In Vitro Study of the Anticancer Effects of Biotechnological Extracts of the Endangered Plant Species Satureja Khuzistanica

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
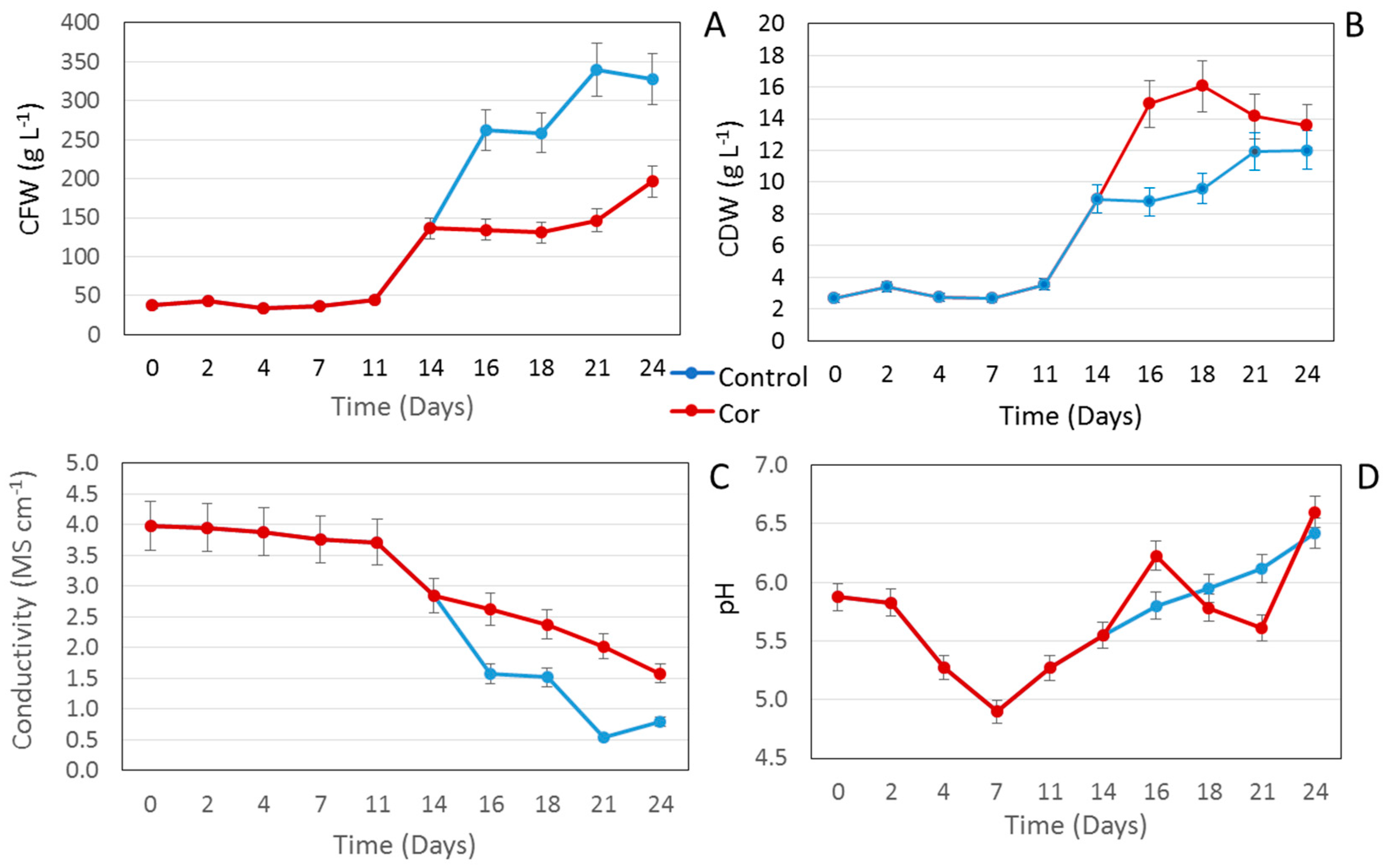
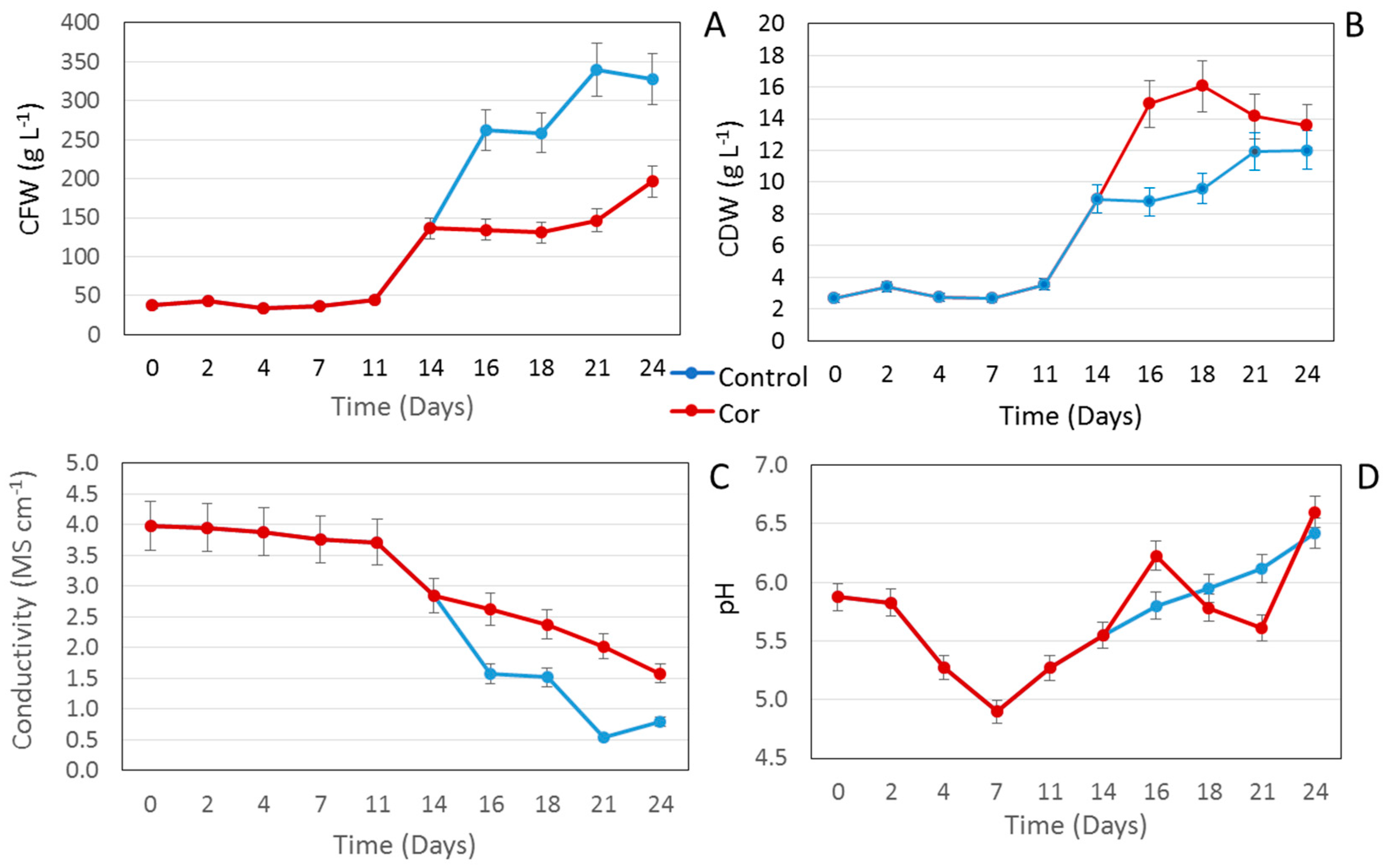
2.1. Effect of Coronatine on a Small Scale
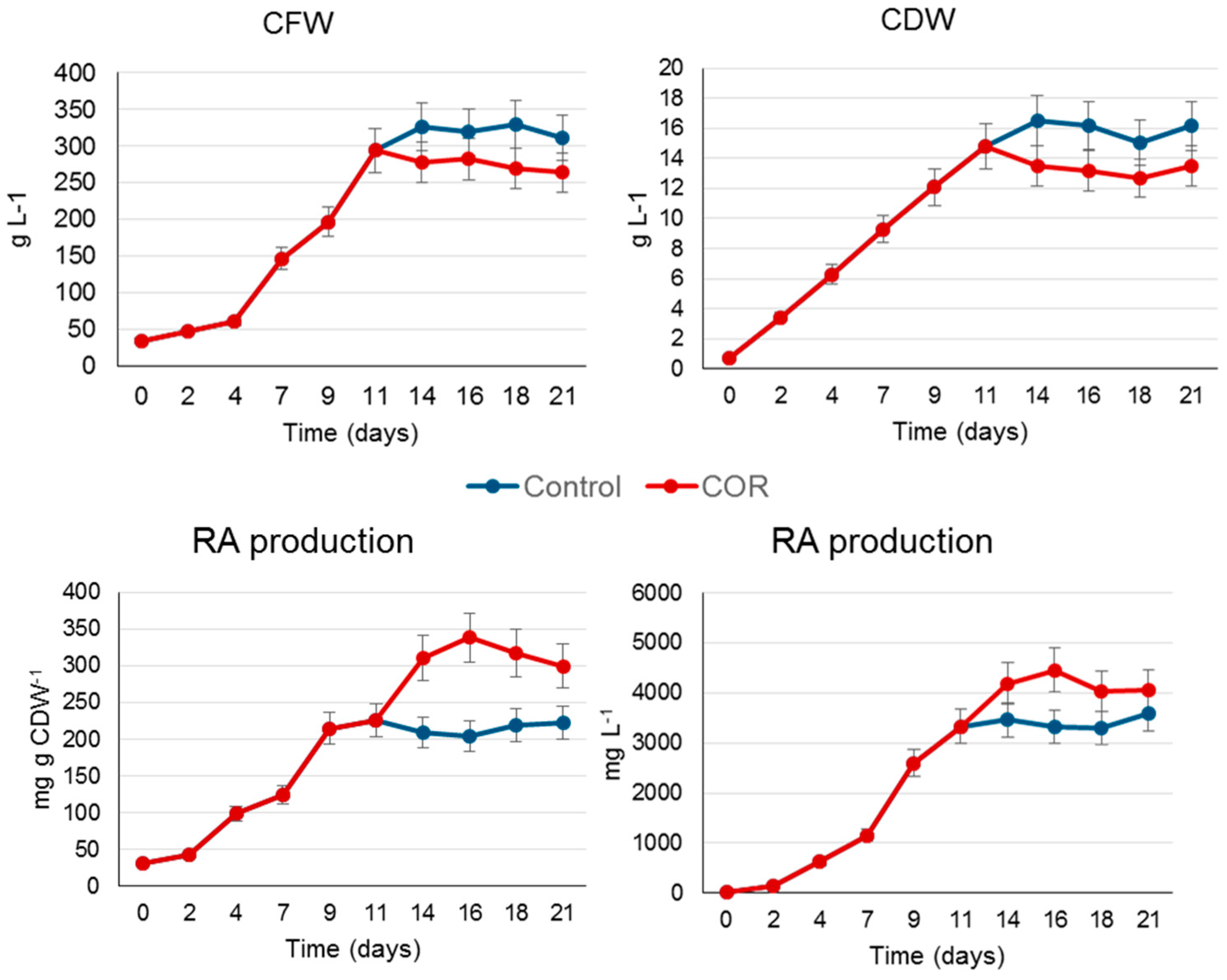
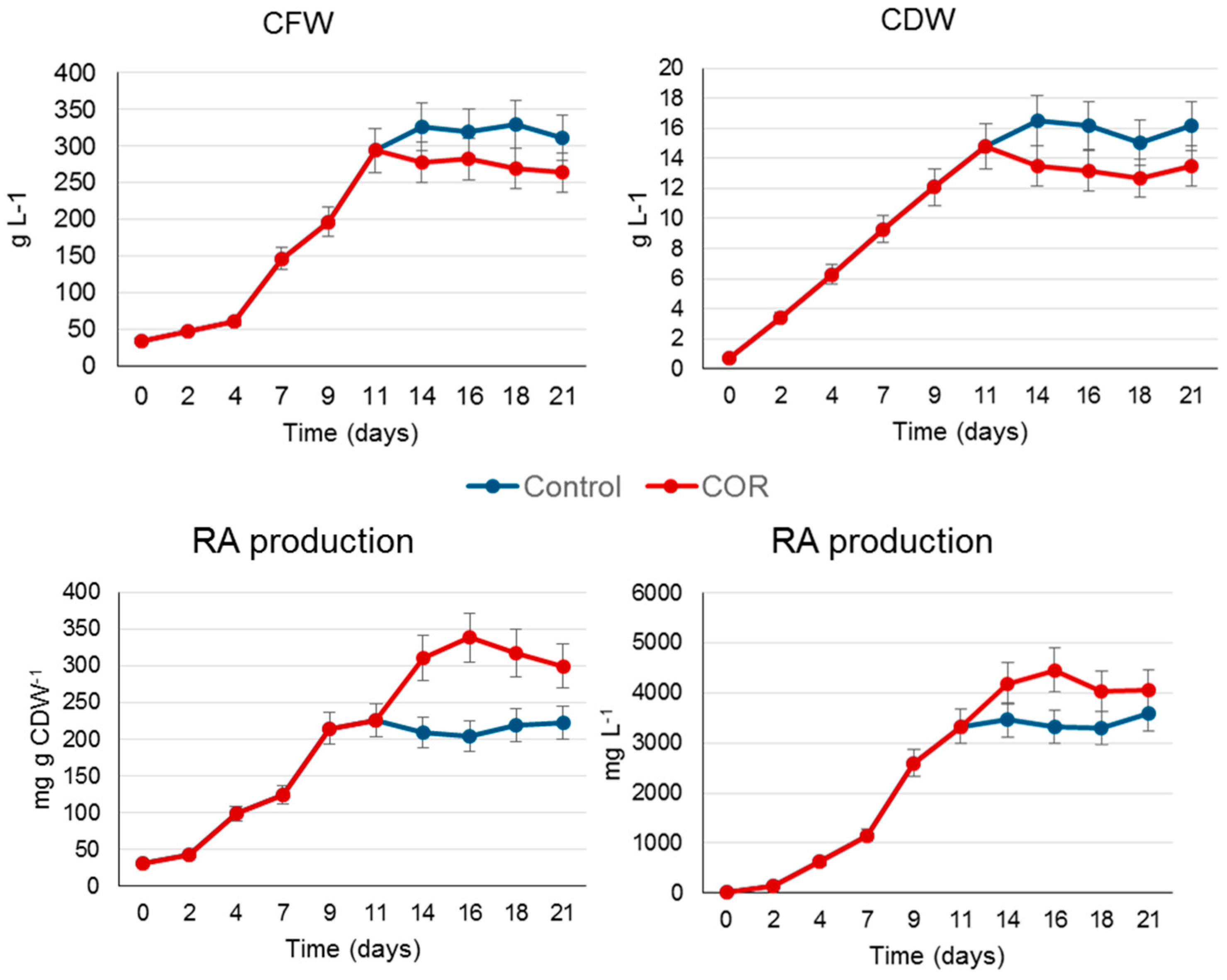
2.2. Scaling Up the Process to a Benchtop Bioreactor
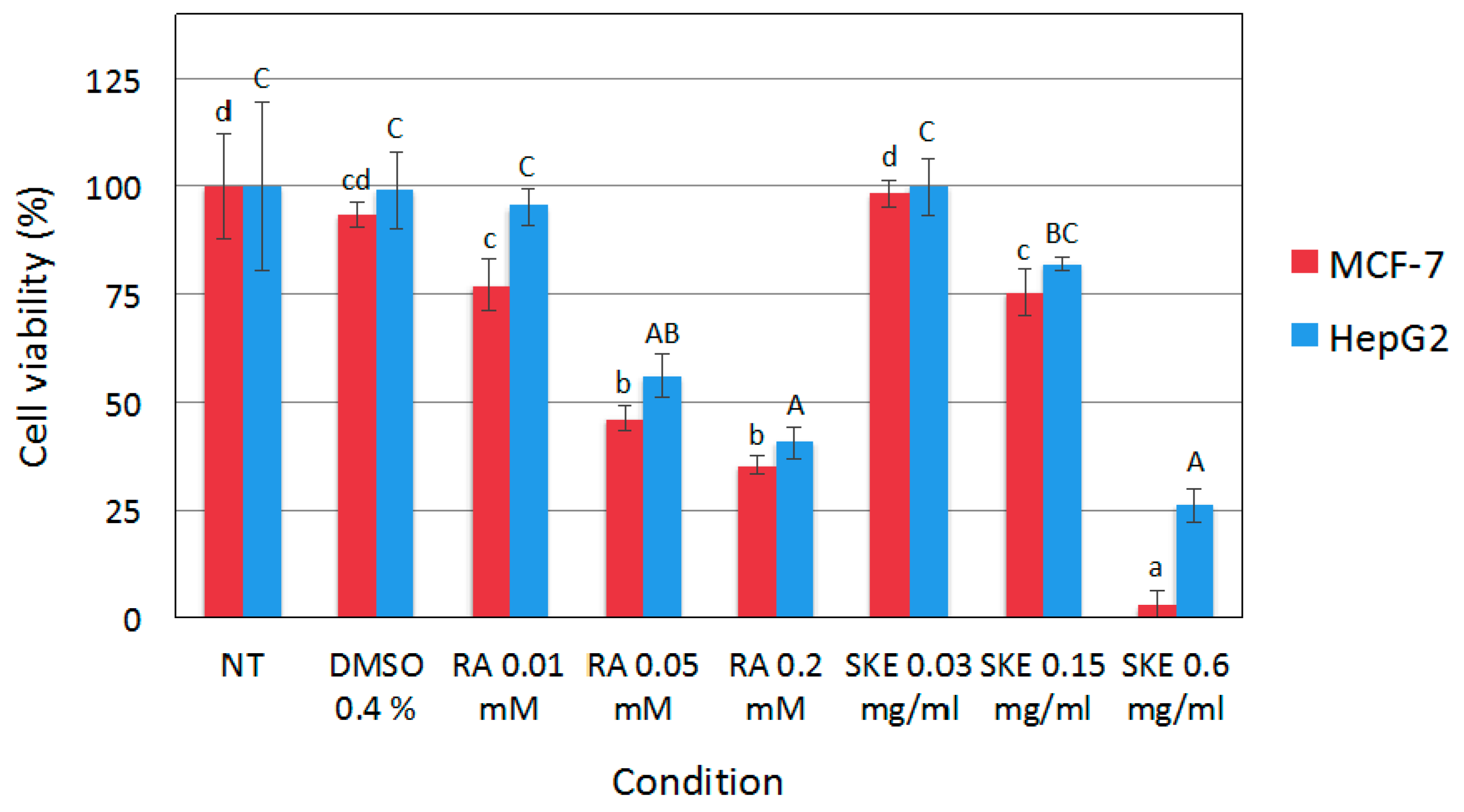
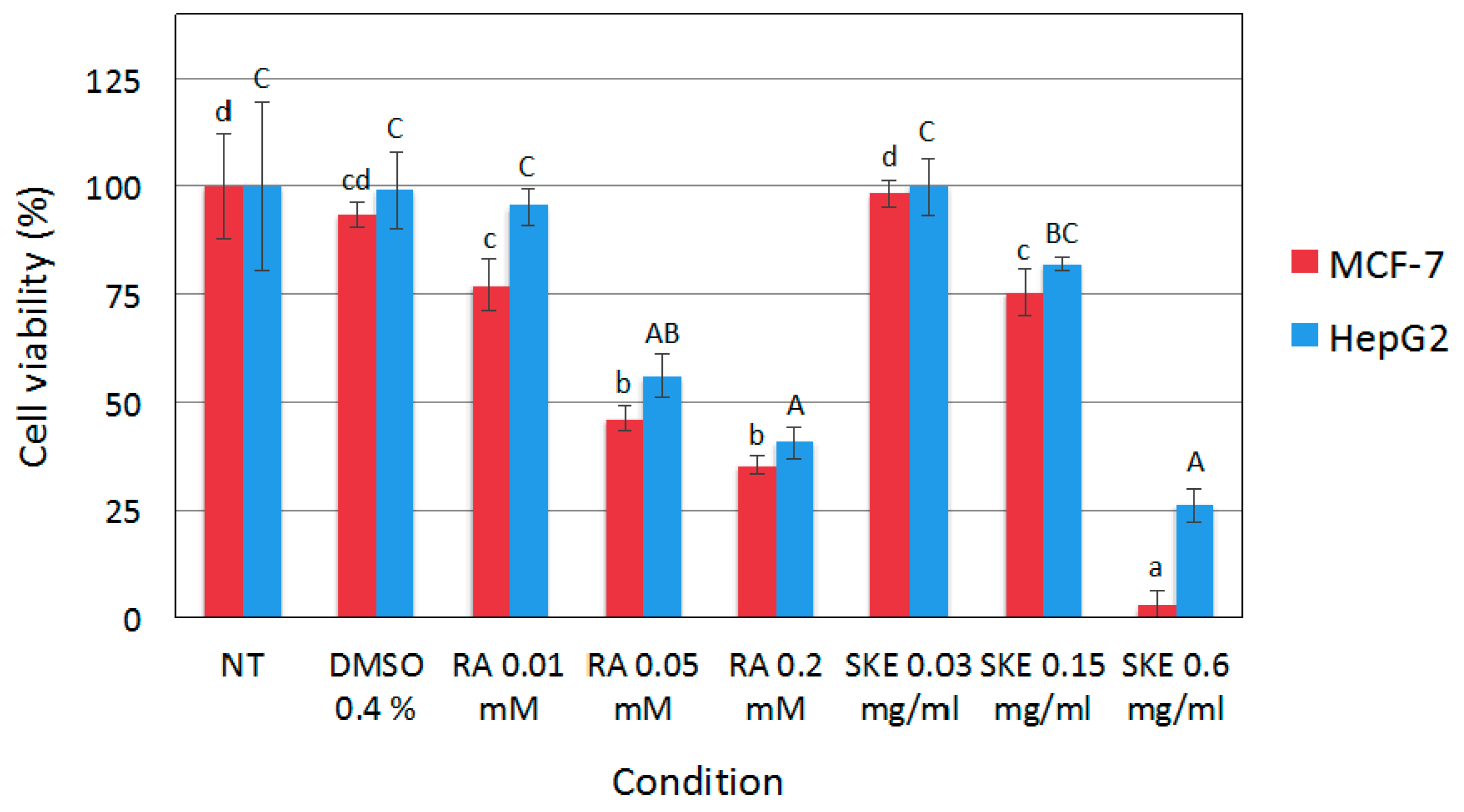
2.3. Reduction of MCF-7 Cell Viability by S. Khuzistanica Extracts
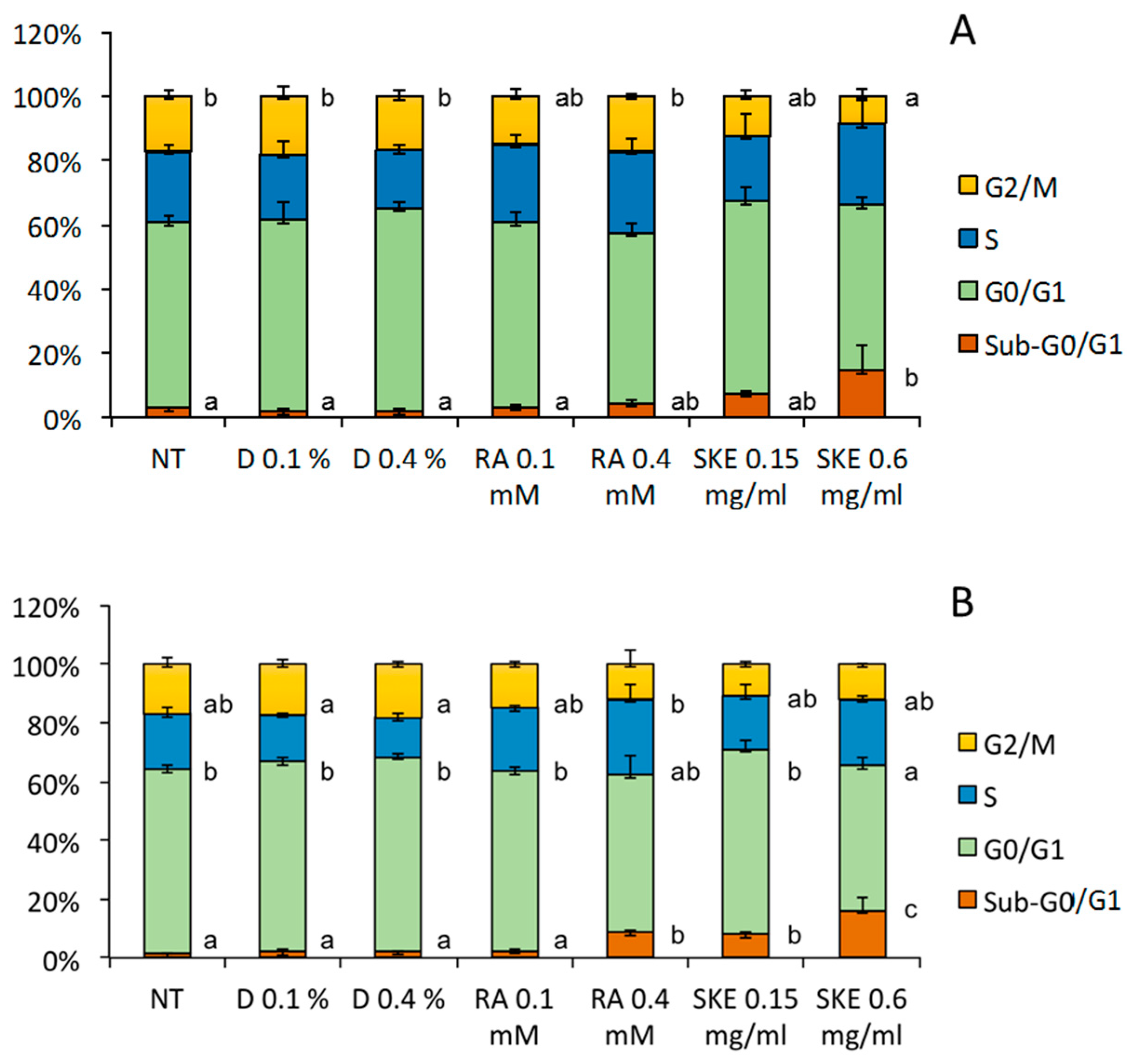
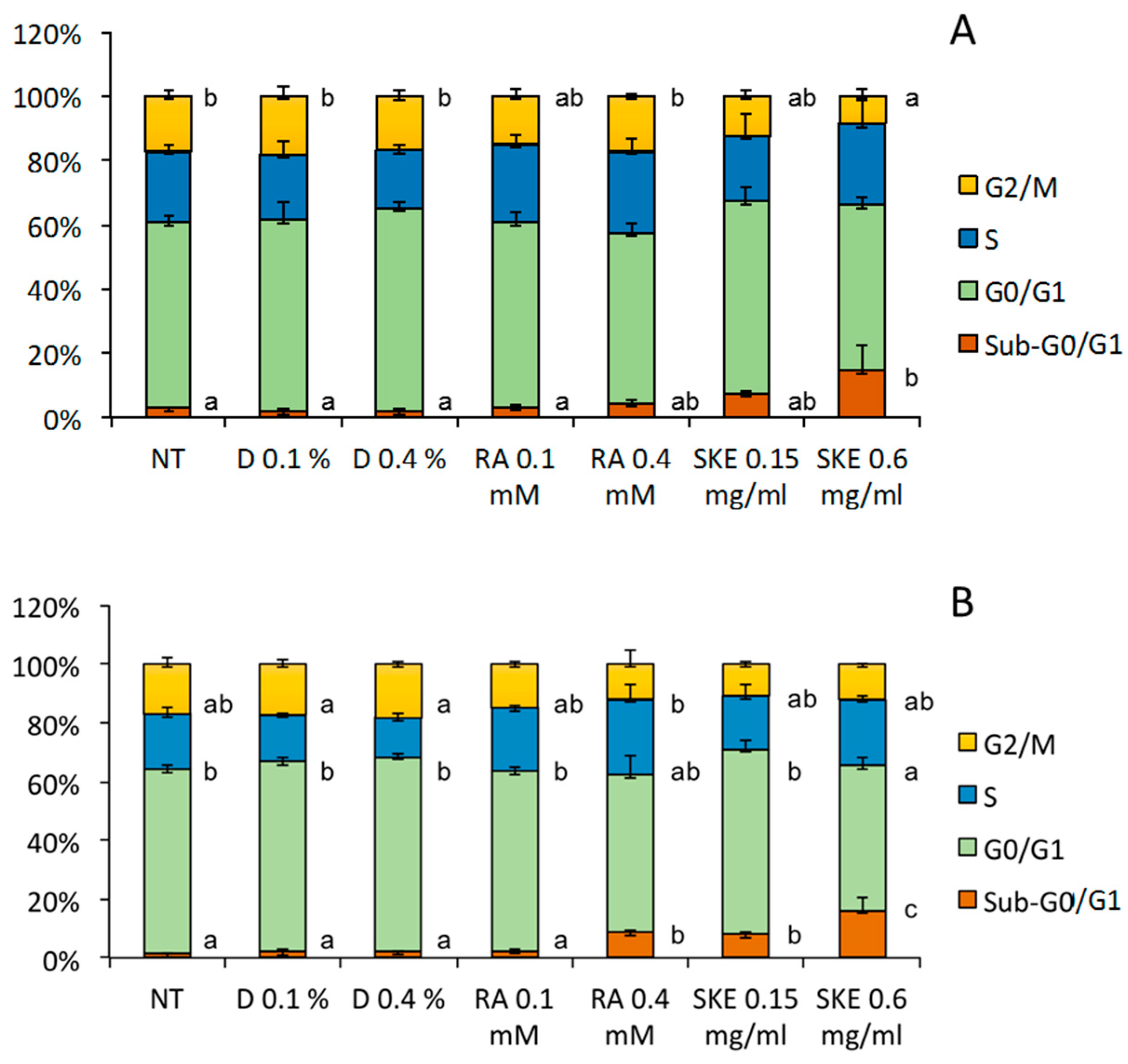
2.4. Induction of MCF-7 Cell Cycle Arrest by S. Khuzistanica Extracts
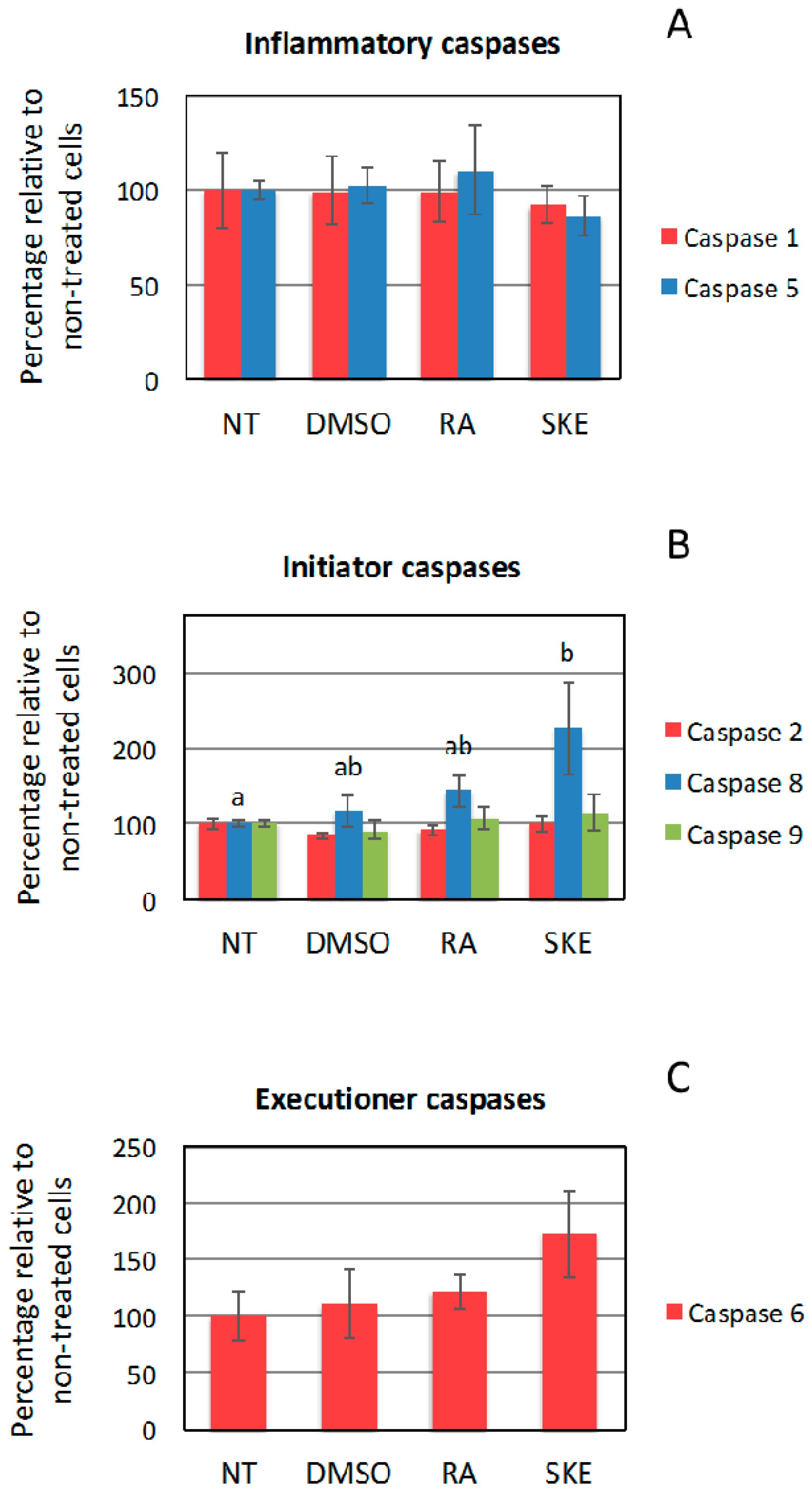
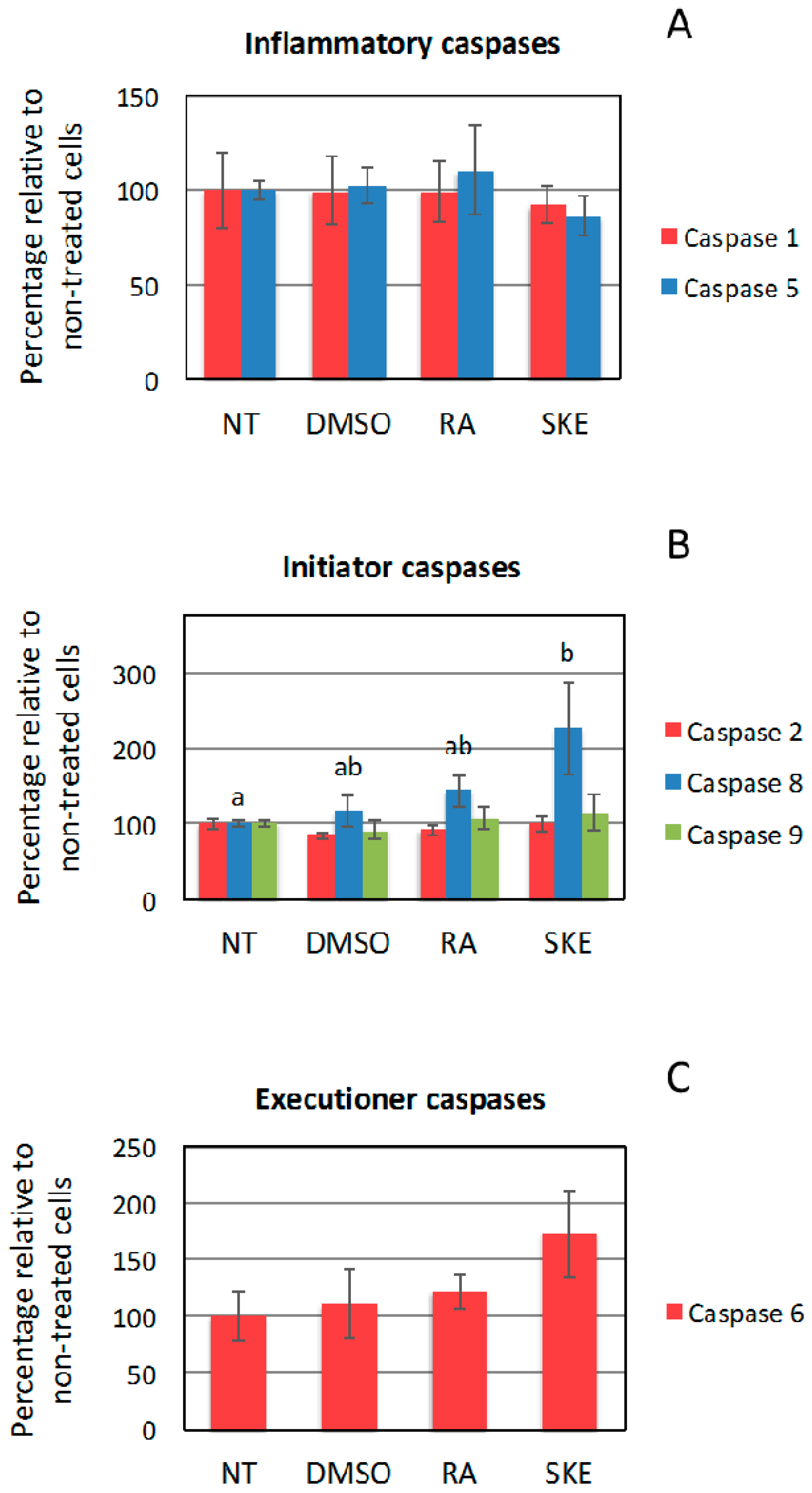
2.5. Effect of S. Khuzistanica Extracts on Caspase Activity of MCF-7 Cells
3. Materials and Methods
3.1. S. Khuzistanica Plant Cell Cultures
3.2. Benchtop Bioreactor Scale
3.3. In-Process Controls
3.4. Rosmarinic Acid Extraction and Quantification
3.5. Preparation of the S. Khuzistanica Extract for Biological Assays
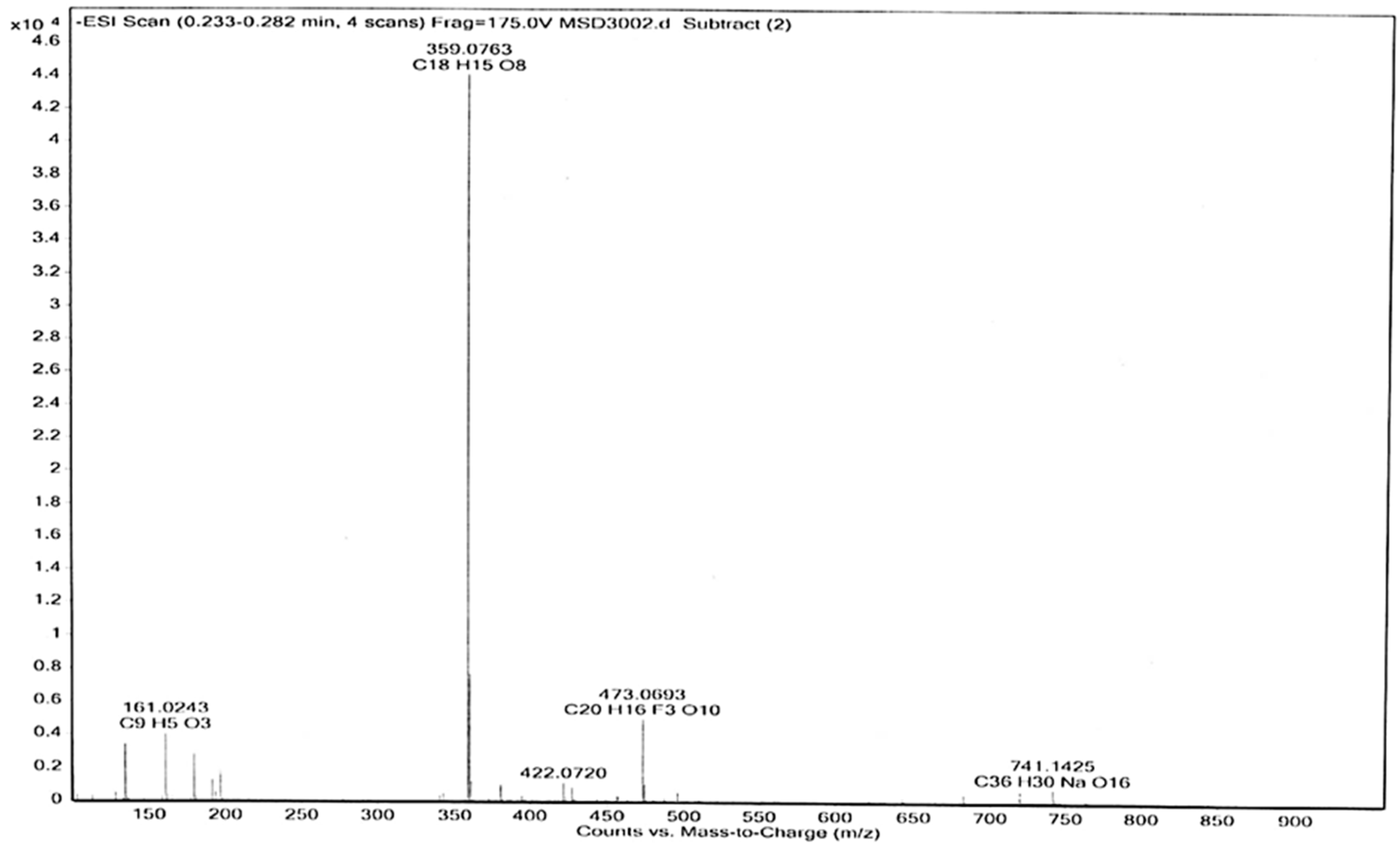
