes Modulate Melanocyte Pigmentation. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Choi, S.H.; Kim, C.H.; Lee, C.H.; Lee, T.R.; Lee, A.Y. Reduced MiR-675 in Exosome in H19 RNA-Related Melanogenesis via MITF as a Direct Target. J. Investig. Dermatol. 2014, 134, 1075–1082. [Google Scholar] [CrossRef]
- Jiang, N.; Xiang, L.; He, L.; Yang, G.; Zheng, J.; Wang, C.; Zhang, Y.; Wang, S.; Zhou, Y.; Sheu, T.J.; et al. Exosomes Mediate Epithelium-Mesenchyme Crosstalk in Organ Development. ACS Nano 2017, 11, 7736–7746. [Google Scholar] [CrossRef]
- Chen, C.C.; Murray, P.J.; Jiang, T.X.; Plikus, M.V.; Chang, Y.T.; Lee, O.K.; Widelitz, R.B.; Chuong, C.M. Regenerative Hair Waves in Aging Mice and Extra-Follicular Modulators Follistatin, Dkk1, and Sfrp4. J. Investig. Dermatol. 2014, 134, 2086–2096. [Google Scholar] [CrossRef]
- Jahoda, C.A.B.; Horne, K.A.; Oliver, R.F. Induction of Hair Growth by Implantation of Cultured Dermal Papilla Cells. Nature 1984, 311, 560–562. [Google Scholar] [CrossRef]
- Reynolds, A.J.; Jahoda, C.A.B. Cultured Dermal Papilla Cells Induce Follicle Formation and Hair Growth by Transdifferentiation of an Adult Epidermis. Development 1992, 115, 587–593. [Google Scholar]
- Plikus, M.V.; Mayer, J.A.; De La Cruz, D.; Baker, R.E.; Maini, P.K.; Maxson, R.; Chuong, C.M. Cyclic Dermal BMP Signalling Regulates Stem Cell Activation during Hair Regeneration. Nature 2008, 451, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Millar, S.E. Molecular Mechanisms Regulating Hair Follicle Development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Myung, P.S.; Takeo, M.; Ito, M.; Atit, R.P. Epithelial Wnt Ligand Secretion Is Required for Adult Hair Follicle Growth and Regeneration. J. Investig. Dermatol. 2013, 133, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt Proteins Are Secreted on Exosomes. Nat. Cell Biol. 2012, 14, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.D.; Rodriguez-Menocal, L.; Guzman, W.; Candanedo, A.; Garcia-Contreras, M.; Badiavas, E.V. Bone Marrow Mesenchymal Stem Cell-Derived CD63 + Exosomes Transport Wnt3a Exteriorly and Enhance Dermal Fibroblast Proliferation, Migration, and Angiogenesis In Vitro. Stem Cells Dev. 2017, 26, 1384–1398. [Google Scholar] [CrossRef] [PubMed]
- Gangoda, L.; Boukouris, S.; Liem, M.; Kalra, H.; Mathivanan, S. Extracellular Vesicles Including Exosomes Are Mediators of Signal Transduction: Are They Protective or Pathogenic? Proteomics 2015, 15, 260–271. [Google Scholar] [CrossRef]
- Menck, K.; Klemm, F.; Gross, J.C.; Pukrop, T.; Wenzel, D.; Binder, C. Induction and Transport of Wnt 5a during Macrophage-Induced Malignant Invasion Is Mediated by Two Types of Extracellular Vesicles. Oncotarget 2015, 4, 2057. [Google Scholar] [CrossRef] [PubMed]
- Dovrat, S.; Caspi, M.; Zilberberg, A.; Lahav, L.; Firsow, A.; Gur, H.; Rosin-Arbesfeld, R. 14-3-3 and β-Catenin Are Secreted on Extracellular Vesicles to Activate the Oncogenic Wnt Pathway. Mol. Oncol. 2014, 8, 894–911. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wu, X.; Zhang, X.; Sun, Y.; Yan, Y.; Shi, H.; Zhu, Y.; Wu, L.; Pan, Z.; Zhu, W.; et al. Human Umbilical Cord Mesenchymal Stem Cell Exosomes Enhance Angiogenesis Through the Wnt4/β-Catenin Pathway. Stem Cells Transl. Med. 2015, 4, 513–522. [Google Scholar] [CrossRef]
