Genetic and Molecular Control of Floral Organ Identity in Cereals

 ,
,  ,
, 
Abstract
1. Introduction
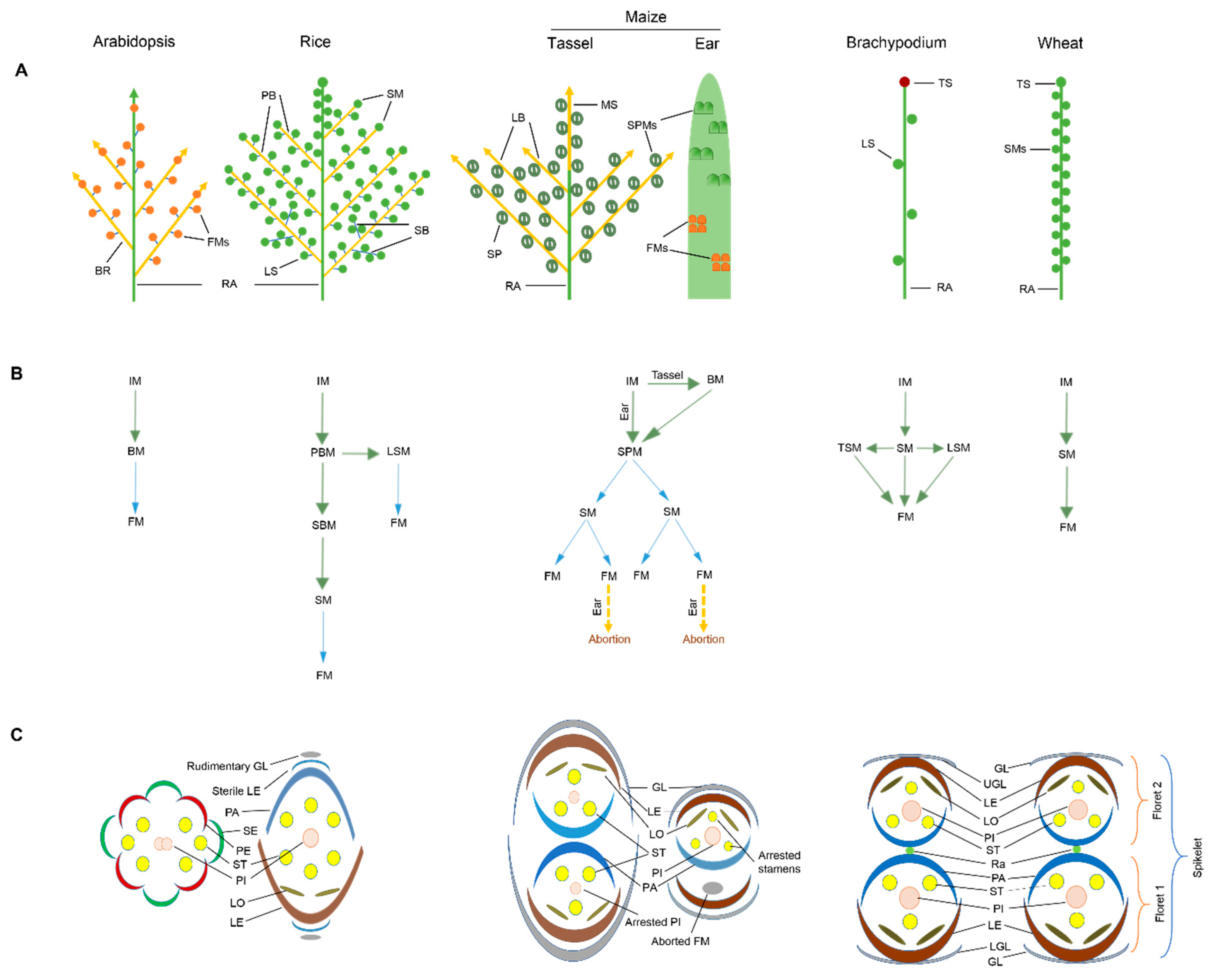
2. Inflorescence Morphology and Development
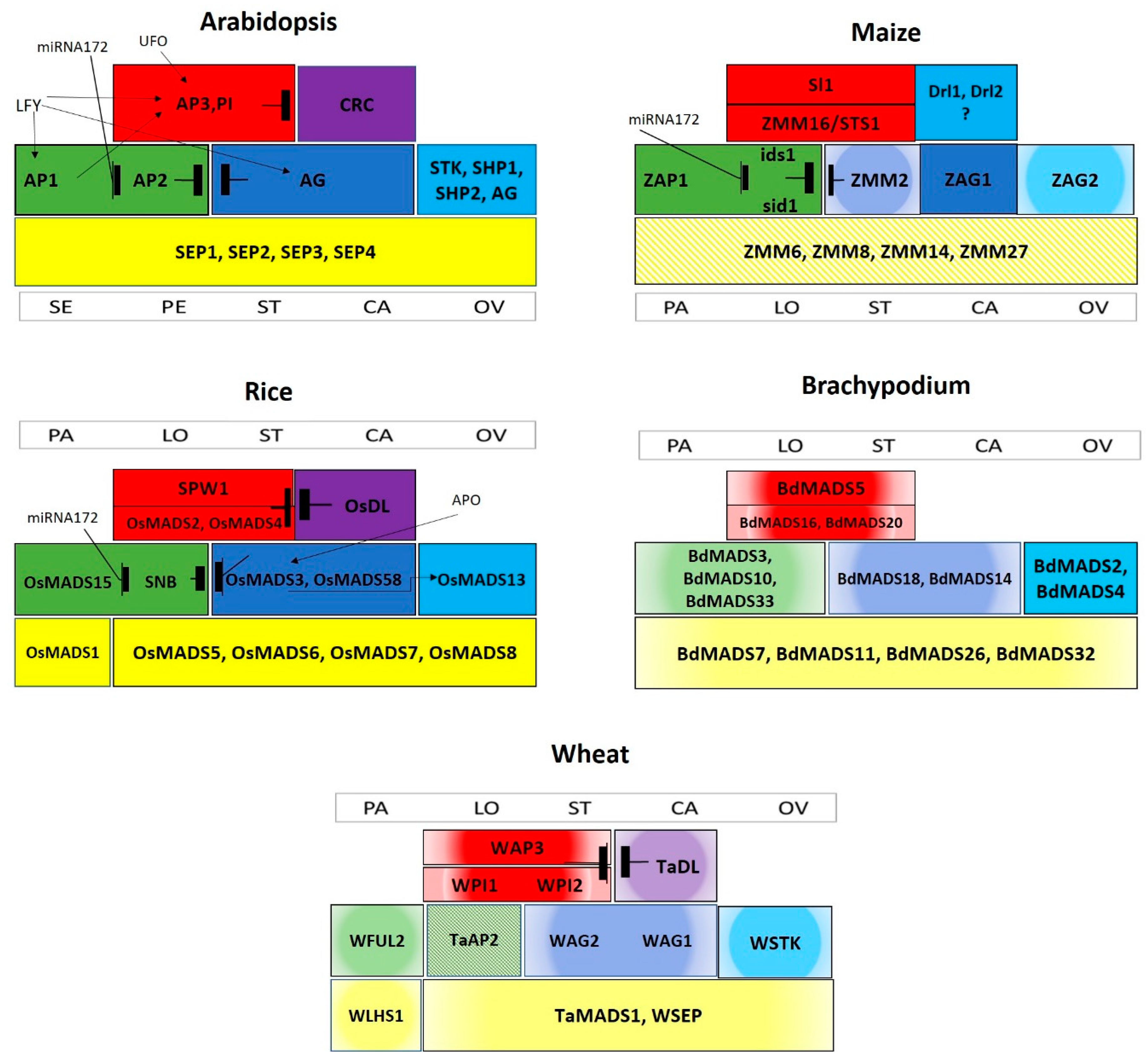
3. Role of MADS-Box Transcription Factors in Floral Organ Identity
3.1. Class A Homeotic Genes
3.2. Class B Homeotic Genes
3.3. Class C and D homeotic genes
3.4. Class E Homeotic Genes
4. Non MADS-Box Genes Involved In Floral Organ Identity
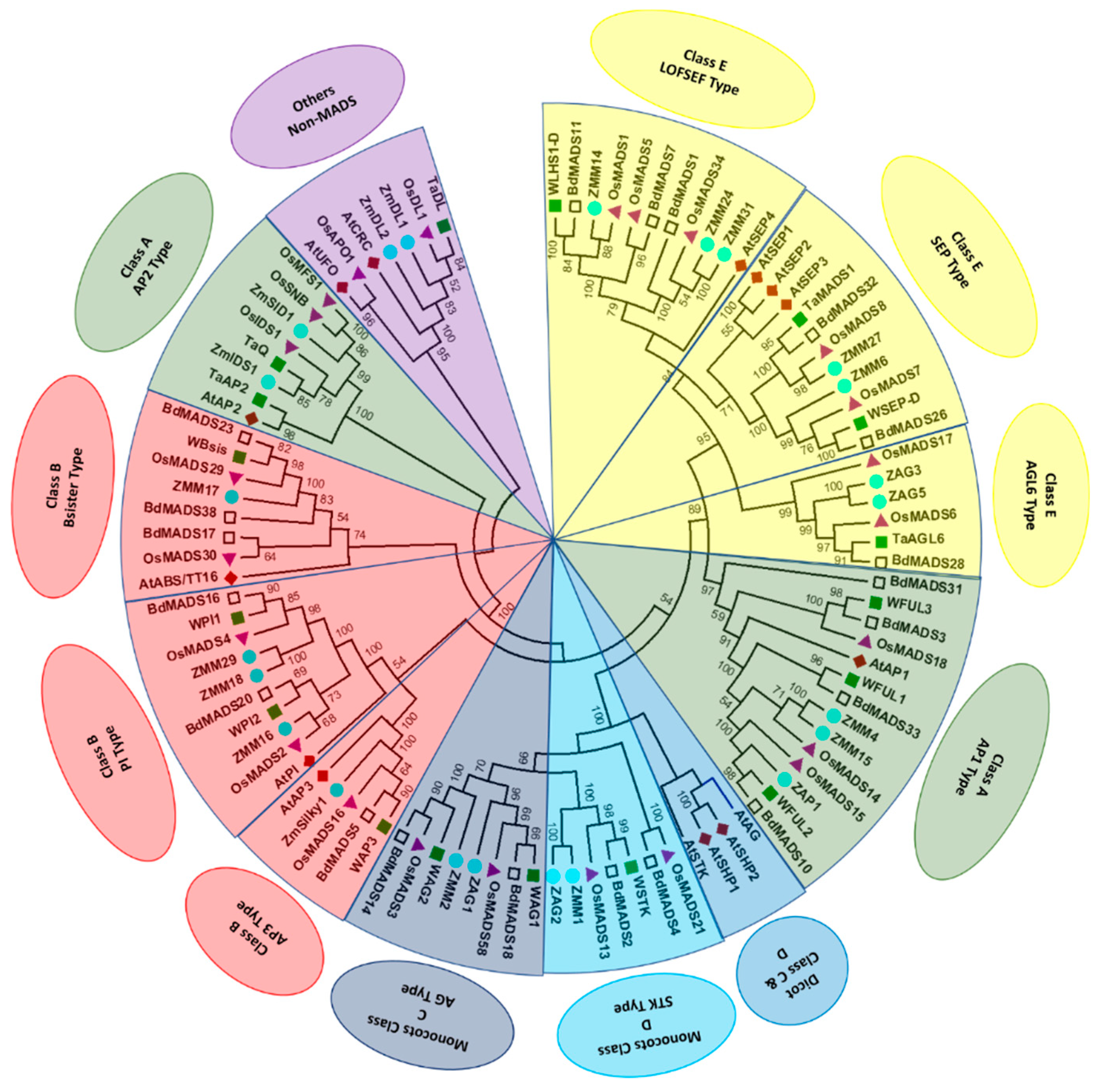
5. Functional Conservation and Diversification between Distinct Floral Specification Systems
6. Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABS | ARABIDOPSIS BSISTER |
| AG | AGAMOUS |
| AGL6 | AGAMOUS-LIKE 6 |
| AP1 | APETALA1 |
| AP2 | APETALA2 |
| AP3 | APETALA3 |
| APO1 | Aberrant panicle organization-1 |
| BD1 | BRANCHED SILKLESS1 |
| BdMADS | Brachypodium MADS |
| BM | Branch meristem |
| BR | Branch |
| CA | Carpel |
| CAL | CAULIFLOWER |
| CFO1 | CHIMERIC FLORAL ORGANS 1 |
| CRC | CRABSCLAW |
| DL | Drooping leaf |
| ELE | ELONGATED EMPTY GLUME |
| FBP11 | FLORAL BINDINGPROTEIN 11 |
| FBP7 | FLORAL BINDINGPROTEIN 7 |
| FLO/LFY | FLORICAULA/ LEAFY |
| FM | Floral meristem |
| FUL1 | FRUITFULL 1 |
| FUL2 | FRUITFULL 2 |
| FUL3 | FRUITFULL 3 |
| FZP | FRIZZY PANICLE |
| G1 | LONG STERILE LEMMA1 |
| GL | Glume |
| GOA | GORDITA |
| HvAP2/Cly1 | Hordeum vulgare APETALA2/Cleistogamy1 |
| IDS1 | INDETERMINATE SPIKELET1 |
| IM | Inflorescence meristem |
| ld-1 | lemma-distortion 1 |
| Le | Lemma |
| LHS1 | LEAFY HULL STERILE 1 |
| LO | Lodicule |
| LS | Lateral spikelet |
| LSM | Lateral spikelet meristem |
| MADS | MINICHROMOSOME MAINTENANCE 1, AGAMOUS, DEFICIENS, and SERUM RESPONSE FACTOR |
| MEGA6 | Molecular Evolutionary Genetic Analysis version 6 |
| MFO1 | MOSAIC FLORAL ORGANS 1 |
| MFS1 | MULTI-FLORET SPIKELET 1 |
| miRNA | micro-RNA |
| MOS1 | MORE SPIKELET 1 |
| mRNA | Messenger RNA |
| NCBI | National Center for Biotechnology Information |
| OsIDS1 | Oryza sativa INDETERMINATE SPIKELET 1 |
| OsMADS | Oryza sative MADS |
| OV | Ovule |
| PA | Palea |
| PAP2 | PANICLE PHYTOMER2 |
| PB | Primary branch |
| PBM/pBM | Primary branch meristem |
| PE | Petal |
| PI | PISTILLATA |
| PI | Pistil |
| RA | Rachis |
| Ra | Rachilla |
| RFL | RICE FLORICAULA |
| RNAi | RNA interference |
| RT-PCR | Reverse transcriptase-polymerase chain reaction |
| SAM | Shoot apical meristem |
| SB | Secondary branch |
| SBM/sBM | Secondary branch meristem |
| SBP/SPL | SQUAMOSA PROMOTER BINDING PROTEIN-LIKE |
| SE | Sepal |
| SEP | SEPALLATA |
| SHP | SHATTERPROOF |
| SID1 | SISTER OF INDETERMINATE SPIKELET1 |
| Sl1 | SILKY1 |
| SM | Spikelet meristem |
| SNB | SUPERNUMERARY BRACT |
| SPM | Spikelet pair meristem |
| SPW1 | SUPERWOMAN1 |
| ST | Stamen |
| STK | SEEDSTICK |
| STS1 | Sterile Tassel Silky Ear1 |
| TaAG3 | Triticum aestivum AGAMOUS3 |
| TaAP3 | Triticum aestivum AP3 |
| TaDL | Triticum aestivum Drooping Leaf |
| TaMADS | Triticum aestivum MADS |
| TFs | Transcription factors |
| TS | Terminal spikelet |
| TSM | Terminal spikelet meristem |
| TT16 | TRANSPARENT TESTA 16 |
| UFO | UNUSUAL FLORAL ORGANS |
| VRN1 | VERNALIZATION 1 |
| WAG-1 | Wheat AGAMOUS-1 |
| WAG-2 | Wheat AGAMOUS-2 |
| WAP1 | Wheat APETALA1 |
| WAP3 | Wheat APETALA3 |
| WBsis | Wheat BSISTER |
| WFUL1 | Wheat FRUITFULL 1 |
| WFUL2 | Wheat FRUITFULL 2 |
| WFUL3 | Wheat FRUITFULL 3 |
| WLHS1 | Wheat LEAFY HULL STERILE 1 |
| WPl1 | Wheat PISTILLATA 1 |
| WPI2 | Wheat PISTILLATA 2 |
| WSEP | Wheat SEPALLATA |
| WSTK | Wheat SEEDSTICK |
| ZAG1 | Zea mays AGAMOUS1 |
| ZAG2 | Zea mays AGAMOUS2 |
| ZAP1 | Zea mays APETALA1 |
| ZmIDS1 | Zea mays INDETERMINATE SPIKELET1 |
| ZMM | Zea mays MADS |
References
- Awika, J.M. Major cereal grains production and use around the world. In Advances in Cereal Science: Implications to Food Processing and Health Promotion; ACS Publications: Washington, DC, USA, 2011; pp. 1–13. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Zuloaga, F.O.; Judziewicz, E.J.; Filgueiras, T.S.; Davis, J.I.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Bell, C.D.; Soltis, D.E.; Soltis, P.S. The age and diversification of the angiosperms re-revisited. Am. J. Bot. 2010, 97, 1296–1303. [Google Scholar] [CrossRef] [PubMed]
- Soltis, D.E.; Bell, C.D.; Kim, S.; Soltis, P.S. Origin and early evolution of angiosperms. Ann. NY Acad. Sci. 2008, 1133, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Ambrose, B.A. The blooming of grass flower development. Curr. Opin. Plant Biol. 1998, 1, 60–67. [Google Scholar] [CrossRef]
- Bommert, P.; Satoh-Nagasawa, N.; Jackson, D.; Hirano, H.-Y. Genetics and evolution of inflorescence and flower development in grasses. Plant Cell Physiol. 2005, 46, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, F.; Yoshida, H. Interpreting lemma and palea homologies: A point of view from rice floral mutants. Front. Plant Sci. 2015, 6, 61. [Google Scholar] [CrossRef]
- Goff, S.A.; Ricke, D.; Lan, T.-H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.-S.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Itoh, J.-I.; Nonomura, K.-I.; Ikeda, K.; Yamaki, S.; Inukai, Y.; Yamagishi, H.; Kitano, H.; Nagato, Y. Rice plant development: From zygote to spikelet. Plant Cell Physiol. 2005, 46, 23–47. [Google Scholar] [CrossRef]