3.6. Electrospray (ESI-MS) Analyses
3.7. Human Cancer-Derived Cell Lines
3.8. Cell Viability Assay
3.9. Cell Cycle Analysis
3.10. Caspase Activity Assays
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Petersen, M.; Abdullah, Y.; Benner, J.; Eberle, D.; Gehlen, K.; Hücherig, S.; Janiak, V.; Kim, K.H.; Sander, M.; Weitzel, C.; et al. Evolution of rosmarinic acid biosynthesis. Phytochemistry 2009, 70, 1663–1679. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M. Rosmarinic acid: New aspects. Phytochem. Rev. 2013, 12, 207–227. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Inyushkina, Y.V.; Fedoreyev, S.A. Rosmarinic acid and its derivatives: Biotechnology and applications. Crit. Rev. Biotechnol. 2012, 32, 203–217. [Google Scholar] [CrossRef]
- Moore, J.; Yousef, M.; Tsiani, E. Anticancer effects of rosemary (Rosmarinus officinalis L.) extract and rosemary extract polyphenols. Nutrients 2016, 8, 731. [Google Scholar] [CrossRef]
- Hadian, J.; Mirjalili, M.H.; Kanani, M.R.; Salehnia, A.; Ganjipoor, P. Phytochemical and morphological characterization of Satureja khuzistanica Jamzad populations from Iran. Chem. Biodiv. 2011, 8, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [PubMed]