- Shi, H.; Xu, X.; Zhang, B.; Xu, J.; Pan, Z.; Gong, A.; Zhang, X.; Li, R.; Sun, Y.; Yan, Y.; et al. 3,3′-Diindolylmethane Stimulates Exosomal Wnt11 Autocrine Signaling in Human Umbilical Cord Mesenchymal Stem Cells to Enhance Wound Healing. Theranostics 2017, 7, 1674–1688. [Google Scholar] [CrossRef]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes Mediate Stromal Mobilization of Autocrine Wnt-PCP Signaling in Breast Cancer Cell Migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Takada, R.; Noda, C.; Kobayashi, S.; Takada, S. Different Populations of Wnt-Containing Vesicles Are Individually Released from Polarized Epithelial Cells. Sci. Rep. 2016, 6, 35562. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.L.; Gangadaran, P.; Bak, S.S.; Oh, J.M.; Kalimuthu, S.; Lee, H.W.; Baek, S.H.; Zhu, L.; Sung, Y.K.; Jeong, S.Y.; et al. Extracellular Vesicles Derived from MSCs Activates Dermal Papilla Cell in Vitro and Promotes Hair Follicle Conversion from Telogen to Anagen in Mice. Sci. Rep. 2017, 7, 15560. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Regulation of Hair Follicle Development by Exosomes Derived from Dermal Papilla Cells. Biochem. Biophys. Res. Commun. 2018, 500, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Gradilla, A.C.; González, E.; Seijo, I.; Andrés, G.; Bischoff, M.; González-Mendez, L.; Sánchez, V.; Callejo, A.; Ibáñez, C.; Guerra, M.; et al. Exosomes as Hedgehog Carriers in Cytoneme-Mediated Transport and Secretion. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.J.; Brown, D.A.; Ibrahim, M.M.; Ramchal, T.D.; Levinson, H. MicroRNAs in Skin Tissue Engineering. Adv. Drug Deliv. Rev. 2015, 88, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Ning, M.S.; Andl, T. Control by a Hair’s Breadth: The Role of MicroRNAs in the Skin. Cell. Mol. Life Sci. 2013, 70, 1149–1169. [Google Scholar] [CrossRef]
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.N.; Botchkareva, N.V. MicroRNA-214 Controls Skin and Hair Follicle Development by Modulating the Activity of the Wnt Pathway. J. Cell Biol. 2014, 207, 549–567. [Google Scholar] [CrossRef]
- Li, X.; Liu, L.; Yang, J.; Yu, Y.; Chai, J.; Wang, L.; Ma, L.; Yin, H. Exosome Derived From Human Umbilical Cord Mesenchymal Stem Cell Mediates MiR-181c Attenuating Burn-Induced Excessive Inflammation. EBioMedicine 2016, 8, 72–82. [Google Scholar] [CrossRef]
- Harel, S.; Higgins, C.A.; Cerise, J.E.; Dai, Z.; Chen, J.C.; Clynes, R.; Christiano, A.M. Pharmacologic Inhibition of JAK-STAT Signaling Promotes Hair Growth. Sci. Adv. 2015, 1, e1500973. [Google Scholar] [CrossRef]
- Luetteke, N.C.; Qiu, T.H.; Peiffer, R.L.; Oliver, P.; Smithies, O.; Lee, D.C. TGFα Deficiency Results in Hair Follicle and Eye Abnormalities in Targeted and Waved-1 Mice. Cell 1993, 73, 263–278. [Google Scholar] [CrossRef]
- Mann, G.B.; Fowler, K.J.; Gabriel, A.; Nice, E.C.; Williams, R.L.; Dunn, A.R. Mice with a Null Mutation of the TGFα Gene Have Abnormal Skin Architecture, Wavy Hair, and Curly Whiskers and Often Develop Corneal Inflammation. Cell 1993, 73, 249–261. [Google Scholar] [CrossRef]