- Kurata, N.; Miyoshi, K.; Nonomura, K.-I.; Yamazaki, Y.; Ito, Y. Rice mutants and genes related to organ development, morphogenesis and physiological traits. Plant Cell Physiol. 2005, 46, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Wu, J.; Kanamori, H.; Katayose, Y. The map-based sequence of the rice genome. Nature 2005, 436, 793. [Google Scholar]
- Jiao, Y.; Peluso, P.; Shi, J.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.; Chin, C.-S. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Initiative, I.B. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763. [Google Scholar]
- Brkljacic, J.; Grotewold, E.; Scholl, R.; Mockler, T.; Garvin, D.F.; Vain, P.; Brutnell, T.; Sibout, R.; Bevan, M.; Budak, H. Brachypodium as a model for the grasses: Today and the future. Plant Physiol. 2011, 157, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Draper, J.; Mur, L.A.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; Routledge, A.P. Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Garvin, D.F.; Mockler, T.C.; Schmutz, J.; Rokhsar, D.; Bevan, M.W.; Barry, K.; Lucas, S.; Harmon-Smith, M.; Lail, K. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Consortium, I.W.G.S. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Jia, J.; Zhao, S.; Kong, X.; Li, Y.; Zhao, G.; He, W.; Appels, R.; Pfeifer, M.; Tao, Y.; Zhang, X. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature 2013, 496, 7443. [Google Scholar] [CrossRef]
- IWGSC, I. Shifting the limits in wheat research and breeding using a fully annotated reference genome by the international wheat genome sequencing consortium (iwgsc). Science 2018, 361. [Google Scholar]
- Derbyshire, P.; Byrne, M.E. MORE SPIKELETS1 is required for spikelet fate in the inflorescence of Brachypodium. Plant Physiol. 2013, 161, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N. FRIZZY PANICLE drives supernumerary spikelets in bread wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Song, G.; Guan, J.; Chen, K.; Jia, M.; Huang, D.; Wu, J.; Zhang, L.; Kong, X.; Geng, S. Transcriptome profiling of wheat inflorescence development from spikelet initiation to floral patterning identified stage-specific regulatory genes. Plant Physiol. 2017, 174, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Gauley, A.; Boden, S.A. Genetic pathways controlling inflorescence architecture and development in wheat and barley. J. Integr. Plant Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Koppolu, R.; Schnurbusch, T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. J. Integr. Plant Biol. 2019, 61, 278–295. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.Y.; Hu, Z.; Yang, R.-C. Genome-wide comparative analysis of flowering-related genes in Arabidopsis, wheat, and barley. Int. J. Plant Genom. 2015, 874361. [Google Scholar] [CrossRef]
- Wei, B.; Liu, D.; Guo, J.; Leseberg, C.H.; Zhang, X.; Mao, L. Functional divergence of two duplicated D-lineage MADS-box genes BdMADS2 and BdMADS4 from Brachypodium distachyon. J. Plant Physiol. 2013, 170, 424–431. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, R.-Z.; Guo, J.-J.; Liu, D.-M.; Li, A.-L.; Fan, R.-C.; Mao, L.; Zhang, X.-Q. Genome-wide analysis of the MADS-box gene family in Brachypodium distachyon. PLoS ONE 2014, 9, e84781. [Google Scholar] [CrossRef]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Colombo, L.; Franken, J.; Koetje, E.; van Went, J.; Dons, H.; Angenent, G.C.; van Tunen, A.J. The petunia MADS box gene FBP11 determines ovule identity. Plant Cell 1995, 7, 1859–1868. [Google Scholar] [CrossRef]
- Ditta, G.; Pinyopich, A.; Robles, P.; Pelaz, S.; Yanofsky, M.F. The SEP4 gene of Arabidopsis thaliana functions in floral organ and meristem identity. Curr. Biol. 2004, 14, 1935–1940. [Google Scholar] [CrossRef]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Theißen, G. Development of floral organ identity: Stories from the MADS house. Curr. Opin. Plant Biol. 2001, 4, 75–85. [Google Scholar] [CrossRef]
- Ciaffi, M.; Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E. Molecular aspects of flower development in grasses. Sex. Plant Reprod. 2011, 24, 247–282. [Google Scholar] [CrossRef]
- Chongloi, G.L.; Prakash, S.; Vijayraghavan, U. Rice shoot and floral meristem development: An overview of developmental regulators of meristem maintenance and organ identity. J. Exp. Bot. 2019, 70, 1719–1736. [Google Scholar] [CrossRef]
- Schnurbusch, T. Wheat and Barley Biology: Towards new frontiers. J. Integr. Plant Biol. 2019, 61, 198–203. [Google Scholar] [CrossRef]
- Zhang, D.; Yuan, Z. Molecular control of grass inflorescence development. Ann. Rev. Plant Biol. 2014, 65, 553–578. [Google Scholar] [CrossRef]
- Benlloch, R.; Berbel, A.; Serrano-Mislata, A.; Madueño, F. Floral initiation and inflorescence architecture: A comparative view. Ann. Bot. 2007, 100, 659–676. [Google Scholar] [CrossRef]
- Doust, A. Architectural evolution and its implications for domestication in grasses. Ann. Bot. 2007, 100, 941–950. [Google Scholar] [CrossRef]
- Kirby, E. Botany of the wheat plant. In Bread Wheat. Improvement and Production; Food and Agriculture Organization of the United Nation: Rome, Italy, 2002; pp. 19–37. [Google Scholar]
- Hirano, H.-Y.; Tanaka, W.; Toriba, T. Grass flower development. In Flower Development; Springer: Berlin, Germany, 2014; pp. 57–84. [Google Scholar]
