Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms


Abstract
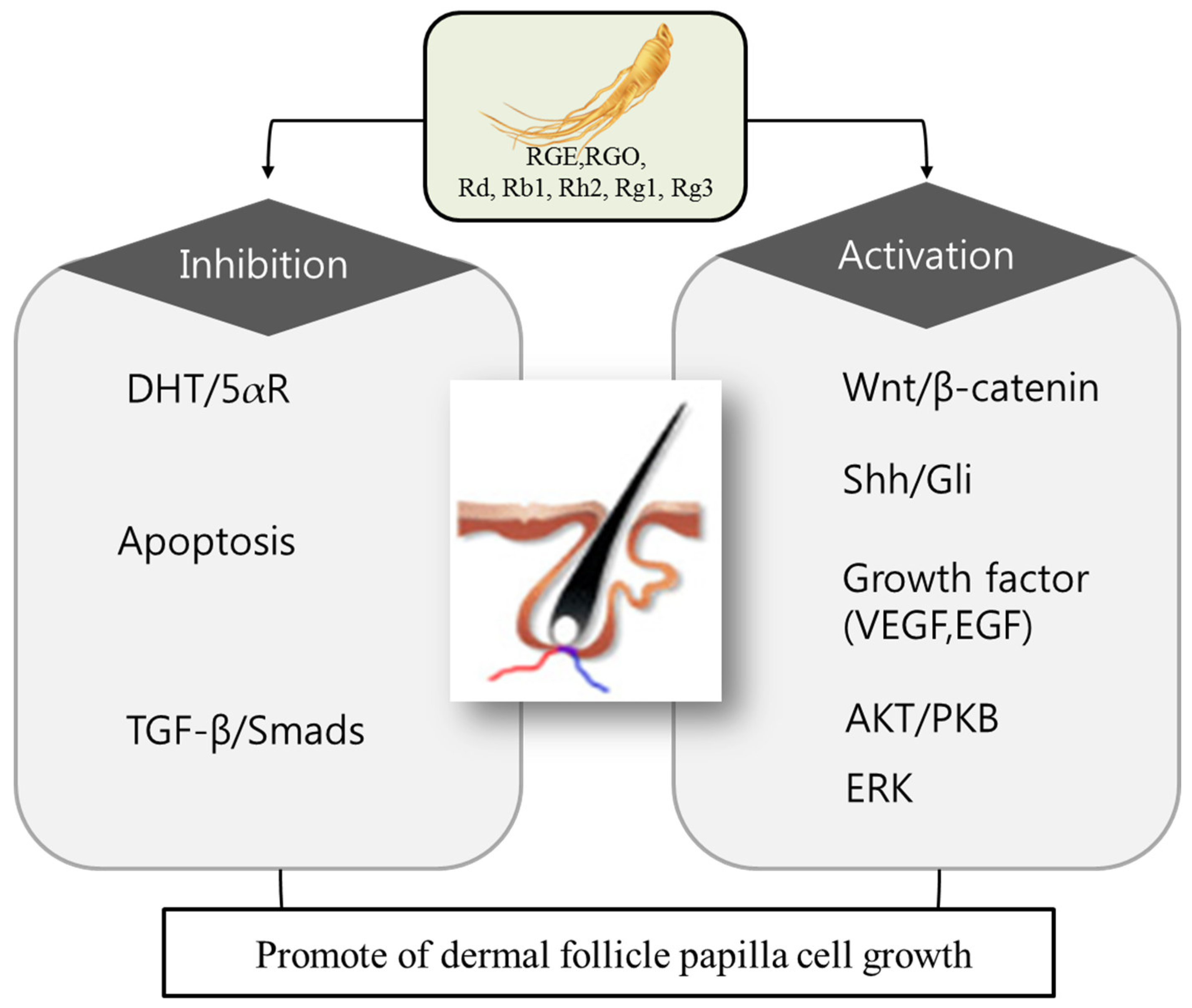
1. Introduction
2. Biochemical Basis of Hair-Growth Promotion by Ginseng
2.1. Prevention of Radiation-Induced Skin Damage
2.2. Antiaging Effects of Ginsenosides
2.3. Modulation of TGF-β Signaling
2.4. Inhibition of 5α-Reductase Enzyme
2.5. Modulation of Wnt/Dickkopf Homologue 1 (DKK1) Signaling
2.6. Modulation of Sonic Hedgehog (Shh) Signaling
2.7. Modulation of JAK2-STAT3 Signaling
2.8. Activation of Dermal Papillary Cell Proliferation
3. Evidence from In Vivo Animal Studies
4. Human Clinical Studies
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Dong Hyun, K. Review: Herbal medicines are activated by intestinal microflora. Nat. Prod. Sci. 2002, 8, 35–43. [Google Scholar]
- Tawab, M.A.; Bahr, U.; Karas, M.; Wurglics, M.; Schubert-Zsilavecz, M. Degradation of ginsenosides in humans after oral administration. Drug Metab. Dispos. 2003, 31, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Rees, J.L. The genetics of sun sensitivity in humans. Am. J. Hum. Genet. 2004, 75, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kim, K.; Lim, C.J. Suppressive properties of ginsenoside Rb2, a protopanaxadiol-type ginseng saponin, on reactive oxygen species and matrix metalloproteinase-2 in UV-B-irradiated human dermal keratinocytes. Biosci. Biotechnol. Biochem. 2015, 79, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Lee, S.; Kho, Y.E.; Kim, K.; Jin, C.D.; Lim, C.J. Stereoselective suppressive effects of protopanaxadiol epimers on uv-b-induced reactive oxygen species and matrix metalloproteinase-2 in human dermal keratinocytes. Can. J. Physiol. Pharmacol. 2015, 93, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Choi, W.Y.; Jung, H.J. Stereoselective skin anti-photoaging properties of ginsenoside Rg3 in UV-B-irradiated keratinocytes. Biol. Pharm. Bull. 2014, 37, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Lee, S.; Choi, W.Y.; Lim, C.J. Skin anti-photoaging properties of ginsenoside Rh2 epimers in UV-B-irradiated human keratinocyte cells. J. Biosci. 2014, 39, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Lee, M.R.; Gu, L.J.; Hossain, M.J.; Kim, H.K.; Sung, C.K. Comparative hair restorer efficacy of medicinal herb on nude (foxn1nu) mice. Biomed Res. Int. 2014, 2014, 319795. [Google Scholar] [CrossRef] [PubMed]
- Gottumukkala, V.R.; Annamalai, T.; Mukhopadhyay, T. Phytochemical investigation and hair growth studies on the rhizomes of nardostachys jatamansi dc. Pharmacogn. Mag. 2011, 7, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Saito, M.; Tabata, M. Bioassay of crude drugs for hair growth promoting activity in mice by a new simple method. Planta Med. 1980, 40, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Jae Hwan, K.; Sang Min, Y.; Jae Eun, C.; Sang Wook, S. Study of the efficacy of Korean red ginseng in the treatment of androgenic alopecia. J. Ginseng Res. 2009, 33, 223–228. [Google Scholar]