- Khojasteh, A.; Mirjalili, M.H.; Palazon, J.; Eibl, R.; Cusido, R.M. Methy-jasmonate enhanced production of rosmarinic acid in cell cultures of Satureja khuzistanica in a bioreactor. Eng. Life Sci. 2016, 16, 740–749. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules. 2016, 21, 182. [Google Scholar] [PubMed]
- Onrubia, M.; Moyano, E.; Bonfill, M.; Cusidó, R.M.; Goossens, A.; Palazón, J. Coronatine, a more powerful elicitor for inducing taxane biosynthesis in Taxus media cell cultures than methyl jasmonate. J. Plant Physiol. 2013, 170, 211–219. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Osuna, L.; Moyano, E.; Bonfill, M.; Tapia, N.; Cusido, R.M.; Palazon, J. Changes in gene transcription and taxane production in elicited cell cultures of Taxus x media and Taxus globosa. Phytochemistry 2015, 117, 174–184. [Google Scholar] [CrossRef]
- Deus-Neumann, B.; Zenk, M.H. Instability of indole alkaloid production in Catharanthus roseus cell suspension cultures. Planta Med. 1984, 50, 427–431. [Google Scholar] [CrossRef]
- Sierra, M.I.; van der Heijden, R.; van der Leer, T.; Verpoorte, R. Stability of alkaloid production in cell suspension cultures of Tabernaemontana divaricata during long- term subculture. Plant Cell Tiss Org Cult. 1992, 28, 59–68. [Google Scholar] [CrossRef]
- Kim, K.H.; Janiak, V.; Petersen, M. Purification, cloning and functional expression of hydroxyphenylpyruvate reductase involved in rosmarinic acid biosynthesis in cell cultures of Coleus blumei. Plant Mol. Biol. 2004, 54, 311–332. [Google Scholar] [CrossRef]
- Sanchez-Muñoz, R.; Bonfill, M.; Cusidó, R.M.; Palazon, J.; Moyano, E. Advances in the regulation of in vitro paclitaxel production: Methylation of a Y-Patch promoter region alters BAPT gene expression in Taxus cell cultures. Plant Cell Physiol. 2018, 59, 2255–2267. [Google Scholar]
- Mustafa, N.R.; de Winter, W.; van Iren, F.; Verpoorte, R. Initiation, growth and cryopreservation of plant cell suspension cultures. Nat. Protoc. 2011, 6, 715–742. [Google Scholar] [CrossRef]
- Fu, C.; Li, L.; Wu, W.; Li, M.; Yu, X.; Yu, L. Assessment of genetic and epigenetic variation during long-term Taxus cell culture. Plant Cell Rep. 2012, 31, 1321–1331. [Google Scholar] [CrossRef]