- Langer, R.H.M.; Hanif, M. A study of floret development in wheat (Triticum aestivum L.). Ann. Bot. 1973, 37, 743–751. [Google Scholar] [CrossRef]
- Hoshikawa, K. The growing rice plant: An anatomical monograph. Nosan Gyoson Bunka 1989, 199–205. [Google Scholar]
- Tanaka, W.; Pautler, M.; Jackson, D.; Hirano, H.-Y. Grass meristems II: Inflorescence architecture, flower development and meristem fate. Plant Cell Physiol. 2013, 54, 313–324. [Google Scholar] [CrossRef]
- Kyozuka, J. Grass Inflorescence: Basic Structure and Diversity. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2014; pp. 191–219. [Google Scholar]
- Liu, C.; Thong, Z.; Yu, H. Coming into bloom: The specification of floral meristems. Development 2009, 136, 3379–3391. [Google Scholar] [CrossRef]
- Gramzow, L.; Theißen, G. Phylogenomics of MADS-box genes in plants—two opposing life styles in one gene family. Biology 2013, 2, 1150–1164. [Google Scholar] [CrossRef]
- Shore, P.; Sharrocks, A.D. The MADS-box family of transcription factors. Eur. J. Bioche. 1995, 229, 1–13. [Google Scholar] [CrossRef]
- Gramzow, L.; Ritz, M.S.; Theißen, G. On the origin of MADS-domain transcription factors. Trends Genet. 2010, 26, 149–153. [Google Scholar] [CrossRef]
- Masiero, S.; Colombo, L.; Grini, P.E.; Schnittger, A.; Kater, M.M. The emerging importance of type I MADS box transcription factors for plant reproduction. Plant Cell 2011, 23, 865–872. [Google Scholar] [CrossRef]
- Smaczniak, C.; Immink, R.G.; Muiño, J.M.; Blanvillain, R.; Busscher, M.; Busscher-Lange, J.; Dinh, Q.P.; Liu, S.; Westphal, A.H.; Boeren, S. Characterization of MADS-domain transcription factor complexes in Arabidopsis flower development. Proc. Nat. Acad. Sci. 2012, 109, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Theissen, G.; Becker, A.; Di Rosa, A.; Kanno, A.; Kim, J.T.; Münster, T.; Winter, K.-U.; Saedler, H. A short history of MADS-box genes in plants. Plant Mol. Biol. 2000, 42, 115–149. [Google Scholar] [CrossRef]
- Malcomber, S.T.; Preston, J.C.; Reinheimer, R.; Kossuth, J.; Kellogg, E.A. Developmental gene evolution and the origin of grass inflorescence diversity. Adv. Bot. Res. 2006, 44, 425–481. [Google Scholar]
- Pinyopich, A.; Ditta, G.S.; Savidge, B.; Liljegren, S.J.; Baumann, E.; Wisman, E.; Yanofsky, M.F. Assessing the redundancy of MADS-box genes during carpel and ovule development. Nature 2003, 424, 85–88. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Meng, X.; Selinger, D.A.; Deschamps, S.; Hermon, P.; Vansant, G.; Gupta, R.; Ananiev, E.V.; Muszynski, M.G. Involvement of the MADS-box gene ZMM4 in floral induction and inflorescence development in maize. Plant Physiol. 2008, 147, 2054–2069. [Google Scholar] [CrossRef]
- Jeon, J.-S.; Lee, S.; Jung, K.-H.; Yang, W.-S.; Yi, G.-H.; Oh, B.-G.; An, G. Production of transgenic rice plants showing reduced heading date and plant height by ectopic expression of rice MADS-box genes. Mol. Breed. 2000, 6, 581–592. [Google Scholar] [CrossRef]
- Kinjo, H.; Shitsukawa, N.; Takumi, S.; Murai, K. Diversification of three APETALA1/FRUITFULL-like genes in wheat. Mol. Genet. Genom. 2012, 287, 283–294. [Google Scholar] [CrossRef]
- Mandel, M.A.; Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar] [CrossRef]
- Münster, T.; Deleu, W.; Wingen, L.; Cacharrón, N.; Ouzunova, M.; Faigl, W. Maize MADS-box genes galore. Maydica 2002, 47, 287–301. [Google Scholar]
- Murai, K.; Miyamae, M.; Kato, H.; Takumi, S.; Ogihara, Y. WAP1, a wheat APETALA1 homolog, plays a central role in the phase transition from vegetative to reproductive growth. Plant Cell Physiol. 2003, 44, 1255–1265. [Google Scholar] [CrossRef]
- Preston, J.C.; Kellogg, E.A. Discrete developmental roles for temperate cereal grass VERNALIZATION1/FRUITFULL-like genes in flowering competency and the transition to flowering. Plant Physiol. 2008, 146, 265–276. [Google Scholar] [CrossRef]
- Wu, F.; Shi, X.; Lin, X.; Liu, Y.; Chong, K.; Theißen, G.; Meng, Z. The ABCs of flower development: Mutational analysis of AP1/FUL-like genes in rice provides evidence for a homeotic (A)-function in grasses. Plant J. 2017, 89, 310–324. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Nat. Acad. Sci. 2003, 100, 6263–6268. [Google Scholar] [CrossRef]
- Bai, X.; Huang, Y.; Mao, D.; Wen, M.; Zhang, L.; Xing, Y. Regulatory role of FZP in the determination of panicle branching and spikelet formation in rice. Sci. Rep. 2016, 6, 19022. [Google Scholar] [CrossRef]
- Chuck, G.; Meeley, R.; Hake, S. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development 2008, 135, 3013–3019. [Google Scholar] [CrossRef]
- Chuck, G.; Muszynski, M.; Kellogg, E.; Hake, S.; Schmidt, R.J. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science 2002, 298, 1238–1241. [Google Scholar] [CrossRef]
- Jofuku, K.D.; Den Boer, B.; Van Montagu, M.; Okamuro, J.K. Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 1994, 6, 1211–1225. [Google Scholar] [CrossRef]
- Komatsu, M.; Chujo, A.; Nagato, Y.; Shimamoto, K.; Kyozuka, J. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 2003, 130, 3841–3850. [Google Scholar] [CrossRef]