- Park, G.H.; Park, K.Y.; Cho, H.I.; Lee, S.M.; Han, J.S.; Won, C.H.; Chang, S.E.; Lee, M.W.; Choi, J.H.; Moon, K.C.; et al. Red ginseng extract promotes the hair growth in cultured human hair follicles. J. Med. Food 2015, 18, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Keum, D.I.; Pi, L.Q.; Hwang, S.T.; Lee, W.S. Protective effect of korean red ginseng against chemotherapeutic drug-induced premature catagen development assessed with human hair follicle organ culture model. J. Ginseng Res. 2016, 40, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Inui, S.; Itami, S. Androgen actions on the human hair follicle: Perspectives. Exp. Dermatol. 2013, 22, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M. Negative impact of testosterone deficiency and 5α-reductase inhibitors therapy on metabolic and sexual function in men. Adv. Exp. Med. Biol. 2017, 1043, 473–526. [Google Scholar] [PubMed]
- Rattan, S.I.; Kryzch, V.; Schnebert, S.; Perrier, E.; Nizard, C. Hormesis-based anti-aging products: A case study of a novel cosmetic. Dose Response 2013, 11, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.J.; Kim, O.Y.; Lee, G.T.; Lee, K.S.; Lee, J.H.; Park, I.C.; Lee, S.J.; Kim, Y.R.; Ahn, K.J.; An, I.S.; et al. Identification of ultraviolet b radiationinduced micrornas in normal human dermal papilla cells. Mol. Med. Rep. 2014, 10, 1663–1670. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.X.; Qin, Z.H.; Zeng, D.; Han, Z.Z.; Zhan, R.T.; Tan, Y.; Chen, W.W. Comparative study on effects of anti-contusion injury, analgesia and anti-inflammation of root and stem of zanthoxylum nitidum. Zhong Yao Cai 2015, 38, 2358–2363. [Google Scholar] [PubMed]
- Hou, C.; Miao, Y.; Wang, J.; Wang, X.; Chen, C.Y.; Hu, Z.Q. Collagenase iv plays an important role in regulating hair cycle by inducing VEGF, IGF-1, and TGF-β expression. Drug Des. Dev. Ther. 2015, 9, 5373–5383. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Sumiyoshi, M.; Sakanaka, M.; Kimura, Y. Effects of ginseng saponins isolated from red ginseng on ultraviolet B-induced skin aging in hairless mice. Eur. J. Pharmacol. 2009, 602, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Cho, S.Y.; Kim, S.J.; Shin, E.S.; Chang, H.K.; Kim, D.H.; Yeom, M.H.; Woe, K.S.; Lee, J.; Sim, Y.C.; et al. Ginsenoside F1 protects human hacat keratinocytes from ultraviolet-B-induced apoptosis by maintaining constant levels of Bcl-2. J. Investig. Dermatol. 2003, 121, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Pham, Q.L.; Jang, H.J.; Kim, K.B. Antiwrinkle effect of fermented black ginseng on human fibroblasts. Int. J. Mol. Med. 2017, 39, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Park, S.Y.; Yin, C.S.; Kim, H.T.; Kim, Y.M.; Yi, T.H. Antiaging effects of the mixture of panax ginseng and crataegus pinnatifida in human dermal fibroblasts and healthy human skin. J. Ginseng Res. 2017, 41, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Park, K.G.; Namgoong, S.; Han, S.K.; Jeong, S.H.; Dhong, E.S.; Kim, W.K. Effects of panax ginseng extract on human dermal fibroblast proliferation and collagen synthesis. Int. Wound J. 2016, 13, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jung, E.; Lee, J.; Huh, S.; Kim, J.; Park, M.; So, J.; Ham, Y.; Jung, K.; Hyun, C.G.; et al. Panax ginseng induces human Type I collagen synthesis through activation of smad signaling. J. Ethnopharmacol. 2007, 109, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lin, Z.; Huang, Y.; Deng, Q.; Xu, H. Study on antiradiation effect of panaxatriol. Zhong Yao Cai 2002, 25, 805–808. [Google Scholar] [PubMed]