- Klöckner, W.; Diederichs, S.; Büchs, J. Orbitally shaken single-use bioreactors. Adv. Biochem. Eng. Biotechnol. 2014, 138, 45–60. [Google Scholar] [PubMed]
- Hidalgo, D.; Steinmetz, V.; Brossat, M.; Tournier-Couturier, L.; Cusido, R.M.; Corchete, P.; Palazon, J. An optimized biotechnological system for the production of centellosides based on elicitation and bioconversion of Centella asiatica cell cultures. Eng. Life Sci. 2017, 17, 413–419. [Google Scholar] [CrossRef]
- Yousefzadi, M.; Riahi-Madvar, A.; Hadian, J.; Rezaee, F.; Rafiee, R.; Biniaz, M. Toxicity of essential oil of Satureja khuzistanica: In vitro cytotoxicity and anti-microbial activity. J. Immunotoxicol. 2014, 11, 50–55. [Google Scholar] [CrossRef]
- Esmaeili-Mahani, S.; Samandari-Bahraseman, M.R.; Yaghoobi, M.M. In-vitro anti-proliferative and pro-apoptotic proterties of Sutureja Khuzestanica on human breast cancer (MCF-7) and its synergic effects with anticancer drug vincristine. Iran J Pharm Res. 2018, 17, 343–352. [Google Scholar]
- Yin, Q.H.; Yan, F.X.; Zu, X.Y.; Wu, Y.H.; Wu, X.P.; Liao, M.C.; Deng, S.W.; Yin, L.L.; Zhuang, Y.Z. Anti-proliferative and pro-apoptotic effect of carvacrol on human hepotocelular carcinoma cell line HepG-2. Cytotechnology. 