- Lee, D.Y.; An, G. Two AP2 family genes, supernumerary bract (SNB) and Osindeterminate spikelet 1 (OsIDS1), synergistically control inflorescence architecture and floral meristem establishment in rice. Plant J. 2012, 69, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.; Wang, N.; Sakuma, S.; Pourkheirandish, M.; Wu, J.; Matsumoto, T.; Koba, T.; Komatsuda, T. Structure, transcription and post-transcriptional regulation of the bread wheat orthologs of the barley cleistogamy gene Cly1. Theor. Appl. Genet. 2013, 126, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Li, Y.; Zhao, F.; Sang, X.; Shi, J.; Wang, N.; Guo, S.; Ling, Y.; Zhang, C.; Yang, Z. MULTI-FLORET SPIKELET1, which encodes an AP2/ERF protein, determines spikelet meristem fate and sterile lemma identity in rice. Plant Physiol. 2013, 162, 872–884. [Google Scholar] [CrossRef]
- Simons, K.J.; Fellers, J.P.; Trick, H.N.; Zhang, Z.; Tai, Y.-S.; Gill, B.S.; Faris, J.D. Molecular characterization of the major wheat domestication gene Q. Genetics 2006, 172, 547–555. [Google Scholar] [CrossRef]
- Ambrose, B.A.; Lerner, D.R.; Ciceri, P.; Padilla, C.M.; Yanofsky, M.F.; Schmidt, R.J. Molecular and genetic analyses of the silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol. Cell 2000, 5, 569–579. [Google Scholar] [CrossRef]
- Hama, E.; Takumi, S.; Ogihara, Y.; Murai, K. Pistillody is caused by alterations to the class-B MADS-box gene expression pattern in alloplasmic wheats. Planta 2004, 218, 712–720. [Google Scholar]
- Jack, T.; Brockman, L.L.; Meyerowitz, E.M. The homeotic gene APETALA3 of Arabidopsis thaliana encodes a MADS box and is expressed in petals and stamens. Cell 1992, 68, 683–697. [Google Scholar] [CrossRef]
- Murai, K.; Takumi, S.; Koga, H.; Ogihara, Y. Pistillody, homeotic transformation of stamens into pistil-like structures, caused by nuclear–cytoplasm interaction in wheat. Plant J. 2002, 29, 169–181. [Google Scholar] [CrossRef]
- Nagasawa, N.; Miyoshi, M.; Sano, Y.; Satoh, H.; Hirano, H.; Sakai, H.; Nagato, Y. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development 2003, 130, 705–718. [Google Scholar] [CrossRef]
- Goto, K.; Meyerowitz, E.M. Function and regulation of the Arabidopsis floral homeotic gene PISTILLATA. Genes Dev. 1994, 8, 1548–1560. [Google Scholar] [CrossRef]
- Murai, K.; Murai, R.; Takumi, S.; Ogihara, Y. Cloning and characterization of cDNAs corresponding to the wheat MADS box genes. In Proceedings of the 9th Int Wheat Genet Symp, Saskatoon, SK, Canada, 2–7 August 1998; pp. 89–94. [Google Scholar]
- Prasad, K.; Vijayraghavan, U. Double-stranded RNA interference of a rice PI/GLO paralog, OsMADS2, uncovers its second-whorl-specific function in floral organ patterning. Genetics 2003, 165, 2301–2305. [Google Scholar]
- Schilling, S.; Gramzow, L.; Lobbes, D.; Kirbis, A.; Weilandt, L.; Hoffmeier, A.; Junker, A.; Weigelt-Fischer, K.; Klukas, C.; Wu, F. Non-canonical structure, function and phylogeny of the Bsister MADS-box gene OsMADS30 of rice (Oryza sativa). Plant J. 2015, 84, 1059–1072. [Google Scholar] [CrossRef]
- Whipple, C.J.; Ciceri, P.; Padilla, C.M.; Ambrose, B.A.; Bandong, S.L.; Schmidt, R.J. Conservation of B-class floral homeotic gene function between maize and Arabidopsis. Development 2004, 131, 6083–6091. [Google Scholar] [CrossRef]
- Yao, S.-G.; Ohmori, S.; Kimizu, M.; Yoshida, H. Unequal genetic redundancy of rice PISTILLATA orthologs, OsMADS2 and OsMADS4, in lodicule and stamen development. Plant Cell Physiol. 2008, 49, 853–857. [Google Scholar] [CrossRef]
- Becker, A.; Kaufmann, K.; Freialdenhoven, A.; Vincent, C.; Li, M.-A.; Saedler, H.; Theissen, G. A novel MADS-box gene subfamily with a sister-group relationship to class B floral homeotic genes. Mol. Genet. Genom. 2002, 266, 942–950. [Google Scholar]
- Nesi, N.; Debeaujon, I.; Jond, C.; Stewart, A.J.; Jenkins, G.I.; Caboche, M.; Lepiniec, L. The TRANSPARENT TESTA16 locus encodes the ARABIDOPSIS BSISTER MADS domain protein and is required for proper development and pigmentation of the seed coat. Plant Cell 2002, 14, 2463–2479. [Google Scholar] [CrossRef]
- Prasad, K.; Zhang, X.; Tobón, E.; Ambrose, B.A. The Arabidopsis B-sister MADS-box protein, GORDITA, represses fruit growth and contributes to integument development. Plant J. 2010, 62, 203–214. [Google Scholar] [CrossRef]
- Yamada, K.; Saraike, T.; Shitsukawa, N.; Hirabayashi, C.; Takumi, S.; Murai, K. Class D and Bsister MADS-box genes are associated with ectopic ovule formation in the pistil-like stamens of alloplasmic wheat (Triticum aestivum L.). Plant Mol. Biol. 2009, 71, 1–14. [Google Scholar] [CrossRef]
- Yang, X.; Wu, F.; Lin, X.; Du, X.; Chong, K.; Gramzow, L.; Schilling, S.; Becker, A.; Theißen, G.; Meng, Z. Live and let die-The B sister MADS-box gene OsMADS29 controls the degeneration of cells in maternal tissues during seed development of rice (Oryza sativa). PLoS ONE 2012, 7, e51435. [Google Scholar] [CrossRef]
- Dreni, L.; Pilatone, A.; Yun, D.; Erreni, S.; Pajoro, A.; Caporali, E.; Zhang, D.; Kater, M.M. Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy. Plant Cell 2011, 23, 2850–2863. [Google Scholar] [CrossRef]
- Gómez-Mena, C.; de Folter, S.; Costa, M.M.R.; Angenent, G.C.; Sablowski, R. Transcriptional program controlled by the floral homeotic gene AGAMOUS during early organogenesis. Development 2005, 132, 429–438. [Google Scholar] [CrossRef]
- Hirabayashi, C.; Murai, K. Class C MADS-box gene AGAMOUS was duplicated in the wheat genome. Wheat Inf. Serv. 2009, 107, 13–16. [Google Scholar]
- Meguro, A.; Takumi, S.; Ogihara, Y.; Murai, K. WAG, a wheat AGAMOUS homolog, is associated with development of pistil-like stamens in alloplasmic wheats. Sex. Plant Reprod. 2003, 15, 221–230. [Google Scholar]