- Kaufman, K.D.; Olsen, E.A.; Whiting, D.; Savin, R.; DeVillez, R.; Bergfeld, W.; Price, V.H.; Van Neste, D.; Roberts, J.L.; Hordinsky, M.; et al. Finasteride in the treatment of men with androgenetic alopecia. Finasteride male pattern hair loss study group. J. Am. Acad. Dermatol. 1998, 39, 578–589. [Google Scholar] [CrossRef]
- Foitzik, K.; Lindner, G.; Mueller-Roever, S.; Maurer, M.; Botchkareva, N.; Botchkarev, V.; Handjiski, B.; Metz, M.; Hibino, T.; Soma, T.; et al. Control of murine hair follicle regression (catagen) by TGF-β1 in vivo. FASEB J. 2000, 14, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Sumiyoshi, M.; Kawahira, K.; Sakanaka, M.; Kimura, Y. Effects of red ginseng extract on ultraviolet b-irradiated skin change in C57BL mice. Phytother. Res. 2008, 22, 1423–1427. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ryu, S.W.; Lee, J.; Choi, K.; Kim, S.; Choi, C. Protopanaxatirol type ginsenoside re promotes cyclic growth of hair follicles via inhibiting transforming growth factor β signaling cascades. Biochem. Biophys. Res. Commun. 2016, 470, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.; Hansen, M.G.; Overall, R.W.; Nakamura, M.; Pertile, P.; Klapp, B.F.; Arck, P.C.; Paus, R. Control of human hair growth by neurotrophins: Brain-derived neurotrophic factor inhibits hair shaft elongation, induces catagen, and stimulates follicular transforming growth factor β2 expression. J. Investig. Dermatol. 2005, 124, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Hibino, T.; Nishiyama, T. Role of tgf-β2 in the human hair cycle. J. Dermatol. Sci. 2004, 35, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.W.; Park, K.H.; Hwang, H.S.; Shin, Y.S.; Oh, Y.T.; Kim, C.H. Protective effects of korean red ginseng against radiation-induced apoptosis in human hacat keratinocytes. J. Radiat. Res. 2014, 55, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Botchkarev, V.A.; Botchkareva, N.V.; Albers, K.M.; Chen, L.H.; Welker, P.; Paus, R. A role for p75 neurotrophin receptor in the control of apoptosis-driven hair follicle regression. FASEB J. 2000, 14, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Matsuura, D.; Kanatani, H.; Yano, S.; Tsunakawa, M.; Matsuyama, S.; Shigemori, H. Inhibitory effects of polyacetylene compounds from panax ginseng on neurotrophin receptor-mediated hair growth. Biol. Pharm. Bull. 2017, 40, 1784–1788. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Cha, Y.J.; Yang, K.E.; Jang, I.S.; Son, C.G.; Kim, B.H.; Kim, J.M. Ginsenoside rg3 up-regulates the expression of vascular endothelial growth factor in human dermal papilla cells and mouse hair follicles. Phytother. Res. 2014, 28, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. Wnt signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Kretzschmar, K.; Cottle, D.L.; Schweiger, P.J.; Watt, F.M. The androgen receptor antagonizes Wnt/β-catenin signaling in epidermal stem cells. J. Investig. Dermatol. 