- Foitzik, K.; Lindner, G.; Mueller-Roever, S.; Maurer, M.; Botchkareva, N.; Botchkarev, V.; Handjiski, B.; Metz, M.; Hibino, T.; Soma, T.; et al. Control of Murine Hair Follicle Regression (Catagen) by TGF-Beta1 in Vivo. FASEB J. 2000, 14, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Soma, T.; Tsuji, Y.; Hibino, T. Involvement of Transforming Growth Factor-Beta2 in Catagen Induction during the Human Hair Cycle. J. Investig. Dermatol. 2002, 118, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.K.; Yeo, M.S.W.; Sheng, C.T.; Chai, L.R. Use of Exosomes to Promote or Enhance Hair Growth. Patent EP2629782A1, 22 January 2015. [Google Scholar]
- Carrasco, E.; Calvo, M.I.; Blázquez-Castro, A.; Vecchio, D.; Zamarrón, A.; De Almeida, I.J.D.; Stockert, J.C.; Hamblin, M.R.; Juarranz, Á.; Espada, J. Photoactivation of ROS Production in Situ Transiently Activates Cell Proliferation in Mouse Skin and in the Hair Follicle Stem Cell Niche Promoting Hair Growth and Wound Healing. J. Invest. Dermatol. 2015, 135, 2611–2622. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yang, Z.; Andl, T.; Cui, C.; Kim, N.; Millar, S.E.; Cotsarelis, G. Wnt-Dependent de Novo Hair Follicle Regeneration in Adult Mouse Skin after Wounding. Nature 2007, 447, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Myung, P.; Ito, M. Dissecting the Bulge in Hair Regeneration. J. Clin. Investig. 2012, 122, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Tong, C.; Hou, Q.; Han, Q.; Zhao, Y.; Liu, H.; et al. Treatment of MSCs with Wnt1a-Conditioned Medium Activates DP Cells and Promotes Hair Follicle Regrowth. Sci. Rep. 2014, 4, 5432. [Google Scholar] [CrossRef] [PubMed]
- Gutkin, A.; Uziel, O.; Beery, E.; Nordenberg, J.; Pinchasi, M.; Goldvaser, H.; Henick, S.; Goldberg, M.; Lahav, M.; Gutkin, A.; et al. Tumor Cells Derived Exosomes Contain HTERT MRNA and Transform Nonmalignant Fibroblasts into Telomerase Positive Cells. Oncotarget 2016, 7, 59173–59188. [Google Scholar] [CrossRef]
- Sarin, K.Y.; Cheung, P.; Gilison, D.; Lee, E.; Tennen, R.I.; Wang, E.; Artandi, M.K.; Oro, A.E.; Artandi, S.E. Conditional Telomerase Induction Causes Proliferation of Hair Follicle Stem Cells. Nature 2005, 436, 1048–1052. [Google Scholar] [CrossRef]
- Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; Mittelbrunn, M.; Sánchez-Madrid, F. Transfer of Extracellular Vesicles during Immune Cell-Cell Interactions. Immunol. Rev. 2013, 251, 125–142. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Deregibus, M.C.; Camussi, G. The Secretome of Mesenchymal Stromal Cells: Role of Extracellular Vesicles in Immunomodulation. Immunol. Lett. 2015, 168, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Gehrmann, M.; Brunet, M.; Multhoff, G.; Garrido, C. Intracellular and Extracellular Functions of Heat Shock Proteins: Repercussions in Cancer Therapy. J. Leukoc. Biol. 2007, 81, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Gay, D.; Kwon, O.; Zhang, Z.; Spata, M.; Plikus, M.V.; Holler, P.D.; Ito, M.; Yang, Z.; Treffeisen, E.; Kim, C.D.; et al. Fgf9 from Dermal Γδ T Cells Induces Hair Follicle Neogenesis after Wounding. Nat. Med. 2013, 19, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Zirak, B.; Rodriguez, R.S.; Pauli, M.L.; Truong, H.A.; Lai, K.; Ahn, R.; Corbin, K.; Lowe, M.M.; Scharschmidt, T.C.; et al. Regulatory T Cells in Skin Facilitate Epithelial Stem Cell Differentiation. Cell 2017, 169, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.M.; Loy, D.E.; Lawson, J.A.; Katseff, A.S.; Fitzgerald, G.A.; Garza, L.A. Prostaglandin D2 Inhibits Wound-Induced Hair Follicle Neogenesis through the Receptor, Gpr44. J. Invest. Dermatol. 