- Mena, M.; Ambrose, B.A.; Meeley, R.B.; Briggs, S.P. Diversification of C-function activity in maize flower development. Science 1996, 274, 1537. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Veit, B.; Mandel, M.A.; Mena, M.; Hake, S.; Yanofsky, M.F. Identification and molecular characterization of ZAG1, the maize homolog of the Arabidopsis floral homeotic gene AGAMOUS. Plant Cell 1993, 5, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Lee, D.Y.; Miyao, A.; Hirochika, H.; An, G.; Hirano, H.-Y. Functional diversification of the two C-class MADS box genes OSMADS3 and OSMADS58 in Oryza sativa. Plant Cell 2006, 18, 15–28. [Google Scholar] [CrossRef]
- Dreni, L.; Jacchia, S.; Fornara, F.; Fornari, M.; Ouwerkerk, P.B.; An, G.; Colombo, L.; Kater, M.M. The D-lineage MADS-box gene OsMADS13 controls ovule identity in rice. Plant J. 2007, 52, 690–699. [Google Scholar] [CrossRef]
- Favaro, R.; Immink, R.; Ferioli, V.; Bernasconi, B.; Byzova, M.; Angenent, G.; Kater, M.; Colombo, L. Ovule-specific MADS-box proteins have conserved protein-protein interactions in monocot and dicot plants. Mol. Genet. Genom. 2002, 268, 152–159. [Google Scholar] [CrossRef]
- Favaro, R.; Pinyopich, A.; Battaglia, R.; Kooiker, M.; Borghi, L.; Ditta, G.; Yanofsky, M.F.; Kater, M.M.; Colombo, L. MADS-box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 2003, 15, 2603–2611. [Google Scholar] [CrossRef]
- Lopez-Dee, Z.P.; Wittich, P.; Pe, M.E.; Rigola, D.; Del Buono, I.; Gorla, M.S.; Kater, M.M.; Colombo, L. OsMADS13, a novel rice MADS-box gene expressed during ovule development. Dev. Genet. 1999, 25, 237–244. [Google Scholar] [CrossRef]
- Yamaki, S.; Nagato, Y.; Kurata, N.; Nonomura, K.-I. Ovule is a lateral organ finally differentiated from the terminating floral meristem in rice. Dev. Biol. 2011, 351, 208–216. [Google Scholar] [CrossRef]
- Cui, R.; Han, J.; Zhao, S.; Su, K.; Wu, F.; Du, X.; Xu, Q.; Chong, K.; Theißen, G.; Meng, Z. Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J. 2010, 61, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Lid, S.E.; Meeley, R.B.; Min, Z.; Nichols, S.; Olsen, O.-A. Knock-out mutants of two members of the AGL2 subfamily of MADS-box genes expressed during maize kernel development. Plant Sci. 2004, 167, 575–582. [Google Scholar] [CrossRef]
- Li, Q.; Liu, B. Genetic regulation of maize flower development and sex determination. Planta 2017, 245, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Shitsukawa, N.; Tahira, C.; Kassai, K.-i.; Hirabayashi, C.; Shimizu, T.; Takumi, S.; Mochida, K.; Kawaura, K.; Ogihara, Y.; Murai, K. Genetic and epigenetic alteration among three homoeologous genes of a class E MADS box gene in hexaploid wheat. Plant Cell 2007, 19, 1723–1737. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Cheng, Z.J.; Zhang, X.S. Overexpression of TaMADS1, a SEPALLATA-like gene in wheat, causes early flowering and the abnormal development of floral organs in Arabidopsis. Planta 2006, 223, 698–707. [Google Scholar] [CrossRef]
- Cacharrón, J.; Saedler, H.; Theißen, G. Expression of MADS box genes ZMM8 and ZMM14 during inflorescence development of Zea mays discriminates between the upper and the lower floret of each spikelet. Dev. Genes Evo. 1999, 209, 411–420. [Google Scholar] [CrossRef]
- Christensen, A.R.; Malcomber, S.T. Duplication and diversification of the LEAFY HULL STERILE1 and Oryza sativa MADS5 SEPALLATA lineages in graminoid Poales. EvoDevo 2012, 3, 4. [Google Scholar] [CrossRef]
- Gao, X.; Liang, W.; Yin, C.; Ji, S.; Wang, H.; Su, X.; Guo, C.; Kong, H.; Xue, H.; Zhang, D. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 2010, 153, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yasuno, N.; Sato, Y.; Yoda, M.; Yamazaki, R.; Kimizu, M.; Yoshida, H.; Nagamura, Y.; Kyozuka, J. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-like MADS box genes and PAP2, a SEPALLATA MADS box gene. Plant Cell 2012, 24, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, W.; Hu, Y.; Zhu, L.; Yin, C.; Xu, J.; Dreni, L.; Kater, M.M.; Zhang, D. Rice MADS6 interacts with the floral homeotic genes SUPERWOMAN1, MADS3, MADS58, MADS13, and DROOPING LEAF in specifying floral organ identities and meristem fate. Plant Cell 2011, 23, 2536–2552. [Google Scholar] [CrossRef]
- Li, H.; Liang, W.; Jia, R.; Yin, C.; Zong, J.; Kong, H.; Zhang, D. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res. 2010, 20, 299–313. [Google Scholar] [CrossRef]
- Ohmori, S.; Kimizu, M.; Sugita, M.; Miyao, A.; Hirochika, H.; Uchida, E.; Nagato, Y.; Yoshida, H. MOSAIC FLORAL ORGANS1, an AGL6-like MADS box gene, regulates floral organ identity and meristem fate in rice. Plant Cell 2009, 21, 3008–3025. [Google Scholar] [CrossRef]
- Reinheimer, R.; Kellogg, E.A. Evolution of AGL6-like MADS box genes in grasses (Poaceae): Ovule expression is ancient and palea expression is new. Plant Cell 2009, 21, 2591–2605. [Google Scholar] [CrossRef]
- Zhao, T.; Ni, Z.; Dai, Y.; Yao, Y.; Nie, X.; Sun, Q. Characterization and expression of 42 MADS-box genes in wheat (Triticum aestivum L.). Mol. Genet. Genom. 2006, 276, 334–350. [Google Scholar] [CrossRef]
- Bowman, J.L.; Smyth, D.R. CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 1999, 126, 2387–2396. [Google Scholar]