2015, 135, 2753–2763. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Park, S.Y.; Hwang, E.S.; Lee, D.G.; Song, H.G.; Mavlonov, G.T.; Yi, T.H. The inductive effect of ginsenoside F2 on hair growth by altering the Wnt signal pathway in telogen mouse skin. Eur. J. Pharmacol. 2014, 730, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Yamazaki, M.; Asanuma, Y.; Kubo, M. Promotion of hair growth by ginseng radix on cultured mouse vibrissal hair follicles. Phytother. Res. 2003, 17, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, S.N.; Hong, Y.D.; Park, B.C.; Na, Y. Panax ginseng extract antagonizes the effect of DKK1-induced catagen-like changes of hair follicles. Int. J. Mol. Med. 2017, 40, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, J.; Burgeson, R.E.; Morgan, B.A. Wnt signaling maintains the hair-inducing activity of the dermal papilla. Genes Dev. 2000, 14, 1181–1185. [Google Scholar] [PubMed]
- Sato, N.; Leopold, P.L.; Crystal, R.G. Effect of adenovirus-mediated expression of sonic hedgehog gene on hair regrowth in mice with chemotherapy-induced alopecia. J. Natl. Cancer Inst. 2001, 93, 1858–1864. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, B.; Dassule, H.R.; Karavanova, I.; Botchkarev, V.A.; Li, J.; Danielian, P.S.; McMahon, J.A.; Lewis, P.M.; Paus, R.; McMahon, A.P. Sonic hedgehog signaling is essential for hair development. Curr. Biol. 1998, 8, 1058–1068. [Google Scholar] [CrossRef]
- Wang, L.C.; Liu, Z.Y.; Gambardella, L.; Delacour, A.; Shapiro, R.; Yang, J.; Sizing, I.; Rayhorn, P.; Garber, E.A.; Benjamin, C.D.; et al. Regular articles: Conditional disruption of hedgehog signaling pathway defines its critical role in hair development and regeneration. J. Investig. Dermatol. 2000, 114, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.L.; Bak, M.J.; Lee, C.; Jun, M.; Jeong, W.S. Hair regenerative mechanisms of red ginseng oil and its major components in the testosterone-induced delay of anagen entry in C57BL/6 mice. Molecules 2017, 22, 1505. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Ito, N.; Takigawa, M.; Ito, T. The hair follicle and immune privilege. J. Investig. Dermatol. Symp. Proc. 2003, 8, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ito, N.; Saatoff, M.; Hashizume, H.; Fukamizu, H.; Nickoloff, B.J.; Takigawa, M.; Paus, R. Maintenance of hair follicle immune privilege is linked to prevention of NK cell attack. J. Investig. Dermatol. 2008, 128, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic t lymphocytes and is reversed by Jak inhibition. Nat. Med. 2014, 20, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Petukhova, L.; Duvic, M.; Hordinsky, M.; Norris, D.; Price, V.; Shimomura, Y.; Kim, H.; Singh, P.; Lee, A.; Chen, W.V.; et al. Genome-wide association study in alopecia areata implicates both innate and adaptive immunity. Nature 2010, 466, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Betz, R.C.; Petukhova, L.; Ripke, S.; Huang, H.; Menelaou, A.; Redler, S.; Becker, T.; Heilmann, S.; Yamany, T.; Duvic, M.; et al. Genome-wide meta-analysis in alopecia areata resolves HLA associations and reveals two new susceptibility loci. Nat. Commun. 