2013, 133, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Bertolini, M.; Zilio, F.; Rossi, A.; Kleditzsch, P.; Emelianov, V.E.; Gilhar, A.; Keren, A.; Meyer, K.C.; Wang, E.; Funk, W.; et al. Abnormal Interactions between Perifollicular Mast Cells and CD8+ T-Cells May Contribute to the Pathogenesis of Alopecia Areata. PLoS ONE 2014, 9, e94260. [Google Scholar] [CrossRef]
- Wang, Z.; Lieberman, P.M. The Crosstalk of Telomere Dysfunction and Inflammation through Cell-Free TERRA Containing Exosomes. RNA Biol. 2016, 13, 690–695. [Google Scholar] [CrossRef]
- Flores, I.; Canela, A.; Vera, E.; Tejera, A.; Cotsarelis, G.; Blasco, M.A. The Longest Telomeres: A General Signature of Adult Stem Cell Compartments. Genes Dev. 2008, 22, 654–667. [Google Scholar] [CrossRef]
- Lindner, G.; Botchkarev, V.A.; Botchkareva, N.V.; Ling, G.; Van der Veen, C.; Paus, R. Analysis of Apoptosis during Hair Follicle Regression (Catagen). Am. J. Pathol. 1997, 151, 1601–1617. [Google Scholar]
- Haake, A.R.; Polakowska, R.R. Cell Death by Apoptosis in Epidermal Biology. J. Investig. Dermatol. 1993, 101, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Magerl, M.; Tobin, D.J.; Müller-Röver, S.; Hagen, E.; Lindner, G.; McKay, I.A.; Paus, R. Patterns of Proliferation and Apoptosis during Murine Hair Follicle Morphogenesis. J. Invest. Dermatol. 2001, 116, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J.; Hagen, E.; Botchkarev, V.A.; Paus, R. Do Hair Bulb Melanocytes Undergo Apotosis during Hair Follicle Regression (Catagen)? J. Investig. Dermatol. 1998, 111, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Higginbotham, J.N.; Jeppesen, D.K.; Yang, Y.-P.; Li, W.; McKinley, E.T.; Graves-Deal, R.; Ping, J.; Britain, C.M.; Dorsett, K.A.; et al. Transfer of Functional Cargo in Exomeres. Cell Rep. 2019, 27, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Deschene, E.R.; Zito, G.; Gonzalez, D.G.; Saotome, I.; Haberman, A.M.; Greco, V. Live Imaging of Stem Cell and Progeny Behaviour in Physiological Hair-Follicle Regeneration. Nature 2012, 487, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Mesa, K.R.; Greco, V. Spatial Organization within a Niche as a Determinant of Stem-Cell Fate. Nature 2013, 502, 513–518. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Signaling Pathway | Molecules Transported via EVs | Source of EVs | Highlights of the Study | Model Used to Test the Effects | Ref. |
|---|---|---|---|---|---|
| Canonical Wnt | β-catenin and 14-3-3 proteins | HEK293T, SW480 | EV-mediated activation of Wnt signaling in recipient cells | In vitro: HEK293T, COS7, SW480 | [48] |
| Wnt4 | HuUC-MSCs | HuUC-MSC exosomes facilitated wound re-epithelization and cell proliferation through the activation of Wnt signaling | In vitro: HaCaT, Ea.hy926, rat dermal fibroblasts In vivo: Rat skin 2nd degree burn injury | [25,49] | |