- Ishikawa, M.; Ohmori, Y.; Tanaka, W.; Hirabayashi, C.; Murai, K.; Ogihara, Y.; Yamaguchi, T.; Hirano, H.-Y. The spatial expression patterns of DROOPING LEAF orthologs suggest a conserved function in grasses. Genes Genet. Sys. 2009, 84, 137–146. [Google Scholar] [CrossRef]
- Strable, J. Functional and Genomic Analyses of the Maize Yabby Transcription Factors Drooping Leaf1 and Drooping Leaf2. Ph.D. Thesis, Low State University, Uganda, Africa, 2015. [Google Scholar]
- Ikeda, K.; Ito, M.; Nagasawa, N.; Kyozuka, J.; Nagato, Y. Rice ABERRANT PANICLE ORGANIZATION 1, encoding an F-box protein, regulates meristem fate. Plant J. 2007, 51, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Nagasawa, N.; Nagato, Y. ABERRANT PANICLE ORGANIZATION 1 temporally regulates meristem identity in rice. Dev. Biol. 2005, 282, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Wolfe, D.S.; Nilsson, O.; Weigel, D. A LEAFY co-regulator encoded by unusual floral organs. Curr. Biol. 1997, 7, 95–104. [Google Scholar] [CrossRef]
- Liu, M.; Li, H.; Su, Y.; Li, W.; Shi, C. G1/ELE Functions in the Development of Rice Lemmas in Addition to Determining Identities of Empty Glumes. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.; Li, Y.; Luo, Z.; Ren, D.; Fang, L.; Wang, N.; Zhao, F.; Ling, Y.; Yang, Z.; Liu, Y. CHIMERIC FLORAL ORGANS1, encoding a monocot-specific MADS box protein, regulates floral organ identity in rice. Plant Physiol. 2012, 160, 788–807. [Google Scholar] [CrossRef]
- Yang, D.; Ye, X.; Zheng, X.; Cheng, C.; Ye, N.; Lu, L.; Huang, F.; Li, Q. Identification and fine mapping of lemma-distortion1, a single recessive gene playing an essential role in the development of lemma in rice. J. Agri. Sci. 2016, 154, 989–1001. [Google Scholar] [CrossRef]
- Müller, B.M.; Saedler, H.; Zachgo, S. The MADS-box gene DEFH28 from Antirrhinum is involved in the regulation of floral meristem identity and fruit development. Plant J. 2001, 28, 169–179. [Google Scholar] [CrossRef]
- Litt, A.; Irish, V.F. Duplication and diversification in the APETALA1/FRUITFULL floral homeotic gene lineage: Implications for the evolution of floral development. Genetics 2003, 165, 821–833. [Google Scholar]
- Chen, A.; Dubcovsky, J. Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLOS Genet. 2012, 8, e1003134. [Google Scholar] [CrossRef]
- Wang, K.; Tang, D.; Hong, L.; Xu, W.; Huang, J.; Li, M.; Gu, M.; Xue, Y.; Cheng, Z. DEP and AFO regulate reproductive habit in rice. PLOS Genet. 2010, 6, e1000818. [Google Scholar] [CrossRef]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Varotto, S.; Ciaffi, M. Molecular and phylogenetic analysis of MADS-box genes of MIKC type and chromosome location of SEP-like genes in wheat (Triticum aestivum L.). Mol. Genet. Genom. 2007, 278, 689–708. [Google Scholar] [CrossRef]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-box gene family in rice: Genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef]
- Pařenicová, L.; de Folter, S.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J.; Cook, H.E.; Ingram, R.M.; Kater, M.M.; Davies, B. Molecular and phylogenetic analyses of the complete MADS-box transcription factor family in Arabidopsis new openings to the MADS world. Plant Cell 2003, 15, 1538–1551. [Google Scholar] [CrossRef]
- Trevaskis, B.; Bagnall, D.J.; Ellis, M.H.; Peacock, W.J.; Dennis, E.S. MADS box genes control vernalization-induced flowering in cereals. Proc. Nat. Acad. Sci. 2003, 100, 13099–13104. [Google Scholar] [CrossRef]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct links between the vernalization response and other key traits of cereal crops. Nature Commun. 2015, 6. [Google Scholar] [CrossRef]
- Greenup, A.G.; Sasani, S.; Oliver, S.N.; Talbot, M.J.; Dennis, E.S.; Hemming, M.N.; Trevaskis, B. ODDSOC2 is a MADS box floral repressor that is down-regulated by vernalization in temperate cereals. Plant Physiol. 2010, 153, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Sys. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evo. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evo. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lee, J.; Moon, S.; Park, S.Y.; An, G. The rice heterochronic gene SUPERNUMERARY BRACT regulates the transition from spikelet meristem to floral meristem. Plant J. 2007, 49, 64–78. [Google Scholar] [CrossRef]
- Nair, S.K.; Wang, N.; Turuspekov, Y.; Pourkheirandish, M.; Sinsuwongwat, S.; Chen, G.; Sameri, M.; Tagiri, A.; Honda, I.; Watanabe, Y. Cleistogamous flowering in barley arises from the suppression of microRNA-guided HvAP2 mRNA cleavage. Proc. Nat. Acad. Sci. 2010, 107, 490–495. [Google Scholar] [CrossRef]
- Jung, J.-H.; Seo, Y.-H.; Seo, P.J.; Reyes, J.L.; Yun, J.; Chua, N.-H.; Park, C.-M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef]
- Yant, L.; Mathieu, J.; Dinh, T.T.; Ott, F.; Lanz, C.; Wollmann, H.; Chen, X.; Schmid, M. Orchestration of the floral transition and floral development in Arabidopsis by the bifunctional transcription factor APETALA2. Plant Cell 2010, 22, 2156–2170. [Google Scholar] [CrossRef]
- Wollmann, H.; Mica, E.; Todesco, M.; Long, J.A.; Weigel, D. On reconciling the interactions between APETALA2, miR172 and AGAMOUS with the ABC model of flower development. Development 2010, 137, 3633–3642. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Upadhyaya, N.M.; Gubler, F.; Helliwell, C.A. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2009, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.-H.; Helliwell, C.A. Regulation of flowering time and floral patterning by miR172. J. Exp. Bot. 2011, erq295. [Google Scholar] [CrossRef]