2015, 6, 5966. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ito, N.; Saathoff, M.; Bettermann, A.; Takigawa, M.; Paus, R. Interferon-gamma is a potent inducer of catagen-like changes in cultured human anagen hair follicles. Br. J. Dermatol. 2005, 152, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Triyangkulsri, K.; Suchonwanit, P. Role of janus kinase inhibitors in the treatment of alopecia areata. Drug Des. Dev. Ther. 2018, 12, 2323–2335. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zeng, K.W.; Ma, X.L.; Jiang, Y.; Tu, P.F.; Wang, X.M. Ginsenoside Rk1 suppresses pro-inflammatory responses in lipopolysaccharide-stimulated RAW264.7 cells by inhibiting the Jak2/Stat3 pathway. Chin. J. Nat. Med. 2017, 15, 751–757. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Nia, J.K.; Hashim, P.W.; Mansouri, Y.; Alia, E.; Taliercio, M.; Desai, P.N.; Lebwohl, M.G. Efficacy and safety of secukinumab treatment in adults with extensive alopecia areata. Arch. Dermatol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Huyghues-Despointes, B.M.; Tsai, J.; Scholtz, J.M. Nmr study and molecular dynamics simulations of optimized β-hairpin fragments of protein g. Proteins 2007, 69, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Seo, W.; Eun, H.S.; Kim, S.Y.; Jo, E.; Kim, M.H.; Choi, W.M.; Lee, J.H.; Shim, Y.R.; Cui, C.H.; et al. Protective effects of ginsenoside F2 on 12-O-tetradecanoylphorbol-13-acetate-induced skin inflammation in mice. Biochem. Biophys. Res. Commun. 2016, 478, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, M.; Barboni, B.; Turriani, M.; Galeati, G.; Zannoni, A.; Castellani, G.; Berardinelli, P.; Scapolo, P.A. Follicle activation involves vascular endothelial growth factor production and increased blood vessel extension. Biol. Reprod. 2001, 65, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Man, X.Y.; Li, C.M.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Lu, Z.F.; Zheng, M. VEGF induces proliferation of human hair follicle dermal papilla cells through VEGFR-2-mediated activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Kwon, O.S.; Chung, J.H.; Cho, K.H.; Eun, H.C.; Kim, K.H. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J. Dermatol. Sci. 2004, 34, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Lindner, G.; Botchkarev, V.A.; Botchkareva, N.V.; Ling, G.; van der Veen, C.; Paus, R. Analysis of apoptosis during hair follicle regression (catagen). Am. J. Pathol. 1997, 151, 1601–1617. [Google Scholar] [PubMed]
- Park, S.; Shin, W.S.; Ho, J. Fructus panax ginseng extract promotes hair regeneration in C57BL/6 mice. J. Ethnopharmacol. 2011, 138, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, J.J.; Gu, L.J.; Zhang, D.L.; Wang, Y.B.; Sung, C.K. Ginsenosides Rb1 and Rd regulate proliferation of mature keratinocytes through induction of p63 expression in hair follicles. Phytother. Res. 2013, 27, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Jeong, K.S.; Ryu, S.Y.; Kim, T.H. Panax ginseng prevents apoptosis in hair follicles and accelerates recovery of hair medullary cells in irradiated mice. In Vivo 1998, 12, 219–222. [Google Scholar] [PubMed]