2012, 64, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Hong, C.; Klauck, S.M.; Lin, Y.L.; Efferth, T. Molecular mechanisms of rosmarinic acid from Salvia miltiorrhiza in acute lymphoblastic leukemia cells. J Ethnopharmacol. 2015, 176, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.G.; Hwang, K.A.; Choi, K.C. Rosmarinic Acid, a component of rosemary tea, induced the cell cycle arrest and apoptosis through modulation of HDAC2 expression in prostate cancer cell lines. Nutrients 2018, 10, 1784. [Google Scholar] [CrossRef]
- Huang, Y.; Cai, Y.; Huang, R.; Zheng, X. Rosmarinic acid combined with adriamycin induces apoptosis by triggering mitochondria-mediated signaling pathway in HepG2 and Bel-7402 Cells. Med. Sci. Monit. 2018, 24, 7898–7908. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, M.; Kuzeva, S.; Pavlov, A.; Kovacheva, E.; Ilieva, M. Enhanced rosmarinic acid production by Lavandula vera MM cell suspension culture through elicitation with vanadyl sulfate. Z. Naturforsch. C. 2006, 61, 241–244. [Google Scholar] [CrossRef]
- Sahraroo, A.; Babalar, M.; Mirjalili, M.H.; Fattahi Moghaddam, M.R.; Nejad Ebrahimi, S. In-vitro callus induction and rosmarinic acid quantification in callus culture of Satureja khuzistanica Jamzad (Lamiaceae). Iran J. Pharm. Res. 2014, 13, 1447–1456. [Google Scholar]
- Gallego, A.; Metón, I.; Baanante, I.V.; Ouazzani, J.; Adelin, E.; Palazon, J.; Bonfill, M.; Moyano, E. Viability-reducing activity of Coryllus avellana L. extracts against human cancer cell lines. Biomed. Pharmacother. 2017, 89, 565–572. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Specific Production (mg·g·CDW−1) | Production (mg·L−1) | ||
|---|---|---|---|---|
| Control | COR | Control | COR | |
| 0 | 67.1 ± 2.1 | 179.4 ± 3.7 | ||
| 2 | 94.6 ± 1.9 | 319.3 ± 3.8 | ||
| 4 | 96.5 ± 2.3 | 264.7 ± 4.6 | ||
| 7 | 117.3 ± 2.4 | 312.0 ± 4.0 | ||
| 11 | 128.4 ± 2.9 | 456.0 ± 4.9 | ||
| 14 | 132.5 ± 2.7 | 1184.2 ± 8.6 | ||
| 16 | 127.4 ± 3.1 | 147.3 ± 2.6 | 1901.8 ± 11.0 | 1288.5 ± 13.1 |
| 18 | 132.0 ± 2.1 | 195.5 ± 3.2 | 2121.3 ± 11.7 | 1873.8 ± 7.6 |
| 21 | 150.9 ± 3.2 | 206.2 ± 3.6 | 2134.0 ± 9.0 | 2460.8 ± 12.1 |