- Chung, Y.-Y.; Kim, S.-R.; Kang, H.-G.; Noh, Y.-S.; Park, M.C.; Finkel, D.; An, G. Characterization of two rice MADS box genes homologous to GLOBOSA. Plant Sci. 1995, 109, 45–56. [Google Scholar] [CrossRef]
- Bartlett, M.E.; Williams, S.K.; Taylor, Z.; DeBlasio, S.; Goldshmidt, A.; Hall, D.H.; Schmidt, R.J.; Jackson, D.P.; Whipple, C.J. The Maize PI/GLO Ortholog Zmm16/sterile tassel silky ear1 interacts with the zygomorphy and sex determination pathways in flower development. Plant Cell 2015, 27, 3081–3098. [Google Scholar] [CrossRef] [PubMed]
- De Folter, S.; Shchennikova, A.V.; Franken, J.; Busscher, M.; Baskar, R.; Grossniklaus, U.; Angenent, G.C.; Immink, R.G. A Bsister MADS-box gene involved in ovule and seed development in petunia and Arabidopsis. Plant J. 2006, 47, 934–946. [Google Scholar] [CrossRef]
- Mizzotti, C.; Mendes, M.A.; Caporali, E.; Schnittger, A.; Kater, M.M.; Battaglia, R.; Colombo, L. The MADS box genes SEEDSTICK and ARABIDOPSIS Bsister play a maternal role in fertilization and seed development. Plant J. 2012, 70, 409–420. [Google Scholar] [CrossRef]
- Kramer, E.M.; Jaramillo, M.A.; Di Stilio, V.S. Patterns of gene duplication and functional evolution during the diversification of the AGAMOUS subfamily of MADS box genes in angiosperms. Genetics 2004, 166, 1011–1023. [Google Scholar] [CrossRef]
- Zahn, L.M.; Leebens-Mack, J.H.; Arrington, J.M.; Hu, Y.; Landherr, L.L.; Depamphilis, C.W.; Becker, A.; Theissen, G.; Ma, H. Conservation and divergence in the AGAMOUS subfamily of MADS-box genes: Evidence of independent sub-and neofunctionalization events. Evo. Dev. 2006, 8, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Yanofsky, M.F.; Ma, H.; Bowman, J.L.; Drews, G.N.; Feldmann, K.A.; Meyerowitz, E.M. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature 1990, 346, 35. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Ditta, G.S.; Eshed, Y.; Savidge, B. SHATTERPROOF MADS-box genes control seed dispersal in Arabidopsis. Nature 2000, 404, 766. [Google Scholar] [CrossRef]
- Yun, D.; Liang, W.; Dreni, L.; Yin, C.; Zhou, Z.; Kater, M.M.; Zhang, D. OsMADS16 genetically interacts with OsMADS3 and OsMADS58 in specifying floral patterning in rice. Mol. Plant 2013, 6, 743–756. [Google Scholar] [CrossRef]
- Angenent, G.C.; Franken, J.; Busscher, M.; van Dijken, A.; van Went, J.L.; Dons, H.; van Tunen, A.J. A novel class of MADS box genes is involved in ovule development in petunia. Plant Cell 1995, 7, 1569–1582. [Google Scholar] [CrossRef]
- Singh, L.N.; Hannenhalli, S. Functional diversification of paralogous transcription factors via divergence in DNA binding site motif and in expression. PLoS ONE 2008, 3, e2345. [Google Scholar] [CrossRef]
- Li, H.; Liang, W.; Yin, C.; Zhu, L.; Zhang, D. Genetic interaction of OsMADS3, DROOPING LEAF, and OsMADS13 in specifying rice floral organ identities and meristem determinacy. Plant Physiol. 2011, 156, 263–274. [Google Scholar] [CrossRef]
- Theissen, G.; Strater, T.; Fischer, A.; Saedler, H. Structural characterization, chromosomal localization and phylogenetic evaluation of two pairs of AGAMOUS-like MADS-box genes from maize. Gene 1995, 156, 155–166. [Google Scholar]
- Mizumoto, K.; Hatano, H.; Hirabayashi, C.; Murai, K.; Takumi, S. Altered expression of wheat AINTEGUMENTA homolog, WANT-1, in pistil and pistil-like transformed stamen of an alloplasmic line with Aegilops crassa cytoplasm. Dev. Genes Evo. 2009, 219, 175–187. [Google Scholar] [CrossRef]
- Theissen, G.; Saedler, H. Plant biology: Floral quartets. Nature 2001, 409, 469–472. [Google Scholar] [CrossRef]
- Malcomber, S.T.; Kellogg, E.A. SEPALLATA gene diversification: Brave new whorls. Trends Plant Sci. 2005, 10, 427–435. [Google Scholar] [CrossRef]
- Prasad, K.; Parameswaran, S.; Vijayraghavan, U. OsMADS1, a rice MADS-box factor, controls differentiation of specific cell types in the lemma and palea and is an early-acting regulator of inner floral organs. Plant J. 2005, 43, 915–928. [Google Scholar] [CrossRef]
- Hu, Y.; Liang, W.; Yin, C.; Yang, X.; Ping, B.; Li, A.; Jia, R.; Chen, M.; Luo, Z.; Cai, Q. Interactions of OsMADS1 with floral homeotic genes in rice flower development. Mol. Plant 2015, 8, 1366–1384. [Google Scholar] [CrossRef]
- Ren, D.; Rao, Y.; Leng, Y.; Li, Z.; Xu, Q.; Wu, L.; Qiu, Z.; Xue, D.; Zeng, D.; Hu, J. Regulatory Role of OsMADS34 in the Determination of Glumes Fate, Grain Yield, and Quality in Rice. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Duan, Y.; Xing, Z.; Diao, Z.; Xu, W.; Li, S.; Du, X.; Wu, G.; Wang, C.; Lan, T.; Meng, Z. Characterization of Osmads6-5, a null allele, reveals that OsMADS6 is a critical regulator for early flower development in rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Malcomber, S.T.; Kellogg, E.A. Heterogeneous expression patterns and separate roles of the SEPALLATA gene LEAFY HULL STERILE1 in grasses. Plant Cell 2004, 16, 1692–1706. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, W.; Toriba, T.; Hirano, H.-Y. Flower development in rice. The Molecular Genetics of Floral Transition and Flower Development, (ed. Fornara, F.) 2014, 221–262.