- Oh, G.N.; Son, S.W. Efficacy of korean red ginseng in the treatment of alopecia areata. J. Ginseng Res. 2012, 36, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.J.; Yoo, M.G.; Son, S.W. The efficacy of 3% minoxidil vs. Combined 3% minoxidil and korean red ginseng in treating female pattern alopecia. Int. J. Dermatol. 2014, 53, e340–e342. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, B.Y.; Cho, S.Y.; Sung, D.S.; Chang, H.K.; Yeom, M.H.; Kim, D.H.; Sim, Y.C.; Lee, Y.S. Compound k induces expression of hyaluronan synthase 2 gene in transformed human keratinocytes and increases hyaluronan in hairless mouse skin. Biochem. Biophys. Res. Commun. 2004, 316, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.G.; Jeon, A.J.; Yoon, J.H.; Song, D.; Kim, J.E.; Kwon, J.Y.; Kim, J.R.; Kang, N.J.; Park, J.S.; Yeom, M.H.; et al. 20-O-β-D-glucopyranosyl-20(S)-protopanaxadiol, a metabolite of ginsenoside Rb1, enhances the production of hyaluronic acid through the activation of ERK and AKT/PKB mediated by Src tyrosin kinase in human keratinocytes. Int. J. Mol. Med. 2015, 35, 1388–1394. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type | Study Model | Dosage | Action Mechanism | Target | Reference |
|---|---|---|---|---|---|
| Fructus panax | Human hair dermal papilla cells | 0.8, 4, 20, 100, 500 μg/mL | FPG elicited the proliferation of DPC by the upregulation of antiapoptotic Bcl-2 accompanied by the inhibition of apoptotic Bax expression | Apoptosis | [4] |
| ginseng extract (FPG) (95% EtOH) | Male six-week-old C57BL/6 mice | 1 mg/mL | |||
| Ginsenoside Re | Male six-week-old C57BL/6 mice | 1 or 5 mg/d | Ginsenoside Re is the effective constituent in Panax ginseng that promotes hair growth through inhibition of transition related TGF-β signaling pathways. | TGF | [5] |
| Cultured C57BL/6 mouse HFs | 10 or 50 mg/L | ||||
| HeLa cells | 10 mg/L | ||||
| Polyacetylenes isolated from P. ginseng | Neurotrophin receptor-binding inhibition assay | sample solution (10, 30, and 100 μM) | Inhibits BDNF-TrkB binding. | Growth | [6] |
| Ginsenoside F2 | Human hair dermal papilla cells | 0.01, 0.1, 1, and 10 μM | (1) Stimulates proliferation of HHDPC and HaCaT; (2) increases β-catenin and Lef-1 expression and decreases DKK-1 expression in HHDPC; (3) hair anagen induction and acceleration of hair growth in mouse model; (4) increases β-catenin expression and decreases DKK-1 expression in mouse tissue. | WNT | [7] |
| Human keratinocyte (HaCaT) cells | |||||
| Male six-week-old C57BL/6 mice | 0. 5 and 2.5 mg/kg | ||||
| Root of PG extract (70% EtOH) | Human ORS keratinocytes | 20 ppm | PG extract may enhance ORS and hDPC stimulation of hair follicle growth despite the presence of DKK-1, a strong catagen inducer | WNT | [29] |
| Anagen HFs from human scalp-skin specimens | |||||
| Korean Red Ginseng (KRG) | Human (patients diagnosed with AA) | Treated with corticosteroid ILI while taking KRG | KRG can result in improved hair regrowth in AA patients. | [20] | |