| Wnt11 | HuUC-MSCs | Exosomal Wnt11 autocrine signaling in response to 3-3′-diindolylmethane increased markers of stemness in MSCs and favored wound healing | In vitro: HaCaT, rat dermal fibroblasts In vivo: Rat skin 2nd degree burn injury | [50] | |
| Wnt3a, Wnt11 | MDCK, HEK293, fibroblast L cells | Different populations of exosomes carrying Wnt factors secreted by epithelial cells depending on the cell polarity and cell type | [52] | ||
| Wnt3a, Wnt5a | Mouse BM-MSCs | EVs contributed to hair growth in mice by promoting telogen to anagen conversion of HFs | In vivo: Mouse skin | [53] | |
| Wnt-planar cell polarity | Wnt11 | Mouse fibroblast L cells | Mouse fibroblast-derived exosomes mobilized Wnt11-mediated autocrine signaling, promoting protrusive activity and motility | In vitro: MDA-MB-231 In vivo: SCID mice | [51] |
| Canonical Wnt; Shh | Not characterized | HuDPCs | Exosomes extended the anagen phase of the hair cycle in mice by inducing the expression of β-catenin and Shh | In vivo: Mouse skin | [54] |
| Hh | Hh | Drosophila | Hh transport via exosomes along cytonems | In vitro: Cl8 | [55] |
| TLR4 | miR-181c | HuUC-MSCs | Exosomes overexpressing miR-181c reduced burn inflammation by downregulating the TLR4 signaling pathway | In vivo: Rat full-thickness burn injury | [59] |
| EGF/EGFR | mi-126-3p | HuS-MSCs | Improvement in the healing capacity of wound dressings by incorporating exosomes derived from miR126-overexpressing HuS-MSCs, which led to the activation of AKT and ERK1/2 through phosphorylation | In vitro: Human dermal fibroblast, HMEC-1 In vivo: Full-thickness excisional skin wound in diabetic rats | [27] |
| ERK1/2 | BM-MSCs | Key pathways for wound healing including Akt, ERK, and STAT3, activated by MSC-exosomes | In vitro: Diabetic versus normal wound patient fibroblasts | [21] | |
| ERK1/2 | HuEPCs | ERK1/2-mediated improved angiogenesis in response to exosomes with beneficial effects on wound healing | In vitro: HMEC-1 In vivo: Full-thickness excisional skin wound in diabetic rats | [28] | |
| TGF-α | HKCs | Stimulation of the secretion of hsp90α in exosomes by HuK-promoted migration of both epidermal and dermal cells | In vitro: Primary neonatal HKCs, dermal cells | [23] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrasco, E.; Soto-Heredero, G.; Mittelbrunn, M. The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics. Int. J. Mol. Sci. 2019, 20, 2758. https://doi.org/10.3390/ijms20112758
Carrasco E, Soto-Heredero G, Mittelbrunn M. The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics. International Journal of Molecular Sciences. 2019; 20(11):2758. https://doi.org/10.3390/ijms20112758
Chicago/Turabian StyleCarrasco, Elisa, Gonzalo Soto-Heredero, and María Mittelbrunn. 2019. "The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics" International Journal of Molecular Sciences 20, no. 11: 2758. https://doi.org/10.3390/ijms20112758
APA StyleCarrasco, E., Soto-Heredero, G., & Mittelbrunn, M. (2019). The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics. International Journal of Molecular Sciences, 20(11), 2758. https://doi.org/10.3390/ijms20112758