- Ikeda-Kawakatsu, K.; Maekawa, M.; Izawa, T.; Itoh, J.I.; Nagato, Y. ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1. Plant J. 2012, 69, 168–180. [Google Scholar] [CrossRef]
- Rao, N.N.; Prasad, K.; Kumar, P.R.; Vijayraghavan, U. Distinct regulatory role for RFL, the rice LFY homolog, in determining flowering time and plant architecture. Proc. Nat. Acad. Sci. 2008, 105, 3646–3651. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L.G.; Dayton, C.B.; Matthew, R.T. Exploring the role of the ovule in cereal grain development and reproductive stress tolerance. Ann. Plant Rev. Online 2018, 1, 1–35. [Google Scholar]
- Rijpkema, A.S.; Zethof, J.; Gerats, T.; Vandenbussche, M. The petunia AGL6 gene has a SEPALLATA-like function in floral patterning. Plant J. 2009, 60, 1–9. [Google Scholar] [CrossRef]
- Raza, Q.; Ali, Z.; Karim, I.; Ajmal, M.; Khan, M.U. Genetic analysis of triple pistil wheat derived two F2 populations to enhance genetic yield potential. Res. Plant Biol. 2019, 9, 1–8. [Google Scholar]
- Albert, V.A.; Soltis, D.E.; Carlson, J.E.; Farmerie, W.G.; Wall, P.K.; Ilut, D.C.; Solow, T.M.; Mueller, L.A.; Landherr, L.L.; Hu, Y. Floral gene resources from basal angiosperms for comparative genomics research. BMC Plant Biol. 2005, 5, 5. [Google Scholar] [CrossRef]
- Matasci, N.; Hung, L.-H.; Yan, Z.; Carpenter, E.J.; Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Ayyampalayam, S.; Barker, M. Data access for the 1,000 Plants (1KP) project. GigaScience 2014, 3, 17. [Google Scholar] [CrossRef]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotec. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef]
- Dinesh-Kumar, S.; Anandalakshmi, R.; Marathe, R.; Schiff, M.; Liu, Y. Virus-induced gene silencing. Plant Funct. Genom. 2003, 287–293. [Google Scholar]
- Gould, B.; Kramer, E.M. Virus-induced gene silencing as a tool for functional analyses in the emerging model plant Aquilegia (columbine, Ranunculaceae). Plant Methods 2007, 3, 6. [Google Scholar] [CrossRef]
- Hidalgo, O.; Bartholmes, C.; Gleissberg, S. Virus-induced gene silencing (VIGS) in Cysticapnos vesicaria, a zygomorphic-flowered Papaveraceae (Ranunculales, basal eudicots). Ann. Bot. 2012, 109, 911–920. [Google Scholar] [CrossRef]
- Sharma, B.; Guo, C.; Kong, H.; Kramer, E.M. Petal-specific subfunctionalization of an APETALA3 paralog in the Ranunculales and its implications for petal evolution. New Phytol. 2011, 191, 870–883. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Arabidopsis Gene | Subfamily | Class | Putative Role | Rice | Maize | Wheat | Brachypodium | References |
|---|---|---|---|---|---|---|---|---|
| AP1 | AP1 | A | Promote floral meristems, sepals, petals or lemma/palea identities | OsMADS14 | ZMM4, 15 | WFUL1, TaAGL25 | BrMADS33 | [31,58,59,60,61,62,63,64,65,66] |
| OsMADS15 | ZAP1 | WFUL2 | BrMADS10 | |||||
| OsMADS18 | - | WFUL3, TaAGL10 | BrMADS3 | |||||
| OsMADS20 | - | - | BrMADS31 | |||||
| AP2 | AP2 | A | Spikelet/floral meristem identity and lodicule identity | IDS1 | ids1 | TaAP2 | - | [24,25,67,68,69,70,71,72,73,74,75] |
| SNB | sid1 | TaQ | - | |||||
| MFS1 | - | - | - | |||||
| FZP | BD1 | WFZP | MOS1 | |||||
| AP3 | AP3 | B | Lodicule and stamen identity | OsMADS16/SPW1 | Silky1 | WAP3 | BrMADS5 | [31,76,77,78,79,80] |
| PI | PI | B | OsMADS2 | ZMM16 | WPI2 | BrMADS20 | [31,77,81,82,83,84,85,86] | |
| OsMADS4 | ZMM18, 29 | WPI1 | BrMADS16 | |||||
| ABS/TT16, GOA | BSISTER | - | Integuments and seed development | OsMADS29, 30 | ZMM17 | WBsis, TaAGL35 | BrMADS17, 23, 38 | [31,84,87,88,89,90,91] |
| AG, SHP1, SHP2 | AG | C | Stamen and carpel identity | OsMADS3 | ZMM2 | WAG-2 | BrMADS14 | [31,57,92,93,94,95,96,97,98] |
| OsMADS58 | ZAG1 | WAG-1 | BrMADS18 | |||||
| STK | AG | D | Ovule identity | OsMADS13 | ZMM1, ZAG2 | WSTK, TaAGL9, 31 | BrMADS2 | [30,31,57,62,90,99,100,101,102,103] |
| OsMADS21 | - | - | BrMADS4 | |||||
| SEP (1-4) | SEP | E | FM determinacy and floral organ identity | OsMADS7/45 | ZMM6 | WSEP, TaAGL16, 28,30 | BrMADS26 | [31,34,104,105,106,107,108] |
| OsMADS8/24 | ZMM7/27 | TaMADS1 | BrMADS32 | |||||
| - | LOFSEP | E | OsMADS1/LHS1 | ZMM8/14 | WLHS1, TaAGL24 | BrMADS11 | [31,104,107,109,110,111,112] | |
| OsMADS5 | - | TaAGL3, 5, 8, 34, 40 | BrMADS7 | |||||
| OsMADS34/PAP2 | ZMM24, 31 | TaAGL27 | BrMADS1 | |||||
| - | AGL6 | E | OsMADS6/MFO1 | ZAG3, 5 | TaAGL6, 37 | BrMADS28 | [31,82,105,113,114,115,116,117] | |
| OsMADS17 | - | - | - | |||||
| CRC | YABBY-like | - | Carpel identity | DL | Drl1, 2 | TaDL | - | [77,80,118,119,120] |
| UFO | F-box | - | Positive regulation of B & C class MADS | APO1 | - | - | - | [121,122,123] |
| - | DoF | - | Lemma and Palea identity? | CFO1, G1/ELE, ld-1 | - | - | - | [124,125,126] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, Z.; Raza, Q.; Atif, R.M.; Aslam, U.; Ajmal, M.; Chung, G. Genetic and Molecular Control of Floral Organ Identity in Cereals. Int. J. Mol. Sci. 2019, 20, 2743. https://doi.org/10.3390/ijms20112743
Ali Z, Raza Q, Atif RM, Aslam U, Ajmal M, Chung G. Genetic and Molecular Control of Floral Organ Identity in Cereals. International Journal of Molecular Sciences. 2019; 20(11):2743. https://doi.org/10.3390/ijms20112743
Chicago/Turabian StyleAli, Zulfiqar, Qasim Raza, Rana Muhammad Atif, Usman Aslam, Muhammad Ajmal, and Gyuhwa Chung. 2019. "Genetic and Molecular Control of Floral Organ Identity in Cereals" International Journal of Molecular Sciences 20, no. 11: 2743. https://doi.org/10.3390/ijms20112743
APA StyleAli, Z., Raza, Q., Atif, R. M., Aslam, U., Ajmal, M., & Chung, G. (2019). Genetic and Molecular Control of Floral Organ Identity in Cereals. International Journal of Molecular Sciences, 20(11), 2743. https://doi.org/10.3390/ijms20112743