| KRG extract | Follicular keratinocytes (FKCs) | 0~1000 μg/mL | KRG may protect against 4-HC-induced premature dystrophy as it occurs in CIA in vivo. Possible mechanisms include the stimulation of hair matrix keratinocyte proliferation and inhibition of hair matrix keratinocyte apoptosis, which are possibly mediated through modulation of p53 and Bax/Bcl-2 expression. | Apotosis | [21] |
| Human anagen hair follicles | 500 μg/mL | ||||
| Red ginseng extract (RGE) | Six-week-old female C57BL/6 mice | 3% | RGE and its ginsenosides may enhance hDPC proliferation, activate the ERK and AKT/PKB signaling pathways in hDPCs, upregulate hair matrix keratinocyte proliferation, and inhibit DHT-induced androgen receptor transcription. | Growth 5aR | [16] |
| RGE, insenoside-Rb1 | Human hair follicles | 100 μg/mL | |||
| Red ginseng oil (RGO) | C57BL/6 mice | RGO 10% | Upregulates Wnt/-catenin and Shh/Gli pathways-mediated expression of genes such as β-catenin, Lef-1, Sonic hedgehog, Smoothened, Gli-1, Cyclin D1, and Cyclin E in TES-treated mice. RGO and its major components reduce the protein level of TGF-β but enhance the expression of antiapoptotic protein Bcl-2. | WNT Shh Growth TGF Apoptosis | [25] |
| KRG | Human (patients with female pattern hair loss) | Oral | Patients about the size of the vertex spot, hair loss on the top of scalp, bitemporal recession, hair shedding, hair quality, and overall satisfaction; group 2 was more satisfied at 24 weeks. | [68] | |
| Ginsenoside Rg3 | Human DP cells | 1, 5, 10 μM | Dose-dependent increases in VEGF, CD8, CD34 Rg3 might increase hair growth through stimulation of hair-follicle stem cells | Growth | [69] |
| Female C57BL/6 mice | 1000 μM | ||||
| Red ginseng (RGE) | B6C3F1 mice | 20, 50 μg/mL | Hair growth-promoting assay using mouse vibrissal follicles in organ culture | Growth | [26] |
| White ginseng (WGE) Ginsenoside-Rb1 (G-Rb1), Rg1 (G-Rg1), -Ro (G-Ro) | Mouse vibrissal hair follicles | 10 μg/mL | |||
| Ginseng rhizome Ginsenoside Ro | C57BL/6 mice | extracts of red ginseng rhizomes (2 mg/mouse) and ginsenoside Ro (0.2 mg/mouse) | Inhibitory activity against 5αR in the androgenetic alopecia model. | 5aR | [27] |
| Ginsenosides Rb1, Re, and Rg1 | Cultured hHFs | 2, 5, and 10 mg/mL PG extracts and 1 mM of the ginsenosides Rb1, Re and Rg1 | PG extract using hHF organ culture, and promoting hair growth through similar mechanisms to those of minoxidil. | 5aR | [36] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.Y. Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 2703. https://doi.org/10.3390/ijms19092703
Choi BY. Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. International Journal of Molecular Sciences. 2018; 19(9):2703. https://doi.org/10.3390/ijms19092703
Chicago/Turabian StyleChoi, Bu Young. 2018. "Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms" International Journal of Molecular Sciences 19, no. 9: 2703. https://doi.org/10.3390/ijms19092703
APA StyleChoi, B. Y. (2018). Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. International Journal of Molecular Sciences, 19(9), 2703. https://doi.org/10.3390/ijms19092703