| 24 | 163.9 ± 3.5 | 221.6 ± 4.0 | 2221.8 ± 7.0 | 2665.6 ± 12.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khojasteh, A.; Metón, I.; Camino, S.; Cusido, R.M.; Eibl, R.; Palazon, J. In Vitro Study of the Anticancer Effects of Biotechnological Extracts of the Endangered Plant Species Satureja Khuzistanica. Int. J. Mol. Sci. 2019, 20, 2400. https://doi.org/10.3390/ijms20102400
Khojasteh A, Metón I, Camino S, Cusido RM, Eibl R, Palazon J. In Vitro Study of the Anticancer Effects of Biotechnological Extracts of the Endangered Plant Species Satureja Khuzistanica. International Journal of Molecular Sciences. 2019; 20(10):2400. https://doi.org/10.3390/ijms20102400
Chicago/Turabian StyleKhojasteh, Abbas, Isidoro Metón, Sergio Camino, Rosa M. Cusido, Regine Eibl, and Javier Palazon. 2019. "In Vitro Study of the Anticancer Effects of Biotechnological Extracts of the Endangered Plant Species Satureja Khuzistanica" International Journal of Molecular Sciences 20, no. 10: 2400. https://doi.org/10.3390/ijms20102400
APA StyleKhojasteh, A., Metón, I., Camino, S., Cusido, R. M., Eibl, R., & Palazon, J. (2019). In Vitro Study of the Anticancer Effects of Biotechnological Extracts of the Endangered Plant Species Satureja Khuzistanica. International Journal of Molecular Sciences, 20(10), 2400. https://doi.org/10.3390/ijms20102400